Rezafungin—Mechanisms of Action, Susceptibility and Resistance: Similarities and Differences with the Other Echinocandins


1
Laboratorio de Micología y Diagnóstico Molecular, Cátedra de Parasitología y Micología, Facultad de Bioquímica y Ciencias Biológicas, Universidad Nacional del Litoral, C.P. 3000 Santa Fe, Argentina
2
Consejo Nacional de Investigaciones Científicas y Tecnológicas, C.P. 3000 Santa Fe, Argentina
J. Fungi 2020, 6(4), 262; https://doi.org/10.3390/jof6040262
Submission received: 30 September 2020
/
Revised: 19 October 2020
/
Accepted: 22 October 2020
/
Published: 1 November 2020
(This article belongs to the Special Issue Antifungal Agents Recently Approved or Under Development)
Abstract
:Rezafungin (formerly CD101) is a new β-glucan synthase inhibitor that is chemically related with anidulafungin. It is considered the first molecule of the new generation of long-acting echinocandins. It has several advantages over the already approved by the Food and Drug Administration (FDA) echinocandins as it has better tissue penetration, better pharmacokinetic/phamacodynamic (PK/PD) pharmacometrics, and a good safety profile. It is much more stable in solution than the older echinocandins, making it more flexible in terms of dosing, storage, and manufacturing. These properties would allow rezafungin to be administered once-weekly (intravenous) and to be potentially administered topically and subcutaneously. In addition, higher dose regimens were tested with no evidence of toxic effect. This will eventually prevent (or reduce) the selection of resistant strains. Rezafungin also has several similarities with older echinocandins as they share the same in vitro behavior (very similar Minimum Inhibitory Concentration required to inhibit the growth of 50% of the isolates (MIC50) and half enzyme maximal inhibitory concentration 50% (IC50)) and spectrum, the same target, and the same mechanisms of resistance. The selection of FKS mutants occurred at similar frequency for rezafungin than for anidulafungin and caspofungin. In this review, rezafungin mechanism of action, target, mechanism of resistance, and in vitro data are described in a comparative manner with the already approved echinocandins.

1. Introduction
Over 1 billion individuals are affected by fungal infections worldwide. It is estimated that 11.5 million suffer serious infections and that more than 1.5 million people die from fungal diseases each year [1,2]. Since the beginning of this century, there have been great advances in medical mycology including an increased number of diagnostic tools, the development of reliable methods for assessing antifungal susceptibility, the increased knowledge in mechanisms of antifungal resistance and fungal virulence factors, and the development of new drugs [1,2]. One of the greatest improvements in this last area was the incorporation of echinocandins into the antifungal armamentarium. Until the appearance of this class of drugs, the therapeutic options for invasive mycoses were limited to polyenes, 5-fluorocytocin, and azoles, which have only two molecular targets, the fungal membrane and the synthesis of nucleic acids [3,4].
2. β-Glucan Synthase Inhibitors Family Tree
There are different chemical classes of fungal glucan synthesis inhibitors including lipopeptides (pneumocandins and echinocandins), glycolipids (papulocandins) [5], and triterpenoids (ibrexafungerp) [6]. Several compounds were evaluated to establish their antifungal potency and toxicity and even some molecules were subjected to clinical trials. Some of the most promising molecules were arbocandins, papulacandins, enfumagungin, arundifungin, and cilofungin [7,8,9,10,11]. Only three echinocandins are currently in clinical use while the safety and efficacy of rezafungin (echinocandin) and ibrexafungerp (triterpenoid) are being evaluated in different clinical trials [12].
The three FDA-approved echinocandin drugs are semisynthetic derivatives of natural products. In 1974, the first echinocandin-like compound was isolated from Aspergillus nidulans var. echinulatus [5,6]. It was named echinocandin B and it became the precedent compound for anidulafungin (LY303366) [5,6,7]. Fifteen year later, pneumocandin was isolated from Lophium arboricola (homotypic synonym: Zalerion arboricola) and it became the compound leading to caspofungin (MK991) [8]. In 1990, three small molecules with antifungal properties were isolated from Coleophoma empetri (homotypic synonym: Thabdostrimina empetri). Of those, WF11899A side chain was chemically modified to obtain micafungin [13]. Rezafungin (CD101, formerly SP3025, Cidara Therapeutics) is the newest member of the echinocandin family and it is the first member of the second generation of this class of antifungals.
3. Chemistry Characteristics and Differences between Rezafungin and the First Generation Echinocandins: Stability, Stability, and Stability
Lipopeptide antifungals are cyclic depsipeptides (oligopeptides that have at least one of its amide groups, -C(O)NHR-, replaced by its ester, -C(O)OR) with different side chains [14]. Echinocandins are lipopeptides with a cyclic hexapeptide core that contains uncommon amino acids in their structure such as 4-hydroxyproline, 3-hydroxy-4-methilproline, dihydroxyornithine, and dihydroxyhomotyrosine [15]. The depsipeptide core is N-linked to different side chains in each of the drugs. Caspofungin, micafungin, and anidulafungin have a fatty-acyl, an isoxazole 3,5-diphenyl-subtituted, and alkoxytriphenyl side chains, respectively [10,15] (Figure 1). When these side chains are removed by deacetylation, the cyclic core showed no antifungal activity, demonstrating its essentiality for echinocandin activity [6].
First-generation echinocandins share several pharmacological problems. They have poor oral absorption and average half-lives of 14 h (mouse model) [10], making daily intravenous administration mandatory [17,18,19]. These drugs also share chemical instability problems such as thermal and hydrolytic degradation for all three echinocandins [17,18,19] plus photodegradation for micafungin [16,17]. These last inconveniences imply manufacturing and storage troubles and several usage problems including the fact that intravenous preparations have to be used within 24–48 h and that other dosage forms are not possible [16].
Rezafungin is being developed to circumvent many of these pharmacological and stability problems. It is a structural analog of anidulafungin. They share the same side chain and a very similar cyclic core in which the hemiaminal region located at the echinocandin cyclic nucleus (C5 ornithine residue) is replaced with a choline aminal ether (Figure 1). This change reduce the chemical degradation that occurs at the hemiaminal region of anidulafungin, increasing its stability and solubility [16] (Figure 1).
After 44 h of incubation in serum and in phosphate buffered saline (PBS) buffer at 37 °C, more than 94% of the initial amount of rezafungin remains active while these percentages are very low for anidulafungin (10% and 50%, respectively). Moreover, after a year of storage in solution (in saline and in 5% dextrose) with exposure to light, high temperature (40 °C), and acidic conditions and being placed in different buffers there was no (or <7%) decrease in rezafungin antifungal activity, and no epimerization (isomer conversion) was produced. Lyophilisate rezafungin is extremely stable. It only showed less than 2% of degradation products after 9 month storage at 40 °C [16]. These facts are beneficial in terms of dosing flexibility and storage of the pharmaceutical product but also in terms of manufacturing. Moreover, rezafungin has a higher half-life (>130 h, >5 times the half-life of the other echinocandins) [20,21,22]. These properties would allow rezafungin to be administered once-weekly (intravenous) [23] and to develop of new forms of administration such as topical and subcutaneous application. In addition, higher dose regimens were tested and no evidence of toxic effect were noticed in hepatocytes of rats exposed to high doses of rezafungin [24]. Oppositely, using anidulafungin in similar doses and regimen, researchers observed hepatocellular necrosis. This will eventually prevent (or reduce) the selection of resistant strains [21,25].
4. Rezafungin and the Older Echinocandin’s Target: The β-Glucan Synthase Complex
Fungal cell wall is an extracellular matrix composed by an inner layer consisting of carbohydrate polymers and an outer layer of glycoproteins [22,26,27,28]. The main carbohydrates of the fungal cell wall are the β-D-1,3-glucans and α-1,3-glucans accompanied by β-D-1,6-glucans and chitin [6,29,30,31]. These carbohydrates constitute the so-called alkali-insoluble nucleus of the cell wall, which is common for most fungi [32]. On the other hand, the outer layer is substantially different in composition in different fungi. In Candida albicans, this layer is mainly formed by mannosylated proteins fixed to the carbohydrate core by glycosylphosphatidylinositol residues [33,34]. In Aspergillus fumigatus hyphae (and other filamentous fungi), the mannan chains are shorter than in Candida spp. (lower molecular weight), are modified with β-(1,5) galactofuran, and are directly attached to the carbohydrate core [35]. In the Basidiomycete Cryptococcus spp., a glucuronoxylomannan-galactoxylomannan capsule is attached to α-1,3-glucans located in the outer cell wall layer while the inner layer is composed of β-D-1,3-glucans, β-D-1,6-glucans, and deacelylated chitin (chitosan) [36,37]. In Ajellomycetaceae such as Blastomyces dermatitidis and Histoplasma capsulatum, the outer layer of their cell walls is composed by α-(1,3) glucans, which reduce the recognition of the β-(1,3) glucans of the inner layers by immune cells [38,39,40] (Figure 2).
Despite the described differences in composition between species, glucans are the most important structural component of the fungal wall. This carbohydrate alone comprises more than 50% of the dry weight of this structure. Of those glucans, the one with glucose units connected by 1,3 links (β-D-1,3-glucans) represents between 65–90% of the glucan polymers [41]. Responsible for the synthesis of this component of the wall is the 1,3-D-glucan synthase complex (EC 2.4.1.34.). It is a transmembrane protein complex of partially known qualitative and quantitative composition. It is formed by at least two main subunits: Rho1p and Fksp. The former is a regulatory element of the complex [42]. It has promiscuous functions since its participation in several biosynthesis pathways has been demonstrated [43,44,45,46]. Rho1p is known to have Guanosine triphsphatase (GTPase) activity (Ras-like GTP-binding protein) [47,48,49], and as a subunit of the 1,3-β-D-glucan synthase complexes, it is estimated that one of its functions is to provide the necessary energy for the 1,3-β-D-glucan bonds to occur [50]. This idea is supported by the fact that GTP is necessary to produce 1,3-β-D-glucans in vitro using semi-purified 1,3-β-D-glucan synthase complexes [51,52,53]. A second function of Rho1p is to sense stress signals due to 1,3-β-D-glucan depletion and to activate as a response a wide variety of effectors ranging from protein kinase C, SLT2 [44], calcineurin/CRZ1/Ca2+, and HOG [54,55] to Hsp90p, Mkc1p, and Sgt1p [46,56,57,58,59].
As a complex, 1,3-β-D-glucan synthase is a glucosyltransferase that generate glycosidic bonds (β-1,3 links). It catalyzes the reaction Uridine diphosphate-glucose (UDP-glucose) + {1,3-β-D-glucosyl}n = UDP+ {1,3-β-D-glucosyl}n+1, incorporating around 6 nmol UDP-glucose/min with a good affinity for its substrate (average Km and Vmax for echinocandin susceptible C. albicans and Candida glabrata: 0.099 ± 0.022 mM and 5.962 ± 0.723 nmol/min and 0.133 ± 0.015 mM and 6.812 ± 0.246 nmol/min, respectively) [51,52]. The Fksp subunits are the real catalyst subunits of the complex and the specific target of echinocandin drugs. This last conclusion is based on the fact that resistance and the subsequent treatment failures is conferred by amino acid substitutions in the Fksp (see below for more details) [60]. Three different homologous genes named FKS1, FKS2, and FKS3 encode these proteins. They were originally identified in the 1990s in Saccharomyces cerevisiae [61]. Later, FKS orthologs were described in several fungal species [53,61,62,63,64,65,66,67] and their particular functions and the way in which their expression is regulated appears to have small differences.
In diploid organisms such as most Candida spp., the disruption of both FKS1 alleles affects cell growth and leads to hypersensitivity to FK-506 (calcineurin phosphatase inhibitor). Thus, FKS2 is considered a backup gene responsible for producing a cell wall in response to glucose starvation and osmotic stress (as happens when 1,3-β-D-glucans of the wall are reduced) through the calcineurin pathway [68]. On the other hand, FKS1 expression predominantly occurs during growth on glucose and is constitutively regulated during the cell cycle [68]. The disruption of both alleles of FKS1 and FKS2 is lethal [68,69], suggesting that (i) both proteins are component of the complex and (ii) that both subunits are essential and have overlapping function [7,68]. In haploid organisms such as C. glabrata, the disruption of FKS1 and FKS2 is also lethal but the disruption of FKS1 and FKS3 or FKS2 and FKS3 is not. Moreover, in this yeast it seems that FKS1 and FKS2 are functionally redundant [70]. In the haploid Aspergillus fumigatus also, FKS1 is non-essential and its knockout leads to a compensatory increase of cell wall galactosaminogalactan and chitin and the depletion of galactomannan [50]. Oppositely, in Yarrowia lipolytica (heterotypic synonym: Candida lipolytica), there is only one FKS homologue (FKS1) that is essential for viability [62].
This enzyme complex is an ideal antifungal target since it is essential for viability of fungi and it is not present in mammal cells.
5. The Place of Rezafungin in the Clinical Setting
Anidulafungin, caspofungin, and micafungin are the drugs of choice to treat Candida spp. deep-seated infections [55] while they are recommended as salvage therapy (either alone or in combination with azole drugs) for different types of aspergillosis [71]. Moreover, some reports described the anti-pneumocystis activity of the first-generation echinocandins since they showed the capacity to inhibit cysts but not trophic cells [72]. However, they were not proposed as a choice to treat Pneumocystis spp. infections.
Everything seems to indicate that rezafungin share the same spectrum and it will be useful in the same clinical scenarios as the first generation of echinocandins. ReSTORE and STRIVE [73] clinical trials are being carried out to prove the usefulness of rezafungin (both compared with caspofungin) as treatment of candidemia and other invasive candidiasis and as treatment of these mycoses followed by oral step-down treatments. Additionally, rezafungin is also being tested as a therapeutic tool to treat acute and recurrent vaginal yeast infections (RADIANT study) [74] and as a prophylaxis of invasive fungal infections (including Candida spp., Pneumocystis spp., and Aspergillus spp.) in adults undergoing allogenic blood and marrow transplantation (ReSPECT study) [75].
