
Farmed Salmon Show No Pathological Alterations When Exposed to Acoustic Treatment for Sea Lice Infestation

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Methods
2.1. Laboratory Experiments
2.1.1. Salmon Specimens Maintenance and Health Assessment
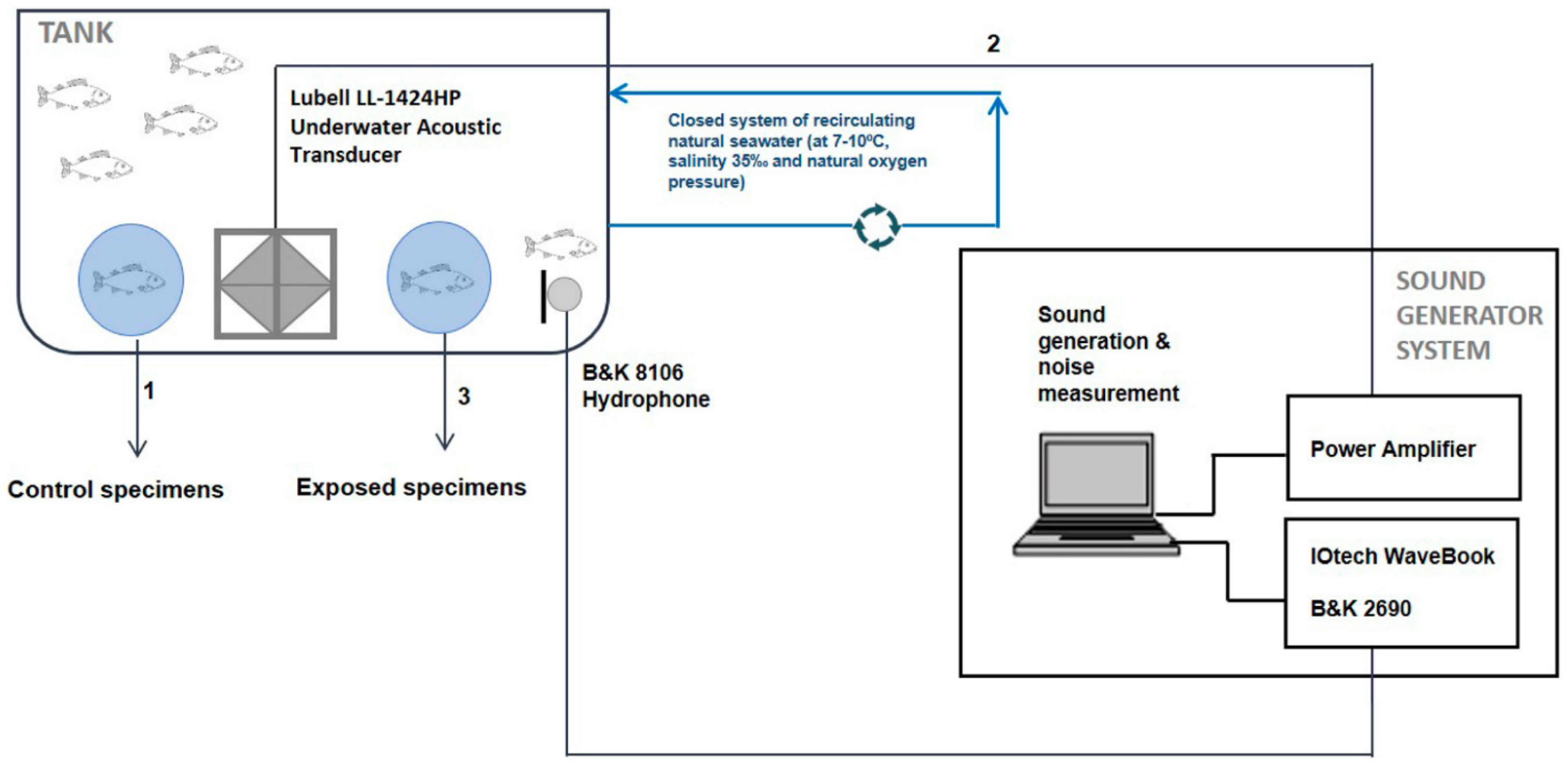
2.1.2. Sound Controlled Exposure Experiment (CEE) Protocol
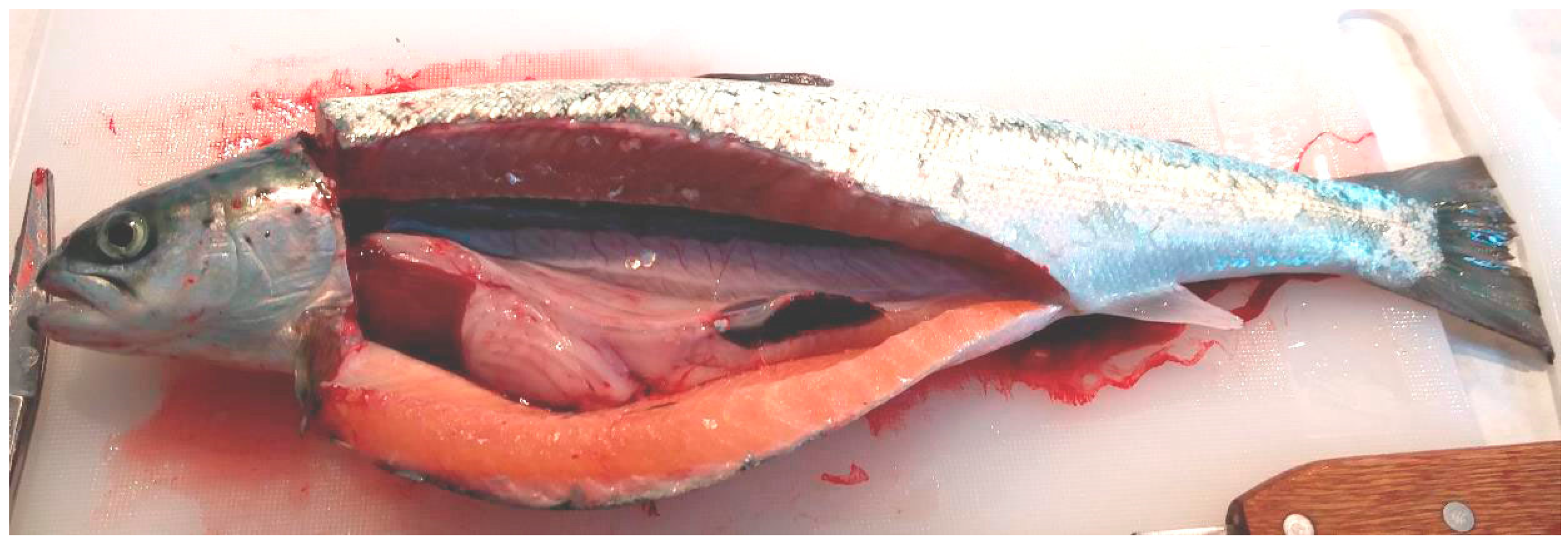
2.1.3. Sample Collection
