
Sneaking into a Hotspot of Biodiversity: Coverage and Integrity of a Rhodolith Bed in the Strait of Sicily (Central Mediterranean Sea)
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
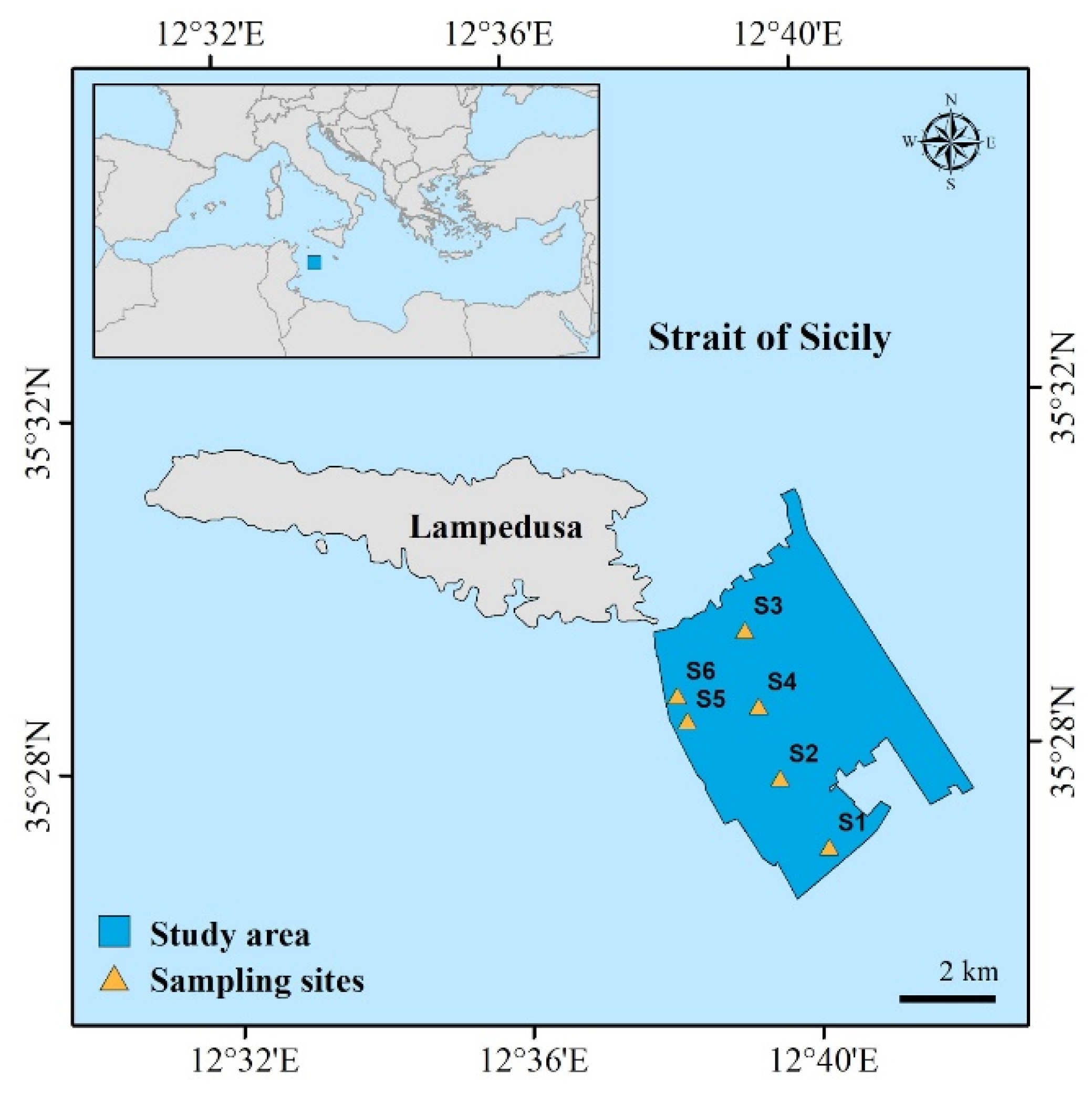
2.1. Study Area
