
In Vitro Effects of Leaf Extracts from Brassica rapa on the Growth of Two Entomopathogenic Fungi

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Species and Fungus-Based Products Used in This Study
2.2. Plant Material
2.3. Leaf Extract Preparation
2.4. Effect of Leaf Extracts on Fungal Growth
2.5. Gas Chromatography–Mass Spectrophotometry Analysis of Leaf Extracts
2.6. Data Analysis
3. Results
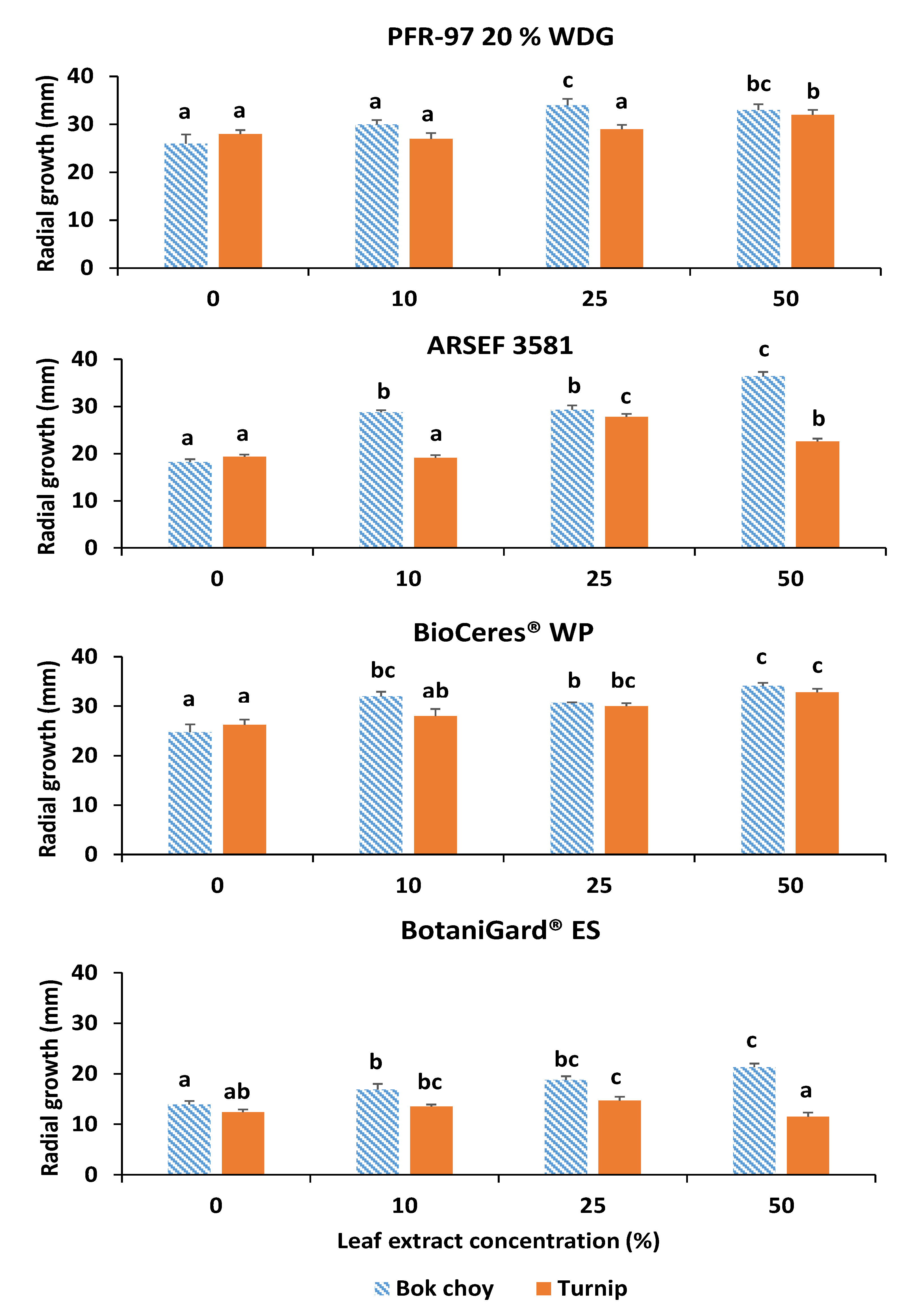
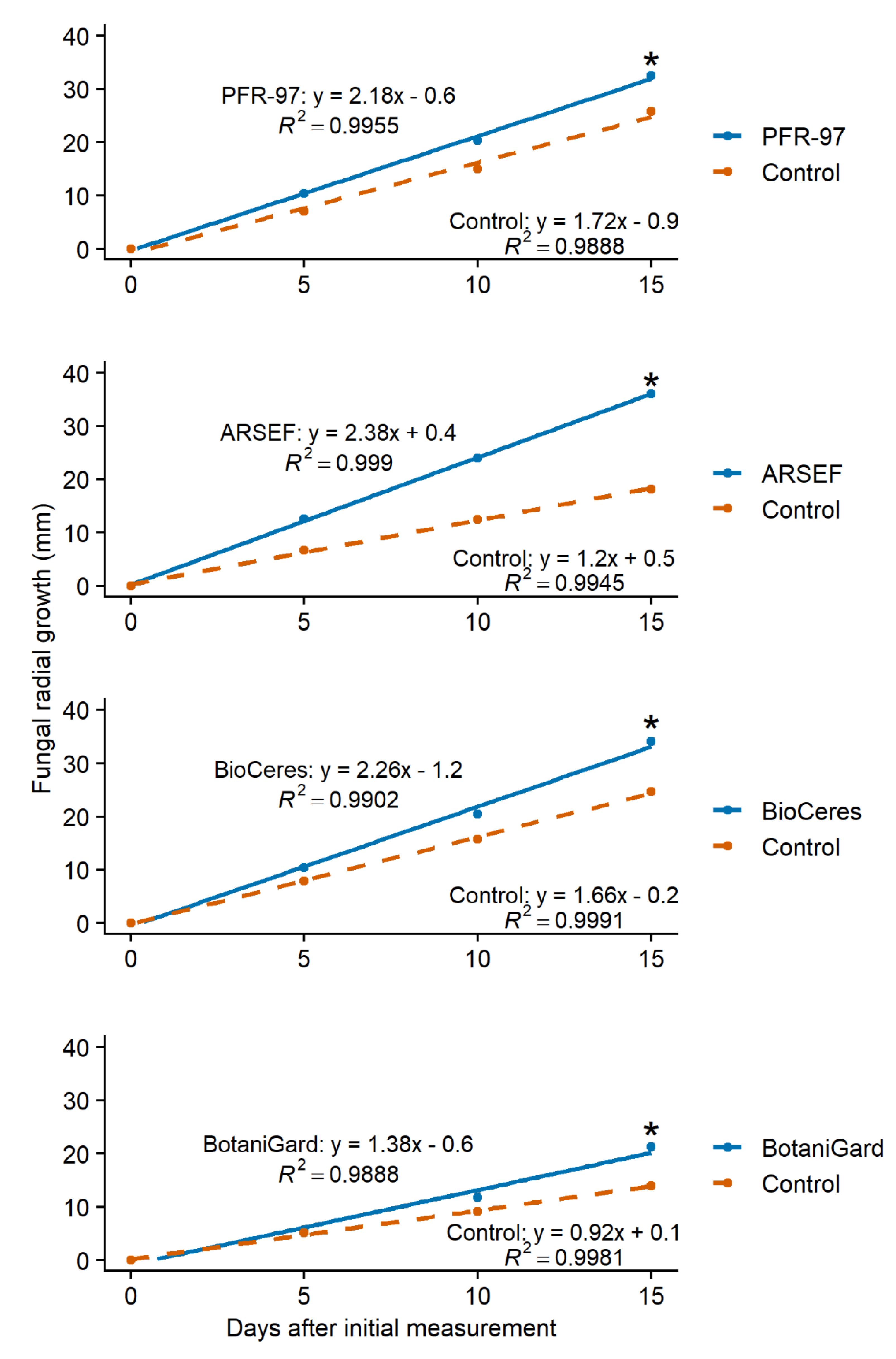
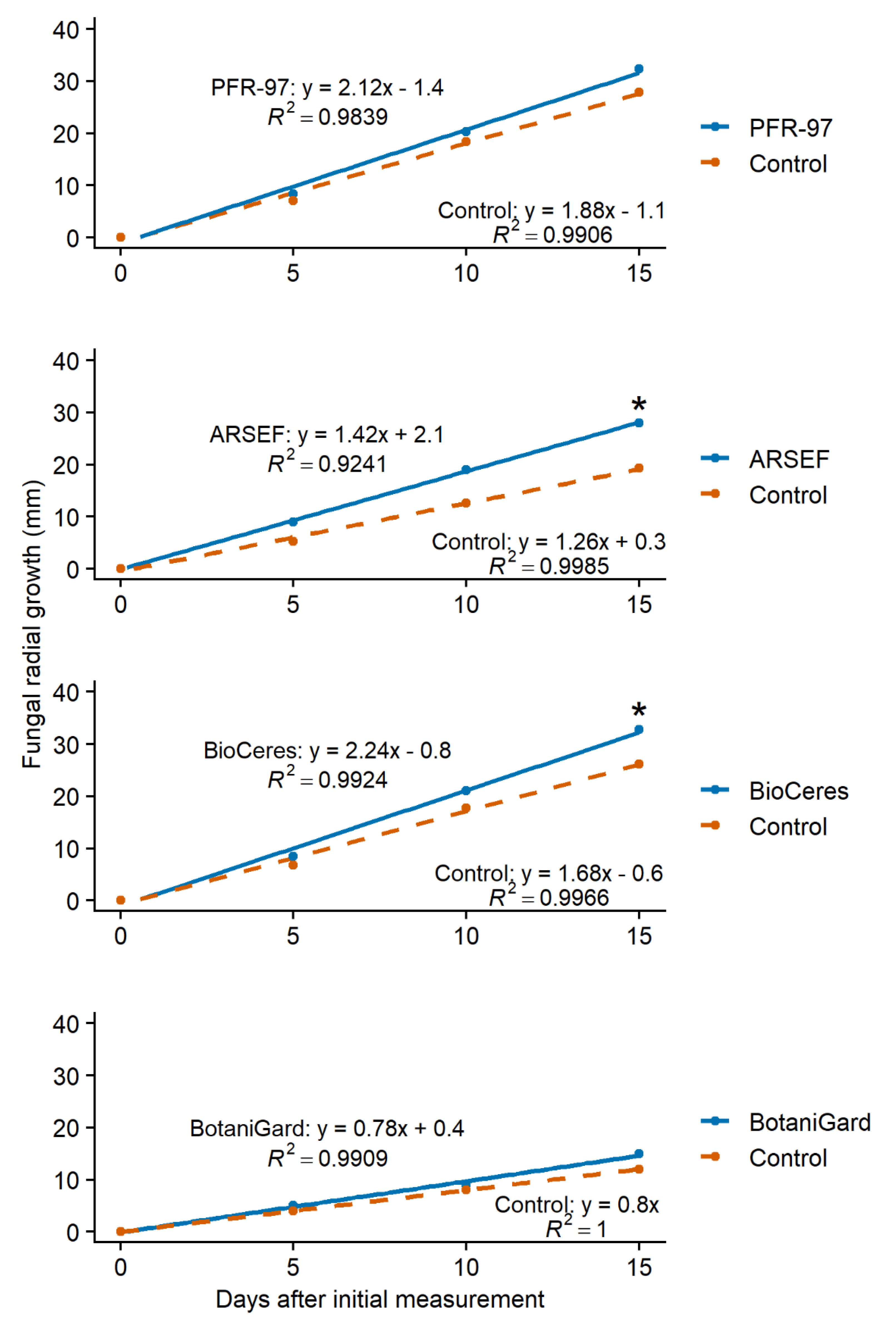
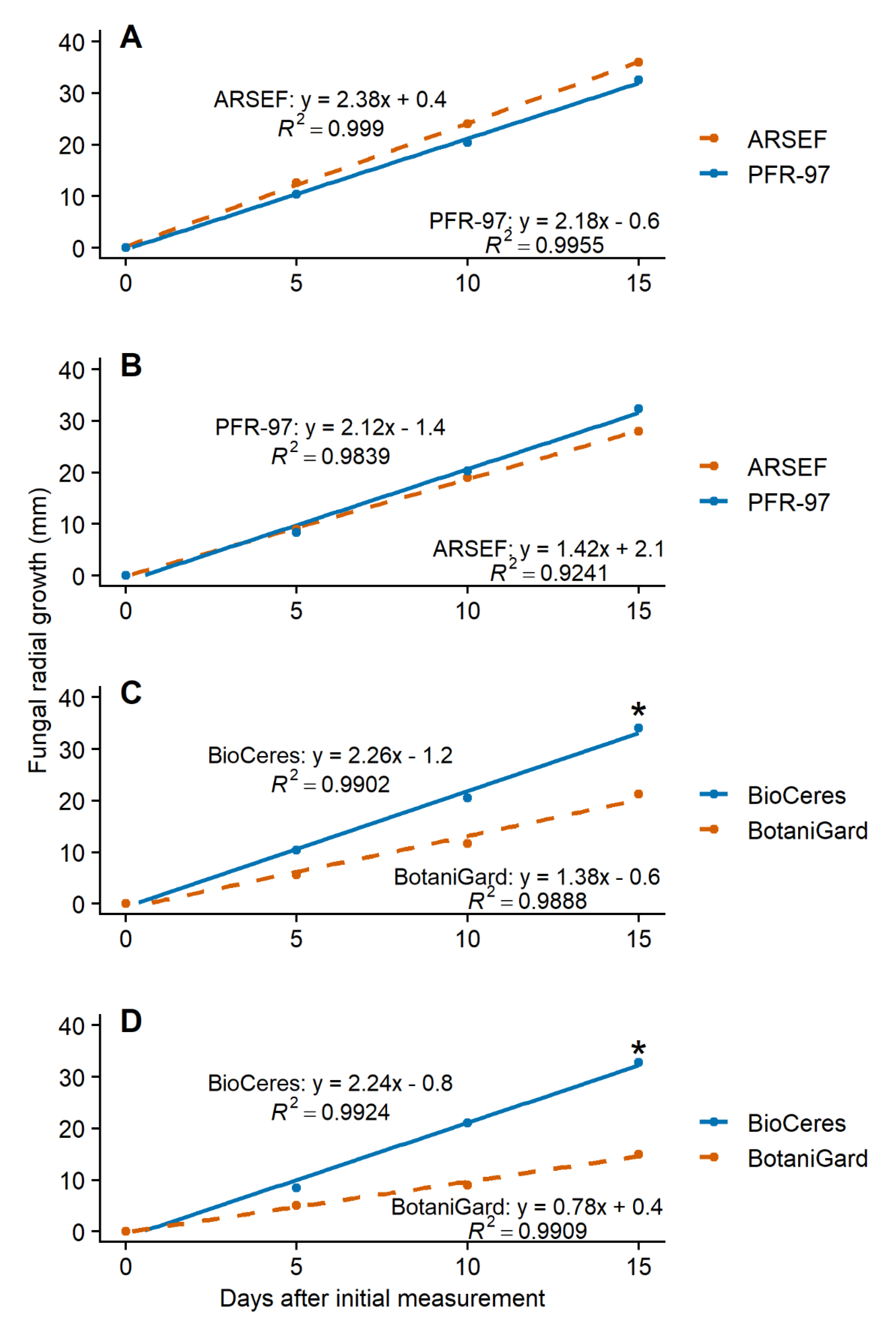
3.1. Effect of Leaf Extracts on Fungal Growth
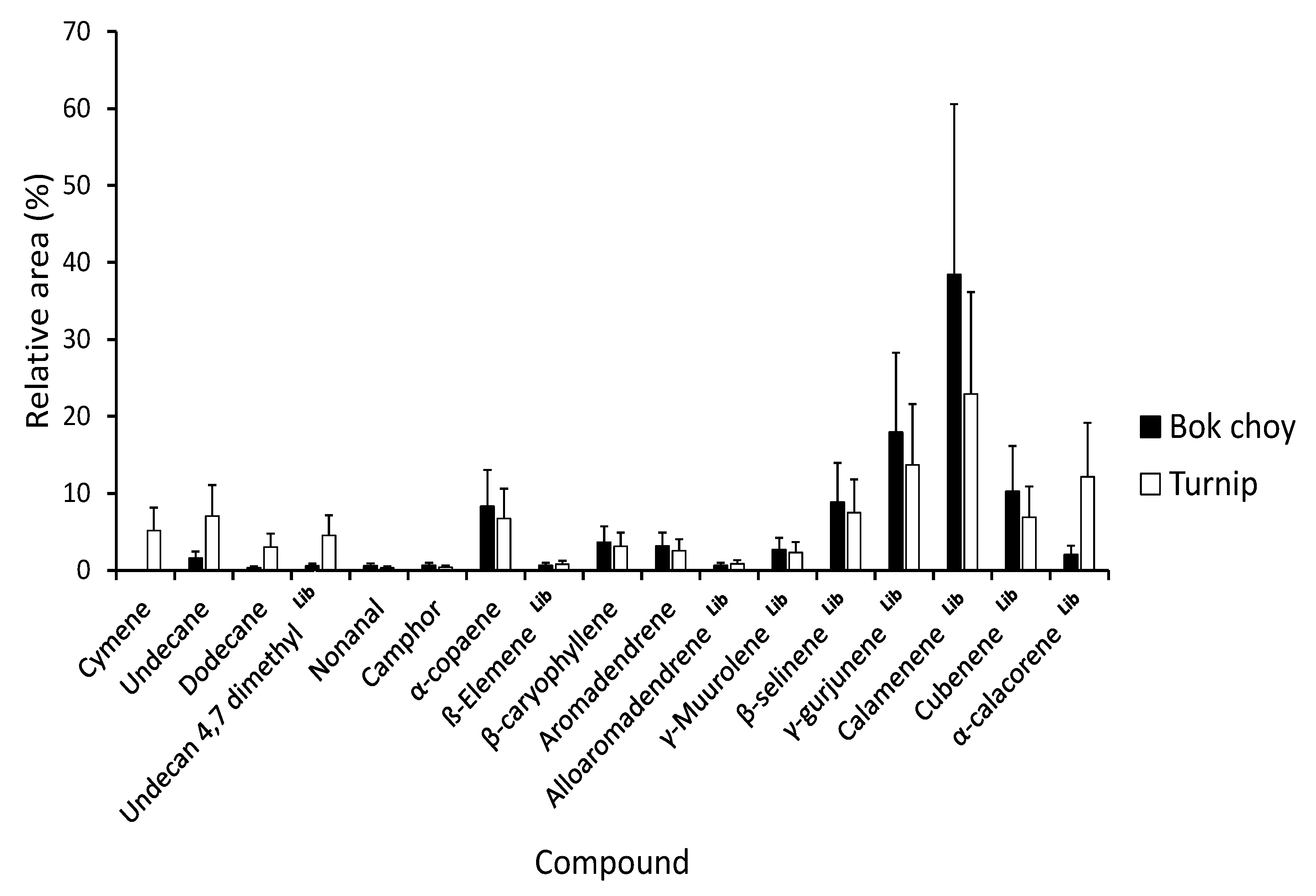
3.2. Gas Chromatography–Mass Spectrophotometry Analysis of Leaf Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The myrosinase-glucosinolate system, its organisation and biochemistry. Physiol. Plant 1996, 97, 194–208. [Google Scholar] [CrossRef]
- Lazzeri, L.; Tacconi, R.; Palmieri, S. In vitro activity of some glucosinolates and their reaction products towards a population of the nematode Heterodera schachtii. J. Agric. Food Chem. 1993, 41, 825–829. [Google Scholar] [CrossRef]
- Yu, Q.; Tsao, R.; Chiba, M.; Potter, J. Selective nematicidal activity of allyl isothiocyanate. JFAE 2005, 3, 218–221. [Google Scholar]
- Manici, L.M.; Lazzeri, L.; Palmieri, S. In vitro fungitoxic activity of some glucosinolates and their enzyme-derived products toward plant pathogenic fungi. J. Agric. Food Chem. 1997, 45, 2768–2773. [Google Scholar] [CrossRef]
- Sotelo, T.; Lema, M.; Soengas, P.; Cartea, M.E.; Velasco, P.; Drake, H.L. In vitro activity of glucosinolates and their degradation products against brassica-pathogenic bacteria and fungi. Appl. Environ. Microbiol. 2014, 81, 432–440. [Google Scholar] [CrossRef] [Green Version]
