
A New Duplex PCR-Assay for the Detection and Identification of Paracoccidioides Species

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Fungal Strains and Culture Conditions
2.3. DNA Extraction
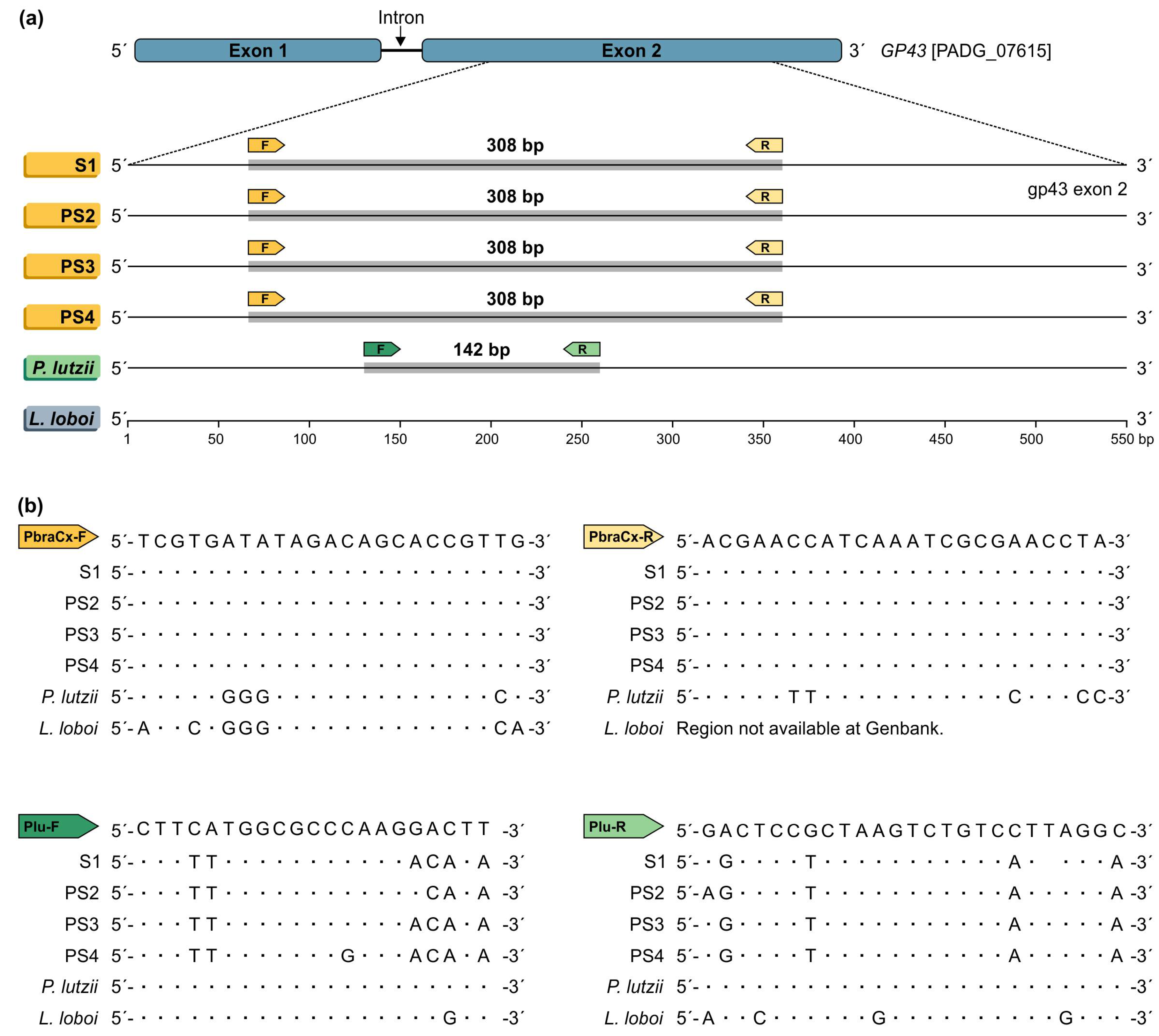
2.4. Primer Design
2.5. In Silico PCR
2.6. PCR Optimization and Gel Electrophoresis
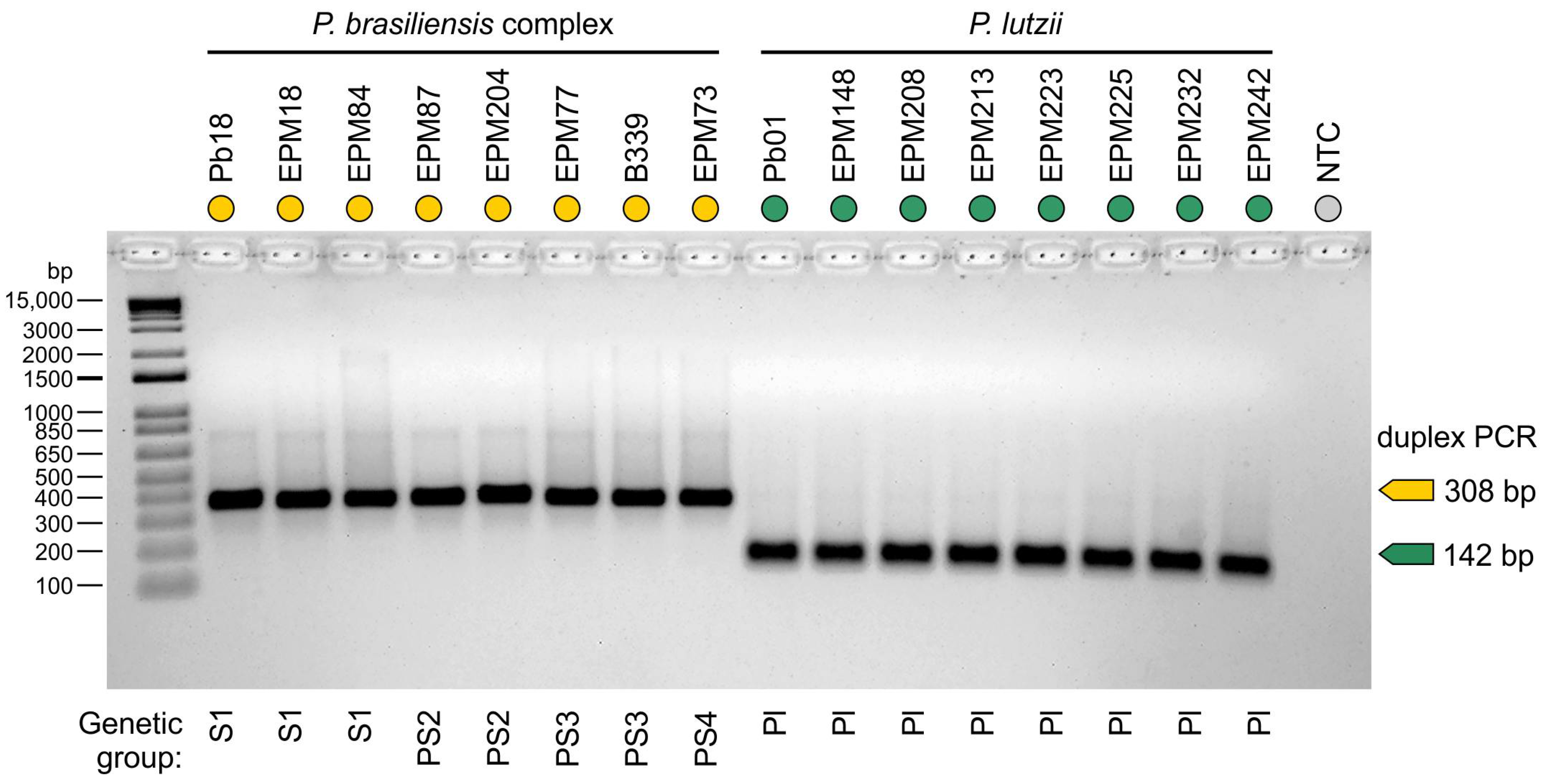
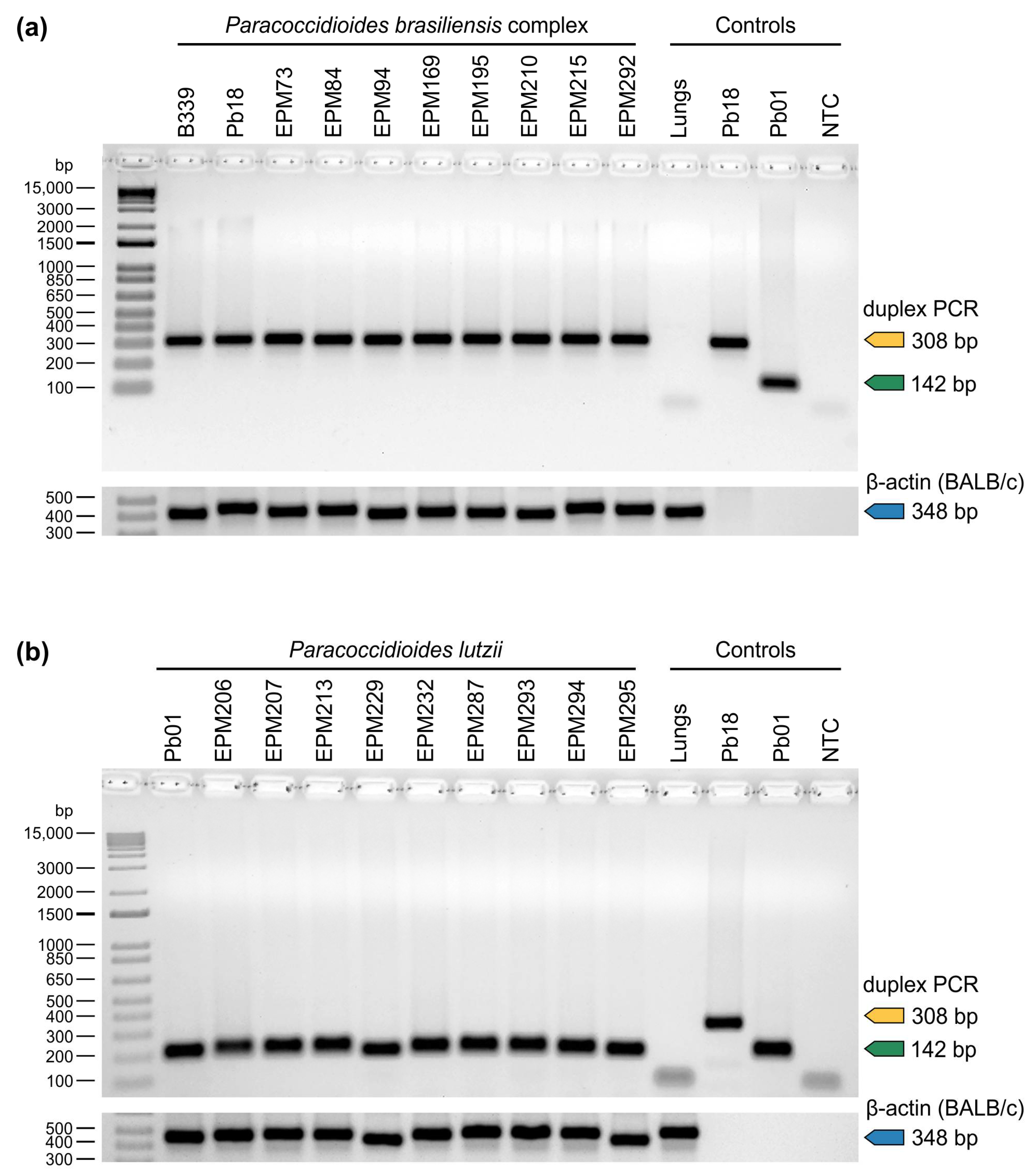
2.7. Assay Specificity
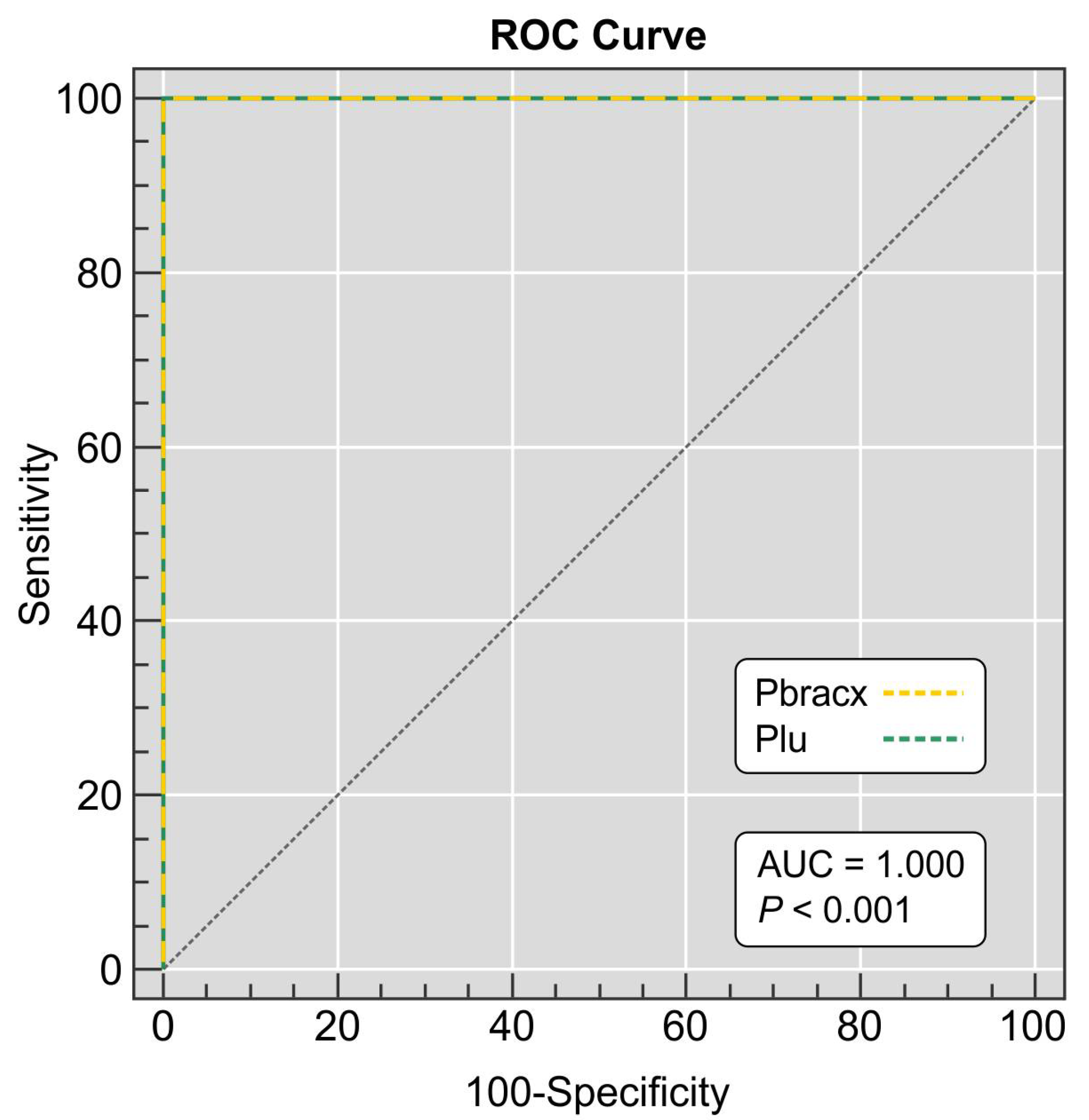
2.8. Assay Sensitivity
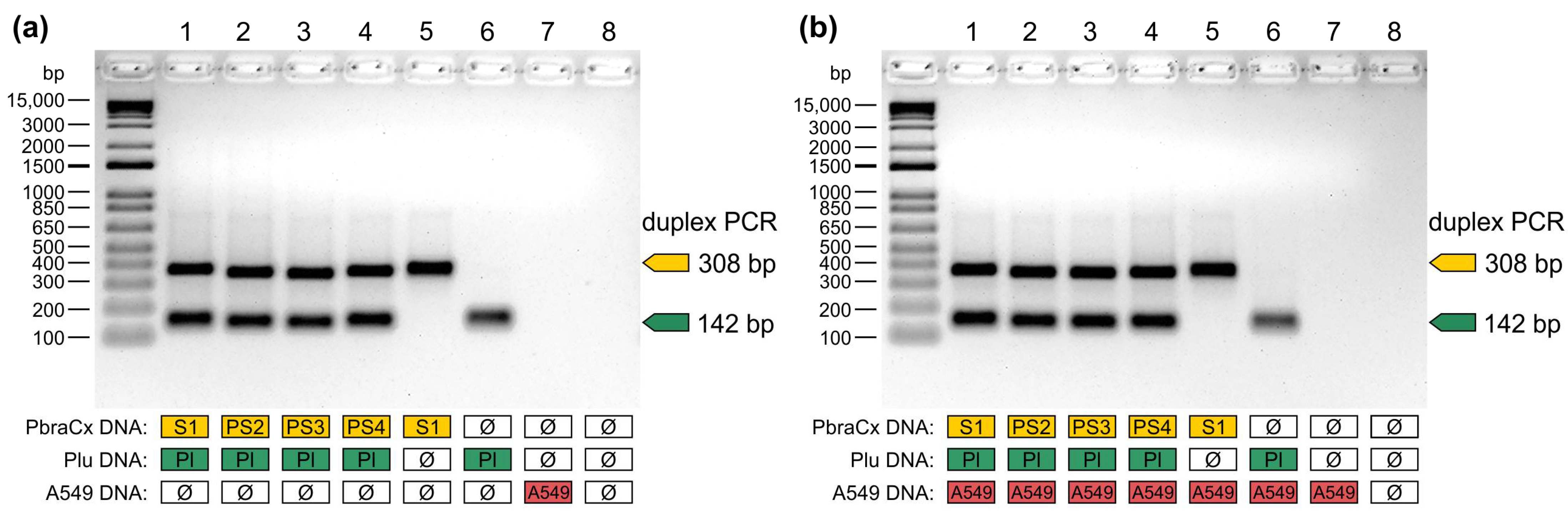
2.9. Non-Target Template Competition
2.10. Detection of Paracoccidioides DNA from BALB/c Lungs and Soil
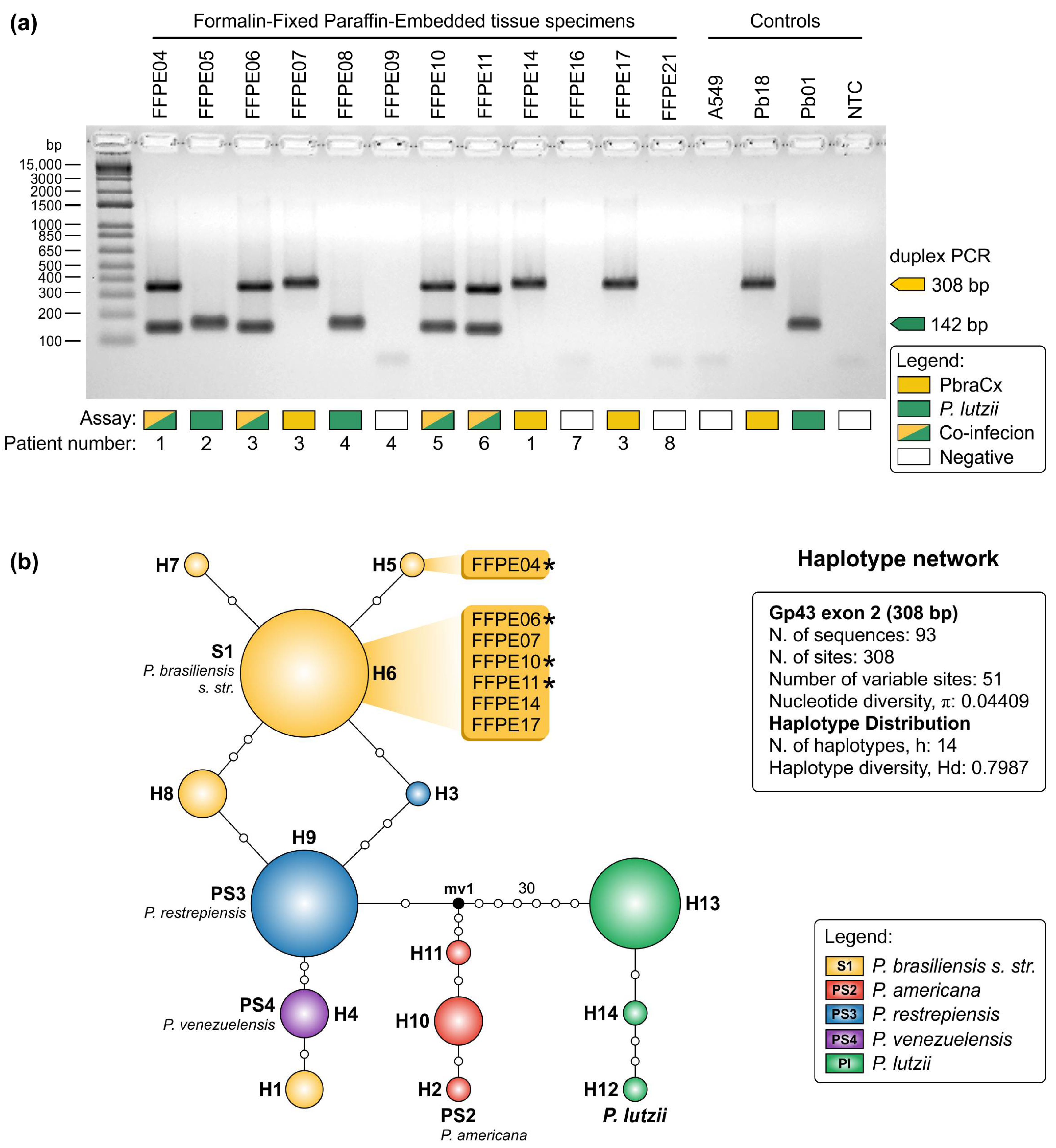
2.11. Detection of Paracoccidioides DNA from Formalin-Fixed, Paraffin-Embedded (FFPE) Tissue Sections
2.12. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- San-Blas, G.; Nino-Vega, G.; Iturriaga, T. Paracoccidioides brasiliensis and paracoccidioidomycosis: Molecular approaches to morphogenesis, diagnosis, epidemiology, taxonomy and genetics. Med. Mycol. 2002, 40, 225–242. [Google Scholar] [CrossRef] [Green Version]
- Lutz, A. Uma micose pseudococídica localizada na boca e observada no Brasil: Contribuição ao conhecimento das hifoblastomicoses americanas. Bras. Med. 1908, 22, 121–124. [Google Scholar]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; Valle, A.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef]
- Fortes, M.R.; Miot, H.A.; Kurokawa, C.S.; Marques, M.E.; Marques, S.A. Immunology of paracoccidioidomycosis. Ann. Bras. Dermatol. 2011, 86, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Hahn, R.C.; Rodrigues, A.M.; Della Terra, P.P.; Nery, A.F.; Hoffmann-Santos, H.D.; Góis, H.M.; Fontes, C.J.; de Camargo, Z.P. Clinical and epidemiological features of paracoccidioidomycosis due to Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2019, 13, e0007437. [Google Scholar] [CrossRef] [Green Version]
- do Amaral, C.C.; Fernandes, G.F.; Rodrigues, A.M.; Burger, E.; de Camargo, Z.P. Proteomic analysis of Paracoccidioides brasiliensis complex isolates: Correlation of the levels of differentially expressed proteins with in vivo virulence. PLoS ONE 2019, 14, e0218013. [Google Scholar] [CrossRef]
- Brummer, E.; Castaneda, E.; Restrepo, A. Paracoccidioidomycosis: An update. Clin. Microbiol. Rev. 1993, 6, 89–117. [Google Scholar] [CrossRef]
- Blotta, M.H.; Mamoni, R.L.; Oliveira, S.J.; Nouer, S.A.; Papaiordanou, P.M.; Goveia, A.; Camargo, Z.P. Endemic regions of paracoccidioidomycosis in Brazil: A clinical and epidemiologic study of 584 cases in the southeast region. Am. J. Trop. Med. Hyg. 1999, 61, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.D. Estudos comparativos do granuloma coccidióidico nos Estados Unidos e no Brasil. Novo gênero para o parasito brasileiro. An. Fac. Med. Sao Paulo 1930, 5, 125–141. [Google Scholar]