After approval, rezafungin will expand the clinical span of echinocandins as a wider ranging prophylaxis tool and as a way to treat recurrent and refractory vaginal candidiasis.
6. Molecular Mechanism of Action of Rezafungin
As mentioned, echinocandins inhibit cell wall formation, specifically 1,3-β-D-glucan synthesis. The consequence of the depletion of 1,3-β-D-glucans in the cell wall are cell morphology changes resulting in osmotic instability and the subsequent cell death and/or inhibition [76]. These antifungals act as fungicidal agents against most Candida spp. (such as C. albicans, C. dubliniensis, C. krusei, and C. tropicalis) [76,77,78]. For other Candida species showing naturally occurring polymorphisms at Fksp that alter its in vitro echinocandin susceptibility (Candida parapsilosis complex and Meyerozima guilliermondii (heterotypic synonym: Candida guilliermondii)) [79,80], these drugs behave differently in vitro, requiring more time and higher doses to reach the fungicidal threshold [81,82], while other emerging species as Candida auris are tolerant in vitro to these drugs [83].
In filamentous fungi, echinocandin effects are dependent on the relative cell position within the hyphae. The inhibition of 1,3-β-D-glucan synthase complexes is produced in apical cells, leaving the subapical cells almost intact [13]. This is possible because the cell wall constitutive polymers are in continuous production, modification, and degradation during the cell cycle and hyphal growth. Specifically, it was demonstrated that 1,3-β-D-glucan synthesis is concentrated where the cell wall is remodeling. Fks1p is co-localized with actin patches that are constantly moving on the membrane cell surface to the locations were this polymer is mostly needed [84]. In apical cells, echinocandins produce a misbalance of the glucan content of the wall. They inhibit 1,3-β-D-glucan production, not altering its degradation speed. This fact leads to the bursting of these cells due to osmotic pressure [85]. This kind of apical inhibition of growth is used to establish the minimal effective concentration (MEC) in susceptibility testing assays for filamentous fungi. The MEC is the lowest echinocandin drug concentration that leads to the growth of compact hyphal forms that provides accurate and reproducible susceptibility data [86].
To comprehend the molecular mechanism of action of echinocandins, we need to understand how these cyclic peptides with lipid tails interact with their targets.
Fungal 1,3-β-D-glucan synthase complexes were never fully purified. Some ideas about how this complex works could be glimpsed when semi-purified enzymes (using a methodology called trapping) are studied. Among the data obtained, a factor that stands out is its non-competitive way of inhibition, confirming the inhibition of this complex by echinocandins and some biochemical parameters of the complex as a whole (Ki, Km, and Vmax) [51,52,87]. Thus, we know that these drugs bind to an allosteric site of the enzymes (different from the active site) and that the substrate (UDP-glucose) is located in the cytosolic surface of the membrane and the product (1,3-β-D-glucans) is external [7,61,68]. Additionally, the description of the molecular mechanisms of clinical echinocandin resistance allowed us to have an insight into how these drugs interact with Fksp. These mechanisms are exclusively related to amino acid substitutions in two limited Fksp regions (named hot spot regions 1 and 2). Another hot spot region was suggested and named hot spot 3, which was linked to intrinsic resistance in Scedosporium spp. [41,51,52,53,88,89]. These facts demonstrated that echinocandins exert their inhibitory activity by binding to these regions. Efforts to effectively demonstrate where the substrate-binding sites are located were made using Neurospora crassa partially purified β-1,3 glucan synthase. The obtained data located this site are in a region of 200 residues (amino acids (aa.) between the residues V1073 and R1277 using C. albicans Fks1p numbering) [90]. In 2012, Johnson and Edlind described the Fksp putative substrate and echinocandin binding sites more accurately using S. cerevisiae as model organisms [91]. They combined in silico and site-directed mutagenesis analysis together with the analysis of C-terminal fusions (Fks1p-hemagglutinin-Suc2-His4C), followed by His4C expression and Suc2 glycosylation assays [92]. They found that the three described hot spot regions are located in the outer layer of the membrane and are adjacent to each other and that they are external or partially embedded to the plasma membrane. Hot spot regions 1 and 3 are located within transmembrane segments 5 and 6 while hot spot region 2 is located between transmembrane segments 7 and 8. They suggested that they would be possibly three-dimensionally arranged, forming a putative echinocandin-binding pocket, and that echinocandin would be interacting outside the cytoplasm, not requiring the entry into the cell (Figure 3). Additionally, they demonstrated that the glycosyltranferase domain (catalytic domain) should be located in the cytoplasm face of the membrane. It is formed mainly by hydrophilic amino acids (between I715 and H1294 for S. cerevisiae Fks1p and amongst the residues I717 and H1298 in C. albicans Fks1p) and it is surrounded by the hot spot regions [91] (Figure 3). Following this model, the lipid tail (essential for echinocandin activity) [93] would interact with the partially membrane-embedded hot spot 3 region (L692-N702 in C. albicans Fks1p) or near the hot spot 1 region while the cyclic hexapeptide core would do so with hot spot 1 and 2 regions. What has been described indicates that the binding of echinocandin to the three hot spot regions reduces or cancels any glycosyltransferase activity, leading to the depletion of beta glucans and osmotic instability of fungal cells. Considering that (i) the putative localization of the hot spot 1 and 2 regions (opposite to each other), (ii) that hot spot 3 regions bind to the echinocandin tail, (iii) that all three hot spot regions are arranged surrounding the hydrophilic catalytic domain and forming an echinocandin binding pocket, (iv) that echinocandin resistance is almost exclusively linked with mutations in the hot spot 1 and 2 regions, and that (v) hot spot regions are located in the extracellular surface of the membrane, we can speculate that the echinocandin cyclic core is binding or interacting with both hot spot 1 and 2. Moreover, knowing that β-1,3-D glucan synthase substrate (UDP-glucose) is mainly found in the cytosol while the glucan polymer is external, we can also speculate the echinocandins are preventing the exit of the formed polymer by some type of steric hindrance (Figure 3).
7. In Vitro Data
The analysis of rezafungin in vitro susceptibility data was performed using a search on pubmed.ncbi.nlm.nih.gov using as keywords “rezafungin” and “CD101 antifungal”. The antifungal susceptibility and surveillance papers were included in the data analysis. Moreover, certain papers with other aims were included, considering the fact that authors published susceptibility data as secondary data or as data that support further studies. Most of the in vitro data were obtained using Clinical and Laboratory Standards Institute (CLSI) reference protocols (documents M27
A3 and M274th ed. for yeasts and M38A2 and M38-3rd ed.) [100,101,102,103,104,105,106,107]. Recently, some differences in rezafungin minimal inhibitory concentration (MIC) were noticed when using the European Committee on Antimicrobial Susceptibility Testing (EUCAST) method and different polystyrene plates were used (culture treated and untreated). MICs values were within three dilutions and untreated polystyrene plates were better to separate wild type and non-wild type strains and also improved the interlaboratory reproducibility [108].
Little difference was seen between reports when MIC/MEC ranges and MIC50 and MIC90 data were evaluated, despite the used reference method and the different laboratory where the tests were performed (Table S1).
The analyzed papers included susceptibility data of 7818 strains of 17 different Candida spp. and 429 isolates of 9 Aspergillus spp. (some of the strains were identified at the section level). In one of the reports, 44 strains of Cryptococcus neoformans var. grubii were studied. All the strains showed very high MIC values (>8 µg/mL), confirming the well-known inactivity of echinocandins in Basidiomycetes [109]. Despite the variety of species included, the published data are focused mainly in the most common Candida spp. (C. albicans, C. glabrata, C. krusei, C. parapsilosis, and C. tropicalis), C. dubliniensis, and C. auris (Table 1). The in vitro rezafungin susceptibility was studied in only 61 echinocandin-resistant strains (FKS mutants). Out of these mutants, the vast majority were C. glabrata and C. albicans (as occurs in clinical settings) (Table 2).
Rezafungin MICs were very similar to the values obtained for the other echinocandins used as comparators. When MIC50 ratios were analyzed (Rezafungin MIC50/comparator MIC50), small differences were found, especially when compared with anidulafungin MIC50. For C. albicans and C. tropicalis, rezafungin MIC50 values were slightly higher than those for anifulafungin (rezafungin MIC50/anidulafungin MIC50: 2.26- and 2.17-fold, respectively), lower for caspofungin (0.38- and 0.65-fold, respectively), and similar for micafungin (1.56- and 1.39-fold, respectively). For C. glabrata, C. krusei, and C. parapsilosis sensu stricto, MIC50 values were almost equal for rezafungin than for anidulafungin (rezafungin MIC50/anidulafungin MIC50: 1.03-, 0.9-, and 0.64- fold, respectively). Rezafungin seems more potent in vitro than caspofungin and micafungin against C. krusei (MIC50 ratios: 0.12 and 0.31, respectively), while caspofungin was more potent than rezafungin when C. parapsilosis was tested (rezafungin MIC50/caspofungin MIC50 ratio: 2.58).
As with the other echinocandins, Candida spp. showing intrinsic reduced echinocandin susceptibility (IRES) phenotype (C. parapsilosis sensu lato, C. guilliermondii, C. lusitaniae, etc.) showed 10- to 50-fold higher MIC50 than those of C. albicans (e.g., rezafungin MIC50 geometric means of 1.12 vs. 0.023 for C. parapsilosis sensu stricto and C. albicans, respectively) (Table 1).
Rezafungin was more potent than caspofungin and micafungin against C. auris (rezafungin MIC50/comparator MIC50 ratios = 0.31 and 0.65, respectively). On the other hand, rezafungin MIC50s were twofold higher than those for anidulafungin (MIC50 ratio 2.55) (Table 1). Rezafungin, anidulafungin, caspofungin, and micafungin MIC values for C. auris were 6.57, 5.81, 8.24, and 16.7 times higher than for C. albicans, respectively.
Rezafungin shares similar potency against FKS mutants than the other echinocandins. The MIC values for mutants were between 6 to 50 times higher for mutants than for those of wild type strains of the same species (Table 2). The highest MIC differences between mutants and wild type strains were observed in C. auris FKS mutants. For this species, only four mutants harboring the same mutation (S6639P equivalent to S645P for C. albicans) were studied. This amino acid substitutions are responsible for the most prominent resistance phenotype in all Candida spp. [60].
Turning to Aspergillus spp., rezafungin was very potent for all the studied species, even for cryptic multi-resistant species of the Aspergillus section Fumigatii. For these intrinsically azole-resistant species and for secondary resistant Aspergillus fumigatus sensu stricto, rezafungin would be a good treatment option, as has been suggested for the other echinocandins (treatment of azole refractory aspergillosis) [71] (Table 3, with more detail in Table S1).
There were other reports that were not included in the initial analysis since they used different antifungal susceptibility testing methods, but it is nonetheless important to analyze them due to the potential use of rezafungin as treatment of vulvovaginal candidiasis (topical formulation) and to act in biofilms. Rezafungin susceptibility of Candida species isolated from patients with acute and recurrent vulvovaginal candidiasis was evaluated using a modified CLSI method (performed at pH = 4 to resemble vaginal pH) [112]. Authors found that rezafungin showed a potent activity against C. albicans, C. glabrata, C. parapsilosis, and C. tropicalis isolated from vulvovaginal infections. Moreover, these MIC values were similar to those obtained for Candida spp. isolated from deep-seated infections [102,103,104]. The obtained MICs (even for C. parapsilosis) were below the intravaginal rezafungin concentration that would be reached after topical administration [112]. To evaluate the activity of rezafungin against C. albicans biofilms, Chandra et al. studied the metabolic activity of the cells in early and mature biofilms (formed on silicone elastomer discs) and the thickness of the formed structures by confocal microscopy [113]. When rezafungin was used at 0.25–1.00 µg/mL, the authors observed a reduction in biofilm thickness (in both early and mature preformed biofilms). In addition, at the same drug concentration, the development of mature biofilms was prevented [113].
8. Mechanisms of Resistance
As with any antibiotic, microbiological resistance to echinocandin drugs can be divided into innate or inherent and secondary or acquired resistance. The first definition includes species that show non-wild type susceptibility patterns of all or almost all strains (susceptibility testing is unnecessary to establish its resistance) [114]. The former designation includes strains that obtain the ability to resist the activity of a particular antimicrobial agent to which it was previously susceptible [115]. Moreover, for this class of antifungals, there is a third group of species that has shown in vitro intrinsic reduced susceptibility [60]. These three types of phenotypes have to be differentiated from low level-resistance and/or tolerance to echinocandins.
This section will be dedicated to summarizing what is known as clinical resistance mechanisms, thus the use of a cited bibliography is suggested to expand on the topic of tolerance and response to stress [44,46,54,55,56,57,58,59]. As a brief summary, we can state that the reduction of the β-1,3-D-glucan content of the cell wall leads to an important cellular stress that is sensed by Rho1p. In response, a complex network of stress response pathways and other compensatory pathways are activated, aiming to alter the qualitative composition of the cell wall to maintain its structural integrity. The main event is the increase of chitin synthesis that replaces in part the structural function of β-1,3-D-glucans, decreasing the cell sensitivity to echinocandins [116,117,118,119]. This shift on the relative composition of the cell wall is responsible (at least in part) for the paradoxical growth effect seen in vitro in several Candida spp. isolates when confronted with high echinocandin doses [120,121,122]. Other tolerance and reduced sensitivity to echinocandin mechanisms include changes in quantity and quality of membrane sphingolipids [123] and chromosomal instability (aneuploidy and other genomic rearrengements) [95,124,125]. As a conclusion, these mechanisms stabilize cells and soothe the effect of the drug against them, giving time for stable mechanisms to be selected [98].