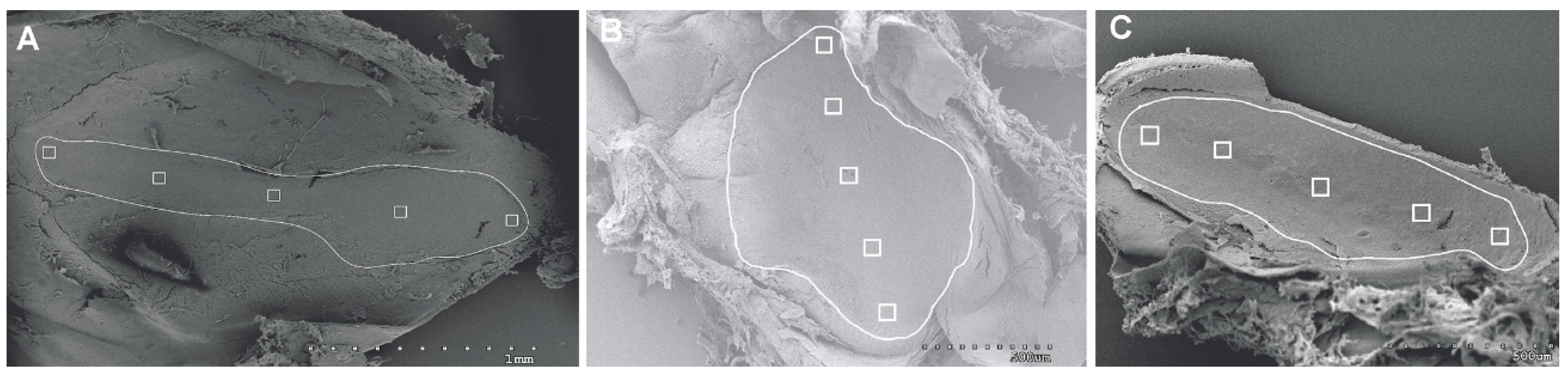
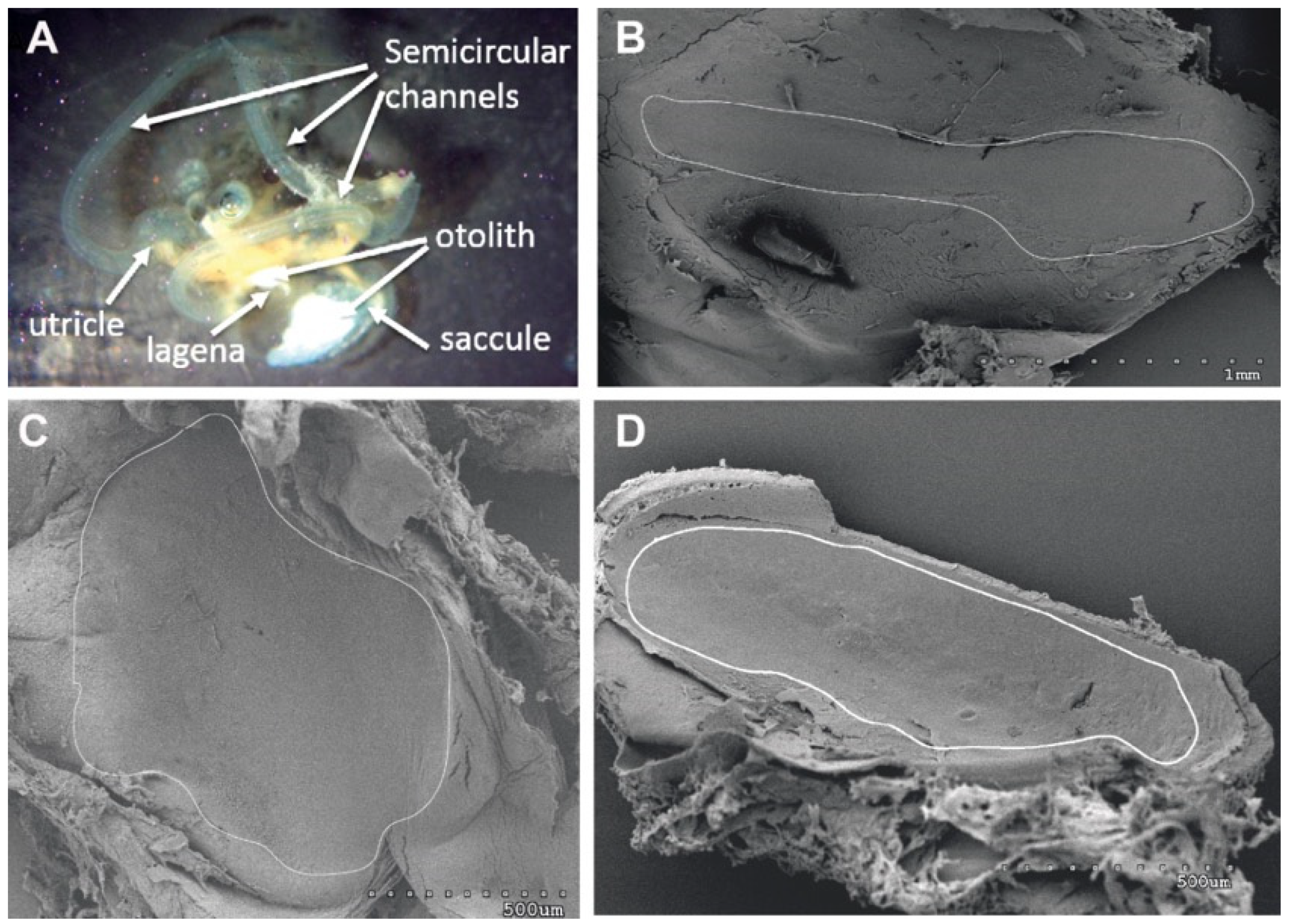
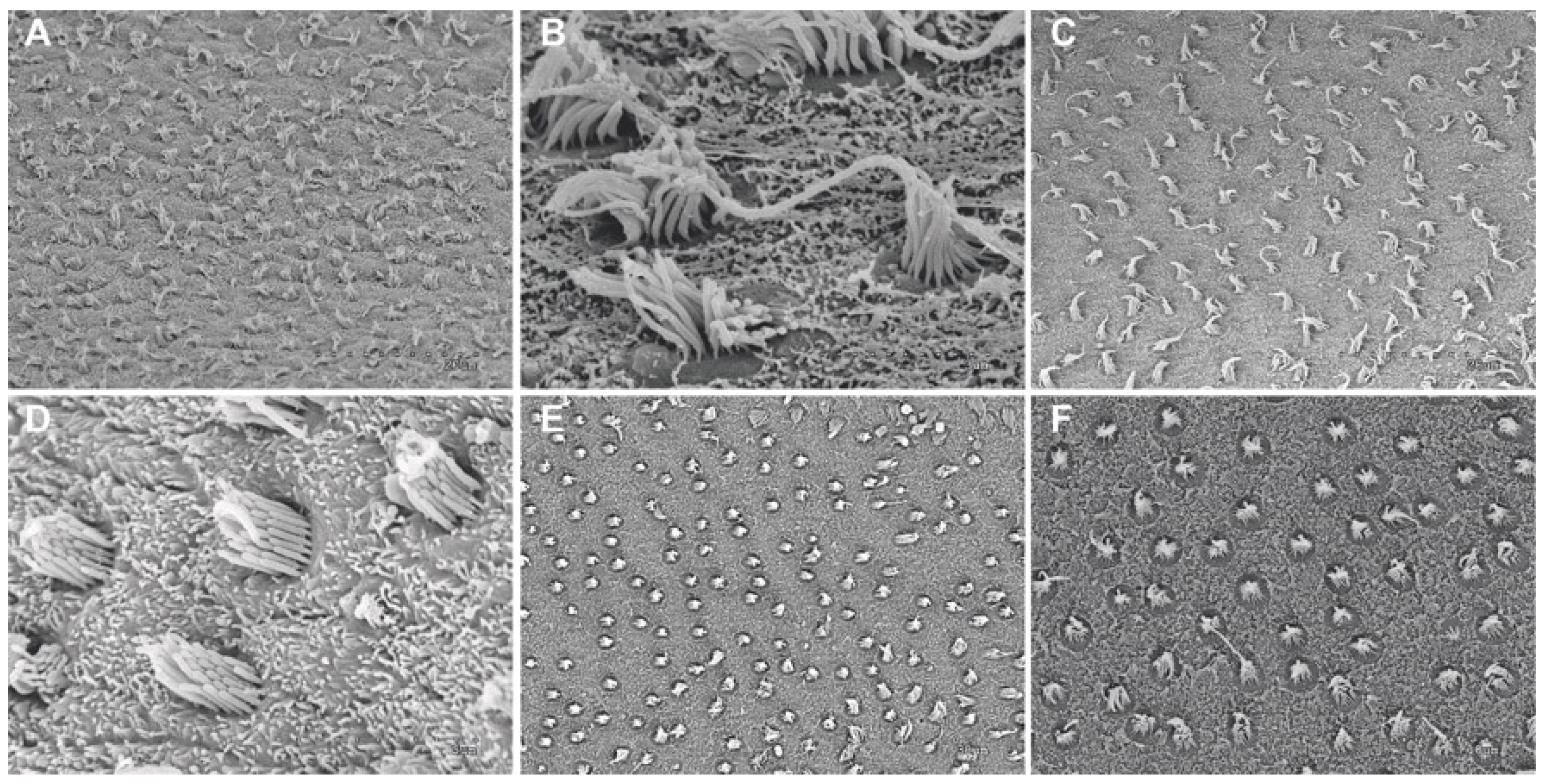
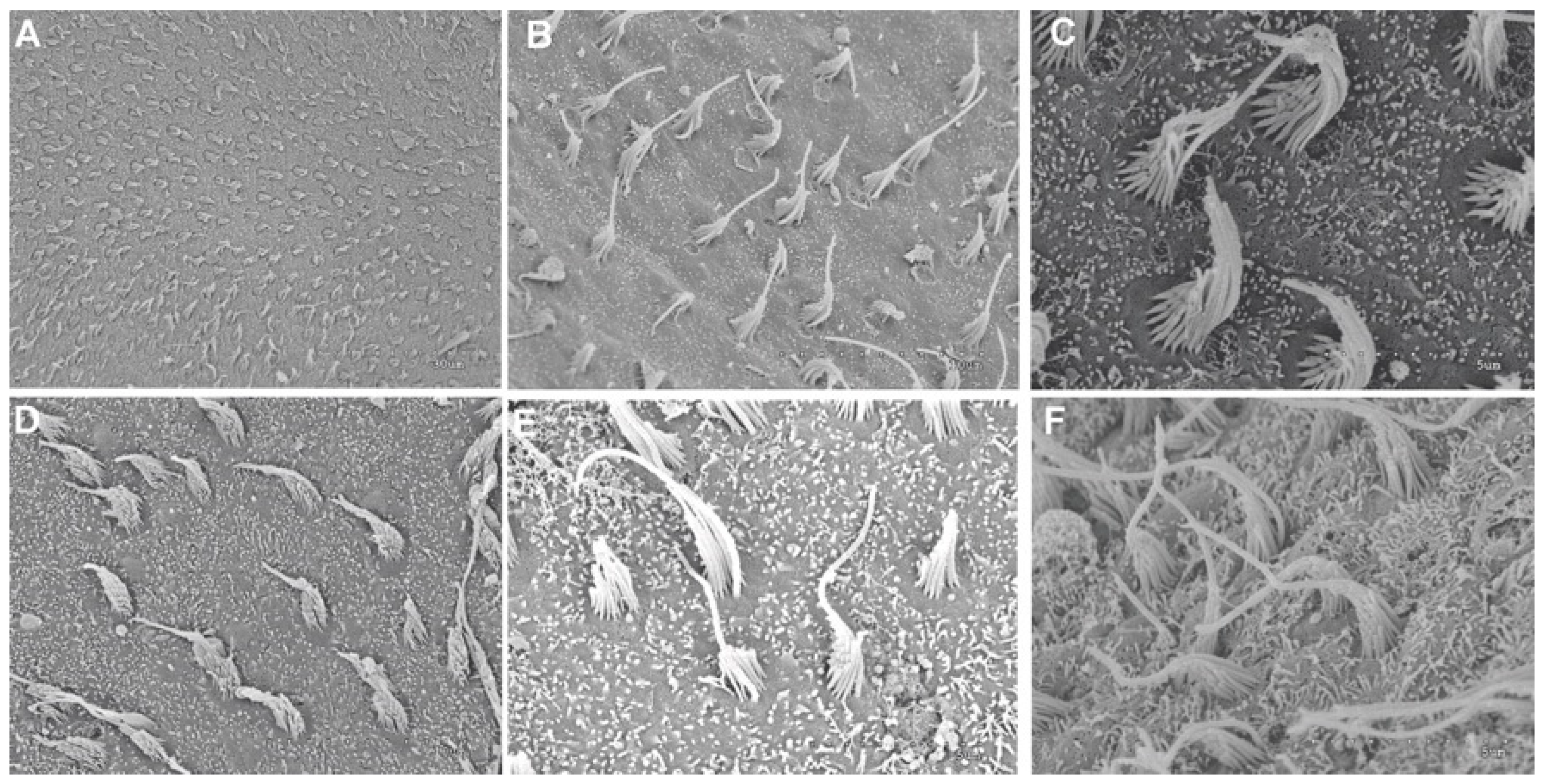
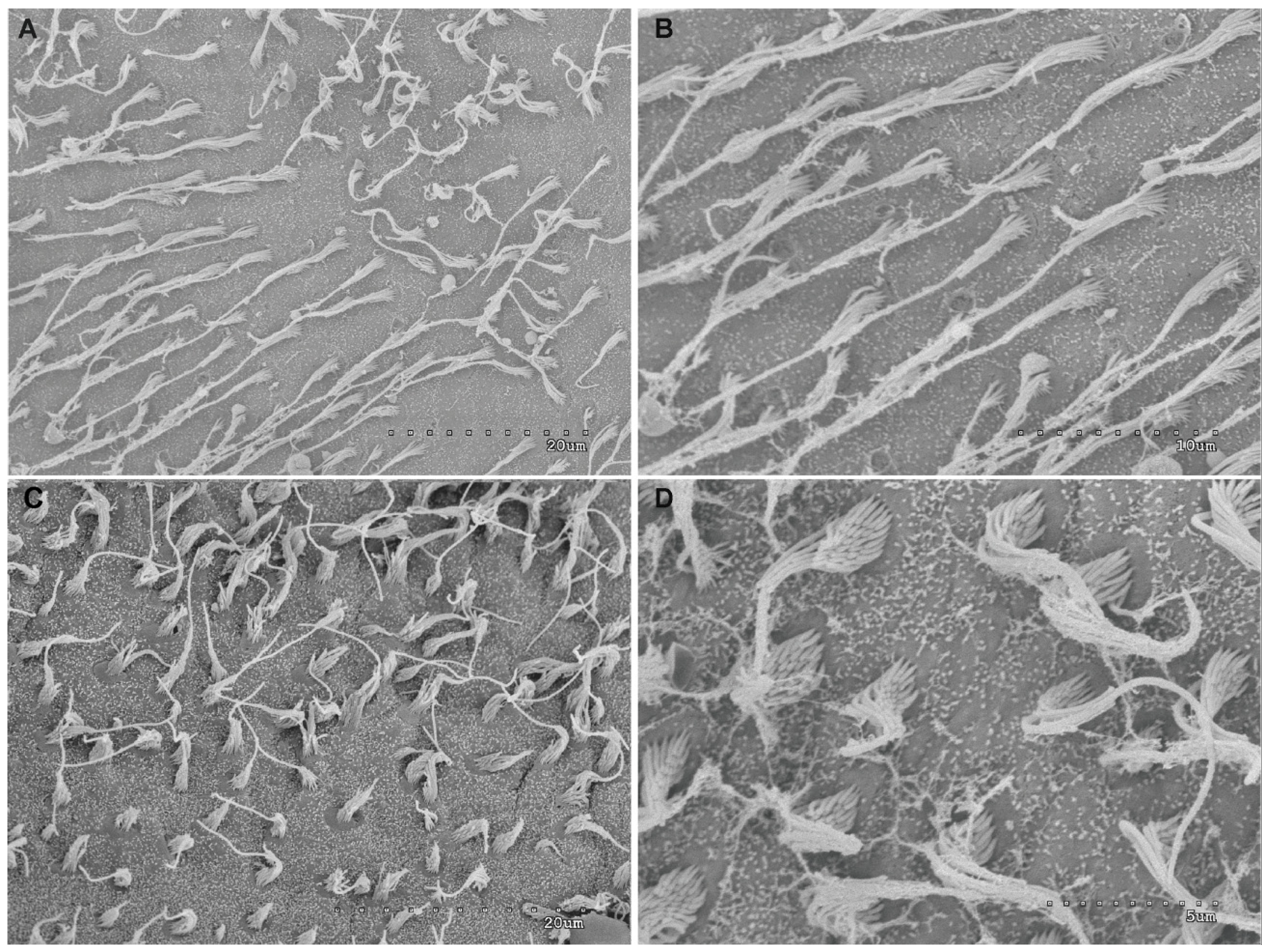
2.1.4. Analysis of Salmon Otolith Organs by Scanning Electron Microscopy
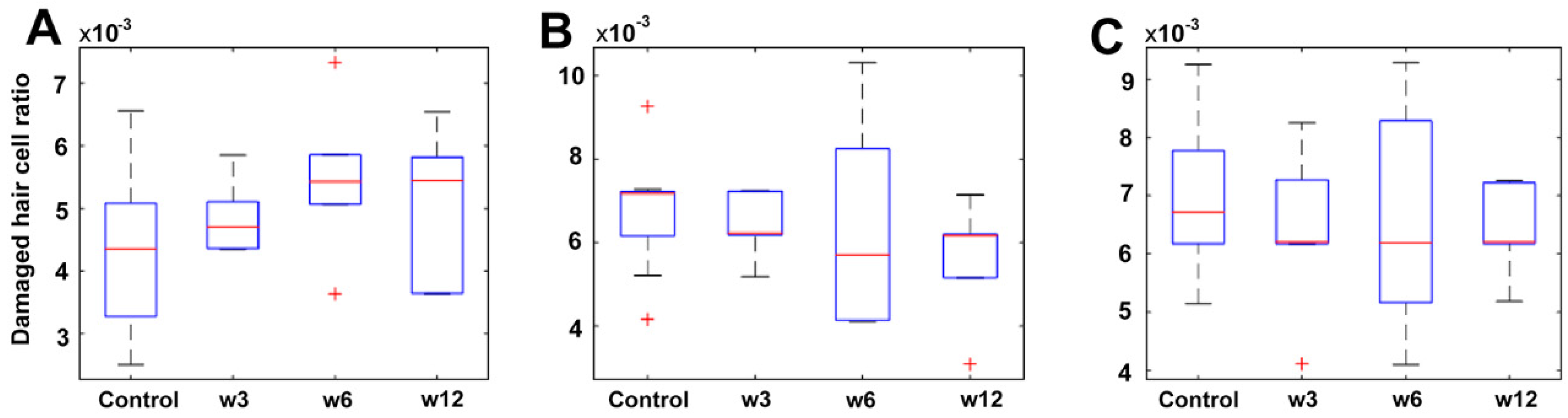
2.1.5. Quantification and Data Analysis
- All controls taken at different times were grouped together into a single control group.
- The two samples that were taken for each animal were combined by summing both the intact hair cells and the extruded or missing hair cells over the samples.
- The hair cells were summed over all the regions, obtaining a single intact hair cells count and single extruded/missing count per animal.
- The extruded/missing count was divided by the intact hair cell count.