- Aires, A.; Mota, V.R.; Saavedra, M.J.; Monteiro, A.A.; Simoes, M.; Rosa, E.A.; Bennett, R.N. Initial in vitro evaluations of the antibacterial activities of glucosinolate enzymatic hydrolysis products against plant pathogenic bacteria. J. Appl. Microbiol. 2009, 106, 2096–2105. [Google Scholar] [CrossRef] [PubMed]
- Choesin, D.N.; Boerner, R.E.J. Allyl isothiocyanate release and the allelopathic potential of Brassica napus (Brassicaceae). Am. J. Bot 1991, 78, 1083–1090. [Google Scholar] [CrossRef]
- Applebaum, S.W.; Ichelczik, D.; Humber, R.A. Three in a boat: Host-plant, insect herbivore, and fungal entomopathogen. In Symbioses and Stress. Cellular Origin, Life in Extreme Habitats and Astrobiology; Seckbach, J., Grube, M., Eds.; Springer: Dordrecht, The Netherlands, 2010; Volume 17, pp. 289–326. [Google Scholar]
- Yang, Z.-L.; Kunert, G.; Sporer, T.; Körnig, J.; Beran, F. Glucosinolate abundance and composition in Brassicaceae influence sequestration in a specialist flea beetle. J. Chem. Ecol. 2020, 46, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, I.; Rohloff, J.; Bones, A.M. Defense mechanisms of Brassicaceae: Implications for plant-insect interactions and potential for integrated pest management. A review. Agron. Sustain. Dev. 2010, 30, 311–348. [Google Scholar] [CrossRef] [Green Version]
- Montemayor, C.; Cave, R.D. Prospects for biological control of the yellow-margined leaf beetle, Microtheca ochroloma Stål. Proc. Fla. State Hort Soc. 2009, 122, 250–252. [Google Scholar]
- Balusu, R.R.; Fadamiro, H.Y. Evaluation of organically acceptable insecticides as stand-alone treatments and in rotation for managing yellowmargined leaf beetle, Microtheca ochroloma (Coleoptera: Chrysomelidae), in organic crucifer production. Pest. Manag. Sci. 2011, 68, 573–579. [Google Scholar] [CrossRef]