The main mechanisms of echinocandin resistance, considered the universal mechanisms, are the FKS hot spot mutations [51,52,53,60,79,80,98]. These mechanisms are the main responsible factors for both intrinsic and secondary resistance to echinocandins with slight differences.
The association between echincandin treatment failures (secondary resistance) with the presence of a mutation in one (or both) of the hot spot regions of the Fksp is considered an independent risk factor for treatment failure for C. glabrata infections [96]. The identification of these mutations is better than MIC as a predictor of clinical response (specially for caspofungin MIC) [96] and as a predictor of the enzyme complex insensitivity to these drugs [51,52].
Subtitutions in Fksp subunits conferring elevated MIC values are limited to two highly conserved amino acid regions named hot spot regions. Early reports consider that these regions should cover the amino acid residues between F641 and P649 (hot spot 1) and D1357 and L1364 (hot spot 2) (C. albicans Fks1p numbering following accession number # D88815) (Figure 3B and Figure 4) [51,52,53,99]. Later, it was suggested that hot spot 2 should be limited to only one amino acid (R1361 following the same aa. #) considering that this is the main amino acid of this hot spot that is linked with echinocandin resistance [98]. These mutations were described in the FKS1 genes of diploid Candida spp. (most of the Candida spp. considered human pathogens, e.g., C. albicans, C. tropicalis, C. krusei) both in homo- and heterozygosity (they are dominant) [51,60,98,99,126,127,128,129,130]. In C. glabrata (haploid Candida spp.), echinocandin resistance was linked in FKS1 and in FKS2 (> prevalence for FKS2 mutants) and it is the only species that would present mutations (or deletions) in both genes at the same time [52,70].
The described amino acid subtitutions reduce the sensitivity of glucan synthase to echinocandins between 30- to 3000-fold (IC50s), which translate into 10- to more than 100-fold MIC increase [51,52,60,70,98,99,127]. These resistance phenotypes have a cost to the fitness of the strains (confirmed with isogenic C. albicans strains using a competitive model of murine candidiasis). It was attributed to a reduction in the catalytic efficacy of the β-1.3-D-glucan synthase complex harboring mutant Fksp subunits (lower Vmax with no Km changes) that led to an altered cell wall composition [131]. As MIC and IC50 values, these Vmax reductions ranged between a 20% and 80% reduction for C. albicans and C. glabrata FKS mutants [51,52]. The described phenotype differences (Vmax/fitness) had a correlation with the prevalence of each of the different affected residues of the hot spots. Thus, in C. albicans, the substitutions at F641 and S645 (aa. #D88815) account for around 75–80% of the mutants. These mutants showed the lowest Vmax reduction with its consequent slight reduction in their fitness (when compared with other mutants). Similarly, in C. glabrata, the most common substitutions are at FKS2, but still are at equivalent positions than for C. albicans Fks1p (at its F659 and S663 residues, aa. # YLR342W). Unfortunately, this higher prevalence and relative virulence is also accompanied by a more marked resistance phenotype [51,52]. Amino acid substitutions in other positions of the hot spot confer no or less-pronounced phenotypes. These differences may be explained through considering the putative echinocandin-binding pocket model described before [91], together with the prediction that the hot spot regions are part of the protein arranged as an α-helix [132]. When these regions are represented as a helical wheel plot (or Edmundson wheel), the amino acid residues of the hot spot regions that are mainly linked with echinocandin resistance group together (Figure 3). These plots represent secondary structures with a helical potential arranging where the amino acids are drawn in a rotating manner with an angle of rotation of around 100° [94]. Thus, the amino acid sequence is represented as viewing the helix from above, showing whether certain amino acids are concentrated in one side of the helix or not. In the Fksp hot spot 1 region, the residues linked with echinocandin resistance (F641, S645, D648, and P649 following C. albicans numbering aa. # D88815) are concentrated in one side of the wheel. Similarly, the hot spot 2 residues linked with echinocandin resistance (W1358 and R1361) are also grouped together in the Edmundson wheel.
As the other echinocandin, rezafungin is inactive against Fusarium spp., Scedosporium spp., Lomentospora prolificans, Mucorales, Histoplasma capsulatum, and Basydiomycetes, including Cryptococcus spp., Trichosporon spp., etc. [60,98,133]. The mechanisms involved in these intrinsic resistances vary depending on the studied species. In the filamentous Ascomycetes Fusarium spp. and L. prolificans, the mechanism of resistance involves naturally occurring polimorfisms at the hot spot regions of the Fks1p. The implicated residues are located at F684Y, being equivalent to those related with high-level echinocandin clinical resistance in C. albicans (F641X) [134] (Figure 3B and Figure 4). For Scedosporium spp., the proposed mechanism involved is a polymorphism located in the so-called hot spot 3 region [89]. Although described in these pathogens and in Saccharomyces cerevisiae laboratory mutants [89], this region was not implicated in acquired clinical echincandin resistance [60,98]. For Mucorales, Ajellomycetaceae, and Basydiomycetes, the molecular mechanisms of their intrinsic echinocandin resistance is not clear. Some reports have suggested that β-1,3-D-glucans are not as important for cell wall integrity in these fungal pathogens in comparison to other fungi (lower content of β-1,3-D-glucans) [32]. Others suggest that Cryptococcus spp. and Histoplasma capsulatum melanization [135] and Cryptococcus neoformans liplid flippase subunit Cdc50 [93] may play a role in their reduced echinocandin susceptibility. Moreover, Cryptococcus neoformans β-1,3-D-glucan synthase complex was partially purified and its FKS subunits were sequenced. No mutations were observed, and the isolated enzyme complex was susceptible in vitro to echinocandins. These two results discard the implication of alterations in Fksp as a molecular mechanism of resistance in this species [65].
As mentioned, there are some Candida spp. that show naturally occurring polymorphisms at FKS hot spot regions. These species were crowded together within the intrinsic reduced echinocandin susceptibility (IRES) group that includes C. parapsilosis complex (C. parapsilosis sensu stricto, C. metapsilosis, and C. orthopsilosis), C. guilliermondii, etc. They showed higher echinocandin MIC values (>10-fold) and their glucan synthase complexes showed higher IC50s than the other Candida spp. The CLSI and EUCAST antifungal susceptibility testing committees propose higher breakpoints and epidemiological cut-off values to consider strains of these species as resistant or non-wild type, respectively [136,137,138]. Early clinical trials showed that the treatment of C. parapsilosis infections showed better results (clinical cure) when treated with fluconazole than with anidulafungin [139]. However, echinocandins are still the drug of choice to treat infections caused by IRES species [55]. Species with IRES phenotype show polymorphisms in residues located in the C-terminal end of the hot spot 1 region or in residues located in the opposite side of the helical wheel of the residues with prominent phenotype (they are grouped together) (Figure 3). This is the case for C. parapsilosis complex (polymorphism P649A, same numbering in C. albicans aa. # D88815) and for C. guilliermondii (L633M and T634A, equivalent to L642 and T643 of aa. # D88815) (Figure 4). The implication of these polymorphism in the IRES phenotype were molecularly confirmed [79,80]. Moreover, echinocandins need more time and higher doses to reduce the starting inocula by three logs (fungicidal threshold). This fact was confirmed using time-killing curves and minimal fungicidal concentration tests [77,81,140].
Turning to Candida auris, this species has some of the characteristics mentioned for species with IRES phenotype and other characteristics of the normally echinocandin-susceptible species. Epidemiological cut-off values for C. auris were firstly established for all echinocandins at 0.25 µg/mL (using Indian isolates) [141]. Later, the U.S. Center for Disease Control and Prevention (CDC) proposed tentative breakpoints on the basis of the MIC modal distributions of more than 100 strains from different geographic locations (anidulafungin ≥ 4 µg/mL, caspofungin ≥ 2 µg/mL, and micafungin ≥ 4 µg/mL) [85]. These breakpoints are very similar to those for C. parapsilosis (anidulafungin ≥ 8 µg/mL and micafungin ≥ 4 µg/mL) [138]. Killing kinetics assays demonstrated that C. auris is tolerant to echinocandins and that there were no fungicidal activity against this species [83]. On the other hand, C. auris shows a “wild type” hot spot region (FLTLSLRDP)) (Figure 4), and echinocandin resistance was related to acquired substitution post-echinocandin treatment [81,130,142].
Turning to rezafungin and considering that it is an anidulafungin analog with extended half-life, it was thought from the start that it would be likely to develop resistance. Locke et al. characterized the in vitro resistance development of four Candida spp. (C. albicans, C. glabrata, C. krusei, and C. parapsilosis) using two different mutant selection methods (spontaneous resistance and resistance after successive passages) and using anidulafungin and caspofungin as comparators [143]. The selection of spontaneous mutants (single-step/high-dose method) occurred at very low frequencies for all the three drugs. Mutations at FKS occurred at similar ranges for rezafungin than for anidulafungin and caspofungin (1 mutant per every 1.5 × 108 cells, per 1.6 × 107–3.9 × 109 cells, and 3.5 × 107–3.9 × 109 cells, respectively) [144]. As for the other echinocandins, rezafungin selected FKS mutants easier in C. glabrata (haploid) than in C. albicans, while C. parapsilosis and C. krusei seemed to have lower potential of FKS-linked resistance development [130,144,145]. Moreover, cross-resistance was observed in FKS mutants independently of the used echinocandin for mutant selection. When using successive passages, 20 passages were needed to obtain strains with increased MIC values. However, most of the mutants selected by passages showed no FKS mutations and those with no hot spot substitutions showed slight MIC increases (2–4-fold) [144]. When rezafungin was used as a selector of spontaneous mutants, only the so-called strong phenotype mutations (which were the most prevalent in clinical settings) were obtained, including S645P in C. albicans Fks1p and F659Δ and S663F in C. glabrata Fks2p [60,98,144,146]. Less common or barely described substitutions were also encountered in C. glabrata Fks2p as R665G, D666H, D666N, D666Y, and R1378S (the latter in hot spot 2) [144,147] (Figure 3). Similar substitutions were obtained when serial passages were used to select mutants. Some were commonly described in clinical isolates, such as S645Y in Fks1p of C. albicans, and D632Y and F659I in C. glabrata Fks1p and Fks2p, respectively. As with spontaneous mutant selection, the same “rare” substitutions were found, such as D666N and D666Y in C. glabrata Fks2p and the never previously described I1366S in the hot spot 2 of the Fks1p of C. krusei [144].
Zhao et al. studied the kinetic inhibition (IC50) of partially purified C. albicans β-1,3-D-glucan synthases by micafungin and rezafungin. They found that both echinocandins were able to inhibit wild type enzyme complexes at very low concentrations (around 15 ng/mL for both drugs). However, rezafungin was shown to be a better inhibitor of mutant complexes harboring Fks1p subunits with F641S substitutions in comparison with micafungin (24-fold vs. 100-fold increase in IC50 values when compared with wild type enzyme complexes). On the other hand, enzyme complexes with homozygous S645P mutation showed similar IC50 values for both echinocandins (>140-fold increase) [100]. When C. glabrata was studied, the semi-purified enzyme complexes harboring Fks2p subunits with F659Δ deletion was very resistant to both echinocandins (IC50 > 10,000 ng/mL), while the one harboring the S663P mutation at Fks2p was more resistant to rezafungin than for micafungin [100].
The same group of researchers compared the mutant prevention concentration of these two echinocandins [100]. This concept was coined for bacteriology at the beginning of this century and is defined as the concentration above which it is possible to inhibit the population of strains able to persist at concentrations above the MIC and that are the source of future mutants with stable resistance phenotypes. Thus, if this concentration is reached, the selection of mutants is less likely [148]. For rezafungin and micafungin, the mutant prevention concentration was 16 µg/mL for C. albicans and C. glabrata for both drugs. Taking all these data together, it seems that rezafungin has a low potential of resistance development that is comparable to other echinocandins. However, rezafungin showed a better tissue penetration [149] and better PK/PD pharmacometrics [150,151]. These facts would allow rezafungin to reach higher tissue concentrations, surpassing the mutation selection threshold.
9. Conclusions
- Rezafungin is a new β-glucan synthase inhibitor that is chemically related to anidulafungin, with better stability, tissue penetration, and PK/PD pharmacometrics.
- Rezafungin shares targets and mechanisms of action with the other echinocandins but with an improved safety profile, allowing the potential administration of higher doses.
- Rezafungin is much more stable in solution than the older echinocandins. This fact is beneficial in terms of dosing flexibility and storage but also in terms of manufacturing.
- Rezafungin is proposed as a drug to be administered once-weekly (intravenous). New forms of administration are in development such as topical and subcutaneous forms.
- Higher dose regimens will eventually prevent (or reduce) the selection of resistant strains.
- Rezafungin MIC50s against Candida spp. mimicked those of anidulafungin. It is very potent in terms of in vitro activity against Aspergillus spp. (including multidrug-resistant cryptic species). Still, its spectrum is narrow. Basidiomycetes, Mucorales, Fusarium spp., and Ajellomycetaceae are intrinsically resistant to rezafungin.
- Rezafungin shares with the older echinocandins the same target and the same mechanisms of resistance. The selection of FKS mutants occurred at a similar frequency for rezafungin than for anidulafungin and caspofungin.
- When rezafungin begins to be used clinically, the appearance of described rare mutations in C. glabrata Fks2p (R665G, D666H, D666N, D666Y, and R1378S) should be monitored as they may be a unique resistance mechanism for this drug. This monitoring will be essential to be able to adapt the few molecular diagnostic methods of echinocandin resistance that have been developed.
