
Combined Culture and DNA Metabarcoding Analysis of Cyanobacterial Community Structure in Response to Coral Reef Health Status in the South China Sea
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
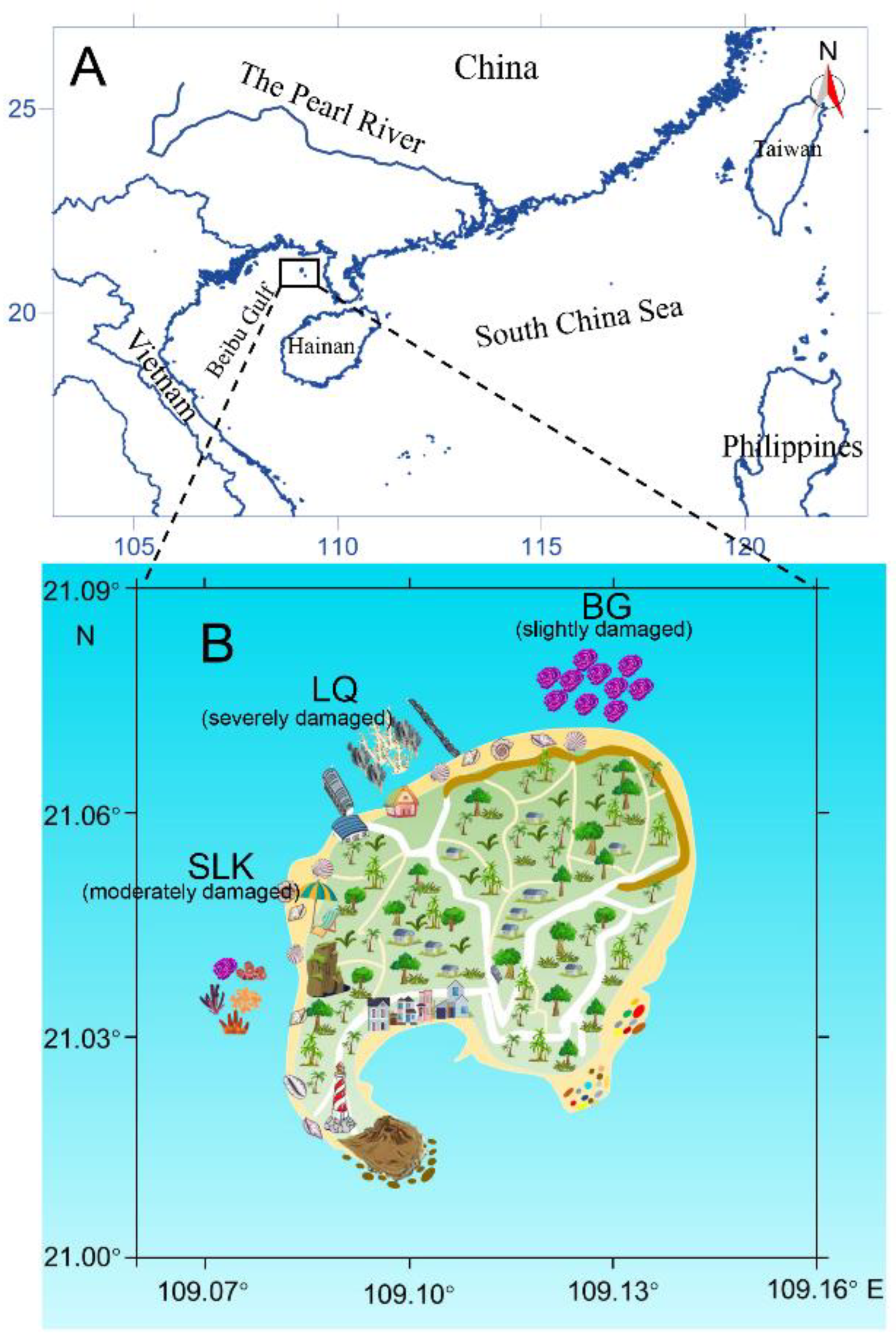
2.1. Study Area
2.2. Seawater, Coral Tissue, and Reef Sediment Sampling
2.3. Environmental DNA Extraction and Metabarcoding Sequencing
2.4. Sequence Data Processing
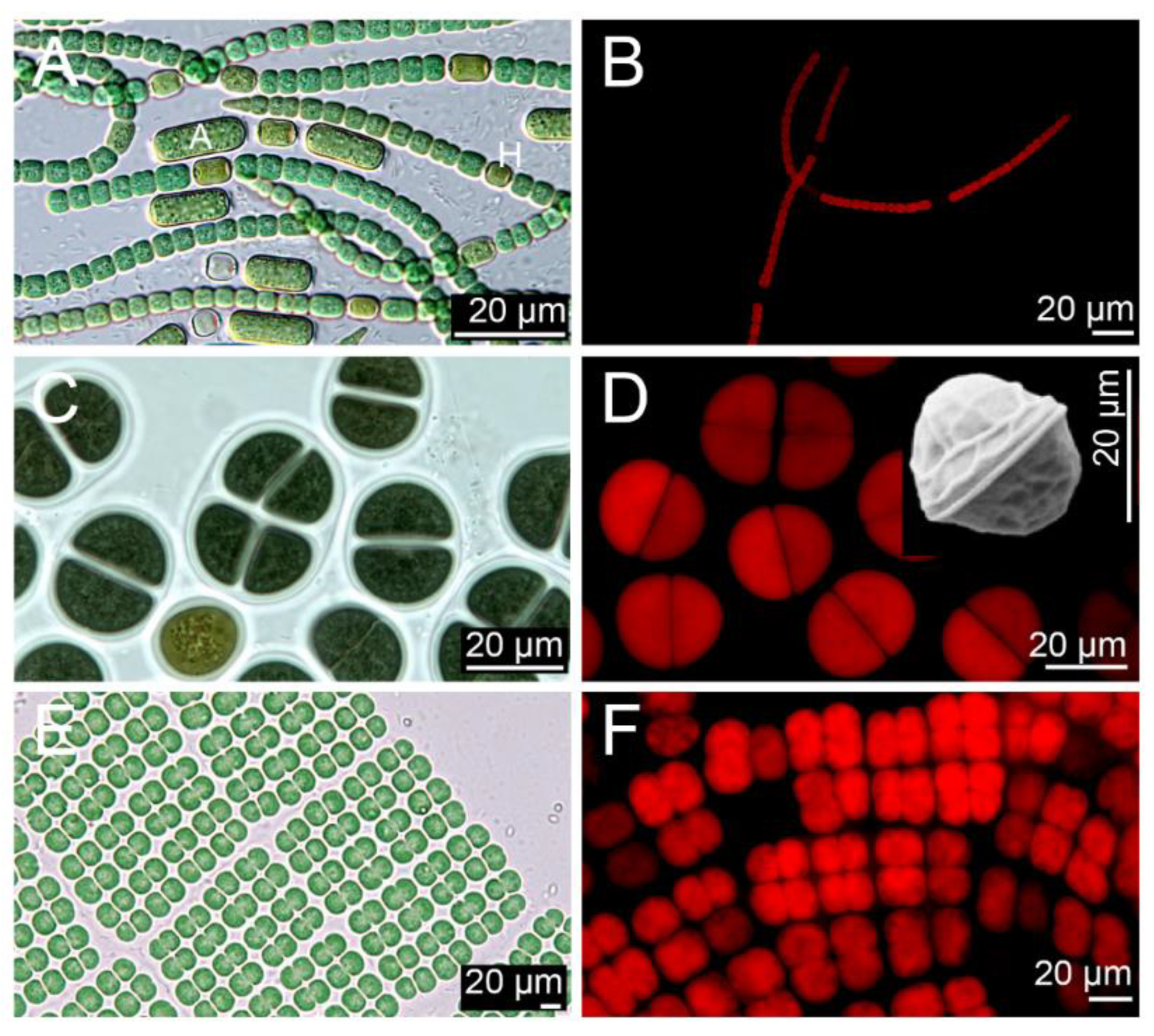
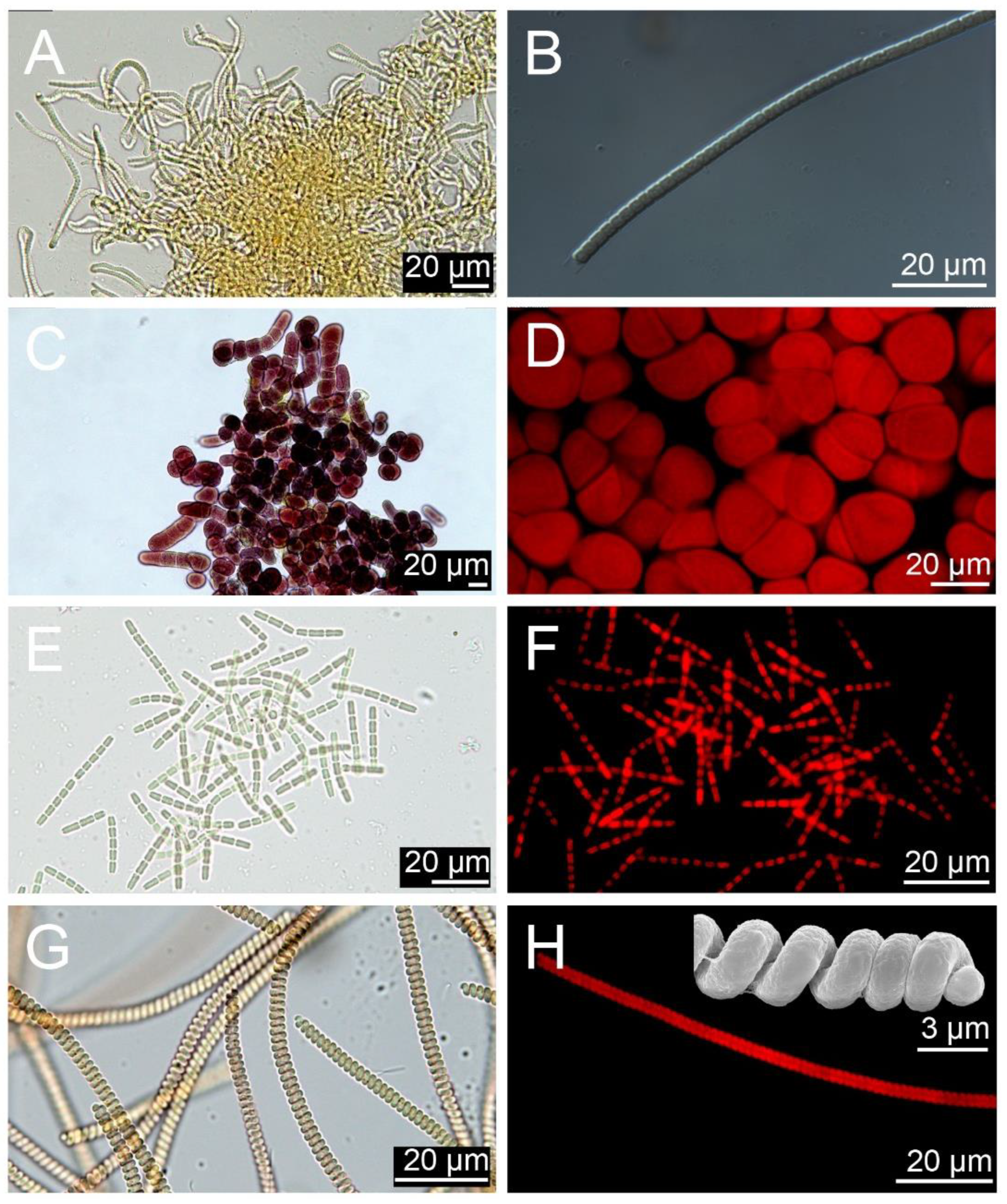
2.5. Clone Strains and Microscopy
3. Results
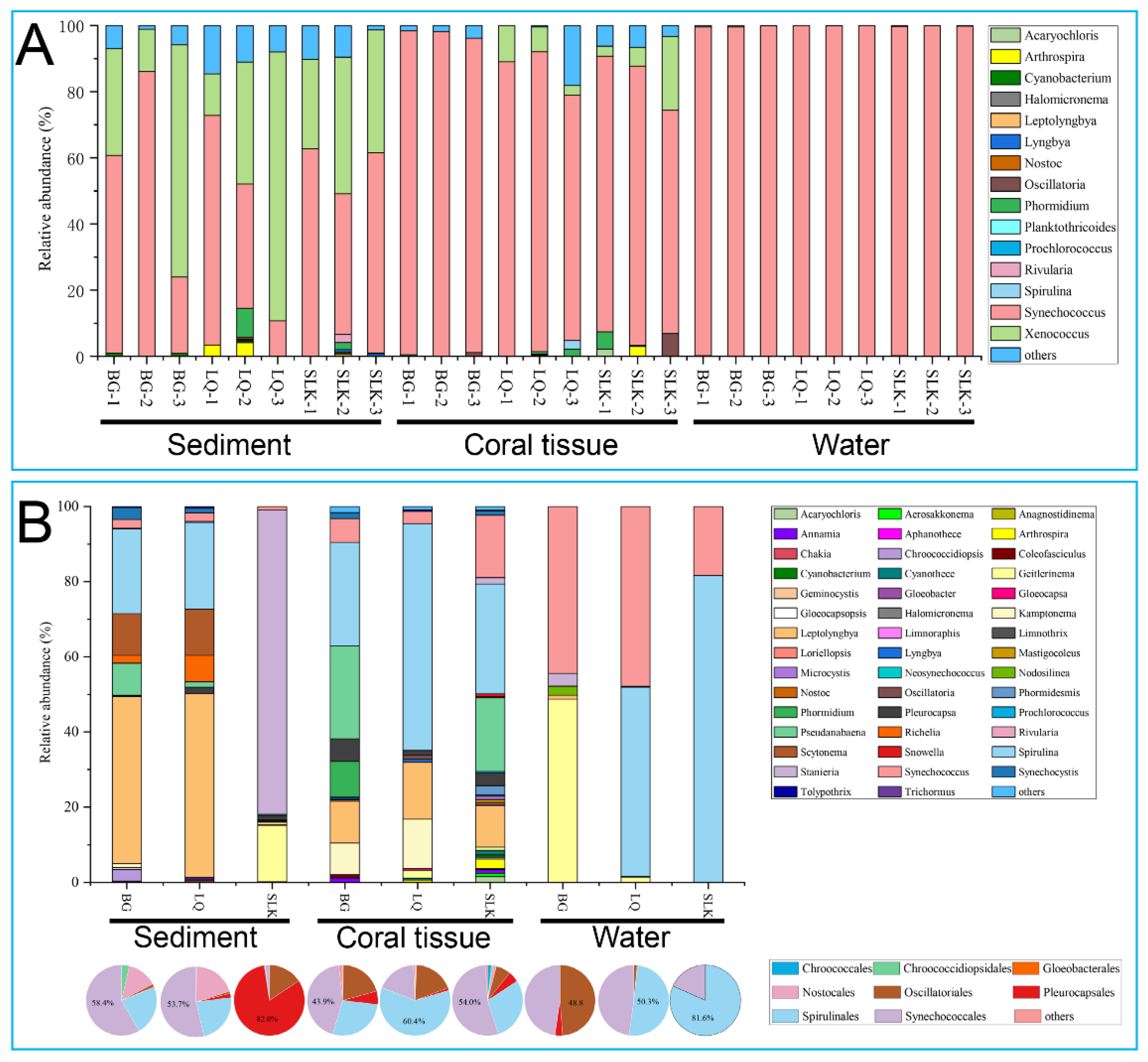
3.1. Diversity of Cyanobacteria Based on Universal Prokaryotic 16S rRNA Gene Primers
3.2. Diversity of Cyanobacteria Based on Cyanobacteria-Specific 16S rRNA Gene Primers
3.3. Diversity of Cyanobacterial Phenotypes
4. Discussion
4.1. Comparison of Different Primer Datasets
4.2. Cyanobacterial Diversity at Weizhou Island
4.3. Biological Nitrogen-Fixing Cyanobacteria
4.4. Harmful Cyanobacterial Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darling, E.S.; McClanahan, T.R.; Maina, J.; Gurney, G.G.; Graham, N.A.J.; Januchowski-Hartley, F.; Cinner, J.E.; Mora, C.; Hicks, C.C.; Maire, E.; et al. Social–environmental drivers inform strategic management of coral reefs in the anthropocene. Nat. Ecol. Evol. 2019, 3, 1341–1350. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef]
- Guo, J.; Yu, K.; Wang, Y.; Xu, D.; Huang, X.; Zhao, M.; Yang, H.; Zhang, R. Nutrient distribution in coral reef degraded areas within sanya bay, South China Sea. J. Coast. Res. 2017, 33, 1148–1160. [Google Scholar] [CrossRef]
- McManus, J.W.; Polsenberg, J.F. Coral–algal phase shifts on coral reefs: Ecological and environmental aspects. Prog. Oceanogr. 2004, 60, 263–279. [Google Scholar] [CrossRef]
- Adam, T.C.; Burkepile, D.E.; Holbrook, S.J.; Carpenter, R.C.; Claudet, J.; Loiseau, C.; Thiault, L.; Brooks, A.J.; Washburn, L.; Schmitt, R.J. Landscape-scale patterns of nutrient enrichment in a coral reef ecosystem: Implications for coral to algae phase shifts. Ecol. Appl. 2021, 31, e2227. [Google Scholar] [CrossRef] [PubMed]
- De Bakker, D.M.; Van Duyl, F.C.; Bak, R.P.M.; Nugues, M.M.; Nieuwland, G.; Meesters, E.H. 40 Years of benthic community change on the Caribbean reefs of Curaçao and Bonaire: The rise of slimy cyanobacterial mats. Coral Reefs 2017, 36, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Reverter, M.; Jackson, M.; Daraghmeh, N.; von Mach, C.; Milton, N. 11-yr of coral community dynamics in reefs around Dahab (Gulf of Aqaba, Red Sea): The collapse of urchins and rise of macroalgae and cyanobacterial mats. Coral Reefs 2020, 39, 1605–1618. [Google Scholar] [CrossRef]
- Ilsa, B.K.; Linda, J.W.; Mikel, A.B.; Valerie, J.P.; Raphael, R.-W.; Kevin, S.B. Inhibition of coral recruitment by macroalgae and cyanobacteria. Mar. Ecol. Prog. Ser. 2006, 323, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Seckbach, J.; Oren, A. Microbial Mats: Modern and Ancient Microorganisms in Stratified Systems, 2010th ed.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; Volume 14. [Google Scholar]