- Balusu, R.R.; Fadamiro, H.Y. Host finding and acceptance preference of the yellowmargined leaf beetle, Microtheca ochroloma (Coleoptera: Chrysomelidae) on cruciferous crops. Environ. Entomol. 2011, 40, 1471–1477. [Google Scholar] [CrossRef]
- Balusu, R.R.; Fadamiro, H.Y. Susceptibility of Microtheca ochroloma (Coleoptera: Chrysomelidae) to botanical and microbial insecticide formulations. Fla. Entomol. 2013, 96, 914–921. [Google Scholar] [CrossRef]
- Balusu, R.R.; Rhodes, E.M.; Liburd, O.E.; Fadamiro, H.Y. Management of yellowmargined leaf beetle Microtheca ochroloma (Coleoptera: Chrysomelidae) using turnip as trap crop. J. Econ. Entomol. 2015, 108, 2691–2701. [Google Scholar] [CrossRef]
- Gámez Herrera, C.; Niño, A.A.; Avery, P.B.; Cave, R.D. Effect of the fungus Isaria fumosorosea Wize on herbivory by adult yellowmargined leaf beetles, Microtheca ochroloma Stål (Coleoptera: Chrysomelidae). Ceiba 2016, 54, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Montemayor, C.; Avery, P.B.; Cave, R.D. Infection and mortality of Microtheca ochroloma (Coleoptera: Chrysomelidae) by Isaria fumosorosea (Hypocreales: Cordycipitaceae) under laboratory conditions. Biocontrol Sci. Technol. 2016, 26, 605–616. [Google Scholar] [CrossRef]
- Senthil-Nathan, S. A review of biopesticides and their mode of action against insect pests. In Environmental Sustainability; Thangavel, P., Sridevi, G., Eds.; Springer: New Dehli, India, 2015; pp. 49–63. [Google Scholar]
- Brownbridge, M.; Buitenhuis, R. Integration of microbial biopesticides in greenhouse floriculture: The Canadian experience. J. Invertebr. Pathol. 2019, 165, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Arthurs, S.; Dara, S.K. Microbial biopesticides for invertebrate pests and their markets in the United States. J. Invertebr. Pathol. 2019, 165, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Mascarin, G.M.; Lopes, R.B.; Delalibera, I., Jr.; Fernandes, E.