
In Silico Characterization of the Secretome of the Fungal Pathogen Thielaviopsis punctulata, the Causal Agent of Date Palm Black Scorch Disease
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Information and Gene Prediction
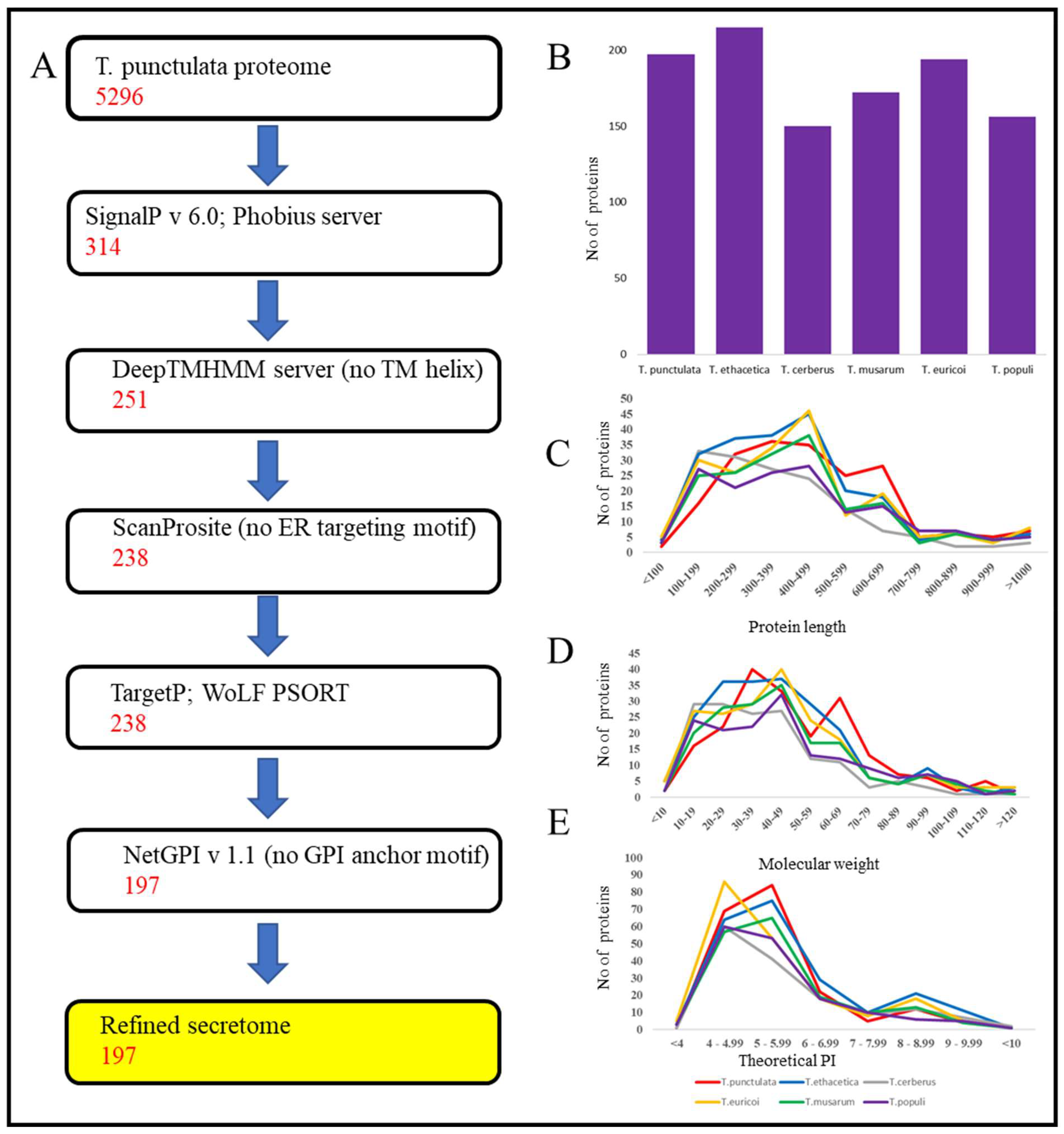
2.2. Prediction of the Secretome
2.3. Annotation of Secretory Proteins
3. Results and Discussion
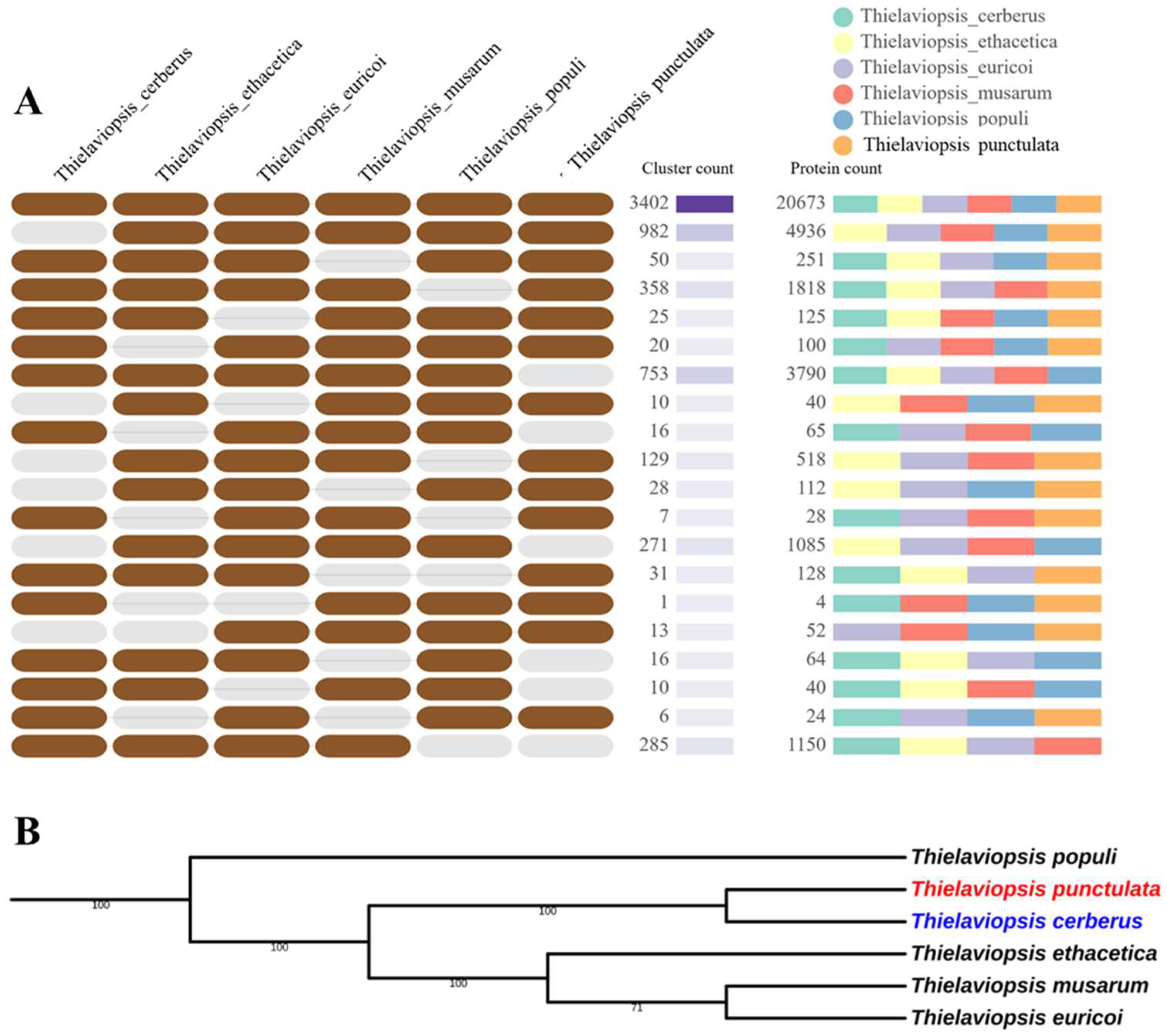
3.1. Prediction of Gene Models and Orthologue Analysis
3.2. Secretome Identification and Analysis
3.3. Structural and Functional Characterization of Secretome
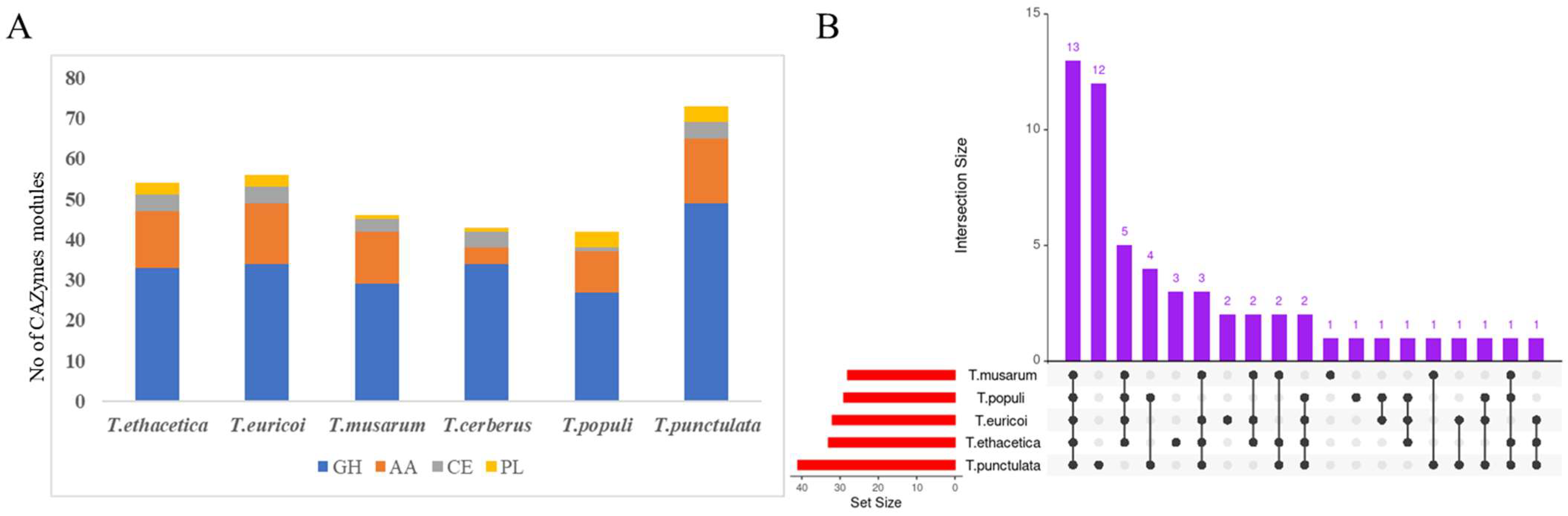
3.4. Carbohydrate Active Enzymes
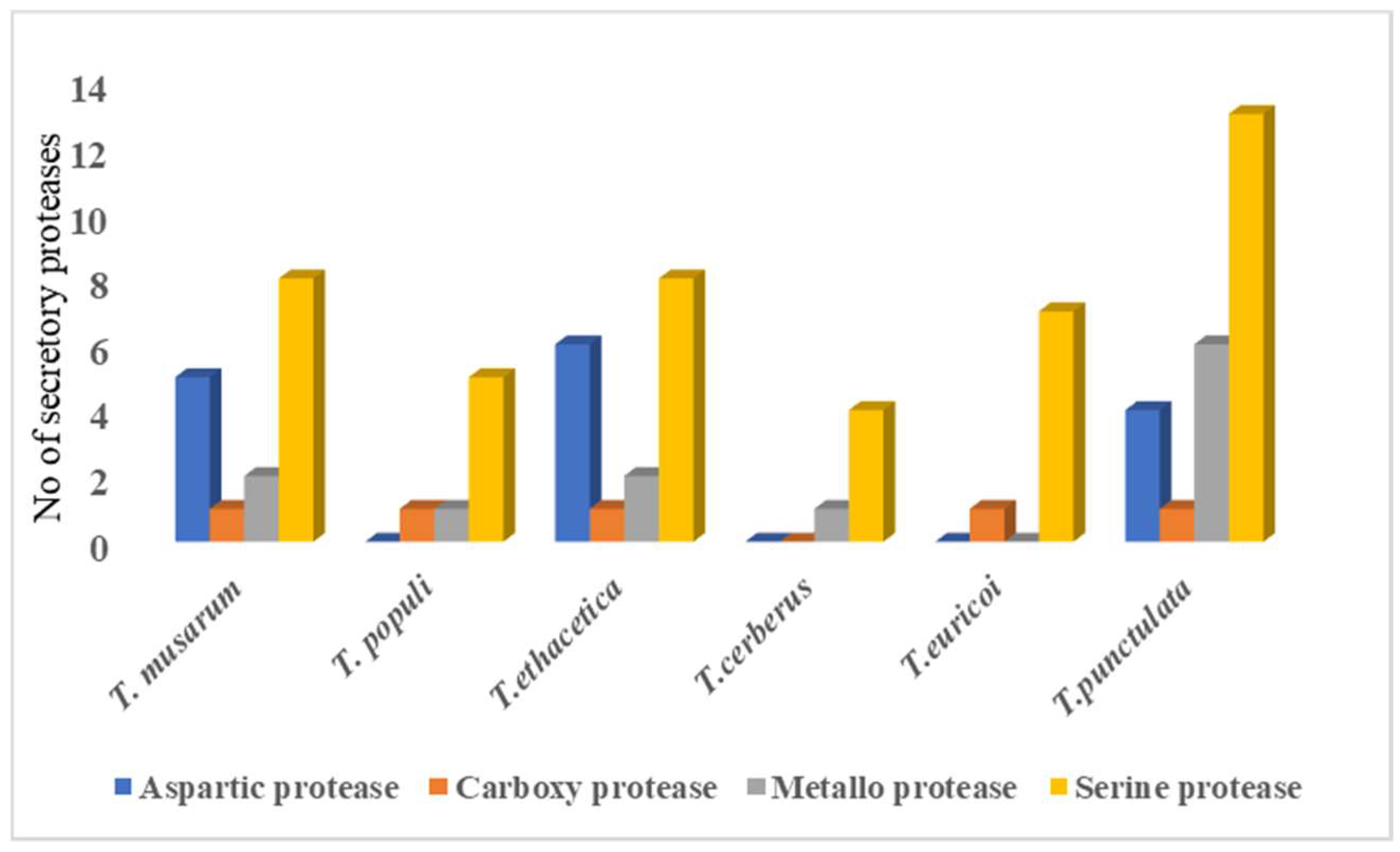
3.5. Secreted Proteases
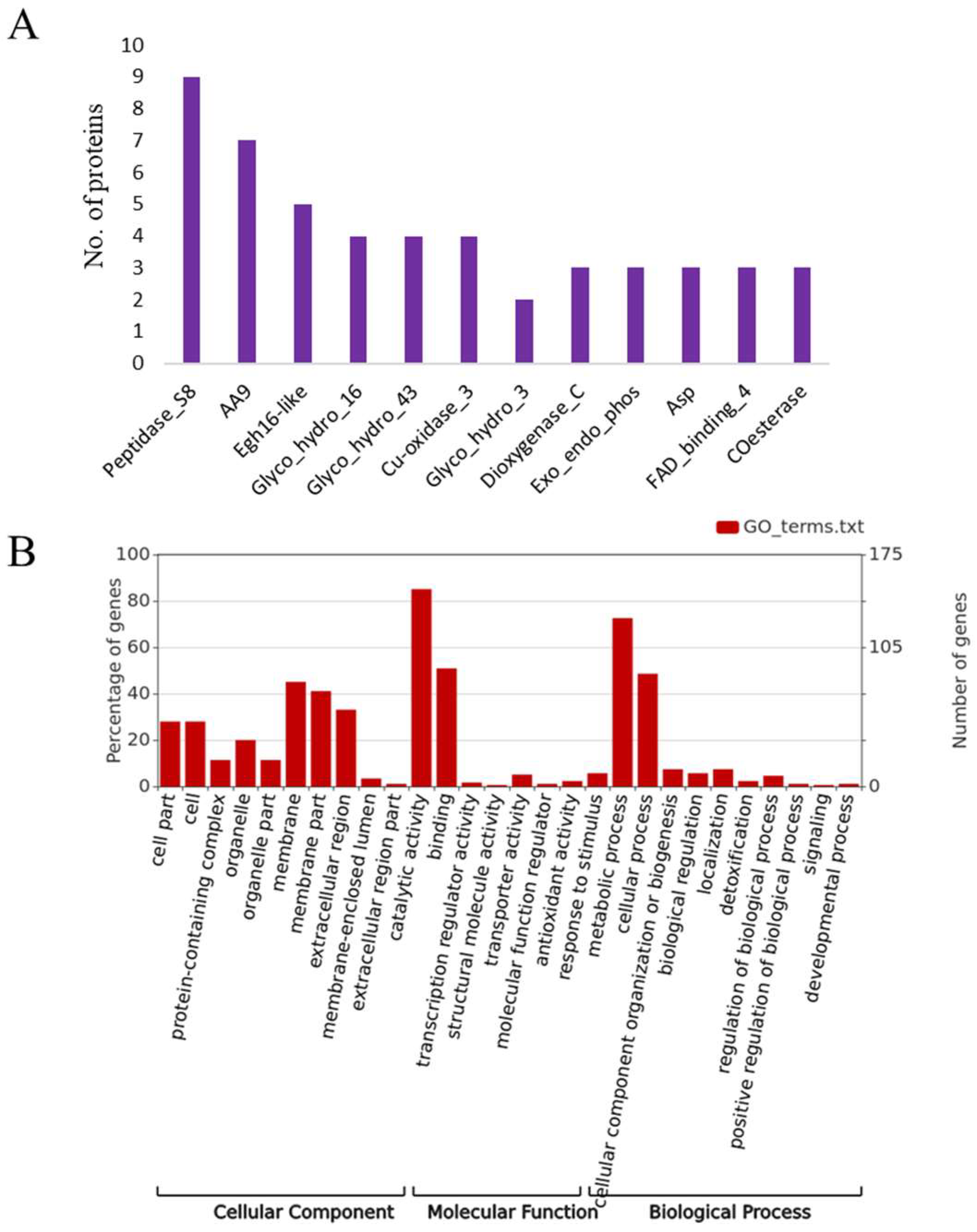
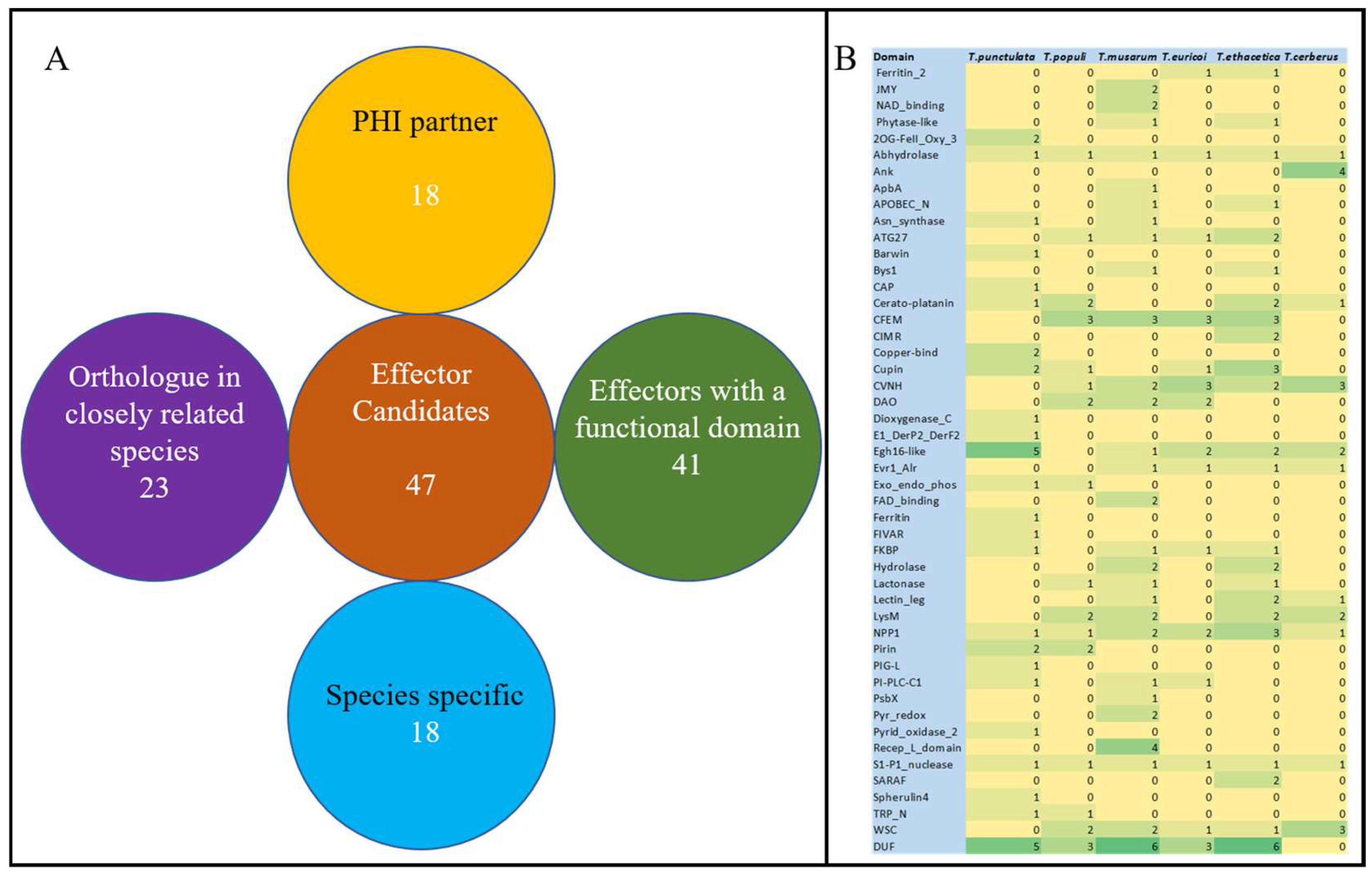
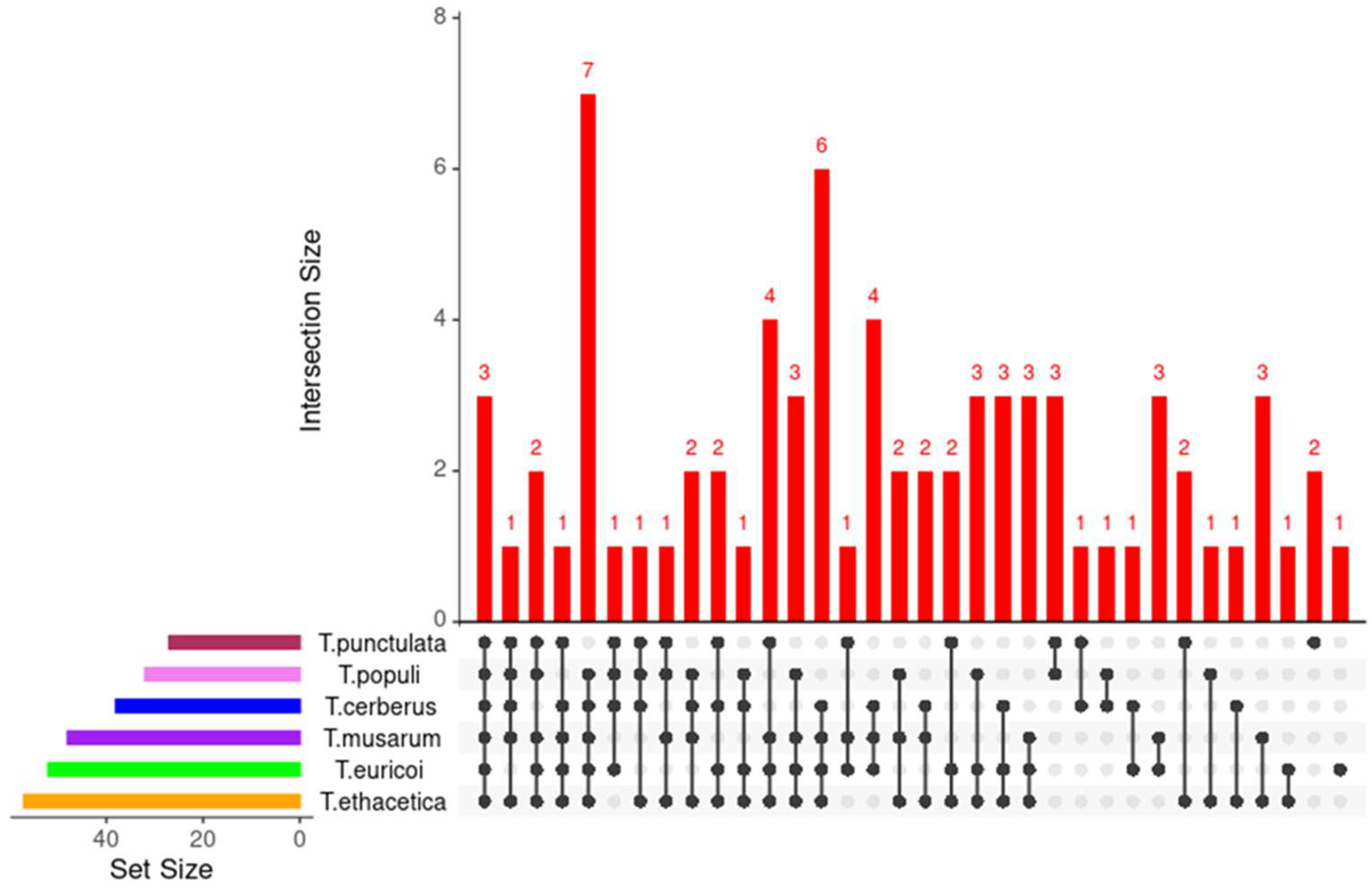
3.6. Putative Effector Proteins
3.7. Putative Virulence Factors
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biju, V.C.; Fokkens, L.; Houterman, P.M.; Rep, M.; Cornelissen, B.J.C. Multiple Evolutionary Trajectories Have Led to the Emergence of Races in Fusarium oxysporum f. sp. lycopersici. Appl. Environ. Microbiol. 2017, 83, e02548-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontes, J.G.D.M.; Fernandes, L.S.; dos Santos, R.V.; Tasic, L.; Fill, T.P. Virulence Factors in the Phytopathogen–Host Interactions: An Overview. J. Agric. Food Chem. 2020, 68, 7555–7570. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-M.; Zhang, Y. Plant Immunity: Danger Perception and Signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Ali, G.S.; Reddy, A. PAMP-triggered immunity. Plant Signal. Behav. 2008, 3, 423–426. [Google Scholar] [CrossRef]
- de Wit, P.J.G.M.; Mehrabi, R.; van den Burg, H.A.; Stergiopoulos, I. Fungal effector proteins: Past, present and future. Mol. Plant Pathol. 2009, 10, 735–747. [Google Scholar] [CrossRef]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal Effectors and Plant Susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Pradhan, A.; Ghosh, S.; Sahoo, D.; Jha, G. Fungal effectors, the double edge sword of phytopathogens. Curr. Genet. 2020, 67, 27–40. [Google Scholar] [CrossRef]
- Todd, J.N.A.; Carreón-Anguiano, K.G.; Islas-Flores, I.; Canto-Canché, B. Fungal Effectoromics: A World in Constant Evolution. Int. J. Mol. Sci. 2022, 23, 13433. [Google Scholar] [CrossRef]
- Ma, W.; Wang, Y.; McDowell, J. Focus on Effector-Triggered Susceptibility. Mol. Plant-Microbe Interact. 2018, 31, 5. [Google Scholar] [CrossRef]
- Hammond-Kosack, K.E.; Jones, J.D. Plant disease resistance genes. Annu. Rev. Plant Biol. 1997, 48, 575–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Cheng, Y. Advances in Fungal Elicitor-Triggered Plant Immunity. Int. J. Mol. Sci. 2022, 23, 12003. [Google Scholar] [CrossRef] [PubMed]
- Laugé, R.; de Wit, P. Fungal Avirulence Genes: Structure and Possible Functions. Fungal Genet. Biol. 1998, 24, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, Y.; Li, B.; Zhang, Z.; Qin, G.; Chen, T.; Tian, S. Molecular mechanisms underlying multi-level defense responses of horticultural crops to fungal pathogens. Hortic. Res. 2022, 9, uhac066. [Google Scholar] [CrossRef] [PubMed]
- Petit-Houdenot, Y.; Fudal, I. Complex Interactions between Fungal Avirulence Genes and Their Corresponding Plant Resistance Genes and Consequences for Disease Resistance Management. Front. Plant Sci. 2017, 8, 1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balint-Kurti, P.; Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takken, F.; Rep, M. The arms race between tomato and Fusarium oxysporum. Mol. Plant Pathol. 2010, 11, 309–314. [Google Scholar] [CrossRef]