- Nino-Vega, G.A.; Calcagno, A.M.; San-Blas, G.; San-Blas, F.; Gooday, G.W.; Gow, N.A. RFLP analysis reveals marked geographical isolation between strains of Paracoccidioides brasiliensis. Med. Mycol. 2000, 38, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Totti, D.O.; Romanha, A.J.; Grisard, E.C.; Simpson, A.J.; Koury, M.C. Random amplified polymorphic DNA (RAPD) analysis of Paracoccidioides brasiliensis isolates. Rev. Latinoam. Microbiol. 1999, 41, 139–143. [Google Scholar] [PubMed]
- Hahn, R.C.; Macedo, A.M.; Santos, N.L.; Resende, J.C.; Hamdan, J.S. Characterization of Paracoccidioides brasiliensis atypical isolates by random amplified polymorphic DNA analysis. Rev. Iberoam. Micol. 2002, 19, 49–51. [Google Scholar]
- Matute, D.R.; Sepulveda, V.E.; Quesada, L.M.; Goldman, G.H.; Taylor, J.W.; Restrepo, A.; McEwen, J.G. Microsatellite analysis of three phylogenetic species of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2006, 44, 2153–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrero, L.L.; Nino-Vega, G.; Teixeira, M.M.; Carvalho, M.J.; Soares, C.M.; Pereira, M.; Jesuino, R.S.; McEwen, J.G.; Mendoza, L.; Taylor, J.W.; et al. New Paracoccidioides brasiliensis isolate reveals unexpected genomic variability in this human pathogen. Fungal Genet. Biol. 2008, 45, 605–612. [Google Scholar] [CrossRef]
- Salgado-Salazar, C.; Jones, L.R.; Restrepo, Á.; McEwen, J.G. The human fungal pathogen Paracoccidioides brasiliensis (Onygenales: Ajellomycetaceae) is a complex of two species: Phylogenetic evidence from five mitochondrial markers. Cladistics 2010, 26, 613–624. [Google Scholar] [CrossRef]
- Feitosa Ldos, S.; Cisalpino, P.S.; dos Santos, M.R.; Mortara, R.A.; Barros, T.F.; Morais, F.V.; Puccia, R.; da Silveira, J.F.; de Camargo, Z.P. Chromosomal polymorphism, syntenic relationships, and ploidy in the pathogenic fungus Paracoccidioides brasiliensis. Fungal Genet. Biol. 2003, 39, 60–69. [Google Scholar] [CrossRef]
- Matute, D.R.; McEwen, J.G.; Puccia, R.; Montes, B.A.; San-Blas, G.; Bagagli, E.; Rauscher, J.T.; Restrepo, A.; Morais, F.; Niño-Vega, G.; et al. Cryptic speciation and recombination in the fungus Paracoccidioides brasiliensis as revealed by gene genealogies. Mol. Biol. Evol. 2006, 23, 65–73. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; Nino-Vega, G.; Bagagli, E.; Felipe, M.S.S. Paracoccidioides species complex: Ecology, phylogeny, sexual reproduction, and virulence. PLoS Pathog. 2014, 10, e1004397. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.M.; Theodoro, R.C.; de Carvalho, M.J.A.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenet Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Cocio, T.A.; Nascimento, E.; von Zeska Kress, M.R.; Bagagli, E.; Martinez, R. Phylogenetic species of Paracoccidioides spp. isolated from clinical and environmental samples in a hyperendemic area of paracoccidioidomycosis in Southeastern Brazil. J. Fungi 2020, 6, 132. [Google Scholar] [CrossRef]
- Cocio, T.A.; Nascimento, E.; Kress, M.R.; Bagagli, E.; Martinez, R. Characterization of a Paracoccidioides spp. strain from southeastern Brazil genotyped as Paracoccidioides restrepiensis (PS3) and review of this phylogenetic species. Genet. Mol. Biol. 2020, 43, e20190201. [Google Scholar] [CrossRef]
- Teixeira, M.d.M.; Cattana, M.E.; Matute, D.R.; Muñoz, J.F.; Arechavala, A.; Isbell, K.; Schipper, R.; Santiso, G.; Tracogna, F.; Sosa, M.d.l.Á.; et al. Genomic diversity of the human pathogen Paracoccidioides across the South American continent. Fungal Genet. Biol. 2020, 140, 103395. [Google Scholar] [CrossRef]
- Theodoro, R.C.; Teixeira, M.d.M.; Felipe, M.S.S.; Paduan, K.d.S.; Ribolla, P.M.; San-Blas, G.; Bagagli, E. Genus Paracoccidioides: Species recognition and biogeographic aspects. PLoS ONE 2012, 7, e37694. [Google Scholar] [CrossRef] [Green Version]
- Tatagiba, L.S.; Pivatto, L.B.; Faccini-Martínez, Á.A.; Peçanha, P.M.; Grão Velloso, T.R.; Gonçalves, S.S.; Rodrigues, A.M.; Camargo, Z.P.; Falqueto, A. A case of paracoccidioidomycosis due to Paracoccidioides lutzii presenting sarcoid-like form. Med. Mycol. Case Rep. 2018, 19, 6–8. [Google Scholar] [CrossRef]
- Marques-da-Silva, S.H.; Rodrigues, A.M.; de Hoog, G.S.; Silveira-Gomes, F.; de Camargo, Z.P. Occurrence of Paracoccidioides lutzii in the Amazon region: Description of two cases. Am. J. Trop. Med. Hyg. 2012, 87, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Martinez, R. New trends in paracoccidioidomycosis epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Macedo, P.M.; Teixeira, M.D.M.; Barker, B.M.; Zancopé-Oliveira, R.M.; Almeida-Paes, R.; do Valle, A.C.F. Clinical features and genetic background of the sympatric species Paracoccidioides brasiliensis and Paracoccidioides americana. PLoS Negl. Trop. Dis. 2019, 13, e0007309. [Google Scholar] [CrossRef]
- Pereira, E.F.; Gegembauer, G.; Chang, M.R.; Camargo, Z.P.D.; Nunes, T.F.; Ribeiro, S.M.; Carvalho, L.R.D.; Maldonado, B.M.; Mendes, R.P.; Paniago, A.M.M. Comparison of clinico-epidemiological and radiological features in paracoccidioidomycosis patients regarding serological classification using antigens from Paracoccidioides brasiliensis complex and Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2020, 14, e0008485. [Google Scholar] [CrossRef] [PubMed]