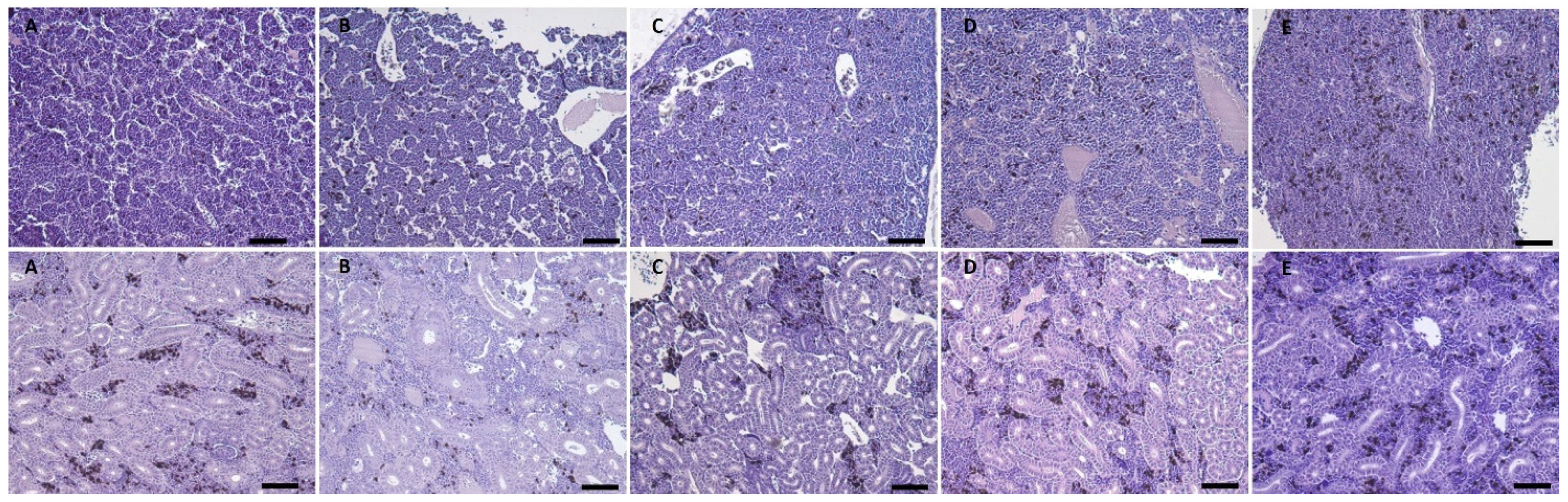
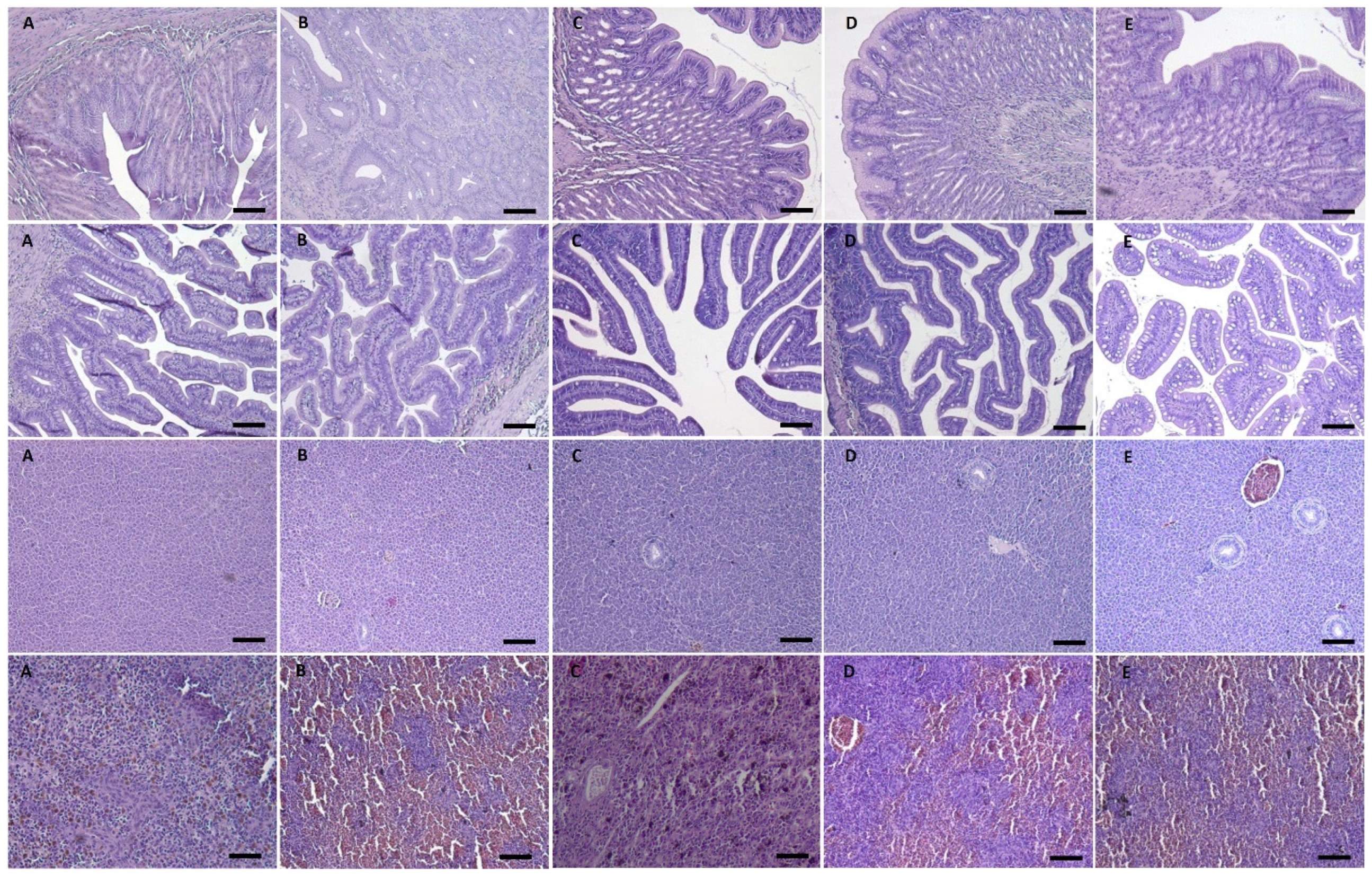
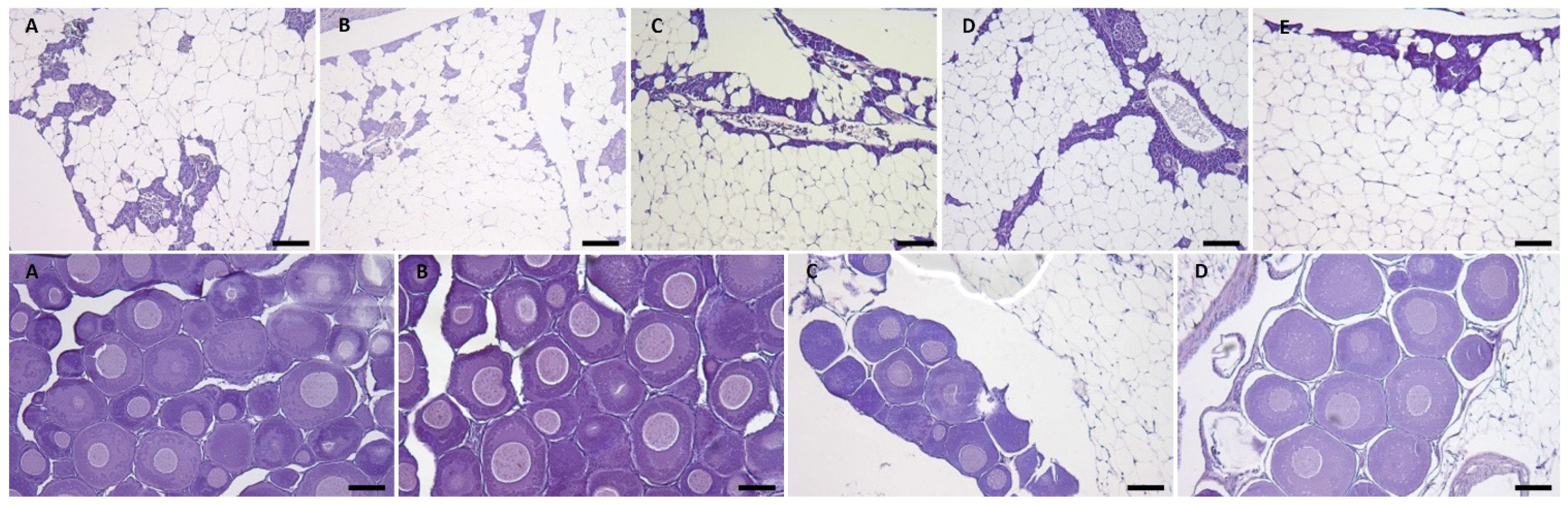
2.1.6. Salmon Gross Pathology and Histopathological Analysis
2.1.7. Behavioural Observations
2.2. Sea Trial Experiments
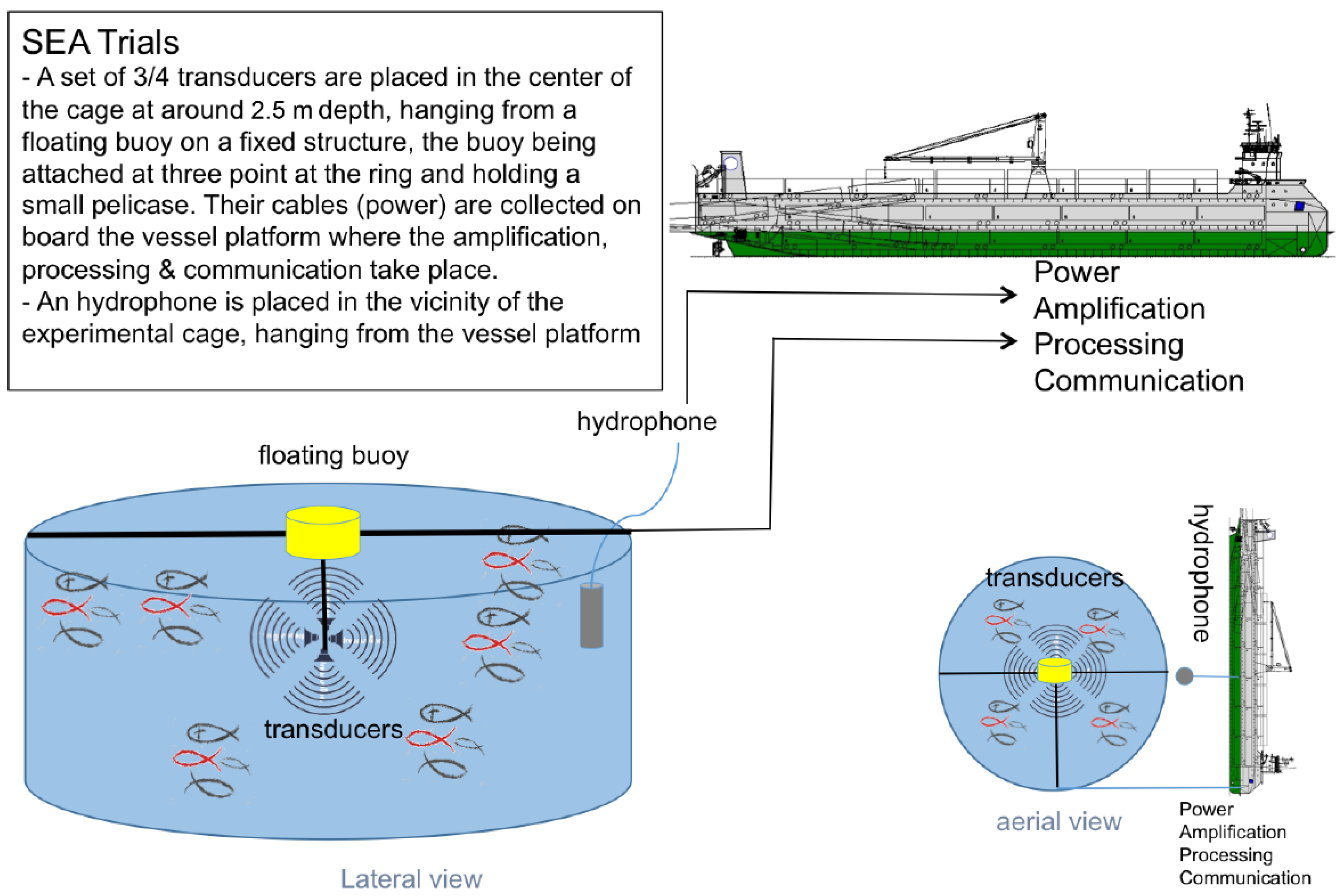
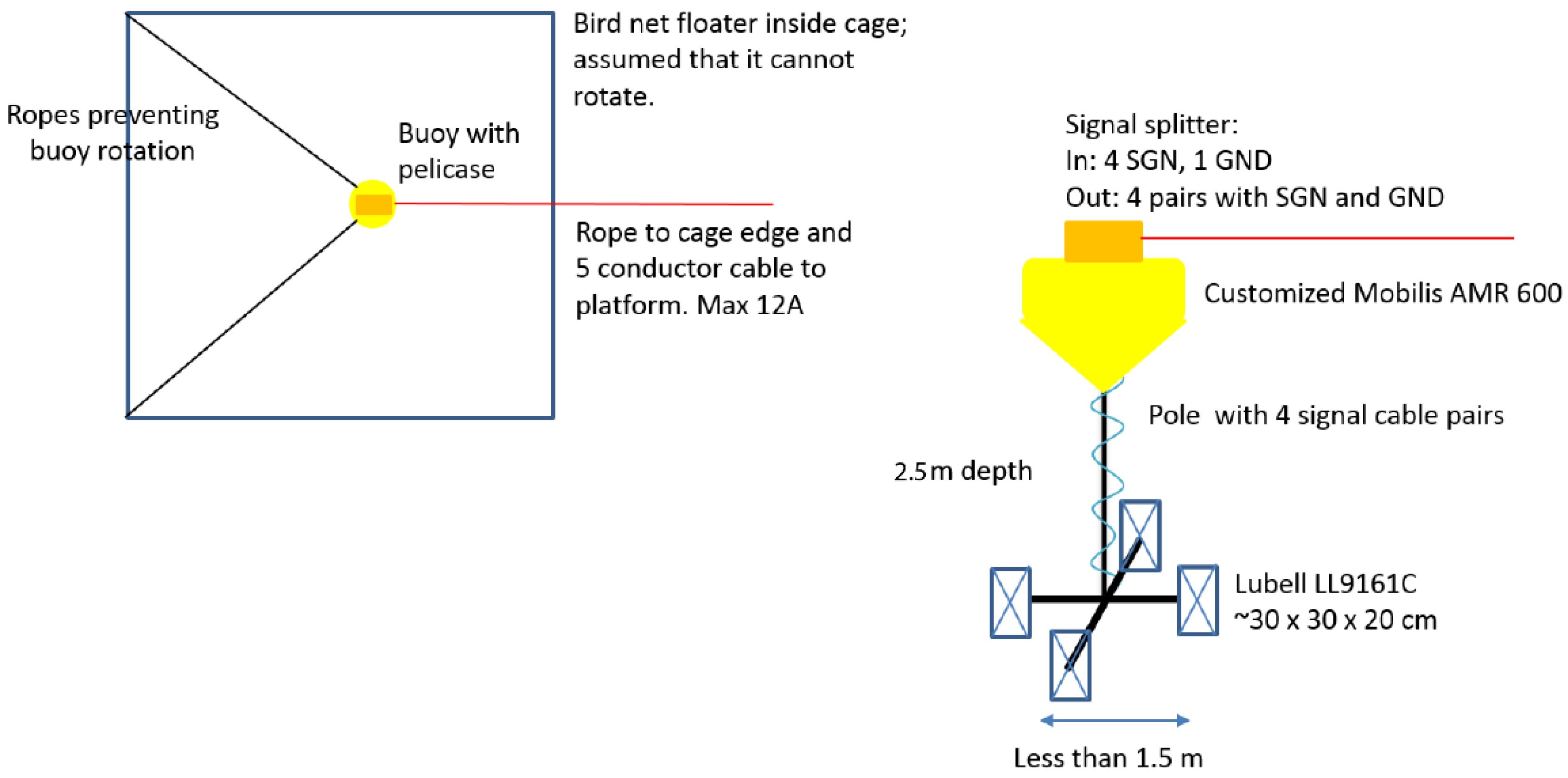
2.2.1. Sound Exposure
2.2.2. Sample Collection
- Week 3 and week 6: three salmon from each cage (taken from the two exposed cages and from the four control cages). Total: six exposed individuals and 12 control individuals per week.
- Week 9 and week 12: three salmon (from the two exposed cages and from the two control cages). Total: six exposed individuals and six control individuals per week.
