
Nephroprotective Effects of Two Ganoderma Species Methanolic Extracts in an In Vitro Model of Cisplatin Induced Tubulotoxicity

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Culture Media
2.2. Sample Preparation
2.3. Cell Culture and Treatments
2.4. Cell Viability Assay (CCK-8)
2.5. Scavenger Activity
2.6. Flux Cytometry Analyses
2.6.1. Apoptosis Detection: Annexin V/PI Assay and Estimation of the p53, Caspase 3 and Cytochrome C Pro-Apoptotic Markers
2.6.2. Oxidative Stress Measurement
2.6.3. Anti-Inflammatory Potential Measurements
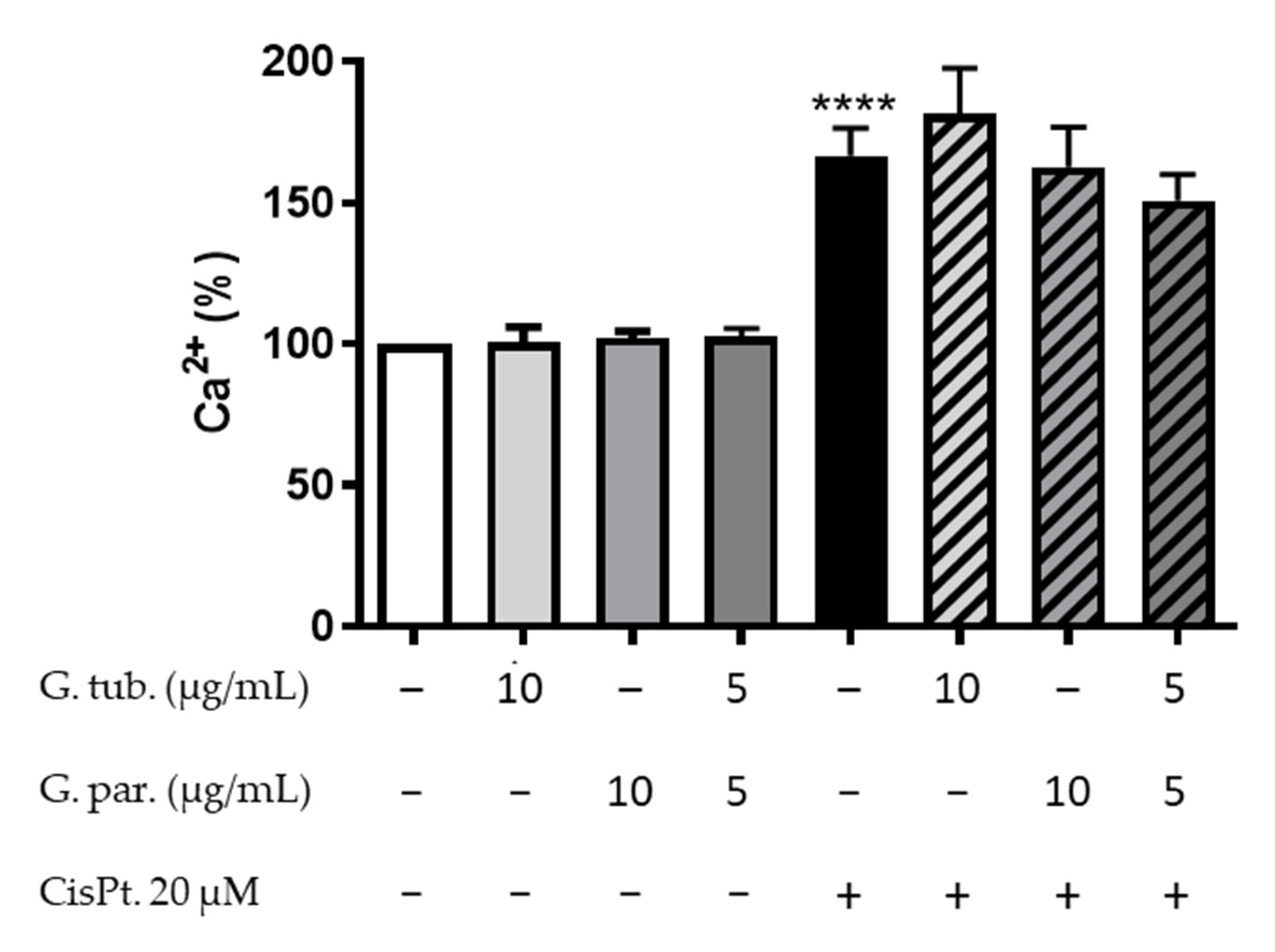
2.6.4. Intracytoplasmic Calcium Estimation
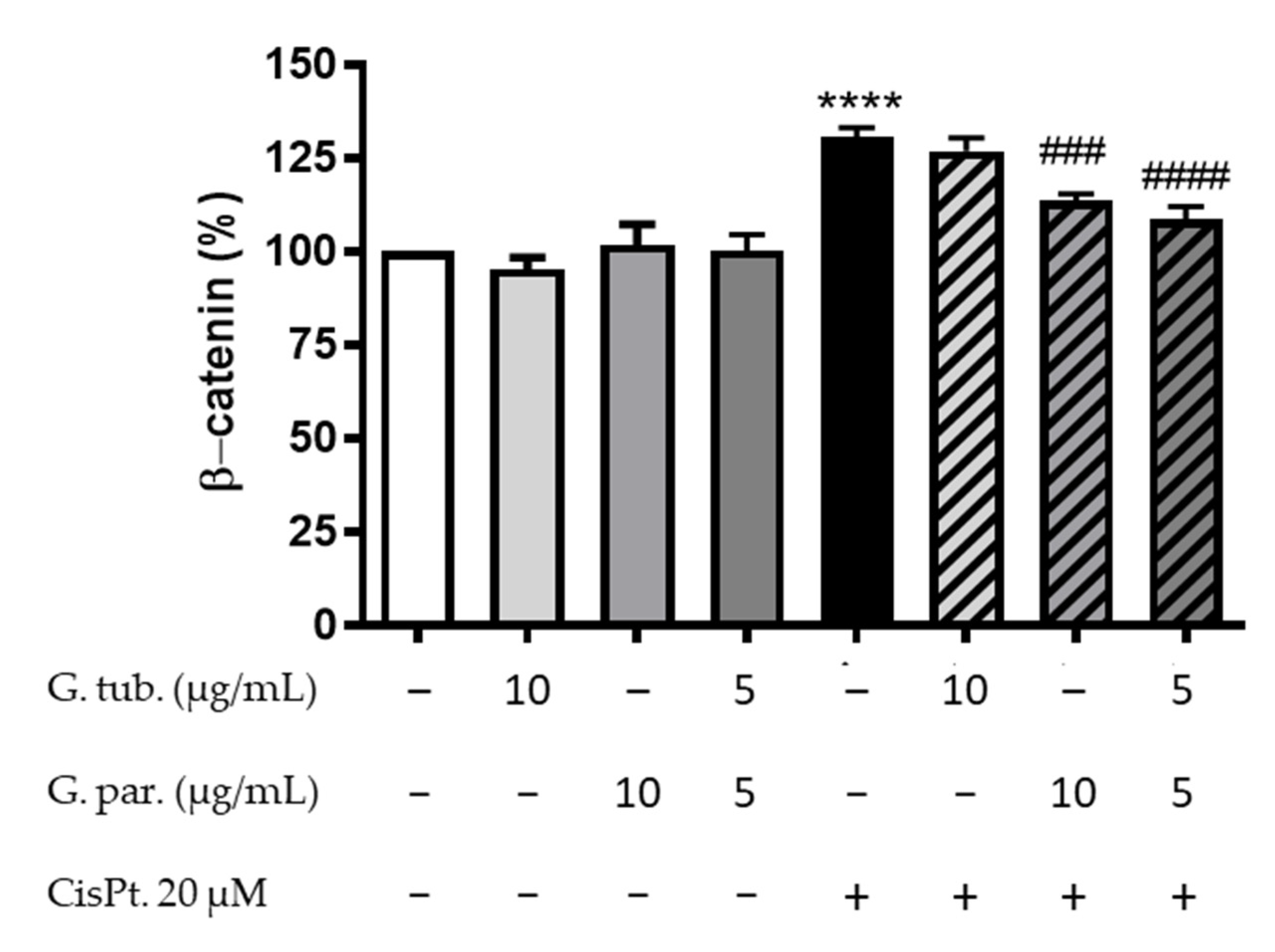
2.6.5. Intracytoplasmic β-catenin Estimation
2.7. Chemical Content
2.7.1. Phytochemical Screening
2.7.2. Total Phenolic Content
2.7.3. Total Triterpenoid Content
2.8. Statistical Analyses
3. Results
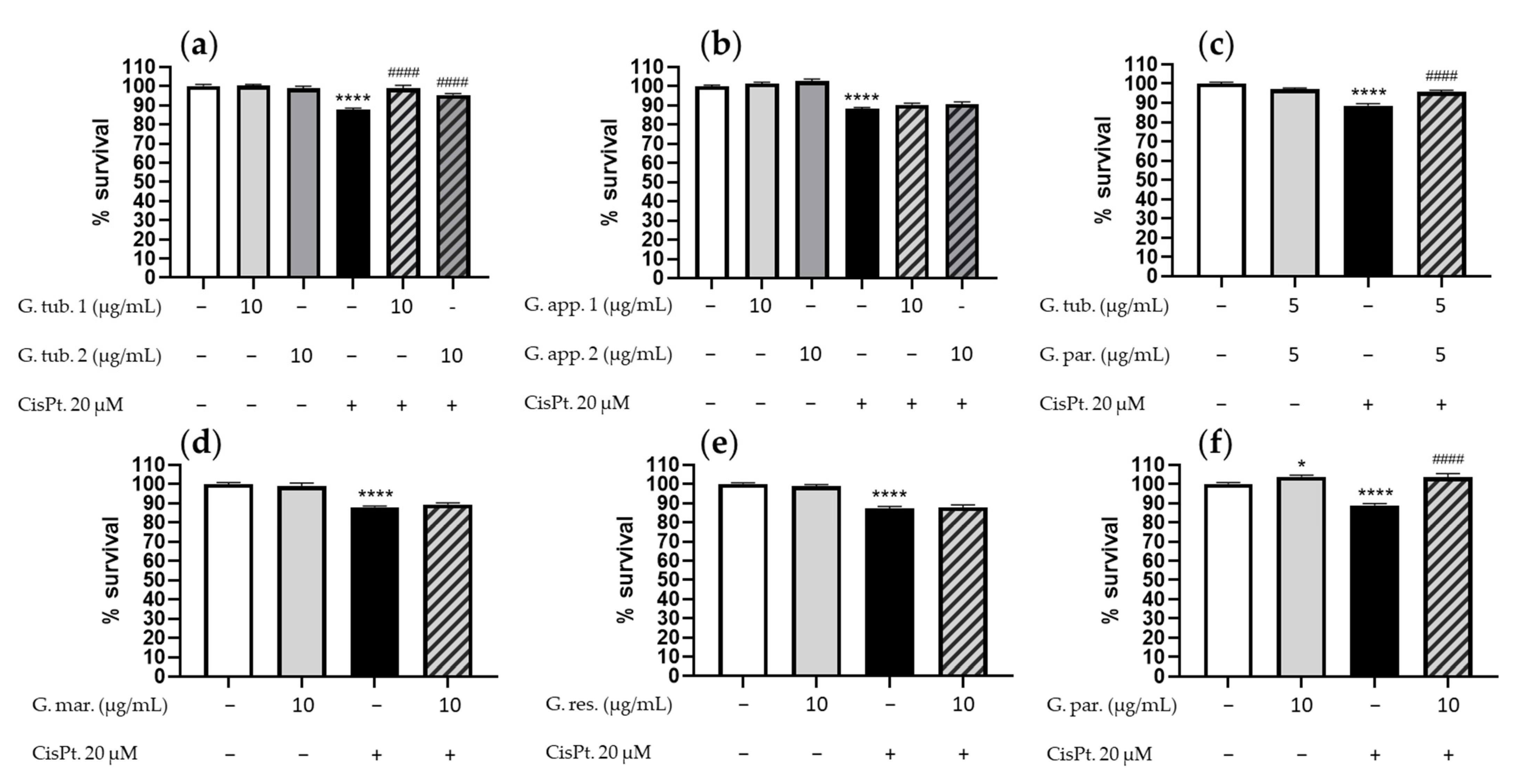
3.1. Cell Viability Assay (CCK-8)
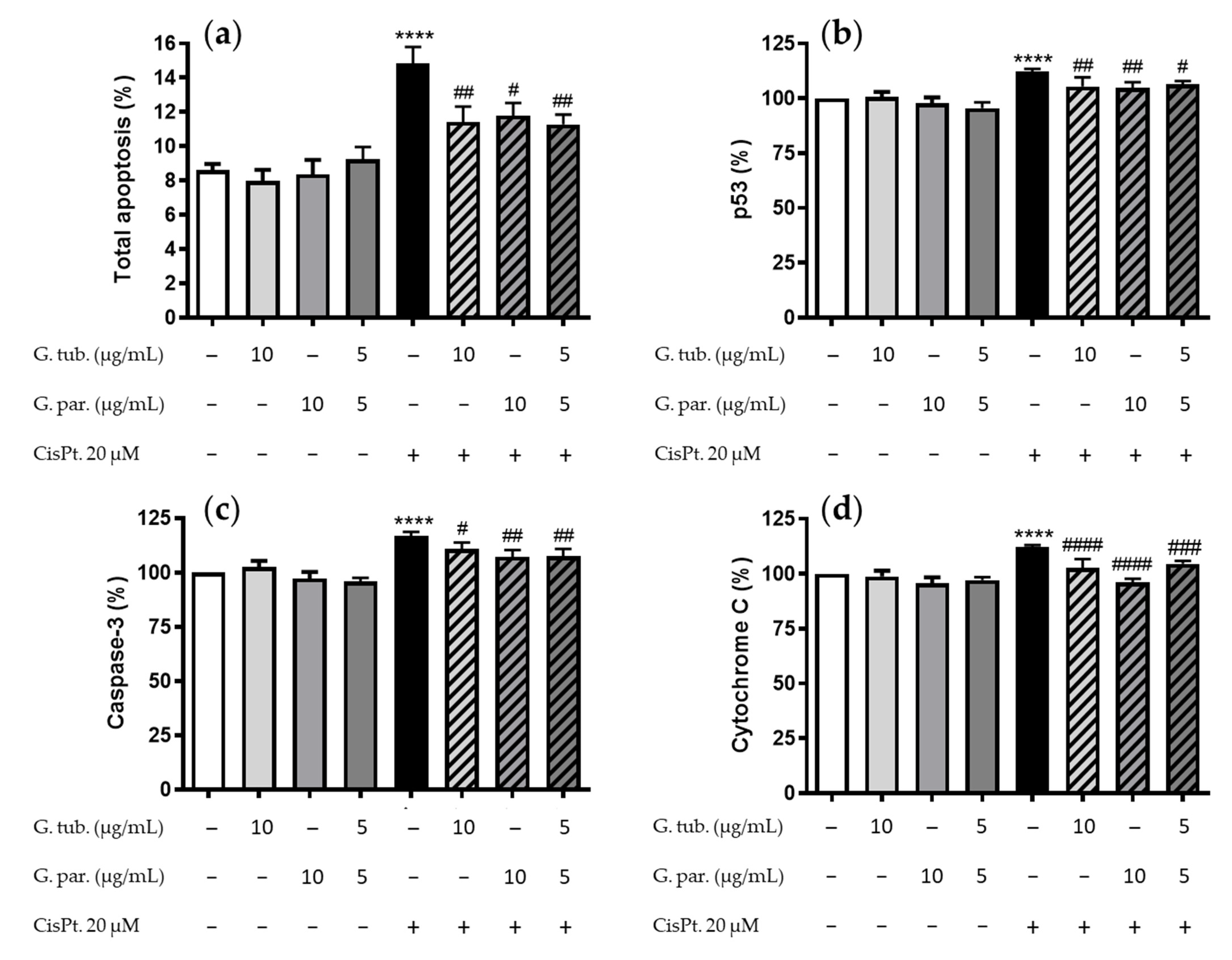
3.2. Apoptosis Detection: Annexin V/PI Assay and Estimation of the p53, Caspase 3, and Cytochrome C pro-Apoptotic Markers
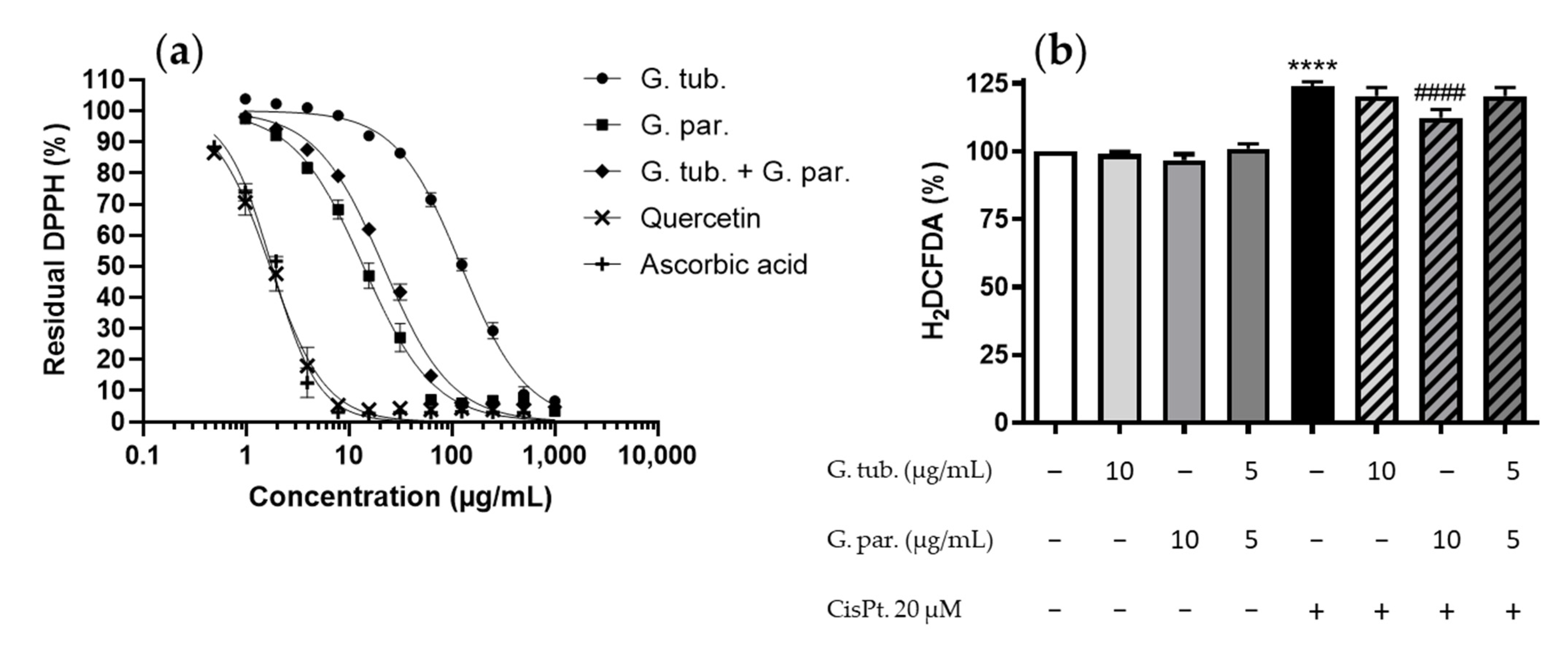
3.3. Antioxidant Potential: Scavenger Activity and ROS Determination (H2DCFDA Assay)
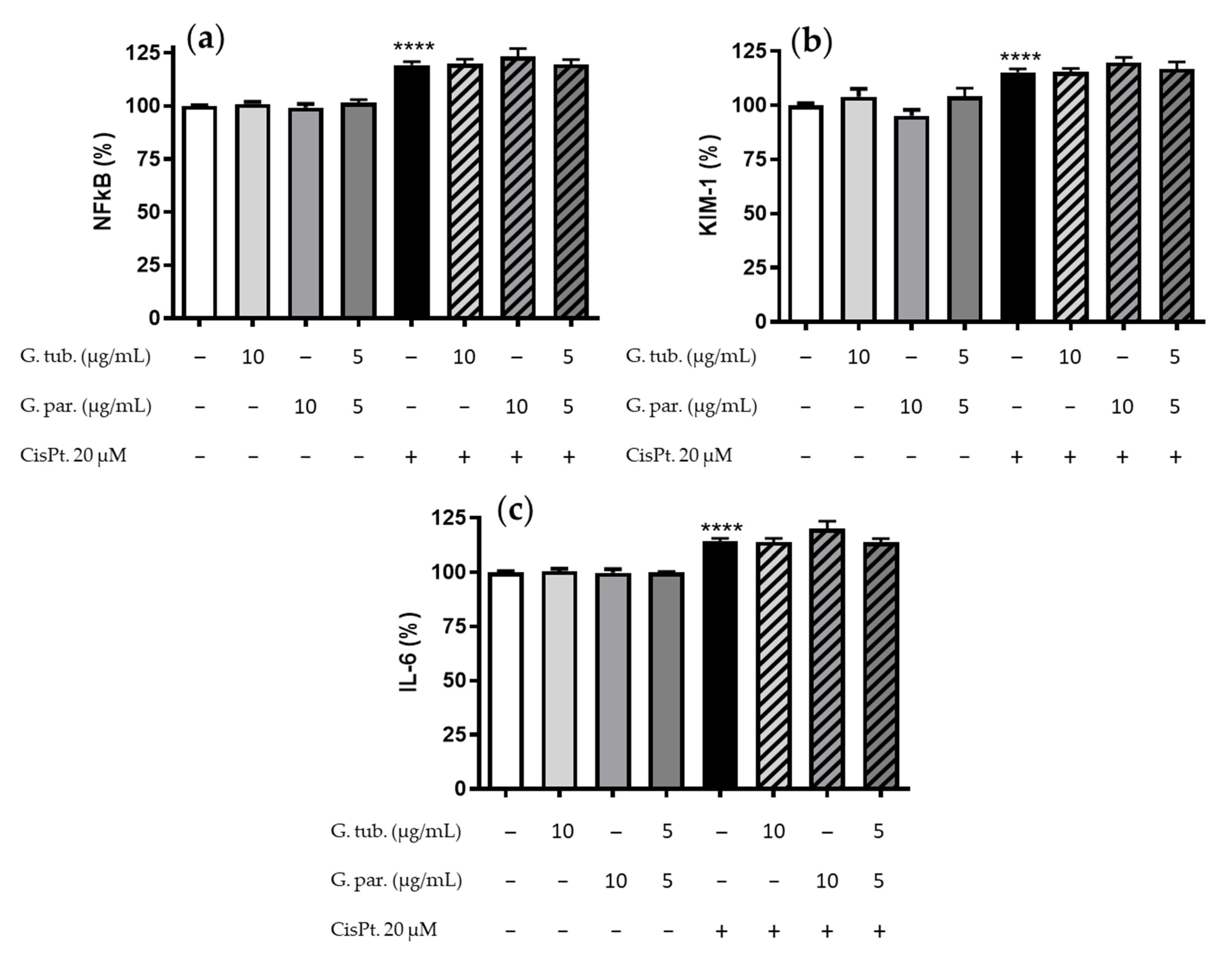
3.4. Anti-Inflammatory Measurement: Impact on the NFkB, KIM-1, IL-6 Markers
3.5. Intracytoplasmic Calcium Estimation
3.6. Intracytoplasmic β-Catenin Determination
3.7. Chemical Content
3.7.1. Phytochemical Screening
3.7.2. Total Phenolic and Triterpenoid Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, S. Cisplatin: The First Metal Based Anticancer Drug. Bioorganic Chem. 2019, 88, 102925. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular Mechanisms of Cisplatin Resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Tchounwou, P.B. Cisplatin in Cancer Therapy: Molecular Mechanisms of Action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Aldossary, S.A. Review on Pharmacology of Cisplatin: Clinical Use, Toxicity and Mechanism of Resistance of Cisplatin. Biomed. Pharmacol. J. 2019, 12, 7–15. [Google Scholar] [CrossRef]