- Anderson, J.; Gleason, C.A.; Foley, R.C.; Thrall, P.H.; Burdon, J.B.; Singh, K.B. Plants versus pathogens: An evolutionary arms race. Funct. Plant Biol. 2010, 37, 499–512. [Google Scholar] [CrossRef]
- Jaswal, R.; Kiran, K.; Rajarammohan, S.; Dubey, H.; Singh, P.K.; Sharma, Y.; Deshmukh, R.; Sonah, H.; Gupta, N.; Sharma, T. Effector Biology of Biotrophic Plant Fungal Pathogens: Current Advances and Future Prospects. Microbiol. Res. 2020, 241, 126567. [Google Scholar] [CrossRef]
- Rafiqi, M.; Jelonek, L.; Diouf, A.M.; Mbaye, A.; Rep, M.; Diarra, A. Profile of the in silico secretome of the palm dieback pathogen, Fusarium oxysporum f. sp. albedinis, a fungus that puts natural oases at risk. PLoS ONE 2022, 17, e0260830. [Google Scholar] [CrossRef]
- Jansson, H.B.; Lopez Lorca, L.V.; Abdullah, S.K. Diseases of date palms (phoenix dactylifera L.). Basrah J. Date Palm Res. 2010, 9, 1–44. [Google Scholar]
- Abdullah, S.; Asensio, L.; Monfort, E.; Gomez-Vidal, S.; Salinas, J.; Lorca, L.; Jansson, H. Incidence of the Two Date Palm Pathogens, Thielaviopsis Paradoxa and T. Punctulata in Soil From Date Palm Plantations in Elx, South-East Spain. J. Plant Prot. Res. 2009, 49. [Google Scholar] [CrossRef]
- Al-Raisi, Y.; B’Chir, M.; Al-Mandhari, A.; Deadman, M.; Gowen, S. First report of Ceratocystis radicicola associated with date palm disease in Oman. New Dis. Rep. 2011, 23, 23. [Google Scholar] [CrossRef] [Green Version]
- Al-Sharidah, A. Report of Black Scorch Disease on Date Palm Trees in the State of Kuwait. Am. Sci. Res. J. Eng. 2017, 28, 14–17. [Google Scholar]
- Alhudaib, K.A.; El-Ganainy, S.M.; Almaghasla, M.I.; Sattar, M.N. Characterization and Control of Thielaviopsis punctulata on Date Palm in Saudi Arabia. Plants 2022, 11, 250. [Google Scholar] [CrossRef] [PubMed]
- Al-Naemi, F.A.; Nishad, R.; Ahmed, T.A.; Radwan, O. First Report of Thielaviopsis punctulata Causing Black Scorch Disease on Date Palm in Qatar. Plant Dis. 2014, 98, 1437. [Google Scholar] [CrossRef] [PubMed]
- Saeed, E.E.; Sham, A.; El-Tarabily, K.; Abu Elsamen, F.; Iratni, R.; AbuQamar, S.F. Chemical Control of Black Scorch Disease on Date Palm Caused by the Fungal Pathogen Thielaviopsis punctulata in United Arab Emirates. Plant Dis. 2016, 100, 2370–2376. [Google Scholar] [CrossRef] [Green Version]
- Al-Naemi, F.A.; Ahmed, T.A.; Nishad, R.; Radwan, O. Antagonistic Effects of Trichoderma harzianum Isolates against Ceratocystis radicicola: Pioneering a Biocontrol Strategy against Black Scorch Disease in Date Palm Trees. J. Phytopathol. 2016, 164, 464–475. [Google Scholar] [CrossRef]
- Saeed, E.E.; Sham, A.; Salmin, Z.; Abdelmowla, Y.; Iratni, R.; El-Tarabily, K.; Abu Qamar, S. Streptomyces globosus UAE1, a Potential Effective Biocontrol Agent for Black Scorch Disease in Date Palm Plantations. Front. Microbiol. 2017, 8, 1455. [Google Scholar] [CrossRef] [Green Version]
- Wingfield, B.D.; Ades, P.K.; Al-Naemi, F.A.; Beirn, L.A.; Bihon, W.; Crouch, J.A.; De Beer, Z.W.; De Vos, L.; Duong, T.A.; Fields, C.J.; et al. Draft genome sequences of Chrysoporthe austroafricana, Diplodia scrobiculata, Fusarium nygamai, Leptographium lundbergii, Limonomyces culmigenus, Stagonosporopsis tanaceti, and Thielaviopsis punctulata. IMA Fungus 2015, 6, 233–248. [Google Scholar] [CrossRef] [Green Version]
- Sonah, H.; Deshmukh, R.; Bélanger, R.R. Computational Prediction of Effector Proteins in Fungi: Opportunities and Challenges. Front. Plant Sci. 2016, 7, 126. [Google Scholar] [CrossRef] [Green Version]