- Bowers, H.A.; Ryan, J.P.; Hayashi, K.; Woods, A.L.; Marin, R.; Smith, G.J.; Hubbard, K.A.; Doucette, G.J.; Mikulski, C.M.; Gellene, A.G.; et al. Diversity and toxicity of Pseudo-nitzschia species in Monterey Bay: Perspectives from targeted and adaptive sampling. Harmful Algae 2018, 78, 129–141. [Google Scholar] [CrossRef]
- Cissell, E.C.; McCoy, S.J. Shotgun metagenomic sequencing reveals the full taxonomic, trophic, and functional diversity of a coral reef benthic cyanobacterial mat from Bonaire, Caribbean Netherlands. Sci. Total Environ. 2021, 755, 142719. [Google Scholar] [CrossRef]
- Ciebiada, M.; Kubiak, K.; Daroch, M. Modifying the cyanobacterial metabolism as a Key to efficient biopolymer production in photosynthetic microorganisms. Int. J. Mol. Sci. 2020, 21, 7204. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.L.; Gunasekera, S.P.; Scott, R.M.; Paul, V.J.; Teplitski, M. Microbiome shifts and the inhibition of quorum sensing by black band disease cyanobacteria. ISME J. 2016, 10, 1204–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cissell, E.C.; Eckrich, C.E.; McCoy, S.J. Cyanobacterial mats as benthic reservoirs and vectors for coral black band disease pathogens. Ecol. Appl. 2022, 32, e2692. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.W.; Meier, O.W. Quantification of loss and change in Floridian reef coral populations. Am. Zool. 2015, 32, 625–640. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, V. Eutrophication, toxic cyanobacteria and cyanotoxins: When ecosystems cry for help. Limnetica 2006, 25, 425–432. [Google Scholar] [CrossRef]
- Regueiras, A.; Pereira, S.; Costa, M.S.; Vasconcelos, V. Differential toxicity of cyanobacteria Isolated from marine sponges towards echinoderms and crustaceans. Toxins 2018, 10, 297. [Google Scholar] [CrossRef] [Green Version]
- Sini, P.; Dang, T.B.C.; Fais, M.; Galioto, M.; Padedda, B.M.; Lugliè, A.; Iaccarino, C.; Crosio, C. Cyanobacteria, cyanotoxins, and neurodegenerative diseases: Dangerous liaisons. Int. J. Mol. Sci. 2021, 22, 8726. [Google Scholar] [CrossRef]
- Assunção, J.; Amaro, H.M.; Lopes, G.; Tavares, T.; Malcata, F.X.; Guedes, A.C. Exploration of marine genus Chroococcidiopsis sp.: A valuable source for antioxidant industry? J. Appl. Phycol. 2021, 33, 2169–2187. [Google Scholar] [CrossRef]
- Huang, I.S.; Zimba, P.V. Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 83, 42–94. [Google Scholar] [CrossRef]
- Du, X.; Liu, H.; Yuan, L.; Wang, Y.; Ma, Y.; Wang, R.; Chen, X.; Losiewicz, M.D.; Guo, H.; Zhang, H. The diversity of cyanobacterial toxins on structural characterization, distribution and Identification: A systematic review. Toxins 2019, 11, 530. [Google Scholar] [CrossRef]
- Siegesmund, M.A.; Johansen, J.R.; Karsten, U.; Friedl, T. Coleofasciculus gen. nov. (cyanobacteria): Morphological and molecular criteria for revision of the genus Microcoleus gomont. J. Phycol. 2008, 44, 1572–1585. [Google Scholar] [CrossRef]