2.2. Habitat Characterization: Analysis of Acoustic and Video Data
2.3. Biological Communities Associated with Rhodolith/Maërl Bed
3. Results
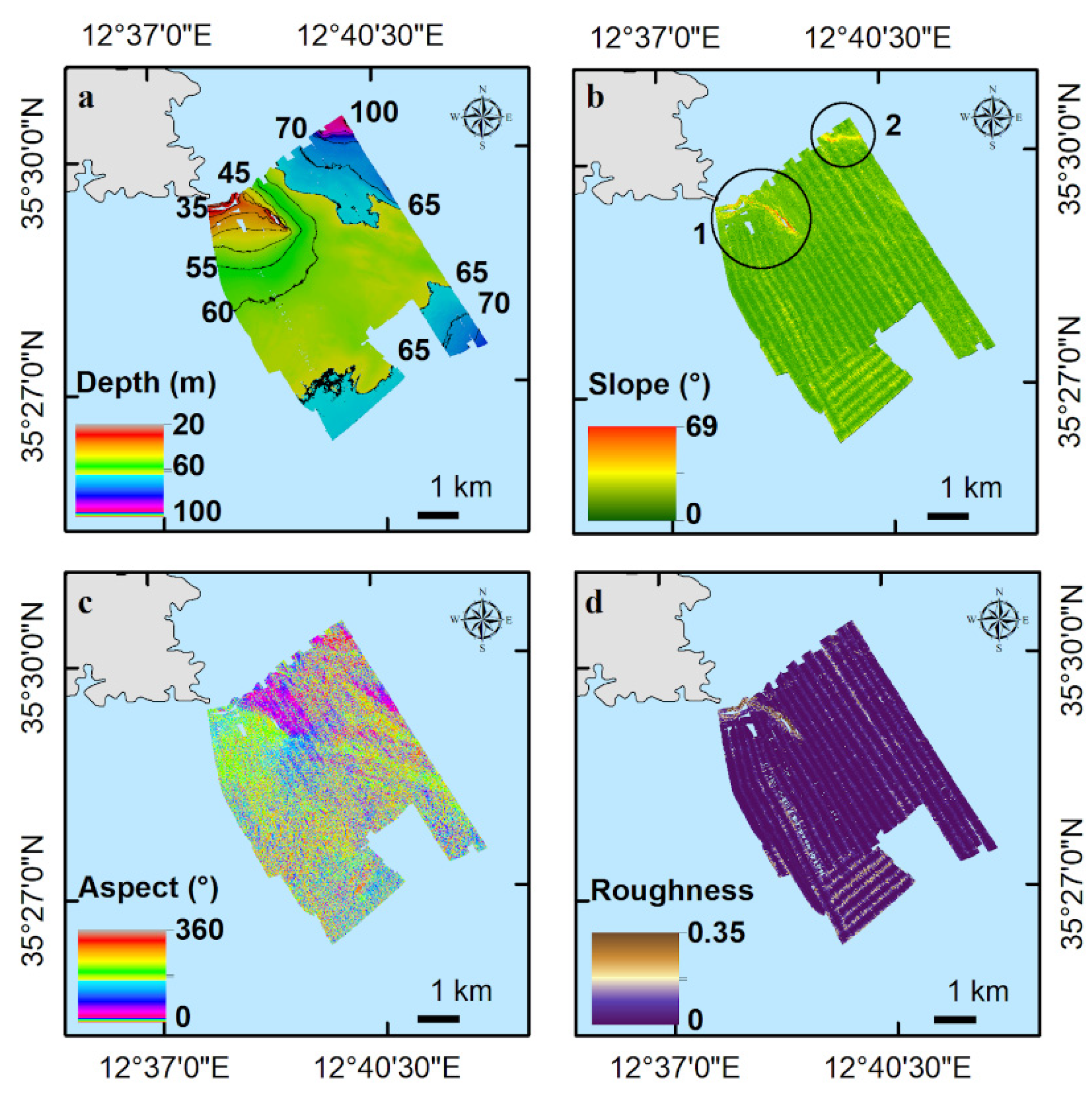
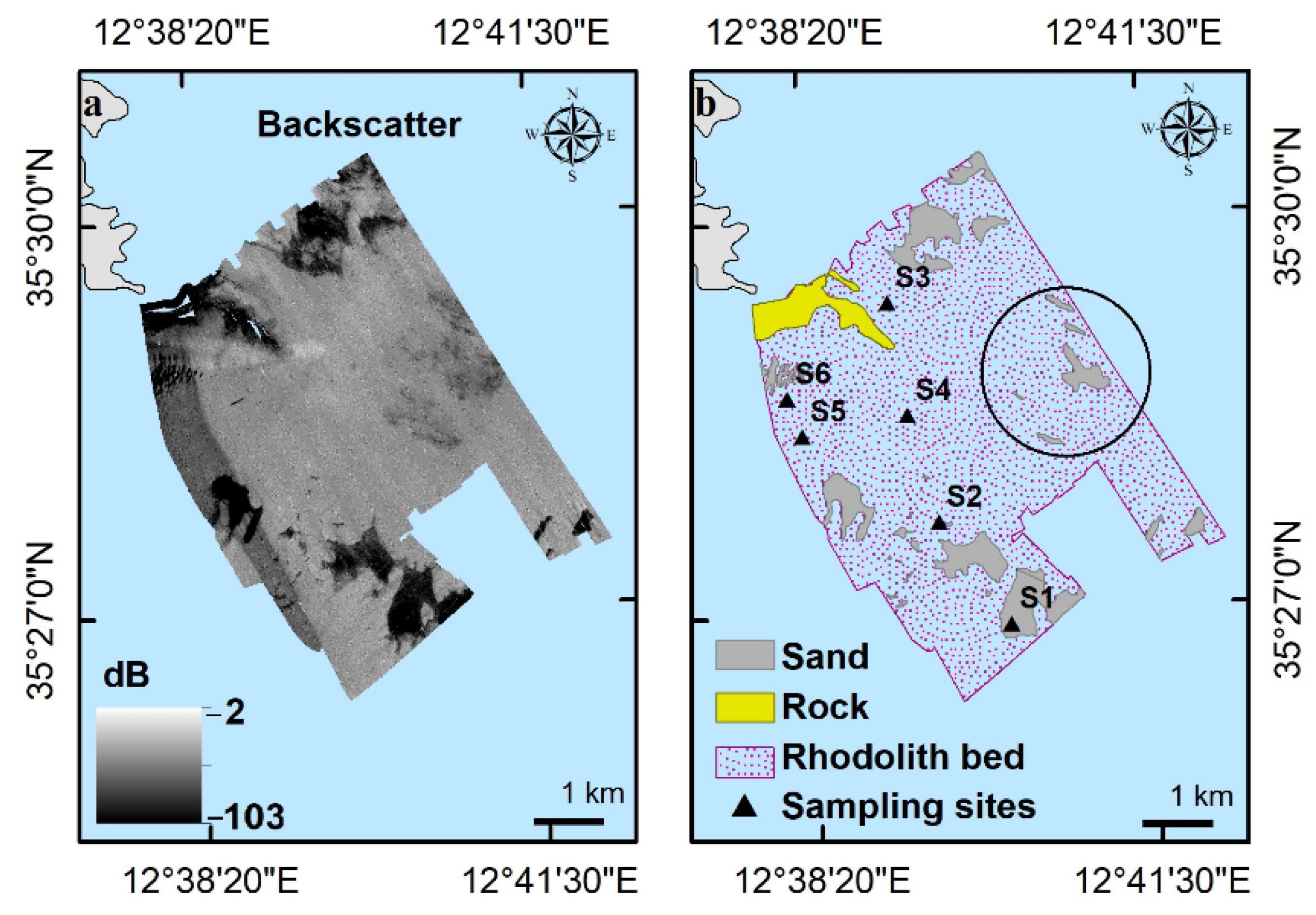