- Rafiqi, M.; Jelonek, L.; Akum, N.F.; Zhang, F.; Kogel, K.-H. Effector candidates in the secretome of Piriformospora indica, a ubiquitous plant-associated fungus. Front. Plant Sci. 2013, 4, 228. [Google Scholar] [CrossRef] [Green Version]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Krogh, A.; Sonnhammer, E.L. A Combined Transmembrane Topology and Signal Peptide Prediction Method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gattiker, A.; Gasteiger, E.; Bairoch, A. ScanProsite: A reference implementation of a PROSITE scanning tool. Appl. Bioinform. 2002, 1, 107–108. [Google Scholar]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Armenteros, J.J.A.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [Green Version]
- Gíslason, M.H.; Nielsen, H.; Armenteros, J.J.A.; Johansen, A.R. Prediction of GPI-anchored proteins with pointer neural networks. Curr. Res. Biotechnol. 2021, 3, 6–13. [Google Scholar] [CrossRef]
- Lees, J.; Yeats, C.; Perkins, J.R.; Sillitoe, I.; Rentzsch, R.; Dessailly, B.H.; Orengo, C. Gene3D: A domain-based resource for comparative genomics, functional annotation and protein network analysis. Nucleic Acids Res. 2011, 40, D465–D471. [Google Scholar] [CrossRef]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2022, 51, D523–D531. [Google Scholar] [CrossRef]
- Yin, Y.; Mao, X.; Yang, J.; Chen, X.; Mao, F.; Xu, Y. dbCAN: A web resource for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2012, 40, W445–W451. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Sperschneider, J.; Dodds, P.N. EffectorP 3.0: Prediction of Apoplastic and Cytoplasmic Effectors in Fungi and Oomycetes. Mol. Plant-Microbe Interact. 2022, 35, 146–156. [Google Scholar] [CrossRef]
- Winnenburg, R.; Urban, M.; Beacham, A.; Baldwin, T.K.; Holland, S.; Lindeberg, M.; Hansen, H.; Rawlings, C.; Hammond-Kosack, K.E.; Köhler, J. PHI-base update: Additions to the pathogen host interaction database. Nucleic Acids Res. 2007, 36, D572–D576. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Benini, S. Carbohydrate-Active Enzymes: Structure, Activity, and Reaction Products. Int. J. Mol. Sci. 2020, 21, 2727. [Google Scholar] [CrossRef] [Green Version]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, Functions, and Mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [Green Version]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant Cell Wall–Degrading Enzymes and Their Secretion in Plant-Pathogenic Fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef] [PubMed]
- Villares, A.; Moreau, C.; Bennati-Granier, C.; Garajova, S.; Foucat, L.; Falourd, X.; Saake, B.; Berrin, J.-G.; Cathala, B. Lytic polysaccharide monooxygenases disrupt the cellulose fibers structure. Sci. Rep. 2017, 7, 40262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, C.M.; Beeson, W.T.; Cate, J.H.; Marletta, M.A. Cellobiose Dehydrogenase and a Copper-Dependent Polysaccharide Monooxygenase Potentiate Cellulose Degradation by Neurospora crassa. ACS Chem. Biol. 2011, 6, 1399–1406. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.J.; Vaaje-Kolstad, G.; Westereng, B.; Eijsink, V.G. Novel enzymes for the degradation of cellulose. Biotechnol. Biofuels 2012, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- van den Brink, J.; De Vries, R.P. Fungal enzyme sets for plant polysaccharide degradation. Appl. Microbiol. Biotechnol. 2011, 91, 1477–1492. [Google Scholar] [CrossRef] [Green Version]