- Malone, C.F.d.S.; Genuário, D.B.; Vaz, M.G.M.V.; Fiore, M.F.; Sant’Anna, C.L. Monilinema gen. nov., a homocytous genus (Cyanobacteria, Leptolyngbyaceae) from saline–alkaline lakes of Pantanal wetlands, Brazil. J. Phycol. 2021, 57, 473–483. [Google Scholar] [CrossRef]
- Komárek, J. A polyphasic approach for the taxonomy of cyanobacteria: Principles and applications. Eur. J. Phycol. 2016, 51, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Temraleeva, A.D.; Dronova, S.A.; Moskalenko, S.V.; Didovich, S.V. Modern methods for isolation, purification, and cultivation of soil cyanobacteria. Microbiology 2016, 85, 389–399. [Google Scholar] [CrossRef]
- Alvarenga, D.O.; Fiore, M.F.; Varani, A.M. A metagenomic approach to cyanobacterial genomics. Front. Microbiol. 2017, 8, 809. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Huo, S.; Zhang, J.; Ma, C.; Xiao, Z.; Zhang, H.; Xi, B.; Xia, X. Metabarcoding reveals a more complex cyanobacterial community than morphological identification. Ecol. Indic. 2019, 107, 105653. [Google Scholar] [CrossRef]
- Curren, E.; Yoshida, T.; Kuwahara, V.S.; Leong, S.C.Y. Rapid profiling of tropical marine cyanobacterial communities. Reg. Stud. Mar. Sci. 2019, 25, 100485. [Google Scholar] [CrossRef]
- Marcelino, V.R.; van Oppen, M.J.H.; Verbruggen, H. Highly structured prokaryote communities exist within the skeleton of coral colonies. ISME J. 2018, 12, 300–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvy, M.; Bélières, A.; Carré, C.; Got, P.; Pagano, M.; Agogué, H.; Bec, B.; Roques, C.; Bigot, L.; Chabanet, P.; et al. Do microbial planktonic communities reflect the ecological changes of Glorieuses coral reefs (Iles Eparses, Western Indian Ocean)? Mar. Pollut. Bull. 2022, 174, 113218. [Google Scholar] [CrossRef] [PubMed]
- Nuryadi, H.; Nguyen, T.T.M.; Ito, M.; Okada, N.; Wakaoji, S.; Maruyama, T.; Nakano, Y.; Fujimura, H.; Takeyama, H.; Suda, S. A metabarcoding survey for seasonal picophytoplankton composition in two coral reefs around Sesoko Island, Okinawa, Japan. J. Appl. Phycol. 2018, 30, 3179–3186. [Google Scholar] [CrossRef]
- MacKeigan, P.W.; Garner, R.E.; Monchamp, M.-È.; Walsh, D.A.; Onana, V.E.; Kraemer, S.A.; Pick, F.R.; Beisner, B.E.; Agbeti, M.D.; da Costa, N.B.; et al. Comparing microscopy and DNA metabarcoding techniques for identifying cyanobacteria assemblages across hundreds of lakes. Harmful Algae 2022, 113, 102187. [Google Scholar] [CrossRef] [PubMed]
- Monchamp, M.E.; Spaak, P.; Domaizon, I.; Dubois, N.; Bouffard, D.; Pomati, F. Homogenization of lake cyanobacterial communities over a century of climate change and eutrophication. Nat. Ecol. Evol. 2018, 2, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Xu, X.; Bian, R.; Wang, Y.; Yu, H.; Xu, Y.; Duan, G.; Bi, L.; Chen, P.; Gao, S.; et al. Sedimentary ancient DNA metabarcoding delineates the contrastingly temporal change of lake cyanobacterial communities. Water Res. 2020, 183, 116077. [Google Scholar] [CrossRef] [PubMed]