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Hatting, J.L.; Moore, S.D.; Mala, A.P. Microbial control of phytophagous invertebrate pests in South Africa: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 54–66. [Google Scholar] [CrossRef]
- Karimi, J.; Dara, S.K.; Arthurs, S. Microbial insecticides in Iran: History, current status, challenges and perspective. J. Invertebr. Pathol. 2019, 165, 67–73. [Google Scholar] [CrossRef]
- Kumar, K.K.; Sridhar, J.; Murali-Baskaran, R.K.; Senthil-Nathan, S.; Kaushal, P.; Dara, S.K.; Arthurs, S. Microbial pesticides for insect pest management in India: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 74–81. [Google Scholar] [CrossRef]
- Glare, T.R.; O’Callaghan, M. Microbial biopesticides for control of invertebrates: Progress from New Zealand. J. Invertebr. Pathol. 2019, 165, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Feng, P.; Wang, C. Fungi that infects insects: Altering host behavior and beyond. PLoS Pathog. 2015, 11, e1005037. [Google Scholar] [CrossRef] [Green Version]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: Insights into host-pathogen interactions. Adv. Genet. 2016, 94, 1–39. [Google Scholar]
- Lei, Y.; Hussain, A.; Guan, Z.; Wang, D.; Jaleel, W.; Lyu, L.; He, Y. Unraveling the mode of action of Cordyceps fumosorosea: Potential biocontrol agent against Plutella xylostella (Lepidoptera: Plutellidae). Insects 2021, 12, 179. [Google Scholar] [CrossRef]
- Poprawski, T.J.; Jones, W.J. Host plant effects on activity of the mitosporic fungi Beauveria bassiana and Paecilomyces fumosoroseus against two populations of Bemisia whiteflies (Homoptera: Aleyrodidae). Mycopathologia 2001, 151, 11–20. [Google Scholar] [CrossRef]