- Manohar, S.; Leung, N. Cisplatin Nephrotoxicity: A Review of the Literature. J. Nephrol. 2018, 31, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.P.; Tadagavadi, R.K.; Ramesh, G.; Reeves, W.B. Mechanisms of Cisplatin Nephrotoxicity. Toxins 2010, 2, 2490–2518. [Google Scholar] [CrossRef] [PubMed]
- Crona, D.J.; Faso, A.; Nishijima, T.F.; McGraw, K.A.; Galsky, M.D.; Milowsky, M.I. A Systematic Review of Strategies to Prevent Cisplatin-Induced Nephrotoxicity. Oncologist 2017, 22, 609–619. [Google Scholar] [CrossRef]
- Hayati, F.; Hossainzadeh, M.; Shayanpour, S.; Abedi-Gheshlaghi, Z.; Beladi Mousavi, S.S. Prevention of Cisplatin Nephrotoxicity. J. Nephropharmacology 2016, 5, 57–60. [Google Scholar]
- Hwang, B.S.; Lee, D.; Choi, P.; Kim, K.S.; Choi, S.-J.; Song, B.G.; Kim, T.; Song, J.H.; Kang, K.S.; Ham, J. Renoprotective Effects of Hypoxylonol C and F Isolated from Hypoxylon Truncatum against Cisplatin-Induced Cytotoxicity in LLC-PK1 Cells. Int. J. Mol. Sci. 2018, 19, 948. [Google Scholar] [CrossRef]
- Lee, S.R.; Lee, D.; Lee, H.-J.; Noh, H.J.; Jung, K.; Kang, K.S.; Kim, K.H. Renoprotective Chemical Constituents from an Edible Mushroom, Pleurotus Cornucopiae in Cisplatin-Induced Nephrotoxicity. Bioorganic Chem. 2017, 71, 67–73. [Google Scholar] [CrossRef]
- Zhou, S.; Zhou, Y.; Yu, J.; Jiang, L.; Xiang, Y.; Wang, J.; Du, Y.; Cui, X.; Ge, F. A Neutral Polysaccharide from Ophiocordyceps Lanpingensis Restrains Cisplatin-Induced Nephrotoxicity. Food Sci. Nutr. 2021, 9, 3602–3616. [Google Scholar] [CrossRef] [PubMed]
- Nitha, B.; Janardhanan, K.K. Aqueous-Ethanolic Extract of Morel Mushroom Mycelium Morchella Esculenta, Protects Cisplatin and Gentamicin Induced Nephrotoxicity in Mice. Food Chem. Toxicol. 2008, 46, 3193–3199. [Google Scholar] [CrossRef] [PubMed]