- Liu, R. Checklist of Marine Biota of China Seas, 2008th ed.; Science Press, Academia Sinica: Beijing, China, 2008. [Google Scholar]
- Huang, B.; Ding, L. The catalogue of marine cyanobacteria of China. Guangxi Sci. 2014, 21, 580–586. [Google Scholar]
- Zubia, M.; Vieira, C.; Palinska, K.A.; Roué, M.; Gaertner, J.-C.; Zloch, I.; Grellier, M.; Golubic, S. Benthic cyanobacteria on coral reefs of Moorea Island (French Polynesia): Diversity response to habitat quality. Hydrobiologia 2019, 843, 61–78. [Google Scholar] [CrossRef]
- Gao, J.; Wu, G.; Ya, H. Review of the circulation in the Beibu Gulf, South China Sea. Cont. Shelf Res. 2017, 138, 106–119. [Google Scholar] [CrossRef]
- Li, F.; Lai, C. Investigations on Red Tide in the Sea Area of Guangxi and Countermeasures. J. Environ. Sci. Manag. 2007, 32, 76–109, (in Chineses with an English abstract). [Google Scholar]
- Chen, T.; Li, S.; Zhao, J.; Feng, Y. Uranium-thorium dating of coral mortality and community shift in a highly disturbed inshore reef (Weizhou Island, northern South China Sea). Sci. Total Environ. 2021, 752, 141866. [Google Scholar] [CrossRef]
- Mohamed, H.F.; Abd-Elgawad, A.; Cai, R.; Luo, Z.; Xu, C. The bacterial signature offers vision into the machinery of coral fitness across high-latitude coral reef in the South China Sea. Environ. Microbiol. Rep. 2022. [Google Scholar] [CrossRef]
- Yuan, J.; Li, M.; Lin, S. An Improved DNA extraction method for efficient and quantitative recovery of phytoplankton diversity in natural assemblages. PLoS ONE 2015, 10, e0133060. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muyzer, G.; Dewaal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Marcelino, V.R.; Verbruggen, H. Multi-marker metabarcoding of coral skeletons reveals a rich microbiome and diverse evolutionary origins of endolithic algae. Sci. Rep. 2016, 6, 31508. [Google Scholar] [CrossRef] [Green Version]
- Monchamp, M.-E.; Walser, J.-C.; Pomati, F.; Spaak, P. Sedimentary DNA reveals cyanobacterial community diversity over 200 years in two Perialpine Lakes. Appl. Environ. Microbiol. 2016, 82, 6472–6482. [Google Scholar] [CrossRef] [Green Version]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Pei, L.; Hu, W.; Wang, P.; Kang, J.; Mohamed, H.F.; Wang, C.; Liu, L.; Luo, Z. Morphologic and phylogenic characterization of two bloom-forming planktonic Prorocentrum (Dinophyceae) species and their potential distribution in the China Sea. Algal Res. 2022, 66, 102788. [Google Scholar] [CrossRef]
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: Chichester, UK, 1991; pp. 115–175. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [Green Version]
- Boc, A.; Diallo, A.B.; Makarenkov, V. T-REX: A web server for inferring, validating and visualizing phylogenetic trees and networks. Nucleic Acids Res. 2012, 40, W573–W579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Meriluoto, J.; Spoof, L.; Codd, G.A. Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar] [CrossRef]