- Kepler, R.M.; Luangsa-Ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Diao, H.; Arthurs, S.; Hao, C.; Mascarin, G.M.; Ma, R. Host plants influence susceptibility of whitefly Bemisia tabaci (Hemiptera: Aleyrodidae) to the entomopathogenic fungus Isaria fumosorosea (Hypocreales: Cordycipitaceae). Biocontrol Sci. Technol. 2016, 26, 528–538. [Google Scholar] [CrossRef]
- Poprawski, T.J.; Greenberg, S.M.; Ciomperlik, M.A. Effect of host plant on Beauveria bassiana- and Paecilomyces fumosoroseus-induced mortality of Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Environ. Entomol. 2000, 29, 1048–1053. [Google Scholar] [CrossRef]
- Vega, F.E.; Dowd, P.F.; Mcguire, M.R.; Jackson, M.A.; Nelsen, T.C. In vitro effects of secondary plant compounds on germination of blastospores of the entomopathogenic fungus Paecilomyces fumosoroseus (Deuteromycotina: Hyphomycetes). J. Invertebr. Pathol. 1997, 70, 209–213. [Google Scholar] [CrossRef]
- Inyang, E.N.; Butt, T.M.; Doughty, K.J.; Todd, A.D.; Archer, S. The effects of isothiocyanates on the growth of the entomo-pathogenic fungus Metarhizium anisopliae and its infection of the mustard beetle. Mycol. Res. 1999, 103, 974–980. [Google Scholar] [CrossRef]
- Klingen, I.; Hajek, A.; Meadow, R.; Renwick, J.A.A. Effect of brassicaceous plants on the survival and infectivity of insect pathogenic fungi. BioControl 2002, 47, 411–425. [Google Scholar] [CrossRef]
- Ujjan, A.A.; Shahzad, S. Use of entomopathogenic fungi for the control of mustard aphid (Lipaphis erysimi) on canola (Brassica napus L.). Pak. J. Bot 2012, 44, 2081–2086. [Google Scholar]
- Dos Anjos, J.; Rosalino, P.; Sausen, C.D.; Do Prado, L.; Egewarth, R.; Soares, V.; Bastos, S.T. Fungos entomopatogênicos em Diabrotica speciosa e Microtheca ochroloma (Col., Chrysomelidae) em hortalizas [Entomopathogenic fungi in Diabrotica speciosa and Microtheca ochroloma (Col.Chrysomelidae) in leafy greens]’. In Informe Técnico; Universidade Federal de Santa Maria: Santa Maria, Brazil, 2007. [Google Scholar]