- Mahran, Y.F.; Hassan, H.M. Ganoderma Lucidum Prevents Cisplatin-Induced Nephrotoxicity through Inhibition of Epidermal Growth Factor Receptor Signaling and Autophagy-Mediated Apoptosis. Oxid. Med. Cell. Longev. 2020, 2020, 4932587. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Yan, R.; Kang, J.; Chen, R. Chemical Components of Ganoderma. Adv. Exp. Med. Biol. 2019, 1181, 59–106. [Google Scholar] [CrossRef]
- Da, J.; Wu, W.-Y.; Hou, J.-J.; Long, H.-L.; Yao, S.; Yang, Z.; Cai, L.-Y.; Yang, M.; Jiang, B.-H.; Liu, X.; et al. Comparison of Two Officinal Chinese Pharmacopoeia Species of Ganoderma Based on Chemical Research with Multiple Technologies and Chemometrics Analysis. J. Chromatogr. A 2012, 1222, 59–70. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Ahmad, F.A.; Khan, M.I.; Alsayegh, A.A.; Wahab, S.; Alam, M.I.; Ahmed, F. Ganoderma Lucidum: A Potential Source to Surmount Viral Infections through β-Glucans Immunomodulatory and Triterpenoids Antiviral Properties. Int. J. Biol. Macromol. 2021, 187, 769–779. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Gabryelska, K.; Grabarczyk, M. Mushrooms of the Genus Ganoderma Used to Treat Diabetes and Insulin Resistance. Molecules 2019, 24, 4075. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Zaghlol, G.M.; El-Garawani, I.M.; Ahmed, E.F.; Rateb, M.E.; Abdel Moneim, A.E. Ganoderma Applanatum Secondary Metabolites Induced Apoptosis through Different Pathways: In Vivo and in Vitro Anticancer Studies. Biomed. Pharmacother. 2018, 101, 264–277. [Google Scholar] [CrossRef]
- Hanyu, X.; Lanyue, L.; Miao, D.; Wentao, F.; Cangran, C.; Hui, S. Effect of Ganoderma Applanatum Polysaccharides on MAPK/ERK Pathway Affecting Autophagy in Breast Cancer MCF-7 Cells. Int. J. Biol. Macromol. 2020, 146, 353–362. [Google Scholar] [CrossRef]
- Kou, R.-W.; Xia, B.; Wang, Z.-J.; Li, J.-N.; Yang, J.-R.; Gao, Y.-Q.; Yin, X.; Gao, J.-M. Triterpenoids and Meroterpenoids from the Edible Ganoderma Resinaceum and Their Potential Anti-Inflammatory, Antioxidant and Anti-Apoptosis Activities. Bioorganic Chem. 2022, 121, 105689. [Google Scholar] [CrossRef]