Supplementary Materials
The following are available online at https://www.mdpi.com/2309-608X/6/4/262/s1, Table S1: In vitro data for rezafungin and the other echinocandins used ass comparators for Candida spp. and Aspergillus spp. determined by CLSI and EUCAST susceptibility testing method.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases—Estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Gobal Action Found for Fungal Infections (GAFFI). Improving Outcomes for Patients with Fungal Infections across the World. A Road Map for the Next Decade. Available online: https://www.gaffi.org/wp-content/uploads/GAFFI_Road_Map_interactive-final0415.pdf (accessed on 29 October 2020).
- Wall, G.; Lopez-Ribot, J.L. Current Antimycotics, New Prospects, and Future Approaches to Antifungal Therapy. Antibiotics 2020, 9, 445. [Google Scholar] [CrossRef] [PubMed]
- Gintjee, T.J.; Donnelley, M.A.; Thompson, G.R. Aspiring Antifungals: Review of Current Antifungal Pipeline Developments. J. Fungi 2020, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onishi, J.; Meinz, M.; Thompson, J.; Curotto, J.; Dreikorn, S.; Rosenbach, M.; Douglas, C.; Abruzzo, G.; Flattery, A.; Kong, L.; et al. Discovery of novel antifungal (1,3)-β-D-glucan synthase inhibitors. Antimicrob. Agents Chemother. 2000, 44, 368–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Złotko, K.; Wiater, A.; Waśko, A.; Pleszczyńska, M.; Paduch, R.; Jaroszuk-Ściseł, J.; Bieganowski, A. A report on fungal (1→3)-α-D-glucans: Properties, functions and application. Molecules 2019, 24, 3972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, C.M. Fungal β(1,3)-D-glucan synthesis. Med. Mycol. Suppl. 2001, 39, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Pelaez, F.; Cabello, A.; Platas, G.; Díez, M.T.; Del Val, A.G.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]
- Cabello, A.M.; Platas, G.; Collado, J.; Díez, T.M.; Martín, I.; Vicente, F.; Meinz, M.; Onishi, J.C.; Douglas, C.; Thompson, J.; et al. Arundifungîn, a novel antifungal compound produced by fungi: Biological activity and taxonomy of the producing organisms. Int. Microbiol. 2001, 4, 93–102. [Google Scholar] [CrossRef]
- Kurtz, M.B.; Rex, J.H. Glucan synthase inhibitors as antifungal agents. Adv. Protein Chem. 2001, 56, 423–475. [Google Scholar]
- Barrett, D. From natural products to clinically useful antifungals. Biochim. Biophys. Acta—Mol. Basis Dis. 2002, 1587, 224–233. [Google Scholar] [CrossRef] [Green Version]
- ClinicalTrials.gov Ibrexafungerp. Available online: https://clinicaltrials.gov/ct2/results?cond=&term=ibrexafungerp&cntry=&state=&city=&dist= (accessed on 12 September 2020).
- Bowman, J.C.; Hicks, P.S.; Kurtz, M.B.; Rosen, H.; Schmatz, D.M.; Liberator, P.A.; Douglas, C.M. The antifungal echinocandin caspofungin acetate kills growing cells of Aspergillus fumigatus in vitro. Antimicrob. Agents Chemother. 2002, 46, 3001–3012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Xue, Y.; Li, X.; Li, J.; Zhao, Z.; Quan, C.; Gao, W.; Zu, X.; Bai, X.; Feng, S. Fungi-derived lipopeptide antibiotics developed since 2000. Peptides 2019, 113, 52–65. [Google Scholar] [CrossRef]
- Denning, D.W. Echinocandin antifungal drugs. Lancet 2003, 362, 1142–1151. [Google Scholar] [CrossRef]
- Pfizer. Anidulafungin Label Information; Pfizer: New York, NY, USA, 2009. [Google Scholar]
- Andes, D.; Diekema, D.J.; Pfaller, M.A.; Prince, R.A.; Marchillo, K.; Ashbeck, J.; Hou, J. In vivo pharmacodynamic characterization of anidulafungin in a neutropenic murine candidiasis model. Antimicrob. Agents Chemother. 2008, 52, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Astellas pharma US Inc. Micafungin Label Information; Astellas Pharma US Inc.: Deerfield, IL, USA, 2009. [Google Scholar]
- Merck and Co. Caspofungin Label, Information; Merck and Co.: Rahway, NJ, USA, 1989. [Google Scholar]
- Krishnan, B.R.; James, K.D.; Polowy, K.; Bryant, B.J.; Vaidya, A.; Smith, S.; Laudeman, C.P. CD101, a novel echinocandin with exceptional stability properties and enhanced aqueous solubility. J. Antibiot. (Tokyo) 2017, 70, 130–135. [Google Scholar] [CrossRef]
- Kofla, G.; Ruhnke, M. Pharmacology and metabolism of anidulafungin, caspofungin and micafungin in the treatment of invasive candidosis—Review of the literature. Eur. J. Med. Res. 2011, 16, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Sandison, T.; Ong, V.; Lee, J.; Thye, D. Safety and pharmacokinetics of CD101 IV, a novel echinocandin, in healthy adults. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, V.; James, K.D.; Smith, S.; Krishnan, B.R. Pharmacokinetics of the novel echinocandin CD101 in multiple animal species. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Bader, J.C.; Lakota, E.A.; Flanagan, S.; Ong, V.; Sandison, T.; Rubino, C.M.; Bhavnani, S.M.; Ambrosea, P.G. Overcoming the resistance hurdle: Pharmacokinetic-pharmacodynamic target attainment analyses for rezafungin (CD101) against Candida albicans and Candida glabrata. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Ong, V.; Hough, G.; Schlosser, M.; Bartizal, K.; Balkovec, J.M.; James, K.D.; Krishnan, B.R. Preclinical evaluation of the stability, safety, and efficacy of CD101, a novel echinocandin. Antimicrob. Agents Chemother. 2016, 60, 6872–6879. [Google Scholar] [CrossRef] [Green Version]
- Sofjan, A.K.; Mitchell, A.; Shah, D.N.; Nguyen, T.; Sim, M.; Trojcak, A.; Beyda, N.D.; Garey, K.W. Rezafungin (CD101), a next-generation echinocandin: A systematic literature review and assessment of possible place in therapy. J. Glob. Antimicrob. Resist. 2018, 14, 58–64. [Google Scholar] [CrossRef]
- Kapteyn, J.C.; Van Den Ende, H.; Klis, F.M. The contribution of cell wall proteins to the organization of the yeast cell wall. Biochim. Biophys. Acta—Gen. Subj. 1999, 1426, 373–383. [Google Scholar] [CrossRef]
- Cid, V.J.; Durán, A.; Del Rey, F.; Snyder, M.P.; Nombela, C.; Sänchez, M. Molecular basis of cell integrity and morphogenesis in Saccharomyces cerevisiae. Microbiol. Rev. 1995, 59, 345–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabib, E.; Arroyo, J. How carbohydrates sculpt cells: Chemical control of morphogenesis in the yeast cell wall. Nat. Rev. Microbiol. 2013, 11, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Huberman, L.B.; Murray, A.W. A model for cell wall dissolution in mating yeast cells: Polarized secretion and restricted diffusion of cell wall remodeling enzymes induces local dissolution. PLoS ONE 2014, 9, e109780. [Google Scholar] [CrossRef] [Green Version]
- Hochstenbach, F.; Klis, F.M.; Den Van Ende, H.; Van Donselaar, E.; Peters, P.J.; Klausner, R.D. Identification of a putative alpha-glucan synthase essential for cell wall construction and morphogenesis in fission yeast. Proc. Natl. Acad. Sci. USA 1998, 95, 9161–9166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latgé, J.P.; Beauvais, A.; Chamilos, G. The Cell Wall of the Human Fungal Pathogen Aspergillus fumigatus: Biosynthesis, Organization, Immune Response, and Virulence. Annu. Rev. Microbiol. 2017, 71, 99–116. [Google Scholar] [CrossRef]
- Gastebois, A.; Clavaud, C.; Aimanianda, V.; Latgé, J.P. Aspergillus fumigatus: Cell wall polysaccharides, their biosynthesis and organization. Future Microbiol. 2009, 4, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.A.R.; Latge, J.-P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. In The Fungal Kingdom; American Society of Microbiology: Washington, DC, USA, 2017; Volume 5, pp. 267–292. [Google Scholar]
- Iorio, E.; Torosantucci, A.; Bromuro, C.; Chiani, P.; Ferretti, A.; Giannini, M.; Cassone, A.; Podo, F. Candida albicans cell wall comprises a branched β-d-(1→6)-glucan with β-d-(1→3)-side chains. Carbohydr. Res. 2008, 343, 1050–1061. [Google Scholar] [CrossRef]
- Klis, F.M.; De Groot, P.; Hellingwerf, K. Molecular organization of the cell wall of Candida albicans. Med. Mycol. Suppl. 2001, 39, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, T.; Simenel, C.; Dubreucq, G.; Adam, O.; Delepierre, M.; Lemoine, J.; Vorgias, C.E.; Diaquin, M.; Latgé, J.P. Molecular organization of the alkali-insoluble fraction of Aspergillus fumigatus cell wall. J. Biol. Chem. 2000, 275, 27594–27607. [Google Scholar] [CrossRef] [Green Version]
- Moyrand, F.; Fontaine, T.; Janbon, G. Systematic capsule gene disruption reveals the central role of galactose metabolism on Cryptococcus neoformans virulence. Mol. Microbiol. 2007, 64, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Klutts, J.S.; Doering, T.L. Cryptococcal xylosyltransferase 1 (Cxt1p) from Cryptococcus neoformans plays a direct role in the synthesis of capsule polysaccharides. J. Biol. Chem. 2008, 283, 14327–14334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snarr, B.D.; Qureshi, S.T.; Sheppard, D.C. Immune recognition of fungal polysaccharides. J. Fungi 2017, 3, 47. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.C.; Rappleye, C.A. Flying under the radar: Histoplasma capsulatum avoidance of innate immune recognition. Semin. Cell Dev. Biol. 2019, 89, 91–98. [Google Scholar] [CrossRef]
- García-Carnero, L.C.; Martínez-Álvarez, J.A.; Salazar-García, L.M.; Lozoya-Pérez, N.E.; González-Hernández, S.E.; Tamez-Castrellón, A.K. Recognition of Fungal Components by the Host Immune System. Curr. Protein Pept. Sci. 2019, 21, 245–264. [Google Scholar] [CrossRef]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef]
- Qadota, H.; Python, C.P.; Inoue, S.B.; Arisawa, M.; Anraku, Y.; Zheng, Y.; Watanabe, T.; Levin, D.E.; Ohya, Y. Identification of yeast Rho1 p GTPase as a regulatory subunit of 1,3-β-glucan synthase. Science (80-.) 1996, 272, 279–281. [Google Scholar] [CrossRef]
- Roelants, F.M.; Leskoske, K.L.; Marshall, M.N.M.; Locke, M.N.; Thorner, J. The TORC2-dependent signaling network in the yeast Saccharomyces cerevisiae. Biomolecules 2017, 7, 66. [Google Scholar] [CrossRef]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, Y.; Mizuuchi, E.; Nogami, S.; Morishita, S.; Ohya, Y. Involvement of Rho-type GTPase in control of cell size in Saccharomyces cerevisiae. FEMS Yeast Res. 2007, 7, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bickle, M.; Delley, P.A.; Schmidt, A.; Hall, M.N. Cell wall integrity modulates RHO1 activity via the exchange factor ROM2. EMBO J. 1998, 17, 2235–2245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, M.J.; Arentshorst, M.; Roos, E.D.; Van Den Hondel, C.A.M.J.J.; Meyer, V.; Ram, A.F.J. Functional characterization of Rho GTPases in Aspergillus niger uncovers conserved and diverged roles of Rho proteins within filamentous fungi. Mol. Microbiol. 2011, 79, 1151–1167. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, O.; Tachibana, Y.; Ohya, Y.; Arisawa, M.; Watanabe, T. Cloning of the RHO1 gene from Candida albicans and its regulation of β-1,3-glucan synthesis. J. Bacteriol. 1997, 179, 7734–7741. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Rocha, A.L.; Roncero, M.I.G.; López-Ramirez, A.; Mariné, M.; Guarro, J.; Martínez-Cadena, G.; Di Pietro, A. Rho1 has distinct functions in morphogenesis, cell wall biosynthesis and virulence of Fusarium oxysporum. Cell. Microbiol. 2008, 10, 1339–1351. [Google Scholar] [CrossRef]
- Dichtl, K.; Helmschrott, C.; Dirr, F.; Wagener, J. Deciphering cell wall integrity signalling in Aspergillus fumigatus: Identification and functional characterization of cell wall stress sensors and relevant Rho GTPases. Mol. Microbiol. 2012, 83, 506–519. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Park, S.; Perlin, D.S. Correlating echinocandin MIC and kinetic inhibition of fks1 mutant glucan synthases for Candida albicans: Implications for interpretive breakpoints. Antimicrob. Agents Chemother. 2009, 53, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Effron, G.; Lee, S.; Park, S.; Cleary, J.D.; Perlin, D.S. Effect of Candida glabrata FKS1 and FKS2 mutations on echinocandin sensitivity and kinetics of 1,3-β-D-glucan synthase: Implication for the existing susceptibility breakpoint. Antimicrob. Agents Chemother. 2009, 53, 3690–3699. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Kelly, R.; Kahn, J.N.; Robles, J.; Hsu, M.J.; Register, E.; Li, W.; Vyas, V.; Fan, H.; Abruzzo, G.; et al. Specific substitutions in the echinocandin target Fks1p account for reduced susceptibility of rare laboratory and clinical Candida sp. isolates. Antimicrob. Agents Chemother. 2005, 49, 3264–3273. [Google Scholar] [CrossRef] [Green Version]
- Munro, C.A.; Selvaggini, S.; De Bruijn, I.; Walker, L.; Lenardon, M.D.; Gerssen, B.; Milne, S.; Brown, A.J.P.; Gow, N.A.R. The PKC, HOG and Ca2+ signalling pathways co-ordinately regulate chitin synthesis in Candida albicans. Mol. Microbiol. 2007, 63, 1399–1413. [Google Scholar] [CrossRef] [Green Version]