- Nuryadi, H.; Suda, S. Revealing the species diversity of Neolyngbya (Cyanobacteria, Oscillatoriales) from subtropical coastal regions of Okinawa, Japan, with descriptions of Neolyngbya intertidalis sp. nov. and Neolyngbya latusa sp. nov. Phycol. Res. 2022, 70, 69–80. [Google Scholar] [CrossRef]
- Ip, Y.C.A.; Chang, J.J.M.; Oh, R.M.; Quek, Z.B.R.; Chan, Y.K.S.; Bauman, A.G.; Huang, D. Seq’ and ARMS shall find: DNA (meta)barcoding of Autonomous Reef Monitoring Structures across the tree of life uncovers hidden cryptobiome of tropical urban coral reefs. Mol. Ecol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Casero, M.C.; Velázquez, D.; Medina-Cobo, M.; Quesada, A.; Cirés, S. Unmasking the identity of toxigenic cyanobacteria driving a multi-toxin bloom by high-throughput sequencing of cyanotoxins genes and 16S rRNA metabarcoding. Sci. Total Environ. 2019, 665, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Biessy, L.; Wood, S.A.; Chinain, M.; Roué, M.; Smith, K.F. Exploring benthic cyanobacterial diversity and co-occurring potentially harmful dinoflagellates in six islands of the South Pacific. Hydrobiologia 2021, 848, 2815–2829. [Google Scholar] [CrossRef]
- Salmaso, N.; Vasselon, V.; Rimet, F.; Vautier, M.; Elersek, T.; Boscaini, A.; Donati, C.; Moretto, M.; Pindo, M.; Riccioni, G.; et al. DNA sequence and taxonomic gap analyses to quantify the coverage of aquatic cyanobacteria and eukaryotic microalgae in reference databases: Results of a survey in the Alpine region. Sci. Total Environ. 2022, 834, 155175. [Google Scholar] [CrossRef]
- Komarek, J.; Johansen, R.J.; Smarda, J.; Strunecky, O. Phylogeny and taxonomy of Synechococcus-like cyanobacteria. Fottea 2020, 20, 171–191. [Google Scholar] [CrossRef]
- Dvořák, P.; Poulíčková, A.; Hašler, P.; Belli, M.; Casamatta, D.A.; Papini, A. Species concepts and speciation factors in cyanobacteria, with connection to the problems of diversity and classification. Biodivers. Conserv. 2015, 24, 739–757. [Google Scholar] [CrossRef]
- Ning, Z.; Yu, K.; Wang, Y.; Huang, X.; Han, M.; Zhang, J. Carbon and nutrient dynamics of permeable carbonate and silicate sands adjacent to coral reefs around Weizhou Island in the northern South China Sea. Estuar. Coast. Shelf Sci. 2019, 225, 106229. [Google Scholar] [CrossRef]
- Brocke, H.J.; Polerecky, L.; de Beer, D.; Weber, M.; Claudet, J.; Nugues, M.M. Organic matter degradation drives benthic cyanobacterial mat abundance on Caribbean coral reefs. PLoS ONE 2015, 10, e0125445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Zhang, Y.; Wang, Y.; Zhang, S.; Wang, H. Spatial and seasonal variations of Cyanobacteria and their nitrogen fixation rates in Sanya Bay, South China Sea. Sci. Mar. 2008, 72, 239–251. [Google Scholar] [CrossRef]
- Hughes, D.J.; Alderdice, R.; Cooney, C.; Kühl, M.; Pernice, M.; Voolstra, C.R.; Suggett, D.J. Coral reef survival under accelerating ocean deoxygenation. Nat. Clim. Chang. 2020, 10, 296–307. [Google Scholar] [CrossRef]