- Margolles-Clark, E.; Tenkanen, M.; Soderlund, H.; Penttila, M. Acetyl Xylan Esterase from Trichoderma reesei Contains an Active-Site Serine Residue and a Cellulose-Binding Domain. JBIC J. Biol. Inorg. Chem. 1996, 237, 553–560. [Google Scholar] [CrossRef]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-degrading enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef]
- Nagel, J.H.; Wingfield, M.J.; Slippers, B. Increased abundance of secreted hydrolytic enzymes and secondary metabolite gene clusters define the genomes of latent plant pathogens in the Botryosphaeriaceae. BMC Genom. 2021, 22, 589. [Google Scholar] [CrossRef]
- Jashni, M.E.; Mehrabi, R.; Collemare, J.; Mesarich, C.; De Wit, P.J.G.M. The battle in the apoplast: Further insights into the roles of proteases and their inhibitors in plant–pathogen interactions. Front. Plant Sci. 2015, 6, 584. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, I.; de Wit, P.J. Fungal Effector Proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Songkumarn, P.; Venu, R.C.; Gowda, M.; Bellizzi, M.; Hu, J.; Liu, W.; Ebbole, D.; Meyers, B.; Mitchell, T.; et al. Identification and Characterization of In planta–Expressed Secreted Effector Proteins from Magnaporthe oryzae That Induce Cell Death in Rice. Mol. Plant-Microbe Interact. 2013, 26, 191–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonio, Á.; Fernández-Ortuño, D.; Vicente, A.; Pérez-García, A. A haustorial-expressed lytic polysaccharide monooxygenase from the cucurbit powdery mildew pathogen Podosphaera xanthii contributes to the suppression of chitin-triggered immunity. Mol. Plant Pathol. 2021, 22, 580–601. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Sánchez, I.; Pejenaute-Ochoa, M.D.; Navarrete, B.; Barrales, R.R.; Ibeas, J.I. Ustilago maydis Secreted Endo-Xylanases Are Involved in Fungal Filamentation and Proliferation on and Inside Plants. J. Fungi 2021, 7, 1081. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Li, T.; Shi, X.; Saleem, M.; Li, B.; Liang, W.; Wang, C. Deletion of Endo-β-1,4-Xylanase VmXyl1 Impacts the Virulence of Valsa mali in Apple Tree. Front. Plant Sci. 2018, 9, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wang, Z.; Li, J.; Wang, Y.; Yuan, J.; Zhan, J.; Wang, P.; Lin, Y.; Li, F.; Ge, X. Verticillium dahliae secreted protein Vd424Y is required for full virulence, targets the nucleus of plant cells, and induces cell death. Mol. Plant Pathol. 2021, 22, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Okuda, S.; Ikeda, K.; Okuno, T.; Takano, Y. LAC2 Encoding a Secreted Laccase Is Involved in Appressorial Melanization and Conidial Pigmentation in Colletotrichum orbiculare. Mol. Plant-Microbe Interact. 2012, 25, 1552–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, A.A.; Park, S.-Y.; Sadat, A.; Kim, S.; Choi, J.; Jeon, J.; Lee, Y.-H. Systematic characterization of the peroxidase gene family provides new insights into fungal pathogenicity in Magnaporthe oryzae. Sci. Rep. 2015, 5, 11831. [Google Scholar] [CrossRef] [Green Version]
- López-Pérez, M.; Ballester, A.-R.; González-Candelas, L. Identification and functional analysis of Penicillium digitatum genes putatively involved in virulence towards citrus fruit. Mol. Plant Pathol. 2014, 16, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Sun, C.; Liu, Y.; Feng, H.; Chang, H.; Cao, S.; Li, G.; Yang, S.; Hou, J.; Zhu-Salzman, K.; et al. Transcriptome analysis and functional validation reveal a novel gene, BcCGF1, that enhances fungal virulence by promoting infection-related development and host penetration. Mol. Plant Pathol. 2020, 21, 834–853. [Google Scholar] [CrossRef] [Green Version]