- Reinoso-Martín, C.; Schüller, C.; Schuetzer-Muehlbauer, M.; Kuchler, K. The Yeast Protein Kinase C Cell Integrity Pathway Mediates Tolerance to the Antifungal Drug Caspofungin through Activation of Slt2p Mitogen-Activated Protein Kinase Signaling. Eukaryot. Cell 2003, 2, 1200–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowen, L.E.; Lindquist, S. Cell biology: Hsp90 potentiates the rapid evolution of new traits: Drug resistance in diverse fungi. Science (80-.) 2005, 309, 2185–2189. [Google Scholar] [CrossRef] [PubMed]
- Singh-Babak, S.D.; Babak, T.; Diezmann, S.; Hill, J.A.; Xie, J.L.; Chen, Y.L.; Poutanen, S.M.; Rennie, R.P.; Heitman, J.; Cowen, L.E. Global analysis of the evolution and mechanism of echinocandin resistance in Candida glabrata. PLoS Pathog. 2012, 8, e1002718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowen, L.E. Hsp90 Orchestrates Stress Response Signaling Governing Fungal Drug Resistance. PLoS Pathog. 2009, 5, e1000471. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, R.S.; Zaas, A.K.; Betancourt-Quiroz, M.; Perfect, J.R.; Cowen, L.E. The Hsp90 Co-Chaperone Sgt1 Governs Candida albicans Morphogenesis and Drug Resistance. PLoS ONE 2012, 7, e44734. [Google Scholar] [CrossRef]
- Singh, S.D.; Robbins, N.; Zaas, A.K.; Schell, W.A.; Perfect, J.R.; Cowen, L.E. Hsp90 Governs Echinocandin Resistance in the Pathogenic Yeast Candida albicans via Calcineurin. PLoS Pathog. 2009, 5, e1000532. [Google Scholar] [CrossRef]
- Perlin, D.S. Echinocandin Resistance in Candida. Clin. Infect. Dis. 2015, 61, S612–S617. [Google Scholar] [CrossRef] [Green Version]
- Douglas, C.M.; Foor, F.; Marrinan, J.A.; Morin, N.; Nielsen, J.B.; Dahl, A.M.; Mazur, P.; Baginsky, W.; Li, W.; El-Sherbeini, M.; et al. The Saccharomyces cerevisiae FKS1 (ETG1) gene encodes an integral membrane protein which is a subunit of 1,3-β-D-glucan synthase. Proc. Natl. Acad. Sci. USA 1994, 91, 12907–12911. [Google Scholar] [CrossRef] [Green Version]
- León, M.; Sentandreu, R.; Zueco, J. A single FKS homologue in Yarrowia lipolytica is essential for viability. Yeast 2002, 19, 1003–1014. [Google Scholar] [CrossRef]
- Pereira, M.; Felipe, M.S.S.; Brígido, M.M.; Soares, C.M.A.; Azevedo, M.O. Molecular cloning and characterization of a glucan synthase gene from the human pathogenic fungus Paracoccidioides brasiliensis. Yeast 2000, 16, 451–462. [Google Scholar] [CrossRef]
- Thompson, J.R.; Douglas, C.M.; Li, W.; Jue, C.K.; Pramanik, B.; Yuan, X.; Rude, T.H.; Toffaletti, D.L.; Perfect, J.R.; Kurtz, M. A glucan synthase FKS1 homolog in Cryptococcus neoformans is single copy and encodes an essential function. J. Bacteriol. 1999, 181, 444–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maligie, M.A.; Selitrennikoff, C.P. Cryptococcus neoformans resistance to echinocandins: (1,3)β-glucan synthase activity is sensitive to echinocandins. Antimicrob. Agents Chemother. 2005, 49, 2851–2856. [Google Scholar] [CrossRef] [Green Version]
- Hori, Y.; Shibuya, K. Role of FKS gene in the susceptibility of pathogenic fungi to echinocandins. Med. Mycol. J. 2018, 59, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, E.M.F.; Garcia-Effron, G.; Park, S.; Perlin, D.S. A Ser678Pro substitution in Fks1p confers resistance to echinocandin drugs in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2007, 51, 4174–4176. [Google Scholar] [CrossRef] [Green Version]
- Mazur, P.; Morin, N.; Baginsky, W.; el-Sherbeini, M.; Clemas, J.A.; Nielsen, J.B.; Foor, F. Differential expression and function of two homologous subunits of yeast 1,3-β-D-glucan synthase. Mol. Cell. Biol. 1995, 15, 5671–5681. [Google Scholar] [CrossRef] [Green Version]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Alastruey-Izquierdo, A.; Healey, K.R.; Johnson, M.E.; Perlin, D.S.; Edlind, T.D. Fks1 and Fks2 are functionally redundant but differentially regulated in Candida glabrata: Implications for echinocandin resistance. Antimicrob. Agents Chemother. 2012, 56, 6304–6309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef]
- Patterson, T.F.; Thompson, G.R.; Denning, D.W.; Fishman, J.A.; Hadley, S.; Herbrecht, R.; Kontoyiannis, D.P.; Marr, K.A.; Morrison, V.A.; Nguyen, M.H.; et al. Practice guidelines for the diagnosis and management of aspergillosis: 2016 update by the infectious diseases society of America. Clin. Infect. Dis. 2016, 63, e1–e60. [Google Scholar] [CrossRef]
- Cushion, M.T.; Linke, M.J.; Ashbaugh, A.; Sesterhenn, T.; Collins, M.S.; Lynch, K.; Brubaker, R.; Walzer, P.D. Echinocandin treatment of Pneumocystis pneumonia in rodent models depletes cysts leaving trophic burdens that cannot transmit the infection. PLoS ONE 2010, 5, e8524. [Google Scholar] [CrossRef]
- Cidara Therapeutics (Sponsor) CD101 Compared to Caspofungin Followed by Oral Step Down in Subjects with Candidemia and/or Invasive Candidiasis-Bridging Extension (STRIVE). Available online: https://clinicaltrials.gov/ct2/show/NCT02734862?term=cd101&draw=2&rank=3 (accessed on 29 October 2020).
- Cidara Therapeutics (Sponsor) RADIANT: CD101 vs Standard of Care in Subjects with Acute Vaginal Yeast Infections. Available online: https://clinicaltrials.gov/ct2/show/NCT02733432?term=cd101&draw=2&rank=4 (accessed on 29 October 2020).
- Cidara Therapeutics (Sponsor) Study of Rezafungin Compared to Standard Antimicrobial Regimen for Prevention of Invasive Fungal Diseases in Adults Undergoing Allogeneic Blood and Marrow Transplantation (ReSPECT). Available online: https://clinicaltrials.gov/ct2/show/NCT04368559?term=cd101&draw=2&rank=10 (accessed on 29 October 2020).
- Cassone, A.; Mason, R.E.; Kerridge, D. Lysis of growing yeast-form cells of Candida albicans by echinocandin: A cytological study. Med. Mycol. 1981, 19, 97–110. [Google Scholar] [CrossRef]
- Gil-Alonso, S.; Jauregizar, N.; Cantón, E.; Eraso, E.; Quindós, G. Comparison of the in vitro activity of echinocandins against Candida albicans, Candida dubliniensis, and Candida africana by time-kill curves. Diagn. Microbiol. Infect. Dis. 2015, 82, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Ernst, E.J.; Klepser, M.E.; Ernst, M.E.; Messer, S.A.; Pfaller, M.A. In vitro pharmacodynamic properties of MK-0991 determined by time-kill methods. Diagn. Microbiol. Infect. Dis. 1999, 33, 75–80. [Google Scholar] [CrossRef]
- Garcia-Effron, G.; Katiyar, S.K.; Park, S.; Edlind, T.D.; Perlin, D.S. A naturally occurring proline-to-alanine amino acid change in Fks1p in Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis accounts for reduced echinocandin susceptibility. Antimicrob. Agents Chemother. 2008, 52, 2305–2312. [Google Scholar] [CrossRef] [Green Version]
- Dudiuk, C.; MacEdo, D.; Leonardelli, F.; Theill, L.; Cabeza, M.S.; Gamarra, S.; Garcia-Effron, G. Molecular confirmation of the relationship between Candida guilliermondii Fks1p naturally occurring amino acid substitutions and its intrinsic reduced echinocandin susceptibility. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Cantón, E.; Pemán, J.; Sastre, M.; Romero, M.; Espinel-Ingroff, A. Killing kinetics of caspofungin, micafungin, and amphotericin B against Candida guilliermondii. Antimicrob. Agents Chemother. 2006, 50, 2829–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Alonso, S.; Quindós, G.; Cantón, E.; Eraso, E.; Jauregizar, N. Killing kinetics of anidulafungin, caspofungin and micafungin against Candida parapsilosis species complex: Evaluation of the fungicidal activity. Rev. Iberoam. Micol. 2019, 36, 24–29. [Google Scholar] [CrossRef]
- Dudiuk, C.; Berrio, I.; Leonardelli, F.; Morales-Lopez, S.; Theill, L.; Macedo, D.; Yesid-Rodriguez, J.; Salcedo, S.; Marin, A.; Gamarra, S.; et al. Antifungal activity and killing kinetics of anidulafungin, caspofungin and amphotericin B against Candida auris. J. Antimicrob. Chemother. 2019, 74, 2295–2302. [Google Scholar] [CrossRef] [PubMed]
- Utsugi, T.; Minemura, M.; Hirata, A.; Abe, M.; Watanabe, D.; Ohya, Y. Movement of yeast 1,3-β-glucan synthase is essential for uniform cell wall synthesis. Genes Cells 2002, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingham, C.J.; Schneeberger, P.M. Microcolony imaging of Aspergillus fumigatus treated with echinocandins reveals both fungistatic and fungicidal activities. PLoS ONE 2012, 7, e35478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Strandards Institute CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; CLSI Standard M38; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Tang, J.; Parr, T.R. W-1 solubilization and kinetics of inhibition by cilofungin of Candida albicans (1,3)-β-D-glucan synthase. Antimicrob. Agents Chemother. 1991, 35, 99–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petraitiene, R.; Petraitis, V.; Groll, A.H.; Candelario, M.; Sein, T.; Bell, A.; Lyman, C.A.; McMillian, C.L.; Bacher, J.; Walsh, T.J. Antifungal activity of LY303366, a novel echinocandin B, in experimental disseminated candidiasis in rabbits. Antimicrob. Agents Chemother. 1999, 43, 2148–2155. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.E.; Katiyar, S.K.; Edlind, T.D. New Fks hot spot for acquired echinocandin resistance in Saccharomyces cerevisiae and its contribution to intrinsic resistance of Scedosporium species. Antimicrob. Agents Chemother. 2011, 55, 3774–3781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schimoler-O’Rourke, R.; Renault, S.; Mo, W.; Selitrennikoff, C.P. Neurospora crassa FKS protein binds to the (1,3)β-glucan synthase substrate, UDP-glucose. Curr. Microbiol. 2003, 46, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.K.; Nguyen, M.H.; Press, E.G.; Kwa, A.L.; Cheng, S.; Du, C.; Clancy, C.J. The presence of an FKS mutation rather than MIC is an independent risk factor for failure of echinocandin therapy among patients with invasive candidiasis due to Candida glabrata. Antimicrob. Agents Chemother. 2012, 56, 4862–4869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.E.; Edlind, T.D. Topological and mutational analysis of Saccharomyces cerevisiae Fks1. Eukaryot. Cell 2012, 11, 952–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengstag, C.; Stirling, C.; Schekman, R.; Rine, J. Genetic and biochemical evaluation of eucaryotic membrane protein topology: Multiple transmembrane domains of Saccharomyces cerevisiae 3-hydroxy-3-methylglutaryl coenzyme A reductase. Mol. Cell. Biol. 1990, 10, 672–680. [Google Scholar] [CrossRef] [Green Version]
- Boeck, L.D.; Fukuda, D.S.; Abbott, B.J.; Debono, M. Deacylation of echinocandin B by actinoplanes utahensis. J. Antibiot. (Tokyo) 1989, 42, 382–388. [Google Scholar] [CrossRef]
- Zhao, Y.; Perez, W.B.; Jiménez-Ortigosa, C.; Hough, G.; Locke, J.B.; Ong, V.; Bartizal, K.; Perlin, D.S. CD101: A novel long-acting echinocandin. Cell. Microbiol. 2016, 18, 1308–1316. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Locke, J.B.; Daruwala, P.; Bartizal, K. Rezafungin (CD101) demonstrates potent in vitro activity against Aspergillus, including azole-resistant Aspergillus fumigatus isolates and cryptic species. J. Antimicrob. Chemother. 2018, 73, 3063–3067. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Carvalhaes, C.; Messer, S.A.; Rhomberg, P.R.; Castanheira, M. Activity of a long-acting echinocandin, rezafungin, and comparator antifungal agents tested against contemporary invasive fungal isolates (SENTRY program, 2016 to 2018). Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Messer, S.A.; Rhomberg, P.R.; Castanheira, M. CD101, a long-acting echinocandin, and comparator antifungal agents tested against a global collection of invasive fungal isolates in the SENTRY 2015 Antifungal Surveillance Program. Int. J. Antimicrob. Agents 2017, 50, 352–358. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Messer, S.A.; Rhomberg, P.R.; Jones, R.N.; Castanheira, M. Activity of a long-acting echinocandin, CD101, determined using CLSI and EUCAST reference methods, against Candida and Aspergillus spp., including echinocandin- and azole-resistant isolates. J. Antimicrob. Chemother. 2016, 71, 2868–2873. [Google Scholar] [CrossRef] [Green Version]
- Berkow, E.L.; Lockhart, S.R. Activity of CD101, a long-acting echinocandin, against clinical isolates of Candida auris. Diagn. Microbiol. Infect. Dis. 2018, 90, 196–197. [Google Scholar] [CrossRef]
- Hall, D.; Bonifas, R.; Stapert, L.; Thwaites, M.; Shinabarger, D.L.; Pillar, C.M. In vitro potency and fungicidal activity of CD101, a novel echinocandin, against recent clinical isolates of Candida spp. Diagn. Microbiol. Infect. Dis. 2017, 89, 205–211. [Google Scholar] [CrossRef]