- El-Sherif, N.F.; Ahmed, S.A.; Ibrahim, A.K.; Habib, E.S.; El-Fallal, A.A.; El-Sayed, A.K.; Wahba, A.E. Ergosterol Peroxide from the Egyptian Red Lingzhi or Reishi Mushroom, Ganoderma Resinaceum (Agaricomycetes), Showed Preferred Inhibition of MCF-7 over MDA-MB-231 Breast Cancer Cell Lines. Int. J. Med. Mushrooms 2020, 22, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Qin, X.; Xu, Z.; Song, Y.; Jiang, H.; Wu, Y.; Ruan, H.; Chen, J. Comparison of Cytotoxicity Evaluation of Anticancer Drugs between Real-Time Cell Analysis and CCK-8 Method. ACS Omega 2019, 4, 12036–12042. [Google Scholar] [CrossRef] [PubMed]
- Hossen, S.M.M.; Hossain, M.S.; Yusuf, A.T.M.; Chaudhary, P.; Emon, N.U.; Janmeda, P. Profiling of Phytochemical and Antioxidant Activity of Wild Mushrooms: Evidence from the in Vitro Study and Phytoconstituent’s Binding Affinity to the Human Erythrocyte Catalase and Human Glutathione Reductase. Food Sci. Nutr. 2022, 10, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Ogidi, O.C.; Oyetayo, V.O. Phytochemical Property and Assessment of Antidermatophytic Activity of Some Selected Wild Macrofungi against Pathogenic Dermatophytes. Mycology 2016, 7, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.C. Trease and Evan’s Pharmacognosy, 13th ed.; Baillière Tendall: London, UK, 1989; ISBN 978-0-7020-1357-7. [Google Scholar]
- Rojo-Poveda, O.; Barbosa-Pereira, L.; El Khattabi, C.; Youl, E.N.H.; Bertolino, M.; Delporte, C.; Pochet, S.; Stévigny, C. Polyphenolic and Methylxanthine Bioaccessibility of Cocoa Bean Shell Functional Biscuits: Metabolomics Approach and Intestinal Permeability through Caco-2 Cell Models. Antioxidants 2020, 9, 1164. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, W.; Yin, L.; Yan, F.; Xu, Y.; Chen, F. Extraction Optimization of Total Triterpenoids from Jatropha Curcas Leaves Using Response Surface Methodology and Evaluations of Their Antimicrobial and Antioxidant Capacities. Electron. J. Biotechnol. 2015, 18, 88–95. [Google Scholar] [CrossRef]