- Naidu, G.P. Antifungal activity in Codiaeum variegatum leaf extract. Curr. Sci. 1988, 57, 502–504. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 12 December 2020).
- Lin, Y.; Hussain, M.; Avery, P.B.; Qasim, M.; Fang, D.; Wang, L. Volatiles from plants induced by multiple aphid attacks promote conidial performance of Lecanicillium lecanii. PLoS ONE 2016, 11, e0151844. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Qasim, M.; Hussain, M.; Akutse, K.S.; Avery, P.B.; Dash, C.K.; Wang, L. The herbivore-induced plant volatiles methyl salicylate and menthol positively affect growth and pathogenicity of entomopathogenic fungi. Sci. Rep. 2017, 7, 40494. [Google Scholar] [CrossRef]
- Elliot, S.L.; Sabelis, M.W.; Janssen, A.; van der Geest, L.P.S.; Beerling, E.A.M.; Fransen, J. Can plants use entomopathogens as body guards? Ecol. Lett. 2000, 3, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Cory, J.S.; Hoover, K. Plant-mediated effects in insect-pathogen interactions. Trends Ecol. Evol. 2006, 21, 278–286. [Google Scholar] [CrossRef]
- Cory, J.S.; Ericsson, J.D. Fungal entomopathogens in a tritrophic context. BioControl 2010, 55, 75–88. [Google Scholar] [CrossRef]
- Meekes, E.T.M.; van Voorst, S.; Joosten, N.N.; Fransen, J.J.; van Lenteren, J.C. Persistence of the fungal whitefly pathogen, Aschersonia aleyrodis, on three different plant species. Mycol. Res. 2000, 104, 1234–1240. [Google Scholar] [CrossRef]
- Avery, P.B.; Queeley, G.L.; Faull, J.; Simmonds, M.S.J. Effect of photoperiod and host distribution on the horizontal transmission of Isaria fumosorosea (Hypocreales: Cordycipitaceae) in greenhouse whitefly assessed using a novel model bioassay. Biocontrol Sci. Technol. 2010, 20, 1097–1111. [Google Scholar] [CrossRef] [Green Version]