- Camargo, Z.P.; Rodrigues, A.M. Paracoccidioides Complex. In Pocket Guide to Mycological Diagnosis; Cordeiro, R.D.A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; Volume 78, pp. 125–134. [Google Scholar]
- Teixeira, M.M.; Theodoro, R.C.; Oliveira, F.F.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Roberto, T.N.; Rodrigues, A.M.; Hahn, R.C.; de Camargo, Z.P. Identifying Paracoccidioides phylogenetic species by PCR-RFLP of the alpha-tubulin gene. Med. Mycol. 2016, 54, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Gomes, G.M.; Cisalpino, P.S.; Taborda, C.P.; de Camargo, Z.P. PCR for diagnosis of paracoccidioidomycosis. J. Clin. Microbiol. 2000, 38, 3478–3480. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, B.G.; Hahn, R.C.; Camargo, Z.P.; Rodrigues, A.M. Molecular Tools for Detection and Identification of Paracoccidioides Species: Current Status and Future Perspectives. J. Fungi 2020, 6, 293. [Google Scholar] [CrossRef] [PubMed]
- Fava-Netto, C. Contribuição para o estudo imunológico da blastomicose de Lutz (blastomicose sul-americana). Rev. Inst. Adolfo Lutz 1961, 21, 99–194. [Google Scholar]
- Fava-Netto, C.; Vegas, V.S.; Sciannamea, I.M.; Guarnieri, D.B. The polysaccharidic antigen from Paracoccidioides brasiliensis. Study of the time of cultivation necessary for the preparation of the antigen. Rev. Inst. Med. Trop. Sao Paulo 1969, 11, 177–181. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hahn, R.C.; Rodrigues, A.M.; Fontes, C.J.; Nery, A.F.; Tadano, T.; de Padua Queiroz Junior, L.; de Camargo, Z.P. Fatal fungemia due to Paracoccidioides lutzii. Am. J. Trop. Med. Hyg. 2014, 91, 394–398. [Google Scholar] [CrossRef] [Green Version]
- Vilela, R.; Mendoza, L.; Rosa, P.S.; Belone, A.F.F.; Madeira, S.; Opromolla, D.V.A.; de Resende, M.A. Molecular model for studying the uncultivated fungal pathogen Lacazia loboi. J. Clin. Microbiol. 2005, 43, 3657–3661. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, A.M.; de Hoog, G.S.; de Camargo, Z.P. Molecular diagnosis of pathogenic Sporothrix species. PLoS Negl. Trop. Dis. 2015, 9, e0004190. [Google Scholar] [CrossRef] [Green Version]
- Giard, D.J.; Aaronson, S.A.; Todaro, G.J.; Arnstein, P.; Kersey, J.H.; Dosik, H.; Parks, W.P. In vitro cultivation of human tumors: Establishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Inst. 1973, 51, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- van Beers, E.H.; Joosse, S.A.; Ligtenberg, M.J.; Fles, R.; Hogervorst, F.B.L.; Verhoef, S.; Nederlof, P.M. A multiplex PCR predictor for aCGH success of FFPE samples. Br. J. Cancer 2006, 94, 333–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, A.M.; Najafzadeh, M.J.; de Hoog, G.S.; de Camargo, Z.P. Rapid identification of emerging human-pathogenic Sporothrix species with rolling circle amplification. Front. Microbiol. 