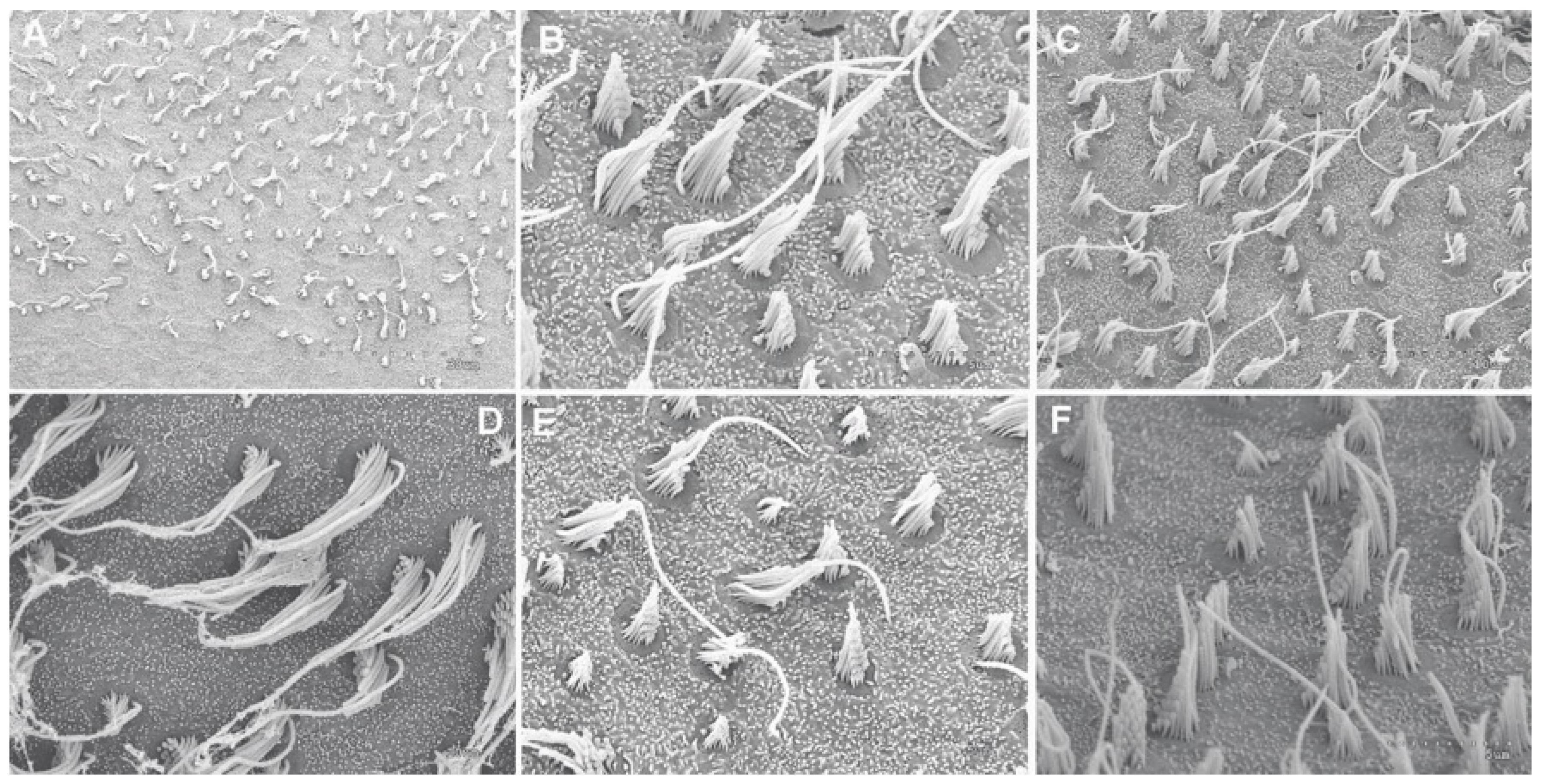
2.2.3. Analysis of Salmon Otolith Organs by Scanning Electron Microscopy
2.2.4. Quantification and Data Analysis
- All controls taken at different times were grouped together into a single control group.
- The two samples that were taken for each animal were combined by summing both the intact hair cells and the extruded or missing hair cells over the samples.
- The hair cells were summed over all the regions, obtaining a single intact hair cells count and single extruded/missing count per animal.
- The extruded/missing count was divided by the intact hair cell count.
2.2.5. Analysis of Presence of Vaterite in Salmon Otolith
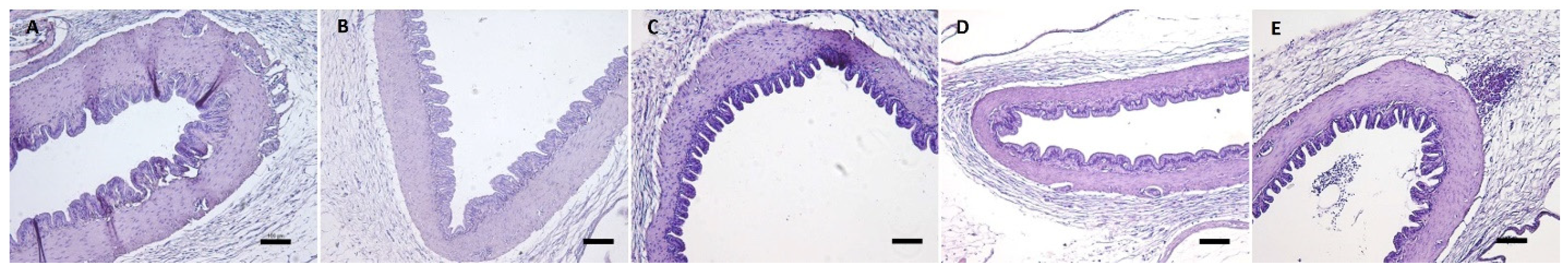
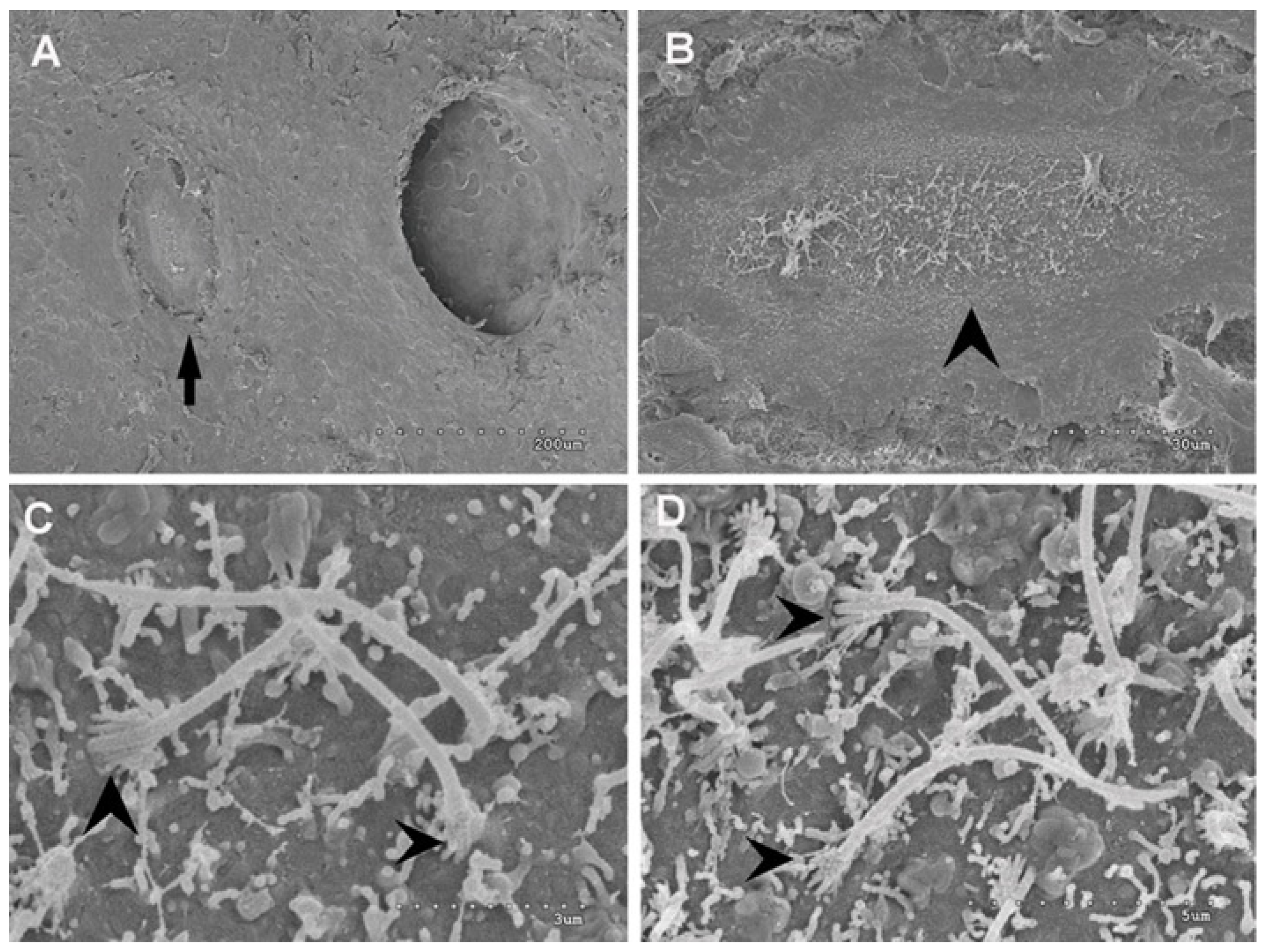
2.2.6. Analysis of Superficial Neuromasts of the Salmon Lateral Line by Scanning Electron Microscopy
2.2.7. Salmon Gross Pathology and Histopathological Analysis
3. Results