- Tóth, Z.; Forgács, L.; Locke, J.B.; Kardos, G.; Nagy, F.; Kovács, R.; Szekely, A.; Borman, A.M.; Majoros, L. In vitro activity of rezafungin against common and rare Candida species and Saccharomyces cerevisiae. J. Antimicrob. Chemother. 2019, 74, 3505–3510. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Jørgensen, K.M.; Hare, R.K.; Cuenca-Estrella, M.; Zaragoza, O. EUCAST reference testing of rezafungin susceptibility and impact of choice of plastic plates. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Messer, S.A.; Rhomberg, P.R.; Castanheira, M. Activity of a long-acting echinocandin (CD101) and seven comparator antifungal agents tested against a global collection of contemporary invasive fungal isolates in the SENTRY 2014 Antifungal Surveillance Program. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boikov, D.A.; Locke, J.B.; James, K.D.; Bartizal, K.; Sobel, J.D. In vitro activity of the novel echinocandin CD101 at pH 7 and 4 against Candida spp. isolates from patients with vulvovaginal candidiasis. J. Antimicrob. Chemother. 2017, 72, 1355–1358. [Google Scholar] [CrossRef] [Green Version]
- Chandra, J.; Ghannoum, M.A. CD101, A novel echinocandin, possesses potent antibiofilm activity against early and mature Candida albicans biofilms. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cota, J.M.; Grabinski, J.L.; Talbert, R.L.; Burgess, D.S.; Rogers, P.D.; Edlind, T.D.; Wiederhold, N.P. Increases in SLT2 expression and chitin content are associated with incomplete killing of Candida glabrata by caspofungin. Antimicrob. Agents Chemother. 2008, 52, 1144–1146. [Google Scholar] [CrossRef] [Green Version]
- Gow, N.A.R.; Netea, M.G.; Munro, C.A.; Ferwerda, G.; Bates, S.; Mora-Montes, H.M.; Walker, L.; Jansen, T.; Jacobs, L.; Tsoni, V.; et al. Immune recognition of Candida albicans β-glucan by dectin-1. J. Infect. Dis. 2007, 196, 1565–1571. [Google Scholar] [CrossRef] [Green Version]
- Plaine, A.; Walker, L.; Da Costa, G.; Mora-Montes, H.M.; McKinnon, A.; Gow, N.A.R.; Gaillardin, C.; Munro, C.A.; Richard, M.L. Functional analysis of Candida albicans GPI-anchored proteins: Roles in cell wall integrity and caspofungin sensitivity. Fungal Genet. Biol. 2008, 45, 1404–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.K.; MacCallum, D.M.; Jacobsen, M.D.; Walker, L.A.; Odds, F.C.; Gow, N.A.R.; Munro, C.A. Elevated cell wall chitin in Candida albicans confers echinocandin resistance in vivo. Antimicrob. Agents Chemother. 2012, 56, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Stevens, D.A.; Ichinomiya, M.; Koshi, Y.; Horiuchi, H. Escape of Candida from caspofungin inhibition at concentrations above the MIC (paradoxical effect) accomplished by increased cell wall chitin; evidence for β-1,6-glucan synthesis inhibition by caspofungin. Antimicrob. Agents Chemother. 2006, 50, 3160–3161. [Google Scholar] [CrossRef] [Green Version]
- Clemons, K.V.; Espiritu, M.; Parmar, R.; Stevens, D.A. Assessment of the paradoxical effect of caspofungin in therapy of candidiasis. Antimicrob. Agents Chemother. 2006, 50, 1293–1297. [Google Scholar] [CrossRef] [Green Version]
- Stevens, D.A.; Espiritu, M.; Parmar, R. Paradoxical effect of caspofungin: Reduced activity against Candida albicans at high drug concentrations. Antimicrob. Agents Chemother. 2004, 48, 3407–3411. [Google Scholar] [CrossRef] [Green Version]
- Healey, K.R.; Katiyar, S.K.; Raj, S.; Edlind, T.D. CRS-MIS in Candida glabrata: Sphingolipids modulate echinocandin-Fks interaction. Mol. Microbiol. 2012, 86, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Harrison, B.D.; Hashemi, J.; Bibi, M.; Pulver, R.; Bavli, D.; Nahmias, Y.; Wellington, M.; Sapiro, G.; Berman, J. A tetraploid intermediate precedes aneuploid formation in yeasts exposed to fluconazole. PLoS Biol. 2014, 12, e1001815. [Google Scholar] [CrossRef]
- Sheltzer, J.M.; Blank, H.M.; Pfau, S.J.; Tange, Y.; George, B.M.; Humpton, T.J.; Brito, I.L.; Hiraoka, Y.; Niwa, O.; Amon, A. Aneuploidy drives genomic instability in yeast. Science (80-.) 2011, 333, 1026–1030. [Google Scholar] [CrossRef] [Green Version]
- Cowen, L.E.; Steinbach, W.J. Stress, drugs, and evolution: The role of cellular signaling in fungal drug resistance. Eukaryot. Cell 2008, 7, 747–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlin, D.S. Mechanisms of echinocandin antifungal drug resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 1–11. [Google Scholar] [CrossRef]
- Arendrup, M.C.; Perlin, D.S. Echinocandin resistance: An emerging clinical problem? Curr. Opin. Infect. Dis. 2014, 27, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, J.N.; Garcia-Effron, G.; Hsu, M.J.; Park, S.; Marr, K.A.; Perlin, D.S. Acquired echinocandin resistance in a Candida krusei isolate due to modification of glucan synthase. Antimicrob. Agents Chemother. 2007, 51, 1876–1878. [Google Scholar] [CrossRef] [Green Version]
- Cleary, J.D.; Garcia-Effron, G.; Chapman, S.W.; Perlin, D.S. Reduced Candida glabrata susceptibility secondary to an FKS1 mutation developed during candidemia treatment. Antimicrob. Agents Chemother. 2008, 52, 2263–2265. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Effron, G.; Chua, D.J.; Tomada, J.R.; DiPersio, J.; Perlin, D.S.; Ghannoum, M.; Bonilla, H. Novel FKS mutations associated with echinocandin resistance in Candida species. Antimicrob. Agents Chemother. 2010, 54, 2225–2227. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Effron, G.; Kontoyiannis, D.P.; Lewis, R.E.; Perlin, D.S. Caspofungin-resistant Candida tropicalis strains causing breakthrough fungemia in patients at high risk for hematologic malignancies. Antimicrob. Agents Chemother. 2008, 52, 4181–4183. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Prakash, A.; Sharma, C.; Kordalewska, M.; Kumar, A.; Sarma, S.; Tarai, B.; Singh, A.; Upadhyaya, G.; Upadhyay, S.; et al. A multicentre study of antifungal susceptibility patterns among 350 Candida auris isolates (2009-17) in India: Role of the ERG11 and FKS1 genes in azole and echinocandin resistance. J. Antimicrob. Chemother. 2018, 73, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Ben-Ami, R.; Garcia-Effron, G.; Lewis, R.E.; Gamarra, S.; Leventakos, K.; Perlin, D.S.; Kontoyiannis, D.P. Fitness and virulence costs of Candida albicans FKS1 hot spot mutations associated with echinocandin resistance. J. Infect. Dis. 2011, 204, 626–635. [Google Scholar] [CrossRef] [Green Version]
- Sen, T.Z.; Jernigan, R.L.; Garnier, J.; Kloczkowski, A. GOR V server for protein secondary structure prediction. Bioinformatics 2005, 21, 2787–2788. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, M.; Edmundson, A.B. Use of helical wheels to represent the structures of proteins and to identify segments with helical potential. Biophys. J. 1967, 7, 121–135. [Google Scholar] [CrossRef] [Green Version]
- Kohler, S.; Wheat, L.J.; Connolly, P.; Schnizlein-Bick, C.; Durkin, M.; Smedema, M.; Goldberg, J.; Brizendine, E. Comparison of the echinocandin caspofungin with amphotericin b for treatment of Histoplasmosis following pulmonary challenge in a murine model. Antimicrob. Agents Chemother. 2000, 44, 1850–1854. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, S.K.; Edlind, T.D. Role for Fks1 in the intrinsic echinocandin resistance of Fusarium solani as evidenced by hybrid expression in Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 2009, 53, 1772–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duin, D.; Casadevall, A.; Nosanchuk, J.D. Melanization of Cryptococcus neoformans and Histoplasma capsulatum reduces their susceptibilities to amphotericin B and caspofungin. Antimicrob. Agents Chemother. 2002, 46, 3394–3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Liao, G.; Baker, G.M.; Wang, Y.; Lau, R.; Paderu, P.; Perlin, D.S.; Xue, C. Lipid flippase subunit Cdc50 mediates drug resistance and virulence in Cryptococcus neoformans. MBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Hamal, P.; Guinea, J.; Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). EUCAST Definitive Document E.DEF 7.3.2. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts. April 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Files/EUCAST_E_Def_7.3.2_Yeast_testing_definitive_revised_2020.pdf (accessed on 29 October 2020).
- Clinical and Laboratory Strandards Institute. Reference Method for Broth Dilution Antifungal Susceptiblity Testing of Yeasts, 4th ed.; CLSI Document M27; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- Clinical and Laboratory Strandards Institute. Epidemiological Cutoff Values for Antifungal Susceptibility Testing, 2nd ed.; CLSI Supplement M59; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Reboli, A.C.; Rotstein, C.; Pappas, P.G.; Chapman, S.W.; Kett, D.H.; Kumar, D.; Betts, R.; Wible, M.; Goldstein, B.P.; Schranz, J.; et al. Anidulafungin versus fluconazole for invasive candidiasis. N. Engl. J. Med. 2007, 356, 2472–2482. [Google Scholar] [CrossRef] [Green Version]
- Gil-Alonso, S.; Jauregizar, N.; Cantón, E.; Eraso, E.; Quindósa, G. In vitro fungicidal activities of anidulafungin, caspofungin, and micafungin against Candida glabrata, Candida bracarensis, and Candida nivariensis evaluated by time-kill studies. Antimicrob. Agents Chemother. 2015, 59, 3615–3618. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Prakash, A.; Meletiadis, J.; Sharma, C.; Chowdhary, A. Comparison of EUCAST and CLSI reference microdilution mics of eight antifungal compounds for Candida auris and associated tentative epidemiological cutoff values. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Centers for Diseases Control and Prevention, C. Antifungal Susceptibility Testing and Interpretation|Candida auris | Fungal Diseases | CDC. Available online: https://www.cdc.gov/fungal/candida-auris/c-auris-antifungal.html (accessed on 23 September 2020).
- Kordalewska, M.; Lee, A.; Zhao, Y.; Perlin, D.S. Detection of Candida auris antifungal drug resistance markers directly from clinical skin swabs. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Hou, X.; Lee, A.; Jiménez-Ortigosa, C.; Kordalewska, M.; Perlin, D.S.; Zhao, Y. Rapid detection of ERG11-associated azole resistance and FKS-associated echinocandin resistance in Candida auris. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Locke, J.B.; Almaguer, A.L.; Zuill, D.E.; Bartizal, K. Characterization of in vitro resistance development to the novel echinocandin CD101 in Candida species. Antimicrob. Agents Chemother. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balashov, S.V.; Park, S.; Perlin, D.S. Assessing resistance to the echinocandin antifungal drug caspofungin in Candida albicans by profiling mutations in FKS1. Antimicrob. Agents Chemother. 2006, 50, 2058–2063. [Google Scholar] [CrossRef] [Green Version]
- Shields, R.K.; Kline, E.G.; Healey, K.R.; Kordalewska, M.; Perlin, D.S.; Hong Nguyen, M.; Clancy, C.J. Spontaneous mutational frequency and fks mutation rates vary by echinocandin agent against Candida glabrata. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jiménez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing echinocandin resistance in Candida glabrata: Clinical failure correlates with presence of FKS mutations and elevated minimum inhibitory concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [Green Version]
- Zimbeck, A.J.; Iqbal, N.; Ahlquist, A.M.; Farley, M.M.; Harrison, L.H.; Chiller, T.; Lockhart, S.R. FKS mutations and elevated echinocandin MIC values among Candida glabrata isolates from U.S. population-based surveillance. Antimicrob. Agents Chemother. 2010, 54, 5042–5047. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Chemical structure of FDA-approved echinocandins (anidulafungin, caspofungin, and micafungin) and rezafungin. In black, the common chemical structure of the four molecules. In red and green, the caspofungin/micafungin and anidulafungin/rezafungin common structures, respectively. In blue, exclusive micafungin chemical structures. In light blue, the choline aminal ether of rezafungin that improves it stability (degradation less likely). Degradation of anidulafungin and rezafungin, modified from [16].
Figure 1.
Chemical structure of FDA-approved echinocandins (anidulafungin, caspofungin, and micafungin) and rezafungin. In black, the common chemical structure of the four molecules. In red and green, the caspofungin/micafungin and anidulafungin/rezafungin common structures, respectively. In blue, exclusive micafungin chemical structures. In light blue, the choline aminal ether of rezafungin that improves it stability (degradation less likely). Degradation of anidulafungin and rezafungin, modified from [16].