- Liang, J.; Yu, K.; Wang, Y.; Huang, X.; Huang, W.; Qin, Z.; Wang, G.; Su, H.; Chen, B.; Wu, Z. Diazotroph diversity associated with scleractinian corals and Its relationships with environmental variables in the South China Sea. Front. Physiol. 2020, 11, 615. [Google Scholar] [CrossRef]
- Lesser, M.P.; Morrow, K.M.; Pankey, S.M.; Noonan, S.H.C. Diazotroph diversity and nitrogen fixation in the coral Stylophora pistillata from the Great Barrier Reef. ISME J. 2018, 12, 813–824. [Google Scholar] [CrossRef]
- Charpy, L.; Palinska, K.A.; Casareto, B.; Langlade, M.J.; Suzuki, Y.; Abed, R.M.M.; Golubic, S. Dinitrogen-fixing cyanobacteria in microbial mats of two shallow coral reef ecosystems. Microb. Ecol. 2010, 59, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Brocke, H.J.; Piltz, B.; Herz, N.; Abed, R.M.M.; Palinska, K.A.; John, U.; Haan, J.d.; de Beer, D.; Nugues, M.M. Nitrogen fixation and diversity of benthic cyanobacterial mats on coral reefs in Curaçao. Coral Reefs 2018, 37, 861–874. [Google Scholar] [CrossRef] [Green Version]
- El-Khaled, Y.C.; Roth, F.; Tilstra, A.; Rädecker, N.; Karcher, D.B.; Kürten, B.; Jones, B.H.; Voolstra, C.R.; Wild, C. In situ eutrophication stimulates dinitrogen fixation, denitrification, and productivity in Red Sea coral reefs. Mar. Ecol. Prog. Ser. 2020, 645, 55–66. [Google Scholar] [CrossRef]
- den Haan, J.; Visser, P.M.; Ganase, A.E.; Gooren, E.E.; Stal, L.J.; van Duyl, F.C.; Vermeij, M.J.A.; Huisman, J. Nitrogen fixation rates in algal turf communities of a degraded versus less degraded coral reef. Coral Reefs 2014, 33, 1003–1015. [Google Scholar] [CrossRef]
- Souza, N.R.; Metcalf, J.S. Cyanobacterial toxins and their effects on human and animal health. In Handbook of Algal Science, Technology and Medicine; Konur, O., Ed.; Academic Press: London, UK; Cambridge, MA, USA, 2020; pp. 561–574. [Google Scholar]
- Kalaitzidou, M.; Papageorgiou, K.; Theodoridis, A.; Economou, V.; Filiousis, G.; Kritas, S.; Angelidis, P.; Petridou, E. Experimental Exposure of the Mediterranean Mussels Mytilus galloprovincialis to Potentially Toxic Cyanobacteria (Synechocystis sp.) and Detection of Microcystins. Acta Vet. Eurasia 2021, 47, 10–19. [Google Scholar] [CrossRef]
- Nagle, D.G.; Paul, V.J. Chemical defense of a marine cyanobacterial bloom. J. Exp. Mar. Biol. Ecol. 1998, 225, 29–38. [Google Scholar] [CrossRef]
- Banack, S.A.; Johnson, H.E.; Cheng, R.; Cox, P.A. Production of the Neurotoxin BMAA by a Marine Cyanobacterium. Mar. Drugs 2007, 5, 180–196. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Civiello, M.; Bell, S.C.; Willis, B.L.; Bourne, D.G. Integrated approach to understanding the onset and pathogenesis of black band disease in corals. Environ. Microbiol. 2016, 18, 752–765. [Google Scholar] [CrossRef]
- Sato, Y.; Ling, E.Y.S.; Turaev, D.; Laffy, P.; Weynberg, K.D.; Rattei, T.; Willis, B.L.; Bourne, D.G. Unraveling the microbial processes of black band disease in corals through integrated genomics. Sci. Rep. 2017, 7, 40455. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Universal Prokaryotic Primers | Cyanobacteria-specific Primers | Cultured Strains | Potential Toxin [58] | Nitrogen Fixation Cyanobacteria |