3.1. Laboratory Experiments
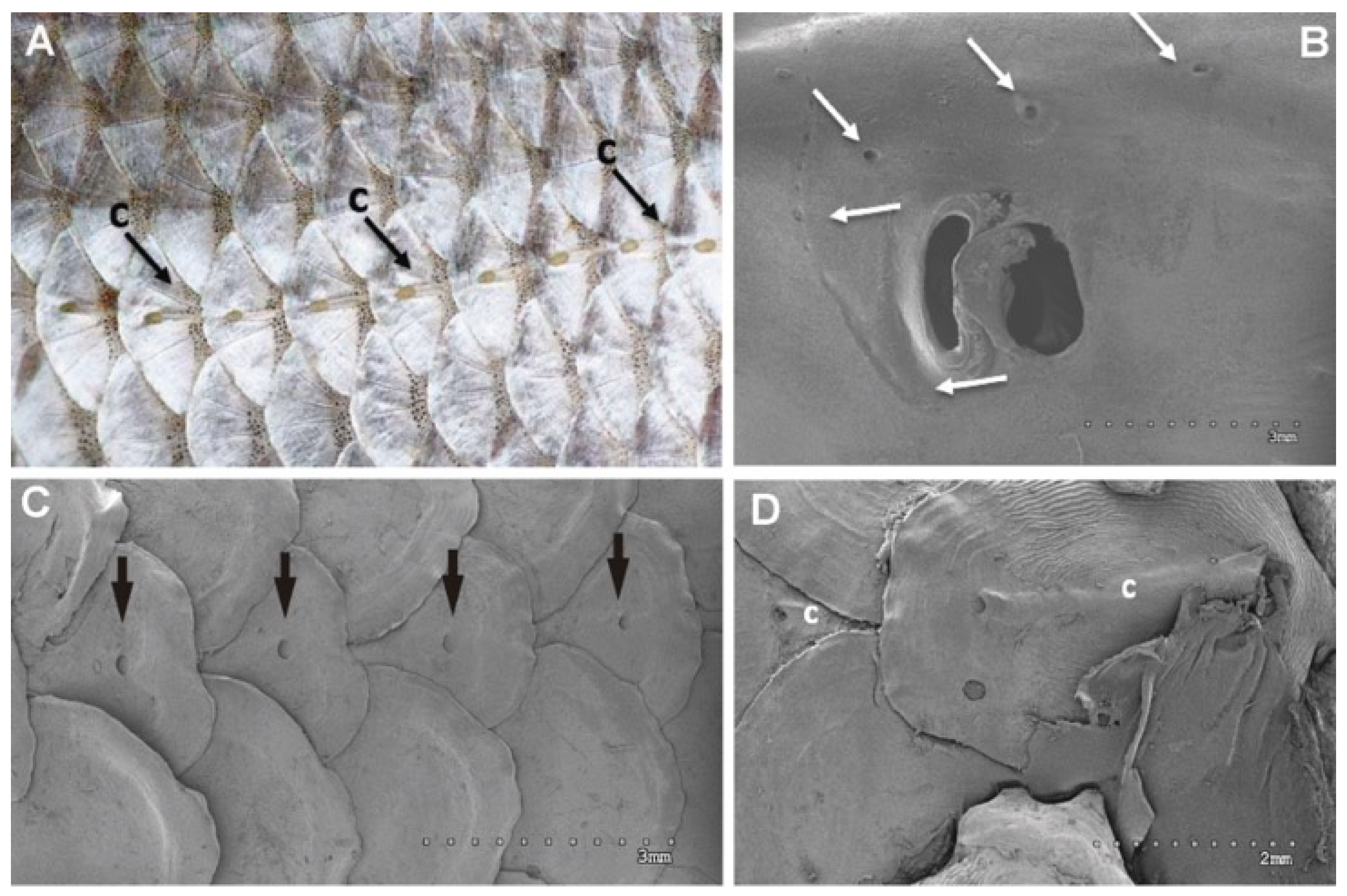
3.1.1. Analysis of Salmon Otolith Organs by Scanning Electron Microscopy
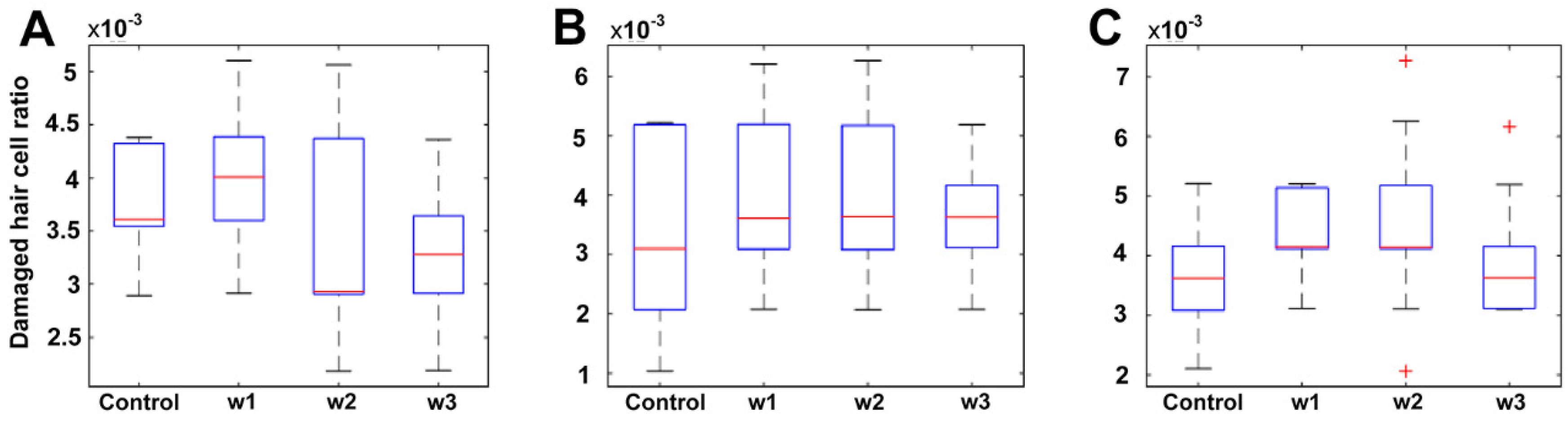
3.1.2. Quantification and Data Analysis
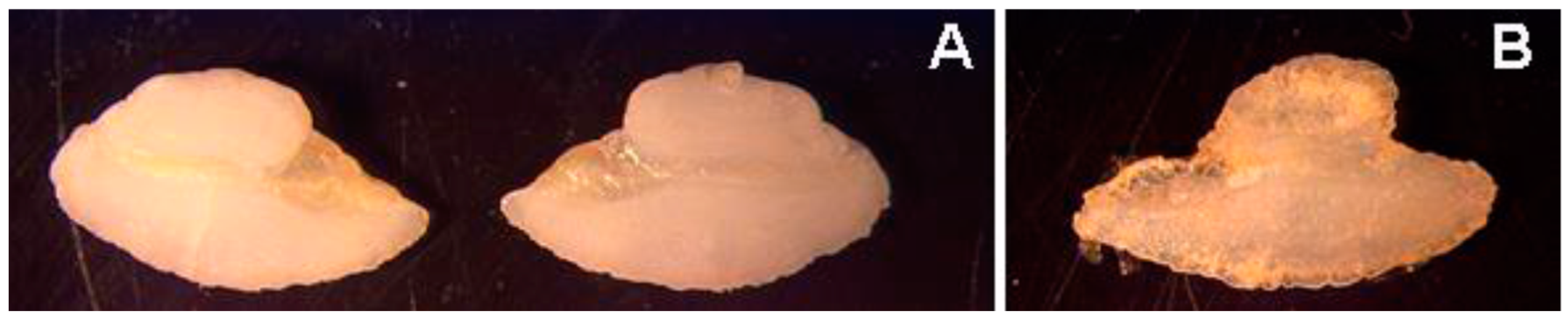
3.1.3. Gross Pathology and Histological Analysis
3.1.4. Behavioural Observations
3.2. Sea Trial Experiments
3.2.1. Analysis of Salmon Otolith Organs by Scanning Electron Microscopy
3.2.2. Quantification and Data Analysis
3.2.3. Analysis of Salmon Otolith
3.2.4. Analysis of Superficial Neuromast of the Salmon Lateral Line by Scanning Electron Microscopy
3.2.5. Gross Pathology and Histological Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Iversen, A.; Hermansen, Ø.; Andreassen, O.; Brandvik, R.K.; Marthinussen, A.; Nystøyl, R. Cost Drivers in Salmon Farming; Trondheim. 2015. Available online: https://nofima.no/en/pub/1281065/ (accessed on 13 October 2021).