2015, 6, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pahl, A.; Kuhlbrandt, U.; Brune, K.; Rollinghoff, M.; Gessner, A. Quantitative detection of Borrelia burgdorferi by real-time PCR. J. Clin. Microbiol. 1999, 37, 1958–1963. [Google Scholar] [CrossRef] [Green Version]
- Bialek, R.; Feucht, A.; Aepinus, C.; Just-Nübling, G.; Robertson, V.J.; Knobloch, J.; Hohle, R. Evaluation of two nested PCR assays for detection of Histoplasma capsulatum DNA in human tissue. J. Clin. Microbiol. 2002, 40, 1644–1647. [Google Scholar] [CrossRef] [Green Version]
- Ricci, G.; Zelck, U.; Mota, F.; Lass-Florl, C.; Franco, M.F.; Bialek, R. Genotyping of Paracoccidioides brasiliensis directly from paraffin embedded tissue. Med. Mycol. 2008, 46, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; Ahn, S.; Hong, M.; Bang, H.; Van Vrancken, M.; Kim, S.; Lee, J.; Park, S.H.; Park, J.O.; Park, Y.S.; et al. Tissue recommendations for precision cancer therapy using next generation sequencing: A comprehensive single cancer center’s experiences. Oncotarget 2017, 8, 42478–42486. [Google Scholar] [CrossRef] [Green Version]
- Blow, N. Tissue preparation: Tissue issues. Nature 2007, 448, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Rickerts, V. Identification of fungal pathogens in Formalin-fixed, Paraffin-embedded tissue samples by molecular methods. Fungal Biol. 2016, 120, 279–287. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, msx248. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; de Hoog, G.S.; Zhang, Y.; Camargo, Z.P. Emerging sporotrichosis is driven by clonal and recombinant Sporothrix species. Emerg. Microbes Infect. 2014, 3, e32. [Google Scholar] [CrossRef] [Green Version]
- Altman, D.G. Practical Statistics for Medical Research; Chapman and Hall: London, UK, 1991; p. 624. [Google Scholar]
- Shikanai-Yasuda, M.A.; Telles Filho Fde, Q.; Mendes, R.P.; Colombo, A.L.; Moretti, M.L. Guidelines in paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2006, 39, 297–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, J.N.; Del Negro, G.M.B.; Grenfell, R.C.; Vidal, M.S.M.; Thomaz, D.Y.; de Figueiredo, D.S.Y.; Bagagli, E.; Juliano Neto, L.; Benard, G. MALDI-TOF mass spectrometry for rapid identification of the dimorphic fungi Paracoccidioides brasiliensis and Paracoccidioides lutzii. J. Clin. Microbiol. 2015, 53, 1383–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peçanha, P.M.; Bahiense, I.C.; Kruschewsky, W.L.L.; Biasutti, C.; Júnior, C.; Pinheiro, B.G.; Maifrede, S.B.; Camargo, Z.P.; Rodrigues, A.M.; Grão-Velloso, T.R.; et al. Paracoccidioidomycosis due to Paracoccidioides brasiliensis S1 associated with acquired immunodeficiency syndrome: A case report. Rev. Iberoam. Micol. 2020, in press. [Google Scholar] [CrossRef]