- Garfoot, A.L.; Shen, Q.; Wüthrich, M.; Klein, B.S.; Rappleye, C.A. The Eng1 β-Glucanase Enhances Histoplasma Virulence by Reducing β-Glucan Exposure. Mbio 2016, 7, e01388-15. [Google Scholar] [CrossRef] [Green Version]
- Jashni, M.K.; Dols, I.H.; Iida, Y.; Boeren, S.; Beenen, H.G.; Mehrabi, R.; Collemare, J.; De Wit, P.J. Synergistic Action of a Metalloprotease and a Serine Protease from Fusarium oxysporum f. sp. lycopersici Cleaves Chitin-Binding Tomato Chitinases, Reduces Their Antifungal Activity, and Enhances Fungal Virulence. Mol. Plant Microbe Interact. 2015, 28, 996–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitoh, H.; Fujisawa, S.; Ito, A.; Mitsuoka, C.; Berberich, T.; Tosa, Y.; Asakura, M.; Takano, Y.; Terauchi, R. SPM1 encoding a vacuole-localized protease is required for infection-related autophagy of the rice blast fungus Magnaporthe oryzae. FEMS Microbiol. Lett. 2009, 300, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira-Garcia, E.; Deising, H.B. The Glycosylphosphatidylinositol Anchor Biosynthesis Genes GPI12, GAA1, and GPI8 Are Essential for Cell-Wall Integrity and Pathogenicity of the Maize Anthracnose Fungus Colletotrichum graminicola. Mol. Plant-Microbe Interact. 2016, 29, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Park, G.; Choi, W.; Zheng, L.; Dean, R.A.; Xu, J.-R. Two Novel Fungal Virulence Genes Specifically Expressed in Appressoria of the Rice Blast Fungus. Plant Cell 2002, 14, 2107–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, G.; Aminedi, R.; Saxena, D.; Gupta, A.; Banerjee, P.; Jain, D.; Chandran, D. Effector mining from the Erysiphe pisi haustorial transcriptome identifies novel candidates involved in pea powdery mildew pathogenesis. Mol. Plant Pathol. 2019, 20, 1506–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genome Size (Mb) | Total Proteins | Refined Secretome | % of Secretome | Cazymes | Proteases | Putative Effectors |
|---|---|---|---|---|---|---|---|
| T. punctulata | 28.1 | 5296 a | 197 | 3.71 | 75 | 24 | 47 |
| T. ethacetica | 29.4 | 7079 b | 215 | 3.03 | 54 | 17 | 68 |
| T. cerberus | 28.6 | 5591 b | 150 | 2.68 | 43 | 5 | 59 |
| T. musarum | 28.4 | 6801 b | 172 | 2.52 | 46 | 16 | 56 |
| T. euricoi | 29.6 | 7004 b | 194 | 2.76 | 56 | 8 | 62 |
| T. populi | 23.9 | 6220 b | 156 | 2.50 | 42 | 7 | 55 |
| CAZy Family | Protein Id | PFAM Id | E.C. Number | Enzyme Name | Substrate |
|---|---|---|---|---|---|
| AA1 | KKA28993.1 | PF07731.17 | 1.10.3.2 | Laccase | lignin |
| KKA29108.1 | PF07732.18 | 1.10.3.2 | Laccase | lignin | |
| KKA30055.1 | PF07732.18 | 1.10.3.2 | Laccase | lignin | |
| AA16 | KKA29756.1 | PF03067.18 | na | Lytic cellulose monooxygenase | cellulose |
| AA2 | KKA28039.1 | PF00141.26 | 1.11.1.16 | versatile peroxidase | lignin |
| KKA29461.1 | PF01822.22 | na | peroxidase | lignin | |
| AA3 | KKA28521.1 | PF16010.8 | 1.1.99.18 | Cellobiose dehydrogenase | cellulose |
| KKA30608.1 | PF16010.8 | 1.1.99.18 | Cellobiose dehydrogenase | cellulose | |
| KKA31082.1 | PF00732.22 | 1.1.3.7 | aryl alcohol oxidase | lignin | |
| AA5 | KKA28638.1 | PF01822.22 | 1.1.3.- | Oxidase with oxygen as acceptor | lignin |
| AA7 | KKA27659.1 | PF01565.26 | 1.1.3.- | glucooligosaccharide oxidase | cellobiose |
| KKA30937.1 | PF01565.26 | 1.1.3.- | glucooligosaccharide oxidase | cellobiose | |
| AA9 | KKA27328.1 | PF03443.17 | 1.14.99.54 | lytic cellulose monooxygenase | cellulose |
| KKA28212.1 | PF03443.17 | 1.14.99.54 | lytic cellulose monooxygenase | cellulose | |
| KKA28497.1 | PF03443.17 | 1.14.99.54 | lytic cellulose monooxygenase | cellulose | |