|---|---|---|---|---|---|---|
| Chroococcales | Aphanothecaceae | Aphanothece | ||||
| Chroococcaceae | Chroococcus | |||||
| Cyanobacteriaceae | Cyanobacterium | Cyanobacterium | ||||
| Geminocystaceae | Geminocystis | |||||
| Microcystaceae | Gloeocapsa | |||||
| Microcystis | Microcystin/Anatoxin-a | |||||
| Chroococcidiopsidales | Chroococcidiopsidaceae | Chroococcidiopsis | ||||
| Gloeocapsopsis | ||||||
| Gloeobacterales | Gloeobacteraceae | Gloeobacter | ||||
| Nostocales | Hapalosiphonaceae | Mastigocoleus | ||||
| Nostocaceae | Anabaena | Microcystin/Cylindrospermopsins/Saxitoxins/Anatoxin-a/BMAA | ||||
| Nostoc | Nostoc | BMAA/Microcystins | Nostoc | |||
| Trichormus | Microcystins | |||||
| Rivulariaceae | Rivularia | Microcystins | ||||
| Scytonemataceae | Chakia | |||||
| Scytonema | Lyngbyatoxin/Saxitoxins | |||||
| Symphyonemataceae | Loriellopsis | |||||
| Tolypothrichaceae | Tolypothrix | Microcystins | ||||
| Oscillatoriales | Coleofasciculaceae | Anagnostidinema | ||||
| Coleofasciculus | ||||||
| Geitlerinema | Microcystin/Saxitoxins/BBD | |||||
| Cyanothecaceae | Cyanothece | |||||
| Microcoleaceae | Annamia | Microcystins | ||||
| Arthrospira | Arthrospira | Microcystins/Anatoxin-a | ||||
| Kamptonema | ||||||
| Planktothricoides | ||||||
| Oscillatoriaceae | Aerosakkonema | |||||
| Lyngbya | Lyngbya | Cylindrospermopsins/Saxitoxins/Lyngbyatoxins/Barbamides/BMAA | Lyngbya | |||
| Oscillatoria | Oscillatoria | Microcystins/Cylindrospermopsins/Anatoxin-a/Lyngbyatoxins/BMAA | ||||
| Phormidium | Phormidium | Microcystins/Anatoxin-a | ||||
| Sirenicapillariaceae | Limnoraphis | |||||
| Pleurocapsales | Dermocarpellaceae | Stanieria | ||||
| Hyellaceae | Pleurocapsa | Pleurocapsa | ||||
| Xenococcaceae | Xenococcus | |||||
| Spirulinales | Spirulinaceae | Spirulina | Spirulina | Spirulina | Microcystins | |
| Synechococcales | Acaryochloridaceae | Acaryochloris | Acaryochloris | |||
| Coelosphaeriaceae | Snowella | |||||
| Leptolyngbyaceae | Leptolyngbya | Leptolyngbya | Microcystin/BMAA/BBD | |||
| Neosynechococcus | ||||||
| Phormidesmis | Phormidesmis | |||||
| Merismopediaceae | Synechocystis | Microcystin/BMAA/LPS | ||||
| Merismopedia | Microcystin | |||||
| Prochlorococcaceae | Prochlorococcus | Prochlorococcus | ||||
| Halomicronema | Halomicronema | |||||
| Nodosilinea | ||||||
| Limnothrix | Microcystin/Saxitoxins | |||||
| Pseudanabaena | Pseudanabaena | Pseudanabaena | Microcystin/Anatoxin-a | |||
| Synechococcaceae | Synechococcus | Synechococcus | icrocystin/BMAA |
| Species | Strains | Collection Date | Latitude (N) | Longtitude (E) | Site | Source | GenBank No. (16S) |
|---|---|---|---|---|---|---|---|
| Anabaena sp. | TIOX110 | 2020.06 | 109.1003 | 21.0644 | LQ | Sediment | OP942405 |
| Chroococcus sp. | TIOX101 | 2020.06 | 109.1250 | 21.0831 | BG | Sediment | OP942406 |
| Merismopedia sp. | TIOX109 | 2020.06 | 109.1003 | 21.0644 | LQ | Water | - |
| Phormidesmis sp. | TIOX96 | 2020.06 | 109.1250 | 21.0831 | BG | Water | OP942407 |