- Gegembauer, G.; Araujo, L.M.; Pereira, E.F.; Rodrigues, A.M.; Paniago, A.M.; Hahn, R.C.; de Camargo, Z.P. Serology of paracoccidioidomycosis due to Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2014, 8, e2986. [Google Scholar] [CrossRef]
- Queiroz Junior Lde, P.; de Camargo, Z.P.; Tadano, T.; Rodrigues, A.M.; Takarara, D.T.; Gegembauer, G.; Araujo, L.M.; Hahn, R.C. Serological and antigenic profiles of clinical isolates of Paracoccidioides spp. from Central Western Brazil. Mycoses 2014, 57, 466–472. [Google Scholar] [CrossRef]
- Batista, J., Jr.; de Camargo, Z.P.; Fernandes, G.F.; Vicentini, A.P.; Fontes, C.J.F.; Hahn, R.C. Is the geographical origin of a Paracoccidioides brasiliensis isolate important for antigen production for regional diagnosis of paracoccidioidomycosis? Mycoses 2010, 53, 176–180. [Google Scholar] [CrossRef]
- Camargo, Z.P. Serology of paracoccidioidomycosis. Mycopathologia 2008, 165, 289–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, P.O.; Rodrigues, A.M.; Fernandes, G.F.; da Silva, S.H.; Burger, E.; de Camargo, Z.P. Immunodiagnosis of paracoccidioidomycosis due to Paracoccidioides brasiliensis using a latex test: Detection of specific antibody anti-gp43 and specific antigen gp43. PLoS Negl. Trop. Dis. 2015, 9, e0003516. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; Kubitschek-Barreira, P.H.; Pinheiro, B.G.; Teixeira-Ferreira, A.; Hahn, R.C.; de Camargo, Z.P. Immunoproteomic analysis reveals novel candidate antigens for the diagnosis of paracoccidioidomycosis due to Paracoccidioides lutzii. J. Fungi 2020, 6, 357. [Google Scholar] [CrossRef]
- Melo, A.S.A.; Santos, D.; Lima, S.L.; Rodrigues, A.M.; de Camargo, Z.P.; Finkelman, M.; Colombo, A.L. Evaluation of (1,3)-beta-D-glucan assay for diagnosing paracoccidioidomycosis. Mycoses 2020, 63, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Batista Junior, J.; Berzaghi, R.; Arnaud, A.D.; Fontes, C.J.; de Camargo, Z.P.; Hahn, R.C. Simultaneous infection of human host with genetically distinct isolates of Paracoccidioides brasiliensis. Mem. Inst. Oswaldo Cruz 2010, 105, 62–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lysen, C.; Silva-Flannery, L.; Zaki, S.R.; Gary, J.M.; Lockhart, S.R. Performance evaluation of fungal DNA PCR amplification from formalin-fixed paraffin-embedded tissue for diagnosis; experience of a tertiary reference laboratory. Mycoses 2021. [Google Scholar] [CrossRef]