| KKA25992.1 | PF03443.17 | 1.14.99.54 | endo-β-1,4-glucanase | cellulose | |
| KKA25994.1 | PF03443.17 | 1.14.99.54 | endo-β-1,4-glucanase | cellulose | |
| KKA29038.1 | PF03443.17 | 1.14.99.54 | lytic cellulose monooxygenase | cellulose | |
| KKA29219.1 | PF03443.17 | 1.14.99.54 | lytic cellulose monooxygenase | cellulose | |
| CE4 | KKA26186.1 | PF01522.24 | 3.5.1.41 | chitin deacetylase | chitin |
| KKA27343.1 | PF01522.24 | 3.5.1.41 | chitin deacetylase | chitin | |
| CE5 | KKA30377.1 | PF01083.25 | 3.1.1.72 | acetyl xylan esterase | Hemi cellulose (xylan) |
| KKA30382.1 | PF01083.25 | 3.1.1.72 | acetyl xylan esterase | Hemi cellulose (xylan) | |
| GH03 | KKA26832.1 | PF01915.25 | 3.2.1.21 | β-glucosidase | Cellulose, Hemi cellulose |
| KKA30767.1 | PF01915.25 | 3.2.1.21 | β-glucosidase | Cellulose, Hemi cellulose | |
| GH05 | KKA26007.1 | PF00150.21 | 3.2.1.4 | endo-β-1,4-glucanase | cellulose |
| KKA26778.1 | PF00150.21 | 3.2.1.4 | endo-β-1,4-glucanase | cellulose | |
| KKA28137.1 | PF00150.21 | 3.2.1.4 | endo-β-1,4-glucanase | cellulose | |
| GH07 | KKA26295.1 | PF00840.23 | 3.2.1.176 | cellulose 1,4-beta-cellobiosidase | cellulose |
| KKA28489.1 | PF00840.23 | 3.2.1.4 | endo-β-1,4-glucanase | cellulose | |
| GH10 | KKA27891.1 | PF00331.23 | 3.2.1.8 | endo-1,4-β-xylanase | Hemi cellulose (xylan) |
| KKA29568.1 | PF00331.23 | 3.2.1.8 | endo-1,4-β-xylanase | Hemi cellulose (xylan) | |
| GH11 | KKA29107.1 | PF00457.20 | 3.2.1.8 | endo-β-1,4-xylanase | Hemi cellulose (xylan) |
| KKA30007.1 | PF00457.20 | 3.2.1.8 | endo-β-1,4-xylanase | Hemi cellulose (xylan) | |
| GH115 | KKA28239.1 | PF15979.8 | 3.2.1.131 | xylan α-1,2-glucuronidase | Hemi cellulose (xylan) |
| GH125 | KKA28305.1 | PF06824.14 | - | exo-α-1,6-mannosidase | mannan (Hemi cellulose) |
| GH128 | KKA29105.1 | PF11790.11 | - | β-1,3-glucanase | β-glucans |
| GH13 | KKA30803.1 | PF00128.27 | 3.2.1.1 | α-amylase | starch |
| GH131 | KKA28951.1 | PF18271.4 | 3.2.1.21 | endo-β-1,4-glucanase | starch |
| KKA29646.1 | PF18271.4 | 3.2.1.21 | endo-β-1,4-glucanase | starch | |
| GH132 | KKA26122.1 | PF03856.16 | 3.2.1.- | Beta-glucosidase | starch |
| GH15 | KKA29558.1 | PF00723.24 | 3.2.1.3 | glucoamylase | starch |
| GH16 | KKA26151.1 | PF00722.24 | 3.2.1.73 | licheninase | Starch |
| KKA27451.1 | PF00722.24 | 3.2.1.73 | licheninase | Starch | |
| KKA28499.1 | PF00722.24 | 3.2.1.73 | licheninase | Starch | |
| KKA30944.1 | PF00722.24 | 3.2.1.181 | endo-β-1,3-galactanase | Pectin (Arabinogalactan) | |
| GH18 | KKA26416.1 | PF03009.20 | N | Polysaccharides | |
| KKA27515.1 | PF00704.31 | 3.2.1.14 | chitinase | chitin | |
| KKA30054.1 | PF00704.31 | 3.2.1.14 | chitinase | chitin | |
| KKA30697.1 | PF00704.31 | 3.2.1.14 | chitinase | chitin | |
| GH20 | KKA30299.1 | PF00728.25 | 3.2.1.52 | β-hexosaminidase | Polysaccharides |
| GH28 | KKA31208.1 | PF00295.20 | 3.2.1.15 | polygalacturonase | Pectin |
| GH30 | KKA27339.1 | PF14587.9 | 3.2.1.164 | endo-β-1,6-galactanase | Pectin (Arabinogalactan) |
| KKA29858.1 | PF02057.18 | 3.2.1.164 | endo-β-1,6-galactanase | Pectin (Arabinogalactan) | |
| GH32 | KKA28220.1 | PF00251.23 | 3.2.1.26 | invertase | sucrose |
| GH37 | KKA30799.1 | PF01204.21 | 3.2.1.28 | α,α-trehalase | Trehalose |
| GH38 | KKA26248.1 | PF01532.23 | 3.2.1.24 | α-mannosidase | Hemi cellulose (mannan) |
| GH43 | KKA27803.1 | PF04616.17 | 3.2.1.99 | endo-α-1,5-L-arabinanase | Hemi cellulose (xylan) |
| KKA28970.1 | PF04616.17 | 3.2.1.145 | exo-β-1,3-galactanase | Hemi cellulose (xylan) | |