- Sohn, S.-J.; Kim, S.Y.; Kim, H.S.; Chun, Y.-J.; Han, S.Y.; Kim, S.H.; Moon, A. In Vitro Evaluation of Biomarkers for Cisplatin-Induced Nephrotoxicity Using HK-2 Human Kidney Epithelial Cells. Toxicol. Lett. 2013, 217, 235–242. [Google Scholar] [CrossRef]
- Pillai, T.G.; John, M.; Sara Thomas, G. Prevention of Cisplatin Induced Nephrotoxicity by Terpenes Isolated from Ganoderma Lucidum Occurring in Southern Parts of India. Exp. Toxicol. Pathol. 2011, 63, 157–160. [Google Scholar] [CrossRef]
- Sheena, N.; Ajith, T.; Janardhanan, K. Prevention of Nephrotoxicity Induced by the Anticancer Drug Cisplatin, Using Ganoderma Lucidum, a Medicinal Mushroom Occurring in South India. Curr. Sci. 2003, 85, 478–482, undefined. [Google Scholar]
- Xue, L.; Wu, W.; You, Y.; Chen, H. A Small Molecule from Ganoderma lucidum Protects against Cisplatin-induced Kidney Injury via Suppressing NLRP3/Caspase-1 Related Pyroptosis. Congress Abstract. Kidney Int. Rep. 2020, 5, S217. [Google Scholar] [CrossRef]
- Xu, Z.Y.; Jin, C.J.; Zhou, C.C.; Wang, Z.Q.; Zhou, W.D.; Deng, H.B.; Zhang, M.; Su, W.; Cai, X.Y. Treatment of Advanced Non-Small-Cell Lung Cancer with Chinese Herbal Medicine by Stages Combined with Chemotherapy. J. Cancer Res. Clin. Oncol. 2011, 137, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.; Singh, A. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, X. Cytochrome C-Mediated Apoptosis. Annu. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef]
- Ow, Y.-L.P.; Green, D.R.; Hao, Z.; Mak, T.W. Cytochrome c: Functions beyond Respiration. Nat. Rev. Mol. Cell Biol. 2008, 9, 532–542. [Google Scholar] [CrossRef]
- Tuppy, H.; Kreil, G. Cytochrome c. In Encyclopedia of Biological Chemistry (Second Edition); Lennarz, W.J., Lane, M.D., Eds.; Academic Press: Waltham, MA, USA, 2013; pp. 599–601. ISBN 978-0-12-378631-9. [Google Scholar]
- Asadi, M.; Taghizadeh, S.; Kaviani, E.; Vakili, O.; Taheri-Anganeh, M.; Tahamtan, M.; Savardashtaki, A. Caspase-3: Structure, Function, and Biotechnological Aspects. Biotechnol. Appl. Biochem. 2021, 69, 1633–1645. [Google Scholar] [CrossRef]