Figure 2.
Schematic representation of fungal cell walls of the main fungal human pathogens following the hypothesis proposed by Gow et al. [32]. Alkali-insoluble core components: chithin and branched β-(1,3) and β-(1,6) glucans (black box).
Figure 2.
Schematic representation of fungal cell walls of the main fungal human pathogens following the hypothesis proposed by Gow et al. [32]. Alkali-insoluble core components: chithin and branched β-(1,3) and β-(1,6) glucans (black box).

Figure 3.
((A) above) Schematic representation (not in scale) of a portion of the Candida albicans Fks1p membrane topology on the basis of the Saccharomyces cerevisiae topology model (98). Black bars represent the transmembrane helices (TMH 4 to THM 9). Hot spot regions are colored in red (hot spot 1), in blue (hot spot 2), and in green (hot spot 3). Hot spot relative positions are not indented to represent the real position or the so-called echinocandin binding pocket. Putative glycosyltranferase domain (catalytic domain and substrate binding sites) is represented by a gray loop (I717-H1298). ((A) below) Helical wheel representation showing the putative position of the amino acid of hot spot 1 (HS1) and hot spot 2 (HS2) of C. albicans Fks1p arranged in a α-helix seen from above [94]. The residues where mutations confer strong echinocandin resistance phenotype are represented in red; in orange, intermediate to low resistance phenotype [95]; and in purple, the residues that show low phenotype and are implicated in the so-called intrinsic reduced echinocandin susceptibility (IRES) phenotype in C. parapsilosis complex (P649A) and C. guilliermondii (L642M) [79,80]. The residues not linked with echinocandin resistance are represented in green. ((B), above) Alignment of the hot spot regions of Candida spp. The residues linked with high level echinocandin resistance are represented in red; in orange, the residues that conferred low to intermediate resistance phenotype [96]; and in purple, the residues confirmed molecularly that are related with IRES phenotype [79,80]. In bold and underlined are the residues where spontaneous mutations were produced after in vitro exposure to high concentrations of rezafungin [97]. ((B), below) Simplified representation of the relative frequency of appearance of each substitutions in clinical isolates together with its main characteristics (highest the size, the highest Minimal Inhibitory Concentration (MIC)/ and half enzyme maximal inhibitory concentration 50% (IC50), Vmax, and better fitness) [51,52,60,98,99].
Figure 3.
((A) above) Schematic representation (not in scale) of a portion of the Candida albicans Fks1p membrane topology on the basis of the Saccharomyces cerevisiae topology model (98). Black bars represent the transmembrane helices (TMH 4 to THM 9). Hot spot regions are colored in red (hot spot 1), in blue (hot spot 2), and in green (hot spot 3). Hot spot relative positions are not indented to represent the real position or the so-called echinocandin binding pocket. Putative glycosyltranferase domain (catalytic domain and substrate binding sites) is represented by a gray loop (I717-H1298). ((A) below) Helical wheel representation showing the putative position of the amino acid of hot spot 1 (HS1) and hot spot 2 (HS2) of C. albicans Fks1p arranged in a α-helix seen from above [94]. The residues where mutations confer strong echinocandin resistance phenotype are represented in red; in orange, intermediate to low resistance phenotype [95]; and in purple, the residues that show low phenotype and are implicated in the so-called intrinsic reduced echinocandin susceptibility (IRES) phenotype in C. parapsilosis complex (P649A) and C. guilliermondii (L642M) [79,80]. The residues not linked with echinocandin resistance are represented in green. ((B), above) Alignment of the hot spot regions of Candida spp. The residues linked with high level echinocandin resistance are represented in red; in orange, the residues that conferred low to intermediate resistance phenotype [96]; and in purple, the residues confirmed molecularly that are related with IRES phenotype [79,80]. In bold and underlined are the residues where spontaneous mutations were produced after in vitro exposure to high concentrations of rezafungin [97]. ((B), below) Simplified representation of the relative frequency of appearance of each substitutions in clinical isolates together with its main characteristics (highest the size, the highest Minimal Inhibitory Concentration (MIC)/ and half enzyme maximal inhibitory concentration 50% (IC50), Vmax, and better fitness) [51,52,60,98,99].

Figure 4.
Clustal alignment of Fksps of different fungal pathogens. HS1: hot spot 1 region; HS2: hot spot 2 region; HS3: hot spot 3 region. Dots represent the same amino acid as C. albicans in its equivalent position.
Figure 4.
Clustal alignment of Fksps of different fungal pathogens. HS1: hot spot 1 region; HS2: hot spot 2 region; HS3: hot spot 3 region. Dots represent the same amino acid as C. albicans in its equivalent position.


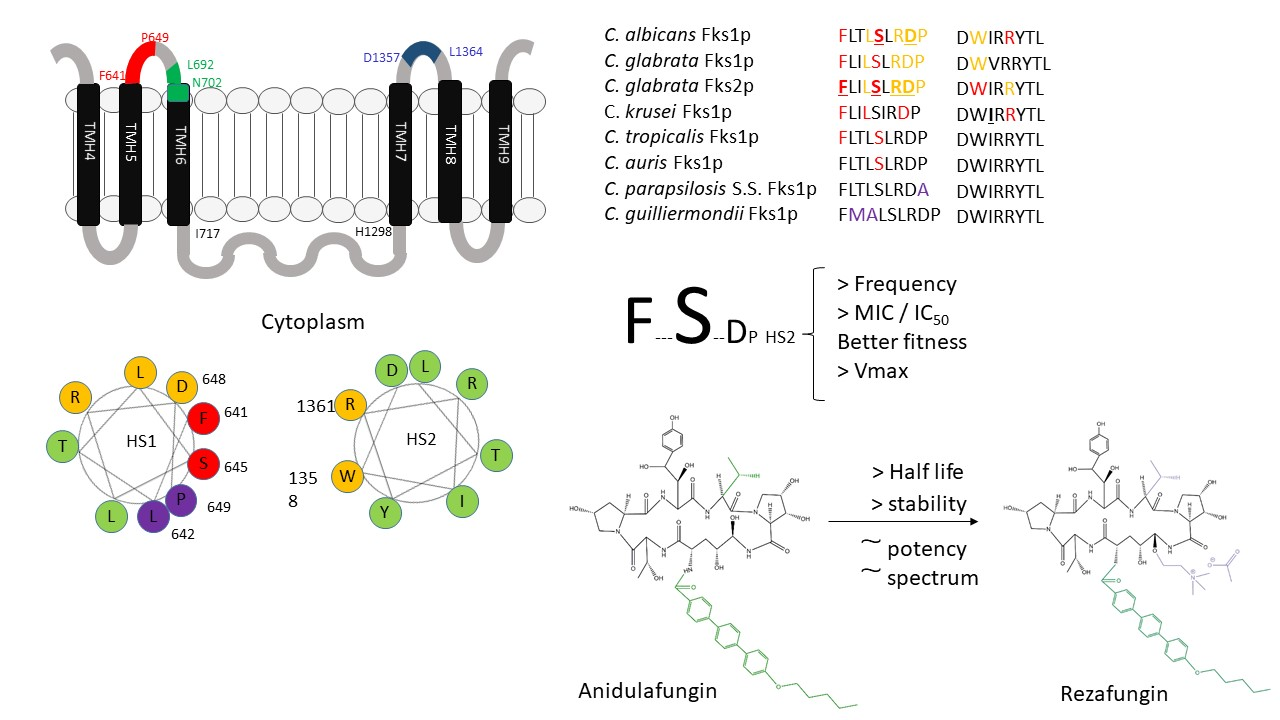{kind=link}
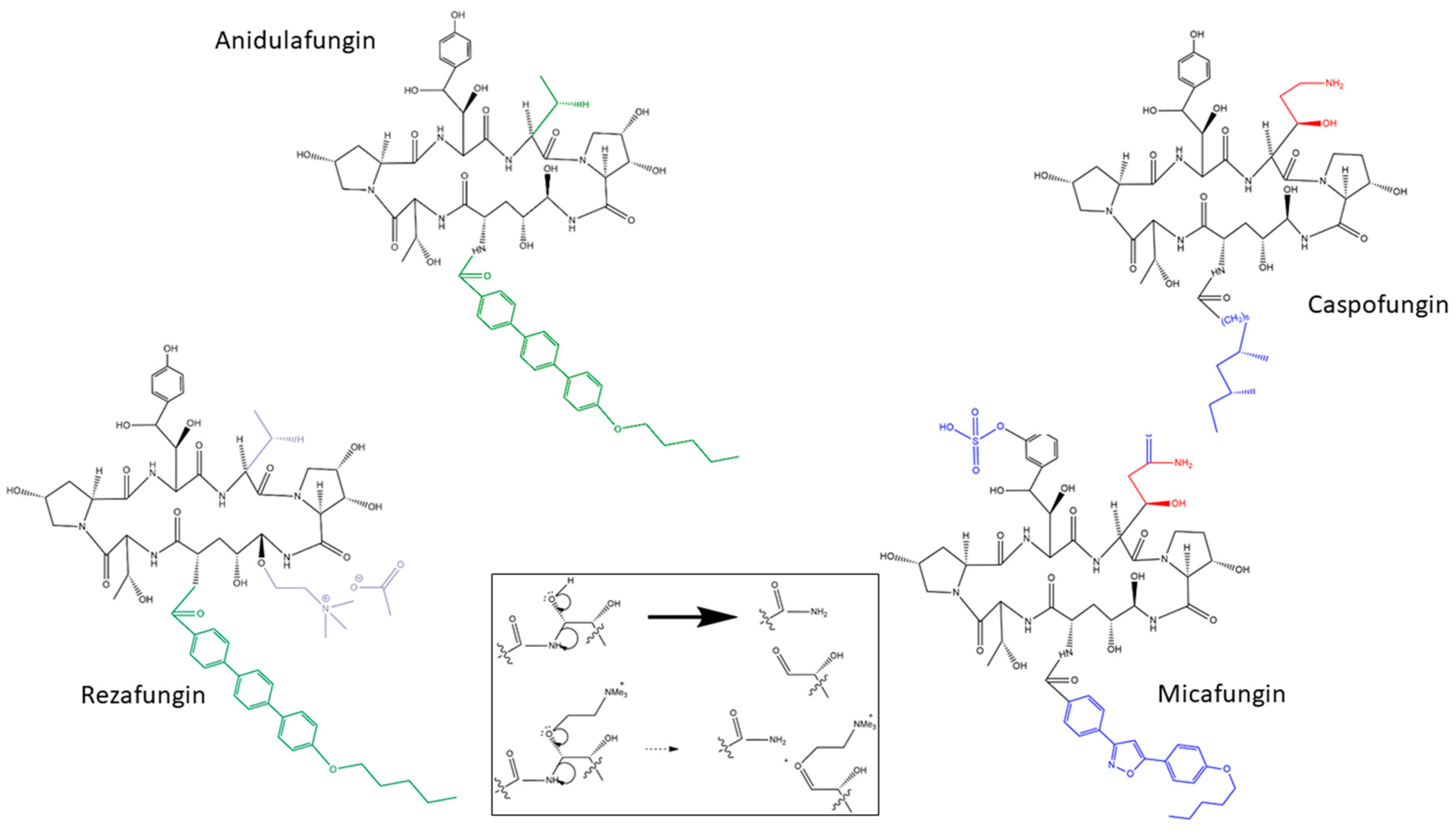{kind=link}
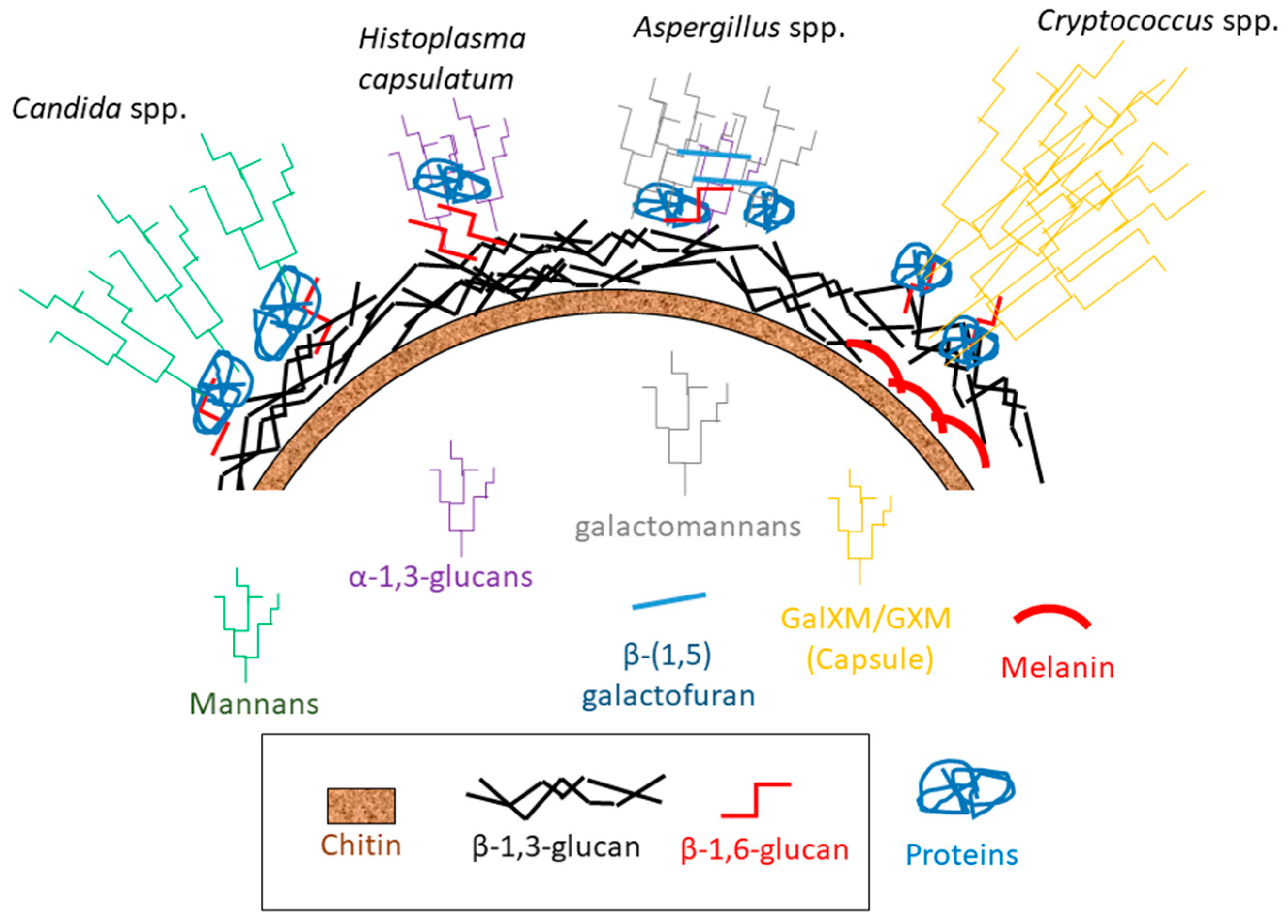{kind=link}
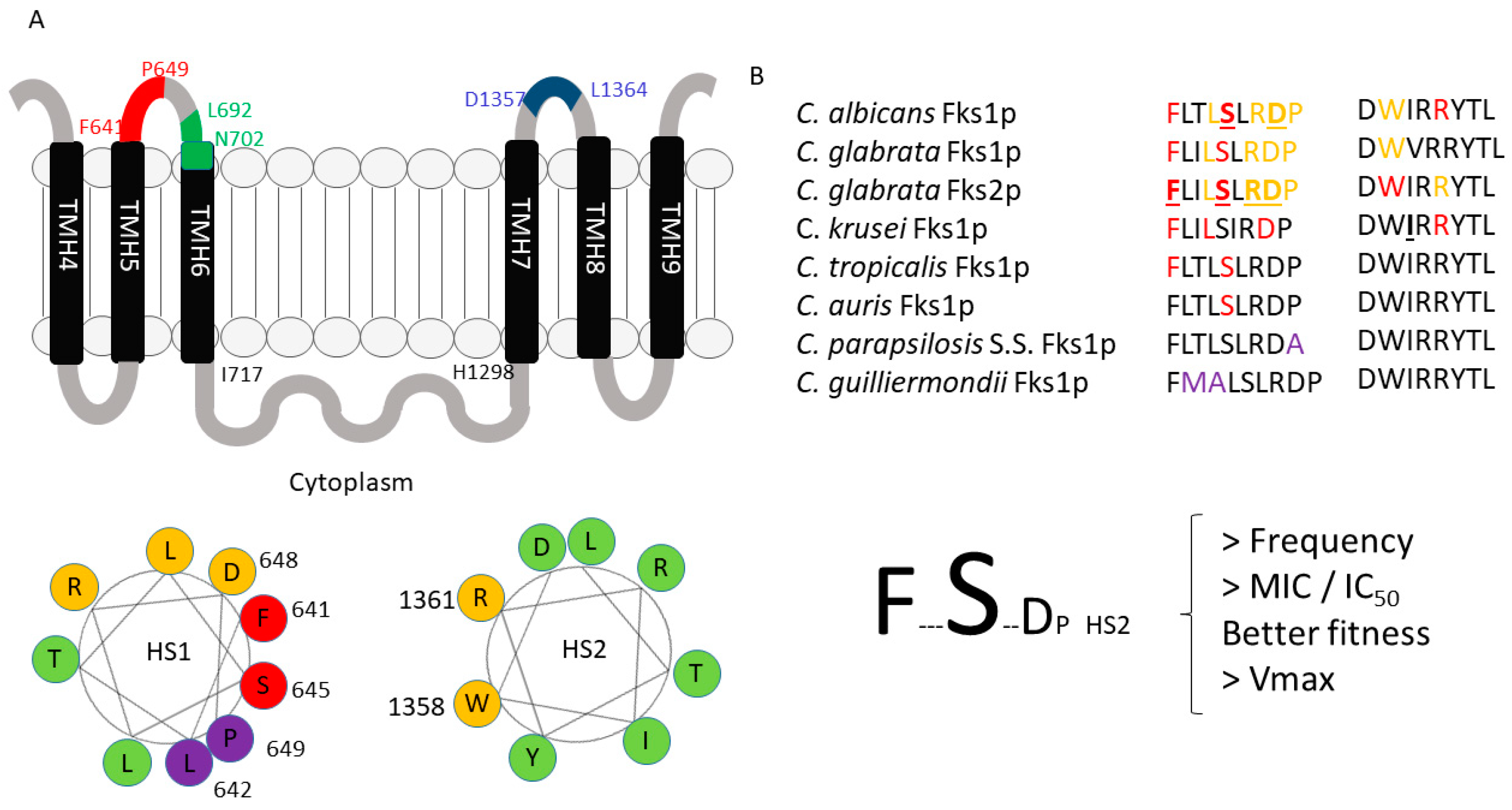{kind=link}
{kind=link}
Table 1.
Rezafungin and FDA-approved echinocandin MIC values for Candida spp. with wild type FKS genes determined by Clinical and Laboratory Standards Institute (CLSI) and/or European Committee on Antimicrobial Susceptibility Testing (EUCAST) microdilution reference methods.
Table 1.
Rezafungin and FDA-approved echinocandin MIC values for Candida spp. with wild type FKS genes determined by Clinical and Laboratory Standards Institute (CLSI) and/or European Committee on Antimicrobial Susceptibility Testing (EUCAST) microdilution reference methods.