- Barrett, L.T.; Oppedal, F.; Robinson, N.; Dempster, T. Prevention not cure: A review of methods to avoid sea lice infestations in salmon aquaculture. Rev. Aquac. 2020, 12, 2527–2543. [Google Scholar] [CrossRef]
- Solé, M.; Lenoir, M.; Fortuño, J.M.; De Vreese, S.; van der Schaar, M.; André, M. Sea Lice are sensitive to low frequency sounds. J. Mar. Sci. Eng. 2021, 9, 765. [Google Scholar] [CrossRef]
- Harding, H.; Bruintjes, R.; Radford, A.N.; Simpson, S.D. Measurement of Hearing in the Atlantic Salmon (Salmo salar) Using Auditory Evoked Potentials, and Effects of Pile Driving Playback on Salmon Behaviour and Physiology; Marine Scotland Science: Aberdeen, UK, 2016; Volume 7, 47p. [CrossRef]
- Smith, M.E.; Kane, A.S.; Popper, A.N. Noise-induced stress response and hearing loss in goldfish (Carassius auratus). J. Exp. Biol. 2004, 207, 427–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popper, A.N.; Smith, M.E.; Cott, P.A.; Hanna, B.W.; MacGillivray, A.O.; Austin, M.E.; Mann, D.A. Effects of exposure to seismic airgun use on hearing of three fish species. J. Acoust. Soc. Am. 2005, 117, 3958–3971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastings, M.C. Coming to terms with the effects of ocean noise on marine animals. Acoust. Today 2008, 4, 22–34. [Google Scholar] [CrossRef]
- McCauley, R.D.; Fewtrell, J.; Popper, A.N. High intensity anthropogenic sound damages fish ears. J. Acoust. Soc. Am. 2003, 113, 638–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uribe, P.M.; Sun, H.; Wang, K.; Asuncion, J.D.; Wang, Q.; Chen, C.W.; Steyger, P.S.; Smith, M.E.; Matsui, J.I. Aminoglycoside-induced hair cell death of inner ear organs causes functional deficits in adult zebrafish (Danio rerio). PLoS ONE 2013, 8, e58755. [Google Scholar] [CrossRef]
- Smith, M.E.; Schuck, J.B.; Gilley, R.R.; Rogers, B.D. Structural and functional effects of acoustic exposure in goldfish: Evidence for tonotopy in the teleost saccule. BMC Neurosci. 2011, 12, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedwell, J.R.; Turnpenny, A.W.H.; Lovell, J.M.; Edwards, B. An investigation into the effects of underwater piling noise on salmonids. J. Acoust. Soc. Am. 2006, 120, 2550–2554. [Google Scholar] [CrossRef]
- Ruggerone, G.T.; Goodman, S.E.; Miner, R. Behavioral Response and Survival of Juvenile Coho Salmon to Pile Driving Sounds; Natural Resources Consultants, Inc.: Seattle, DC, USA, 2008; Available online: http://home.comcast.net/∼ruggerone/FishTerminal%0APileDriveStudy.pdf (accessed on 13 October 2021).
- Casper, B.M.; Smith, M.E.; Halvorsen, M.B.; Sun, H.; Carlson, T.J.; Popper, A.N. Effects of exposure to pile driving sounds on fish inner ear tissues. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2013, 166, 352–360. [Google Scholar] [CrossRef]
- Reimer, T.; Dempster, T.; Wargelius, A.; Fjelldal, P.G.; Hansen, T.; Glover, K.A.; Solberg, M.F.; Swearer, S.E. Rapid growth causes abnormal vaterite formation in farmed fish otoliths. J. Exp. Biol. 2017, 220, 2965–2969. [Google Scholar] [CrossRef] [Green Version]
- Skalski, J.R.; Pearson, W.H.; Malme, C.I. Effects of sounds from a geophysical survey device on catch-per-unit-effort in a hook-and-line fishery for rockfish (Sebastes spp.). Can. J. Fish. Aquat. Sci. 1992, 49, 1357–1365. [Google Scholar] [CrossRef]
- Wardle, C.S.; Carter, T.J.; Urquhart, G.G.; Johnstone, A.D.F.; Ziolkowski, A.M.; Hampson, G.; Mackie, D. Effects of seismic air guns on marine fish. Cont. Shelf Res. 2001, 21, 1005–1027. [Google Scholar] [CrossRef]
- Boeger, W.A.; Pie, M.R.; Ostrensky, A.; Cardoso, M.F. The effect of exposure to seismic prospecting on coral reef fishes. Brazilian J. Oceanogr. 2006, 54, 235–239. [Google Scholar] [CrossRef]
- Herbert-Read, J.E.; Kremer, L.; Bruintjes, R.; Radford, A.N.; Ioannou, C.C. Anthropogenic noise pollution from pile-driving disrupts the structure and dynamics of fish shoals. Proc. R. Soc. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weilgart, L.S. The Impact of Ocean Noise Pollution on Fish and Invertebrate; Ocean Care and Dalhousie University. 2018, p. 34. Available online: https://www.oceancare.org/wp-content/uploads/2017/10/OceanNoise_FishInvertebrates_May2018.pdf (accessed on 13 October 2021).
- Nedelec, S. Impacts of Anthropogenic Noise on Behaviour, Development and Fitness of Fishes and Invertebrates. Ecosystems. École Pratique Des Hautes Études-EPHE PARIS. University of Bristol. 2015. Available online: https://tel.archives-ouvertes.fr/tel-02099631/document (accessed on 13 October 2021).
- Wysocki, L.E.; Davidson, J.W.I.; Smith, M.E.; Frankel, A.S.; Ellison, W.T.; Mazik, P.M.; Popper, A.N.; Bebak, J. Effects of aquaculture production noise on hearing, Growth, and disease resistance of rainbow trout. Oncorhynchus Mykiss. Aquac. 2007, 272, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Popper, A.N.; Hawkins, A.D.; Halvorsen, M.B. Anthropogenic Sound and Fishes; 2019. Available online: https://www.wsdot.wa.gov/research/reports/fullreports/891-1.pdf (accessed on 13 October 2021).
- Angelica, M.D.; Fong, Y. Cell proliferation follows acoustically-induced hair cell bundle loss in the zebrafish saccule. Hear. Res. 2009, 253, 67–76. [Google Scholar] [CrossRef]
- Oxman, D.S.; Barnett-Johnson, R.; Smith, M.E.; Coffin, A.; Miller, D.L.; Josephson, R.; Popper, A.N. The effect of vaterite deposition on sound reception, otolith morphology, and inner ear sensory epithelia in hatchery-reared Chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. Sci. 2007, 64, 1469–1478. [Google Scholar] [CrossRef] [Green Version]
- Uribe, P.M.; Villapando, B.K.; Lawton, K.J.; Fang, Z.; Gritsenko, D.; Bhandiwad, A.; Sisneros, J.A.; Xu, J.; Coffin, A.B. Larval zebrafish lateral line as a model for acoustic trauma. eNeuro 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halvorsen, M.B.; Casper, B.M.; Woodley, C.M.; Carlson, T.J.; Popper, A.N. Threshold for onset of injury in chinook salmon from exposure to impulsive pile driving sounds. PLoS ONE 2012, 7, e38968. [Google Scholar] [CrossRef] [PubMed]
- Whelan, K. A review of the impacts of the salmon louse, Lepeophtheirus salmonis (Krøyer, 1837) on wild salmonids. Atl. Salmon Trust 2010. Available online: https://atlanticsalmontrust.org/wp-content/uploads/2016/12/ast-sea-lice-impacts-review1.pdf (accessed on 13 October 2021).
- Urbina, M.A.; Cumillaf, J.P.; Paschke, K.; Gebauer, P. Effects of pharmaceuticals used to treat salmon lice on non-target species: Evidence from a systematic review. Sci. Total Environ. 2018, 649, 1124–1136. [Google Scholar] [CrossRef] [PubMed]
- Hannisdal, R.; Nøstbakken, O.J.; Hove, H.; Madsen, L.; Horsberg, T.E.; Lunestad, B.T. Anti-sea lice agents in Norwegian aquaculture; surveillance, treatment trends and possible implications for food safety. Aquaculture 2020, 521, 735044. [Google Scholar] [CrossRef]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Vaterite Incidence Rate (%) |
|---|---|
| Week 3 | 75 |
| Week 6 | 83.3 |
| Week 12 | 16.6 |
| Total Incidence | - |
| Vateritic otolith | 65.6 |
| No vateritic otolith | 34.3 |
| Location | - |
| Right otolith | 52 |
| Left otolith | 48 |
| Cages | - |
| Control | 75 |
| Exposed | 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solé, M.; Constenla, M.; Padrós, F.; Lombarte, A.; Fortuño, J.-M.; van der Schaar, M.; André, M. Farmed Salmon Show No Pathological Alterations When Exposed to Acoustic Treatment for Sea Lice Infestation. J. Mar. Sci. Eng. 2021, 9, 1114. https://doi.org/10.3390/jmse9101114
Solé M, Constenla M, Padrós F, Lombarte A, Fortuño J-M, van der Schaar M, André M. Farmed Salmon Show No Pathological Alterations When Exposed to Acoustic Treatment for Sea Lice Infestation. Journal of Marine Science and Engineering. 2021; 9(10):1114. https://doi.org/10.3390/jmse9101114
Chicago/Turabian StyleSolé, Marta, Maria Constenla, Francesc Padrós, Antoni Lombarte, José-Manuel Fortuño, Mike van der Schaar, and Michel André. 2021. "Farmed Salmon Show No Pathological Alterations When Exposed to Acoustic Treatment for Sea Lice Infestation" Journal of Marine Science and Engineering 9, no. 10: 1114. https://doi.org/10.3390/jmse9101114