- Jiang, M.; Qi, L.; Li, L.; Li, Y. The Caspase-3/GSDME Signal Pathway as a Switch between Apoptosis and Pyroptosis in Cancer. Cell Death Discov. 2020, 6, 112. [Google Scholar] [CrossRef]
- Teng, X.; Zhang, W.; Song, Y.; Wang, H.; Ge, M.; Zhang, R. Protective Effects of Ganoderma Lucidum Triterpenoids on Oxidative Stress and Apoptosis in the Spleen of Chickens Induced by Cadmium. Environ. Sci. Pollut. Res. 2019, 26, 23967–23980. [Google Scholar] [CrossRef]
- Cummings, B.S.; Schnellmann, R.G. Cisplatin-Induced Renal Cell Apoptosis: Caspase 3-Dependent and -Independent Pathways. J. Pharmacol. Exp. Ther. 2002, 302, 8–17. [Google Scholar] [CrossRef]
- Somogyi, A.; Rosta, K.; Pusztai, P.; Tulassay, Z.; Nagy, G. Antioxidant Measurements. Physiol. Meas. 2007, 28, R41–R55. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB System. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Nennig, S.E.; Schank, J.R. The Role of NFkB in Drug Addiction: Beyond Inflammation. Alcohol Alcohol. 2017, 52, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Bonventre, J.V. Kidney Injury Molecule-1 (KIM-1): A Urinary Biomarker and Much More. Nephrol. Dial. Transplant. 2009, 24, 3265–3268. [Google Scholar] [CrossRef]
- Han, W.K.; Bailly, V.; Abichandani, R.; Thadhani, R.; Bonventre, J.V. Kidney Injury Molecule-1 (KIM-1): A Novel Biomarker for Human Renal Proximal Tubule Injury. Kidney Int. 2002, 62, 237–244. [Google Scholar] [CrossRef]
- Gleichmann, M.; Mattson, M.P. Neuronal Calcium Homeostasis and Dysregulation. Antioxid. Redox Signal. 2011, 14, 1261–1273. [Google Scholar] [CrossRef]
- Carlström, M.; Wilcox, C.S.; Arendshorst, W.J. Renal Autoregulation in Health and Disease. Physiol. Rev. 2015, 95, 405–511. [Google Scholar] [CrossRef]
- Katoh, M. Multi-Layered Prevention and Treatment of Chronic Inflammation, Organ Fibrosis and Cancer Associated with Canonical WNT/β-Catenin Signaling Activation (Review). Int. J. Mol. Med. 2018, 42, 713–725. [Google Scholar] [CrossRef]