- Kotepui, M.; Kotepui, K.U.; De Jesus Milanez, G.; Masangkay, F.R. Plasmodium spp. mixed infection leading to severe malaria: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 11068. [Google Scholar] [CrossRef] [PubMed]
- Bastrenta, B.; Mita, N.; Buitrago, R.; Vargas, F.; Flores, M.; Machane, M.; Yacsik, N.; Torrez, M.; Le Pont, F.; Brenière, F. Human mixed infections of Leishmania spp. and Leishmania-Trypanosoma cruzi in a sub Andean Bolivian area: Identification by polymerase chain reaction/hybridization and isoenzyme. Mem. Inst. Oswaldo Cruz 2003, 98, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Guarro, J. Fusariosis, a complex infection caused by a high diversity of fungal species refractory to treatment. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1491–1500. [Google Scholar] [CrossRef]
- Buitrago, M.J.; Aguado, J.M.; Ballen, A.; Bernal-Martinez, L.; Prieto, M.; Garcia-Reyne, A.; Garcia-Rodriguez, J.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Efficacy of DNA amplification in tissue biopsy samples to improve the detection of invasive fungal disease. Clin. Microbiol. Infect. 2013, 19, E271–E277. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Li, L.; Wan, Z.; Chen, W.; Liu, H.; Li, R. Simultaneous detection and identification of Aspergillus and Mucorales species in tissues collected from patients with fungal rhinosinusitis. J. Clin. Microbiol. 2011, 49, 1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarro, J.; Nucci, M.; Akiti, T.; Gené, J. Mixed infection caused by two species of Fusarium in a human immunodeficiency virus-positive patient. J. Clin. Microbiol. 2000, 38, 3460–3462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, G.; Santos, D.W.; Kovacs, J.A.; Nishikaku, A.S.; de Sandes-Freitas, T.V.; Rodrigues, A.M.; Kutty, G.; Affonso, R.; Silva, H.T.; Medina-Pestana, J.O.; et al. Genetic diversity of Pneumocystis jirovecii from a cluster of cases of pneumonia in renal transplant patients: Cross-sectional study. Mycoses 2018, 61, 845–852. [Google Scholar] [CrossRef]
- Ponzio, V.; Chen, Y.; Rodrigues, A.M.; Tenor, J.L.; Toffaletti, D.L.; Medina-Pestana, J.O.; Colombo, A.L.; Perfect, J.R. Genotypic diversity and clinical outcome of cryptococcosis in renal transplant recipients in Brazil. Emerg. Microbes Infect. 2019, 8, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migheli, Q.; Balmas, V.; Harak, H.; Sanna, S.; Scherm, B.; Aoki, T.; O’Donnell, K. Molecular phylogenetic diversity of dermatologic and other human pathogenic fusarial isolates from hospitals in northern and central Italy. J. Clin. Microbiol. 2010, 48, 1076–1084. [Google Scholar] [CrossRef] [Green Version]