| KKA29859.1 | PF04616.17 | 3.2.1.145 | exo-β-1,3-galactanase | Hemi cellulose (xylan) | |
| KKA30545.1 | PF04616.17 | 3.2.1.145 | exo-β-1,3-galactanase | Hemi cellulose (xylan) | |
| GH45 | KKA28018.1 | PF02015.19 | 3.2.1.4 | endo-β-1,4-glucanase | cellulose |
| GH51 | KKA29147.1 | PF06964.15 | 3.2.1.55 | α-L-arabinofuranosidase | Hemicellulose |
| GH53 | KKA29651.1 | PF07745.16 | 3.2.1.89 | endo-β-1,4-galactanase | Hemicellulose |
| GH55 | KKA30026.1 | PF12708.10 | 3.2.1.58 | glucan β-1,3-glucosidase | callose |
| KKA30509.1 | PF12708.10 | 3.2.1.58 | glucan β-1,3-glucosidase | callose | |
| GH64 | KKA27366.1 | PF16483.8 | 3.2.1.39 | glucan endo-1,3-β-D-glucosidase | callose |
| GH76 | KKA26192.1 | PF03663.17 | 3.2.1.101 | α-1,6-mannanase | mannan |
| GH78 | KKA27604.1 | PF17390.5 | 3.2.1.40 | α-L-rhamnosidase | mannan |
| GH93 | KKA30496.1 | PF06964.15 | 3.2.1.- | exo-α-L-1,5-arabinanase | Hemicellulose |
| GT4 | KKA26489.1 | PF04488.18 | 2.4.1.257 | α-1,6-mannosyltransferase | mannan |
| PL1 | KKA26877.1 | PF00544.22 | 4.2.2.10 | pectin lyase | pectin |
| KKA27238.1 | PF00544.22 | 4.2.2.10 | pectin lyase | pectin | |
| PL3 | KKA30830.1 | PF03211.16 | 4.2.2.2 | pectate lyase | pectin |
| PL4 | KKA28462.1 | PF09284.13 | 4.2.2.23 | rhamnogalacturonan endolyase | pectin |
| CAZyme Class | Class Members | Thielaviopsis Species | |||||
|---|---|---|---|---|---|---|---|
| T. punctulata | T. ethacetica | T. cerberus | T. euricoi | T. musarum | T. populi | ||
| Auxiliary Activities | AA1 | 3 | 1 | 1 | 2 | 1 | 2 |
| AA2 | 2 | 0 | 0 | 1 | 0 | 0 | |
| AA3 | 3 | 3 | 0 | 3 | 2 | 1 | |
| AA5 | 1 | 1 | 1 | 1 | 1 | 1 | |
| AA7 | 2 | 0 | 0 | 0 | 2 | 0 | |
| AA8 | 0 | 1 | 1 | 1 | 2 | ||
| AA9 | 7 | 5 | 0 | 5 | 5 | 3 | |
| AA11 | 0 | 2 | 1 | 2 | 2 | 1 | |
| AA12 | 0 | 1 | 0 | 0 | 0 | 0 | |
| AA16 | 1 | 0 | 0 | 0 | 0 | 0 | |
| Carbohydrate esterases | CE1 | 0 | 1 | 1 | 2 | 2 | 1 |
| CE3 | 0 | 1 | 0 | 0 | 0 | 0 | |
| CE4 | 2 | 1 | 1 | 1 | 0 | 0 | |
| CE5 | 2 | 1 | 2 | 1 | 1 | 0 | |
| Glycoside Hydrolases | GH03 | 2 | 2 | 2 | 2 | 2 | |
| GH05 | 3 | 0 | 0 | 0 | 1 | 1 | |
| GH06 | 0 | 0 | 0 | 1 | 0 | 0 | |
| GH07 | 2 | 1 | 1 | 0 | 1 | 0 | |
| GH10 | 2 | 1 | 1 | 2 | 2 | 1 | |
| GH11 | 2 | 3 | 3 | 3 | 2 | 1 | |
| GH12 | 0 | 0 | 1 | 0 | 0 | 0 | |
| GH13 | 0 | 0 | 0 | 0 | 0 | 1 | |
| GH15 | 1 | 0 | 0 | 0 | 0 | 1 | |
| GH16 | 4 | 5 | 5 | 5 | 3 | 3 | |
| GH17 | 4 | 3 | 4 | 3 | 4 | 4 | |
| GH20 | 1 | 1 | 1 | 1 | 1 | 1 | |
| GH28 | 1 | 0 | 1 | 0 | 0 | 0 | |
| GH30 | 2 | 2 | 1 | 1 | 1 | 0 | |
| GH31 | 0 | 0 | 0 | 0 | 1 | 0 | |
| GH32 | 1 | 1 | 1 | 1 | 1 | 1 | |
| GH37 | 1 | 0 | 1 | 0 | 0 | 0 | |
| GH38 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GH43 | 4 | 4 | 4 | 5 | 3 | 2 | |
| GH45 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GH51 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GH53 | 1 | 0 | 0 | 0 | 0 | 1 | |
| GH55 | 2 | 1 | 1 | 1 | 0 | 1 | |
| GH63 | 0 | 1 | 0 | 1 | 1 | 0 | |
| GH64 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GH72 | 0 | 1 | 2 | 2 | 2 | 3 | |
| GH74 | 0 | 0 | 1 | 1 | 0 | 0 | |
| GH76 | 1 | 2 | 2 | 2 | 2 | 2 | |
| GH78 | 1 | 0 | 1 | 0 | 0 | 0 | |
| GH92 | 0 | 1 | 0 | 1 | 1 | 1 | |
| GH93 | 1 | 1 | 0 | 0 | 1 | 0 | |
| GH115 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GH125 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GH128 | 1 | 0 | 0 | 0 | 0 | 0 | |
| GH131 | 2 | 0 | 0 | 0 | 0 | 1 | |
| GH132 | 1 | 0 | 0 | 0 | 0 | 0 | |
| Glycosyl Transferases | GT4 | 1 | 0 | 0 | 0 | 0 | 0 |
| GT8 | 0 | 0 | 0 | 1 | 0 | 1 | |
| GT32 | 0 | 0 | 0 | 0 | 0 | 1 | |
| GT34 | 0 | 1 | 0 | 1 | 1 | 0 | |