- Valenta, T.; Hausmann, G.; Basler, K. The Many Faces and Functions of β-Catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef]
- Silva-García, O.; Valdez-Alarcón, J.J.; Baizabal-Aguirre, V.M. Wnt/β-Catenin Signaling as a Molecular Target by Pathogenic Bacteria. Front. Immunol. 2019, 10, 2135. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Z.; Ji, H. Direct Targeting of β-Catenin in the Wnt Signaling Pathway: Current Progress and Perspectives. Med. Res. Rev. 2021, 41, 2109–2129. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xiao, L.; Sun, L.; Liu, F. Wnt/Beta-Catenin Signaling: A Promising New Target for Fibrosis Diseases. Physiol. Res. 2012, 61, 337–346. [Google Scholar] [CrossRef]
- Zhang, D.; Pan, Y.; Zhang, C.; Yan, B.; Yu, S.; Wu, D.; Shi, M.; Shi, K.; Cai, X.; Zhou, S.; et al. Wnt/β-Catenin Signaling Induces the Aging of Mesenchymal Stem Cells through Promoting the ROS Production. Mol. Cell. Biochem. 2013, 374, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Lin, J.; Yin, Y.; Zhao, J.; Sun, X.; Tang, K. Ganodermataceae: Natural Products and Their Related Pharmacological Functions. Am. J. Chin. Med. 2007, 35, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Singh, V.P.; Singh, N.K. A Review on Phytochemistry and Pharmacology of Medicinal as Well as Poisonous Mushrooms. Mini Rev. Med. Chem. 2018, 18, 1095–1109. [Google Scholar] [CrossRef]
- Hsin, I.-L.; Ou, C.-C.; Wu, M.-F.; Jan, M.-S.; Hsiao, Y.-M.; Lin, C.-H.; Ko, J.-L. GMI, an Immunomodulatory Protein from Ganoderma Microsporum, Potentiates Cisplatin-Induced Apoptosis via Autophagy in Lung Cancer Cells. Mol. Pharm. 2015, 12, 1534–1543. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xu, J.; Jiang, L.-L.; Huang, J.-Q.; Yan, J.-Y.; Chen, Y.-W.; Yang, Q. Improved Antitumor Activity of Cisplatin Combined with Ganoderma Lucidum Polysaccharides in U14 Cervical Carcinoma-Bearing Mice. Kaohsiung J. Med. Sci. 2019, 35, 222–229. [Google Scholar] [CrossRef]