- Altre, J.A.; Vandenberg, J.D. Penetration of cuticle and proliferation in hemolymph by Paecilomyces fumosoroseus isolates that differ in virulence against lepidopteran larvae. J. Invertebr. Pathol. 2001, 78, 81–86. [Google Scholar] [CrossRef]
- Costa, S.D.; Gaugler, R. Influence of Solanum host plants on Colorado potato beetle (Coleoptera: Chrysomelidae) susceptibility to the entomopathogen Beauveria bassiana. Environ. Entomol. 1989, 18, 531–536. [Google Scholar] [CrossRef]
- Costa, S.D.; Gaugler, R. Sensitivity of Beauveria bassiana to solanine and tomatine: Plant defensive chemicals inhibit an insect pathogen. J. Chem. Ecol. 1989, 15, 697–706. [Google Scholar] [CrossRef]
- Gasmi, L.; Martínez-Solís, M.; Frattini, A.; Ye, M.; Collado, M.C.; Turlings, T.C.J.; Erb, M.; Herrero, S. Can herbivore-induced volatiles protect plants by increasing the herbivores’ susceptibility to natural pathogens? Appl. Environ. Microbiol. 2019, 85, e01468-18. [Google Scholar] [CrossRef] [Green Version]
- Ocampo-Hernández, J.A.; Tamayo-Mejía, F.; Tamez-Guerra, P.; Gao, Y.; Ariel, W.; Guzmán-Franco, A.W. Different host plant species modifies the susceptibility of Bactericera cockerelli to the entomopathogenic fungus Beauveria bassiana. J. Appl. Entomol. 2019, 143, 984–991. [Google Scholar] [CrossRef]
- Robertson, G.W.; Griffiths, D.W.; Smith, M.W.; Butcher, R.D. The application of thermal desorption-gas chromatography-mass spectrometry to the analyses of flower volatiles from five varieties of oilseed rape (Brassica napus spp. oleifera). Phytochem. Anal. 1993, 4, 152–157. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Haubruge, E.; De Moraes, C.M.; Mescher, M.C. Aphid responses to volatile cues from turnip plants (Brassica rapa) infested with phloem-feeding and chewing herbivores. Arthropod-Plant. Interact. 2013, 7, 567–577. [Google Scholar] [CrossRef]