| Candida spp. | n | RZF a | ANF a | CSF a | MCF a | References | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| MIC50 | MIC90 | MIC50 | MIC90 | MIC50 | MIC90 | MIC50 | MIC90 | |||
| C. albicans | 2612 | 0.022 | 0.050 | 0.012 | 0.027 | 0.053 | 0.069 | 0.022 | 0.021 | [100,101,102,103,104,105,106,107,109,110,111] |
| C. glabrata | 1541 | 0.044 | 0.085 | 0.045 | 0.085 | 0.080 | 0.140 | 0.027 | 0.030 | [100,101,102,103,104,105,106,107,109,110,111] |
| C. krusei | 773 | 0.033 | 0.078 | 0.045 | 0.085 | 0.280 | 0.248 | 0.108 | 0.153 | [100,101,102,103,104,105,106,107,109,110,111] |
| C. parapsilosis sensu stricto | 1156 | 1.260 | 2.000 | 1.219 | 2.245 | 0.435 | 0.758 | 1.122 | 1.414 | [100,101,102,103,104,105,106,107,109,110,111] |
| C. tropicalis | 959 | 0.030 | 0.072 | 0.012 | 0.034 | 0.046 | 0.092 | 0.030 | 0.050 | [100,101,102,103,104,105,106,107,109,110,111] |
| C. dubliniensis | 207 | 0.060 | 1.360 | 0.034 | 0.270 | 0.036 | 0.370 | 0.030 | 0.105 | [100,101,102,103,104,105,107,109,110] |
| C. auris | 237 | 0.153 | 0.500 | 0.391 | 0.250 | 0.707 | 1.000 | 0.630 | 0.500 | [105,107,109,110] |
| C. lusitaniae | 66 | 0.120 | 0.250 | 0.042 | 0.060 | 0.500 | 1.000 | 0.038 | 0.250 | [107,110] |
| C. kefyr | 52 | 0.06 | 0.12 | 0.03 | 0.06 | 0.25 | 0.50 | 0.06 | 0.12 | [107] |
| C. guilliermondii | 27 | 1.00 | 1.00 | 1.00 | 2.00 | 0.50 | 1.00 | 1.00 | 2.00 | [107] |
| C. orthopsilosis | 25 | 0.500 | 1.000 | 0.707 | 1.000 | 0.354 | 0.707 | 0.500 | 1.000 | [107,109] |
| C. metapsilosis | 15 | 0.50 | 0.50 | 0.25 | 0.50 | 0.25 | 0.50 | 0.25 | 0.50 | [107] |
| C. fabianii | 15 | 0.06 | 0.12 | 0.06 | 0.12 | 1.00 | 1.00 | 0.06 | 0.12 | [107] |
| C. insconspicua | 41 | 0.06 | 0.06 | 0.008 | 0.015 | 0.25 | 0.50 | 0.03 | 0.06 | [107] |
| C. sojae | 10 | 0.06 | 0.06 | 0.015 | 0.03 | 0.25 | 0.50 | 0.03 | 0.06 | [107] |
| C. lipolytica | 10 | 0.06 | 0.06 | 0.06 | 0.12 | 0.25 | 0.50 | 0.25 | 1.00 | [107] |
| C. pulcherrima | 10 | 0.03 | 0.06 | 0.015 | 0.06 | 0.50 | 1.00 | 0.06 | 0.25 | [107] |
RZF: rezafungin, ANF: anidulafungin, CSF: caspofungin, MCF: micafungin. a Geometric means of the data published in the cited references expressed in µg/mL. ND: no data available.
Table 2.
Rezafungin and FDA-approved echinocandin MIC values for Candida spp. with mutant FKS genes determined by CLSI microdilution reference methods.
Table 2.
Rezafungin and FDA-approved echinocandin MIC values for Candida spp. with mutant FKS genes determined by CLSI microdilution reference methods.
| Candida spp. FKS Mutants | n | RZF a | ANF a | CSF a | MCF a | References | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| MIC50 | MIC90 | MIC50 | MIC90 | MIC50 | MIC90 | MIC50 | MIC90 | |||
| C. albicans | 20 | 0.71 | 1.00 | 0.50 | 1.00 | 0.50 | 1.00 | 1.00 | ND | [100,104] |
| C. glabrata | 21 | 0.50 | 1.00 | 0.25 | 1.00 | 0.50 | 1.00 | 1.00 | ND | [100,104] |
| C. krusei | 6 | 0.35 | 1.00 | 0.50 | 2.00 | 1.00 | 16.00 | 1.00 | ND | [100,104] |
| C. tropicalis | 9 | 0.71 | 1.00 | 0.50 | 1.00 | 1.00 | 2.00 | 2.00 | ND | [100,104] |
| C. dubliniensis | 1 | 0.03 | ND | ND | ND | ND | ND | 0.03 | ND | [100] |
| C. auris | 4 | 8.00 | 8.00 | 8.00 | ND | 4.00 | ND | 4.00 | ND | [105] |
RZF: rezafungin, ANF: anidulafungin, CSF: caspofungin, MCF: micafungin. a Geometric means of the data published in the cited references expressed in µg/mL. ND: no data available.
Table 3.
Rezafungin and FDA-approved echinocandin MEC values for Aspergillus spp. determined by CLSI microdilution reference methods.
Table 3.
Rezafungin and FDA-approved echinocandin MEC values for Aspergillus spp. determined by CLSI microdilution reference methods.
| Aspergillus spp. | n | RZF a,b | ANF a,b | CSF a,b | MCF a,b | References | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| MEC50 | MEC90 | MEC50 | MEC90 | MEC50 | MEC90 | MEC50 | MEC90 | |||
| A. fumigatus sensu stricto | 305 | 0.018 | 0.025 | 0.010 | 0.019 | 0.036 | 0.050 | 0.009 | 0.015 | [101,102,104,109] |
| A. calidoustus | 11 | 0.060 | 0.060 | ND | ND | 0.120 | 4.000 | 0.008 | 0.030 | [101] |
| A. lentulus | 11 | 0.080 | 0.080 | ND | ND | 0.060 | 0.250 | 0.008 | 0.030 | [101] |
| A. thermomutatus | 5 | 0.060 | ND | ND | ND | 0.060 | ND | 0.030 | ND | [101] |
| A. udagawae | 5 | 0.015 | ND | ND | ND | 0.060 | ND | 0.008 | ND | [101] |
| A. section Terrei | 19 | 0.015 | 0.015 | 0.015 | 0.015 | 0.120 | 0.250 | ND | ND | [104] |
| A. flavus | 12 | 0.015 | 0.015 | 0.015 | 0.015 | 0.120 | 0.250 | ND | ND | [104] |
| A. section Flavii | 57 | 0.004 | 0.015 | 0.004 | 0.015 | 0.015 | 0.030 | 0.015 | 0.030 | [102] |
| A. niger | 16 | 0.008 | 0.030 | 0.008 | 0.008 | 0.060 | 0.120 | ND | ND | [104] |
RZF: rezafungin, ANF: anidulafungin, CSF: caspofungin, MCF: micafungin. a Geometric means of the data published in the cited references expressed in µg/mL. ND: no data available. b Susceptibility shown are minimal effective concentration (MEC) values.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Garcia-Effron, G. Rezafungin—Mechanisms of Action, Susceptibility and Resistance: Similarities and Differences with the Other Echinocandins. J. Fungi 2020, 6, 262. https://doi.org/10.3390/jof6040262
AMA Style
Garcia-Effron G. Rezafungin—Mechanisms of Action, Susceptibility and Resistance: Similarities and Differences with the Other Echinocandins. Journal of Fungi. 2020; 6(4):262. https://doi.org/10.3390/jof6040262
Chicago/Turabian StyleGarcia-Effron, Guillermo. 2020. "Rezafungin—Mechanisms of Action, Susceptibility and Resistance: Similarities and Differences with the Other Echinocandins" Journal of Fungi 6, no. 4: 262. https://doi.org/10.3390/jof6040262
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.