- Hahn, R.C.; Macedo, A.M.; Fontes, C.J.; Batista, R.D.; Santos, N.L.; Hamdan, J.S. Randomly amplified polymorphic DNA as a valuable tool for epidemiological studies of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2003, 41, 2849–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.A.; et al. Genome diversity, recombination, and virulence across the major lineages of Paracoccidioides. mSphere 2016, 1, e00213-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, F.L.; Ribeiro, M.A.; Hahn, R.C.; de Melo Teixeira, M.; de Camargo, Z.P.; Cisalpino, P.S.; Marini, M.M. Transposable elements and two other molecular markers as typing tools for the genus Paracoccidioides. Med. Mycol. 2015, 53, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Koishi, A.C.; Vituri, D.F.; Dionízio Filho, P.S.R.; Sasaki, A.A.; Felipe, M.S.S.; Venancio, E.J. A semi-nested PCR assay for molecular detection of Paracoccidioides brasiliensis in tissue samples. Rev. Soc. Bras. Med. Trop. 2010, 43, 728–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaviria, M.; Rivera, V.; Munoz-Cadavid, C.; Cano, L.E.; Naranjo, T.W. Validation and clinical application of a nested PCR for paracoccidioidomycosis diagnosis in clinical samples from Colombian patients. Braz. J. Infect. Dis. 2015, 19, 376–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Species | Primer | Primer Sequence (5’-3’) | Amplicon Size |
|---|---|---|---|
| P. brasiliensis complex | PbraCx-F | TCG TGA TAT AGA CAG CAC CGT TG | 308 bp |
| PbraCx-R | ACG AAC CAT CAA ATC GCG AAC CTA | ||
| P. lutzii | Plu-F | CTT CAT GGC GCC CAA GGA CTT | 142 bp |
| Plu-R | GAC TCC GCT AAG TCT GTC CTT AGG C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, B.G.; Pôssa, A.P.; Della Terra, P.P.; de Carvalho, J.A.; Ricci, G.; Nishikaku, A.S.; Hahn, R.C.; Camargo, Z.P.d.; Rodrigues, A.M. A New Duplex PCR-Assay for the Detection and Identification of Paracoccidioides Species. J. Fungi 2021, 7, 169. https://doi.org/10.3390/jof7030169
Pinheiro BG, Pôssa AP, Della Terra PP, de Carvalho JA, Ricci G, Nishikaku AS, Hahn RC, Camargo ZPd, Rodrigues AM. A New Duplex PCR-Assay for the Detection and Identification of Paracoccidioides Species. Journal of Fungi. 2021; 7(3):169. https://doi.org/10.3390/jof7030169
Chicago/Turabian StylePinheiro, Breno Gonçalves, Ana Paula Pôssa, Paula Portella Della Terra, Jamile Ambrósio de Carvalho, Giannina Ricci, Angela Satie Nishikaku, Rosane Christine Hahn, Zoilo Pires de Camargo, and Anderson Messias Rodrigues. 2021. "A New Duplex PCR-Assay for the Detection and Identification of Paracoccidioides Species" Journal of Fungi 7, no. 3: 169. https://doi.org/10.3390/jof7030169