| GT61 | 0 | 1 | 0 | 0 | 0 | 0 | |
| Polysaccharide Lyases | PL1 | 2 | 2 | 1 | 1 | 1 | 2 |
| PL3 | 1 | 1 | 0 | 1 | 0 | 1 | |
| PL4 | 1 | 0 | 0 | 0 | 0 | 1 | |
| Protein Id | Length | Cysteines | Domain1 | Name | Domain2 | Name |
|---|---|---|---|---|---|---|
| KKA28270.1 | 275 | 4 | PF13640 | 2OG-FeII_Oxy_3 | ||
| KKA29775.1 | 392 | 4 | PF12697 | Abhydrolase_6 | ||
| KKA30577.1 | 359 | 7 | PF00733 | Asn_synthase | ||
| KKA26543.1 | 316 | 5 | PF00967 | Barwin | ||
| KKA27620.1 | 348 | 4 | PF00188 | CAP | ||
| KKA28836.1 | 133 | 4 | PF07249 | Cerato-platanin | ||
| KKA26938.1 | 356 | 6 | PF00127 | Copper-bind | ||
| KKA28390.1 | 177 | 2 | PF00127 | Copper-bind | ||
| KKA28890.1 | 234 | 4 | PF00190 | Cupin_1 | ||
| KKA27979.1 | 369 | 3 | PF00775 | Dioxygenase_C | ||
| KKA28872.1 | 203 | 4 | PF07510 | DUF1524 | ||
| KKA26687.1 | 278 | 4 | PF10057 | DUF2294 | PF13640 | 2OG-FeII_Oxy_3 |
| KKA27329.1 | 237 | 3 | PF10901 | DUF2690 | PF07883 | Cupin_2 |
| KKA26724.1 | 253 | 4 | PF11693 | DUF2990 | PF11937 | DUF3455 |
| KKA26892.1 | 247 | 3 | PF11937 | DUF3455 | ||
| KKA27739.1 | 118 | 7 | PF15371 | DUF4599 | ||
| KKA29708.1 | 260 | 7 | PF05359 | DUF748 | ||
| KKA31236.1 | 179 | 4 | PF02221 | E1_DerP2_DerF2 | ||
| KKA26926.1 | 345 | 6 | PF11327 | Egh16-like | PF09716 | ETRAMP |
| KKA26947.1 | 171 | 6 | PF11327 | Egh16-like | ||
| KKA27553.1 | 158 | 4 | PF11327 | Egh16-like | ||
| KKA27672.1 | 272 | 4 | PF11327 | Egh16-like | ||
| KKA29410.1 | 381 | 8 | PF11327 | Egh16-like | PF12230 | PRP21_like_P |
| KKA25960.1 | 307 | 2 | PF03372 | Exo_endo_phos | ||
| KKA29592.1 | 316 | 4 | PF13668 | Ferritin_2 | PF06140 | Ifi-6-16 |
| KKA29712.1 | 289 | 3 | PF07554 | FIVAR | PF00775 | Dioxygenase_C |
| KKA26457.1 | 200 | 2 | PF00254 | FKBP_C | ||
| KKA27476.1 | 255 | 2 | PF05630 | NPP1 | ||
| KKA29596.1 | 296 | 2 | PF02585 | PIG-L | ||
| KKA28719.1 | 290 | 8 | PF16670 | PI-PLC-C1 | ||
| KKA28033.1 | 266 | 2 | PF13883 | Pyrid_oxidase_2 | ||
| KKA29951.1 | 304 | 5 | PF02265 | S1-P1_nuclease | ||
| KKA28667.1 | 316 | 5 | PF12138 | Spherulin4 | PF15862 | Coilin_N |
| KKA29465.1 | 127 | 4 | PF14558 | TRP_N | ||
| KKA26744.1 | 291 | 2 | No domain | |||
| KKA27434.1 | 157 | 5 | No domain | |||
| KKA27537.1 | 98 | 6 | No domain | |||
| KKA27913.1 | 179 | 4 | No domain | |||
| KKA28484.1 | 339 | 8 | No domain | |||
| KKA29066.1 | 211 | 6 | No domain | |||
| KKA29758.1 | 78 | 7 | No domain | |||
| KKA30719.1 | 189 | 2 | No domain | |||
| KKA30870.1 | 169 | 4 | No domain | |||
| KKA30907.1 | 140 | 2 | No domain | |||
| KKA30938.1 | 371 | 8 | No domain | |||
| KKA31074.1 | 140 | 2 | No domain | |||
| KKA31087.1 | 113 | 8 | No domain |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chellappan, B.V.; El-Ganainy, S.M.; Alrajeh, H.S.; Al-Sheikh, H. In Silico Characterization of the Secretome of the Fungal Pathogen Thielaviopsis punctulata, the Causal Agent of Date Palm Black Scorch Disease. J. Fungi 2023, 9, 303. https://doi.org/10.3390/jof9030303
Chellappan BV, El-Ganainy SM, Alrajeh HS, Al-Sheikh H. In Silico Characterization of the Secretome of the Fungal Pathogen Thielaviopsis punctulata, the Causal Agent of Date Palm Black Scorch Disease. Journal of Fungi. 2023; 9(3):303. https://doi.org/10.3390/jof9030303
Chicago/Turabian StyleChellappan, Biju Vadakkemukadiyil, Sherif Mohamed El-Ganainy, Hind Salih Alrajeh, and Hashem Al-Sheikh. 2023. "In Silico Characterization of the Secretome of the Fungal Pathogen Thielaviopsis punctulata, the Causal Agent of Date Palm Black Scorch Disease" Journal of Fungi 9, no. 3: 303. https://doi.org/10.3390/jof9030303