| Phormidesmis sp. | TIOX95 | 2020.06 | 109.1250 | 21.0831 | BG | Water | - |
| Phormidesmis sp. | TIOX89 | 2020.08 | 109.1250 | 21.0831 | BG | Water | - |
| Pleurocapsa sp. | TIOX102 | 2020.06 | 109.0835 | 21.0400 | SLK | Sediment | OP942408 |
| Pseudanabaena sp. | TIOX98 | 2021.04 | 109.1003 | 21.0644 | LQ | Water | OP942409 |
| Spirulina sp. | TIOX113 | 2021.04 | 109.1003 | 21.0644 | LQ | Water | OP942410 |
| Site BG | Site LQ | Site SLK |
|---|---|---|
| Annamia | Annamia | Annamia |
| Chroococcidiopsis | Chroococcidiopsis | Chroococcidiopsis |
| Coleofasciculus | ||
| Cyanobacterium | Cyanobacterium | Cyanobacterium |
| Geitlerinema | Geitlerinema | Geitlerinema |
| Gloeocapsopsis | ||
| Kamptonema | ||
| Leptolyngbya | Leptolyngbya | Leptolyngbya |
| Limnoraphis | Limnoraphis | |
| Loriellopsis | Loriellopsis | |
| Lyngbya | Lyngbya | |
| Nodosilinea | Nodosilinea | |
| Oscillatoria | Oscillatoria | Oscillatoria |
| Phormidesmis | Phormidesmis | Phormidesmis |
| Phormidium | Phormidium | Phormidium |
| Planktothricoides | ||
| Pleurocapsa | Pleurocapsa | Pleurocapsa |
| Pseudanabaena | Pseudanabaena | Pseudanabaena |
| Richelia | Richelia | Rivularia |
| Scytonema | Scytonema | |
| Spirulina | Spirulina | Spirulina |
| Stanieria | Stanieria | Stanieria |
| Synechococcus | Synechococcus | Synechococcus |
| Synechocystis | Synechocystis | Synechocystis |
| Tolypothrix | Tolypothrix | |
| Xenococcus | Xenococcus | Xenococcus |
| Acaryochloris | Acaryochloris | |
| Aerosakkonema | Aerosakkonema | |
| Anagnostidinema | ||
| Arthrospira | Arthrospira | |
| Chakia | ||
| Cyanothece | Cyanothece | |
| Geminocystis | ||
| Gloeobacter | ||
| Gloeocapsa | ||
| Halomicronema | Halomicronema | |
| Kamptonema | ||
| Limnothrix | ||
| Microcystis | ||
| Lyngbya | ||
| Nostoc | Nostoc | |
| Prochlorococcus | Prochlorococcus | |
| Aphanothece | ||
| Mastigocoleus | ||
| Neosynechococcus | ||
| Snowella | ||
| Trichormus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.; Mohamed, H.F.; Liu, X.; Pei, L.; Huang, S.; Lin, X.; Zheng, X.; Luo, Z. Combined Culture and DNA Metabarcoding Analysis of Cyanobacterial Community Structure in Response to Coral Reef Health Status in the South China Sea. J. Mar. Sci. Eng. 2022, 10, 1984. https://doi.org/10.3390/jmse10121984
Kang J, Mohamed HF, Liu X, Pei L, Huang S, Lin X, Zheng X, Luo Z. Combined Culture and DNA Metabarcoding Analysis of Cyanobacterial Community Structure in Response to Coral Reef Health Status in the South China Sea. Journal of Marine Science and Engineering. 2022; 10(12):1984. https://doi.org/10.3390/jmse10121984
Chicago/Turabian StyleKang, Jianhua, Hala F. Mohamed, Xinming Liu, Lulu Pei, Shuhong Huang, Xiangyuan Lin, Xinqing Zheng, and Zhaohe Luo. 2022. "Combined Culture and DNA Metabarcoding Analysis of Cyanobacterial Community Structure in Response to Coral Reef Health Status in the South China Sea" Journal of Marine Science and Engineering 10, no. 12: 1984. https://doi.org/10.3390/jmse10121984