- Jiang, Y.; Chang, Y.; Liu, Y.; Zhang, M.; Luo, H.; Hao, C.; Zeng, P.; Sun, Y.; Wang, H.; Zhang, L. Overview of Ganoderma Sinense Polysaccharide–an Adjunctive Drug Used during Concurrent Chemo/Radiation Therapy for Cancer Treatment in China. Biomed. Pharmacother. 2017, 96, 865–870. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Secondary Metabolite | G. tub. 1 | G. app. 1 | G. tub. 2 | G. par. | G. mar. | G. res. | G. app. 2 |
|---|---|---|---|---|---|---|---|
| Simple carbohydrates | + | ++ | + | ++ | − | − | ++ |
| Terpenes | + | + | + | + | + | + | + |
| Saponins | − | + | − | + | − | − | − |
| Alkaloids | − | − | − | − | − | − | − |
| Flavonoids | − | − | − | − | − | − | − |
| Tannins | − | ± | − | ± | − | − | ± |
| Total Phenolic Content 1 | Total Triterpenoid Content 2 | |
|---|---|---|
| G. tub. 1 | 47 ± 4 | 212 ± 12 |
| G. app. 1 | 121 ± 14 | 110 ± 13 |
| G. tub. 2 | 40 ± 3 | 256 ± 13 |
| G. par. | 201 ± 12 | 261 ± 16 |
| G. mar. | 26 ± 2 | 138 ± 12 |
| G. res. | 30 ± 2 | 169 ± 14 |
| G. app. 2 | 77 ± 7 | 157 ± 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinaeve, S.; Husson, C.; Antoine, M.-H.; Welti, S.; Stévigny, C.; Nortier, J. Nephroprotective Effects of Two Ganoderma Species Methanolic Extracts in an In Vitro Model of Cisplatin Induced Tubulotoxicity. J. Fungi 2022, 8, 1002. https://doi.org/10.3390/jof8101002
Sinaeve S, Husson C, Antoine M-H, Welti S, Stévigny C, Nortier J. Nephroprotective Effects of Two Ganoderma Species Methanolic Extracts in an In Vitro Model of Cisplatin Induced Tubulotoxicity. Journal of Fungi. 2022; 8(10):1002. https://doi.org/10.3390/jof8101002
Chicago/Turabian StyleSinaeve, Sébastien, Cécile Husson, Marie-Hélène Antoine, Stéphane Welti, Caroline Stévigny, and Joëlle Nortier. 2022. "Nephroprotective Effects of Two Ganoderma Species Methanolic Extracts in an In Vitro Model of Cisplatin Induced Tubulotoxicity" Journal of Fungi 8, no. 10: 1002. https://doi.org/10.3390/jof8101002