- Akiyama, K.; Matsuzaki, K.I.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar] [CrossRef] [PubMed]
- Berrocal, A.; Oviedo, C.; Nickerson, K.W.; Navarrete, J. Quorum sensing activity and control of yeast-mycelium dimorphism in Ophiostoma floccosum. Biotechnol. Lett. 2014, 36, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Kramer, R.; Abraham, W.R. Volatile sesquiterpenes from fungi: What are they good for? Phytochem. Rev. 2012, 11, 15–37. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Radial Growth a | |||
|---|---|---|---|
| Fungal product | Species (strain) | Bok choy b | Turnip c |
| PFR-97 | Cordyceps fumosorosea (Apopka) | 33 ± 1.2 b | 32 ± 1.0 c |
| ARSEF 3581 | Cordyceps fumosorosea (3581-TX) | 36 ± 0.9 b | 28 ± 0.6 b |
| BioCeres WP | Beauveria bassiana (ANT-03) | 34 ± 0.6 b | 33 ± 0.7 c |
| BotaniGard ES | Beauveria bassiana (GHA) | 21 ± 0.7 a | 15 ± 0.7 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerritos-Garcia, D.G.; Avery, P.B.; Martini, X.; Candian, V.; Cano, L.M.; Cave, R.D. In Vitro Effects of Leaf Extracts from Brassica rapa on the Growth of Two Entomopathogenic Fungi. J. Fungi 2021, 7, 779. https://doi.org/10.3390/jof7090779
Cerritos-Garcia DG, Avery PB, Martini X, Candian V, Cano LM, Cave RD. In Vitro Effects of Leaf Extracts from Brassica rapa on the Growth of Two Entomopathogenic Fungi. Journal of Fungi. 2021; 7(9):779. https://doi.org/10.3390/jof7090779
Chicago/Turabian StyleCerritos-Garcia, Daniel G., Pasco B. Avery, Xavier Martini, Valentina Candian, Liliana M. Cano, and Ronald D. Cave. 2021. "In Vitro Effects of Leaf Extracts from Brassica rapa on the Growth of Two Entomopathogenic Fungi" Journal of Fungi 7, no. 9: 779. https://doi.org/10.3390/jof7090779