3.1. Terrain and Textural Analysis of Rhodolith/Maërl Bed
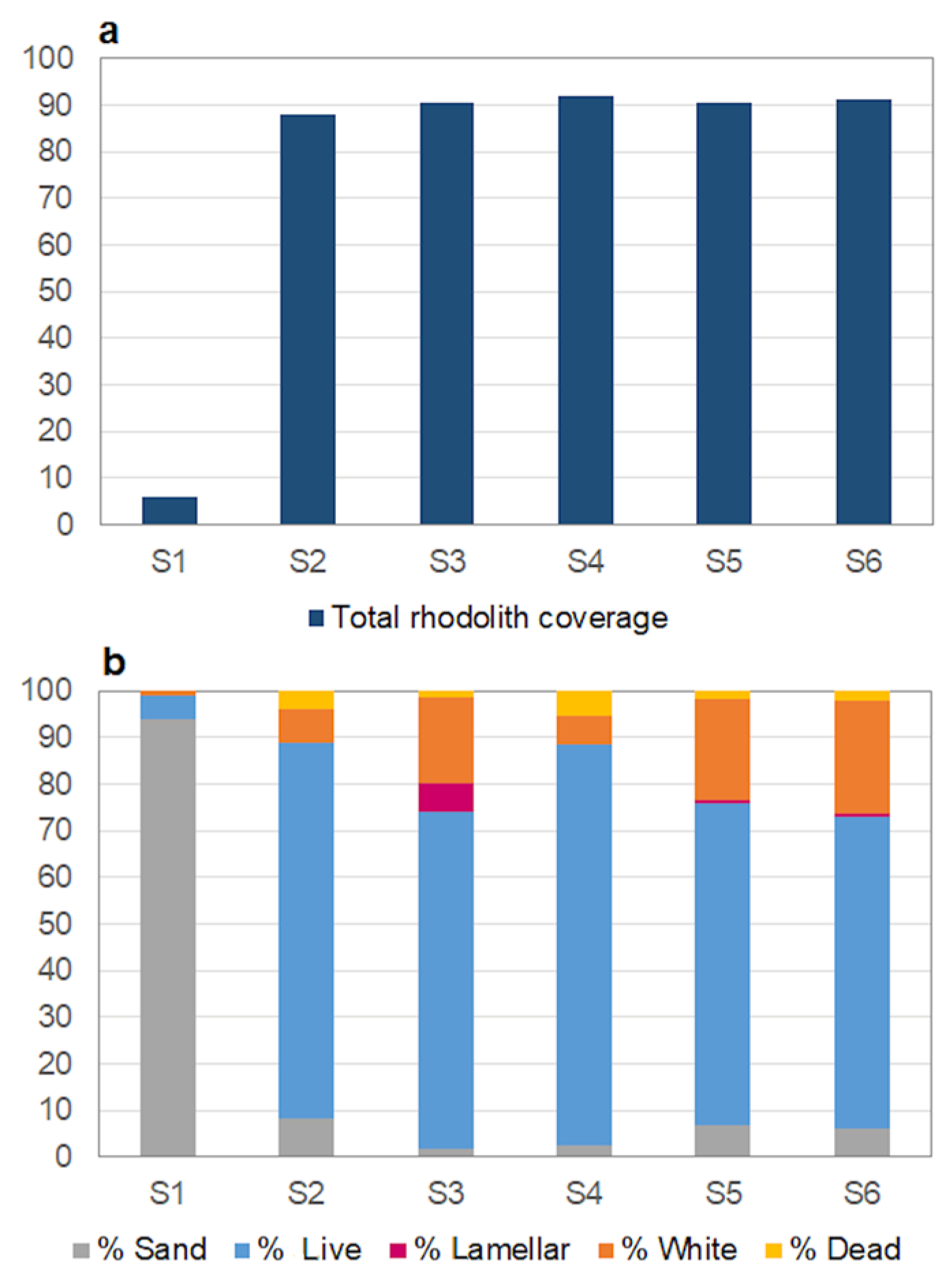
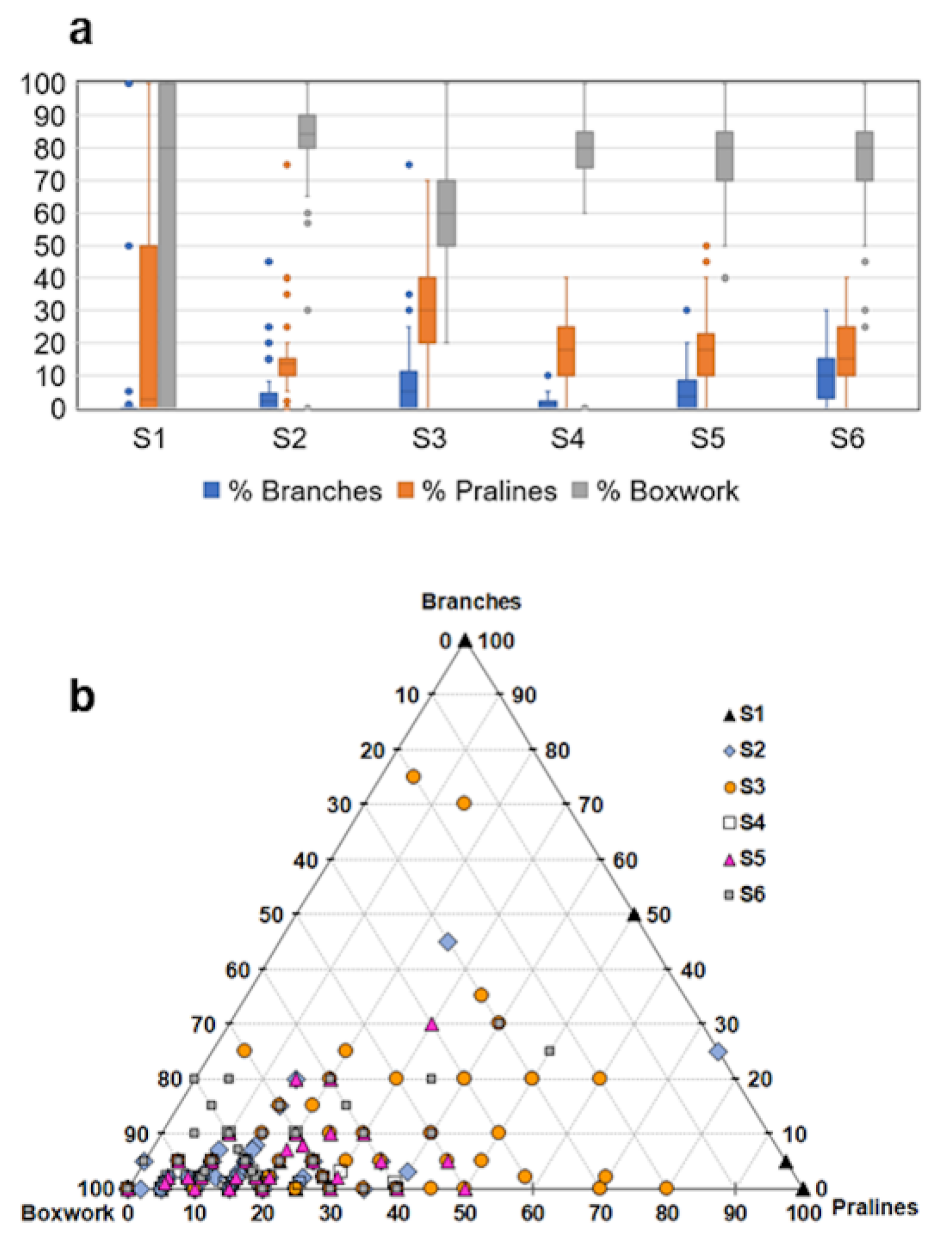
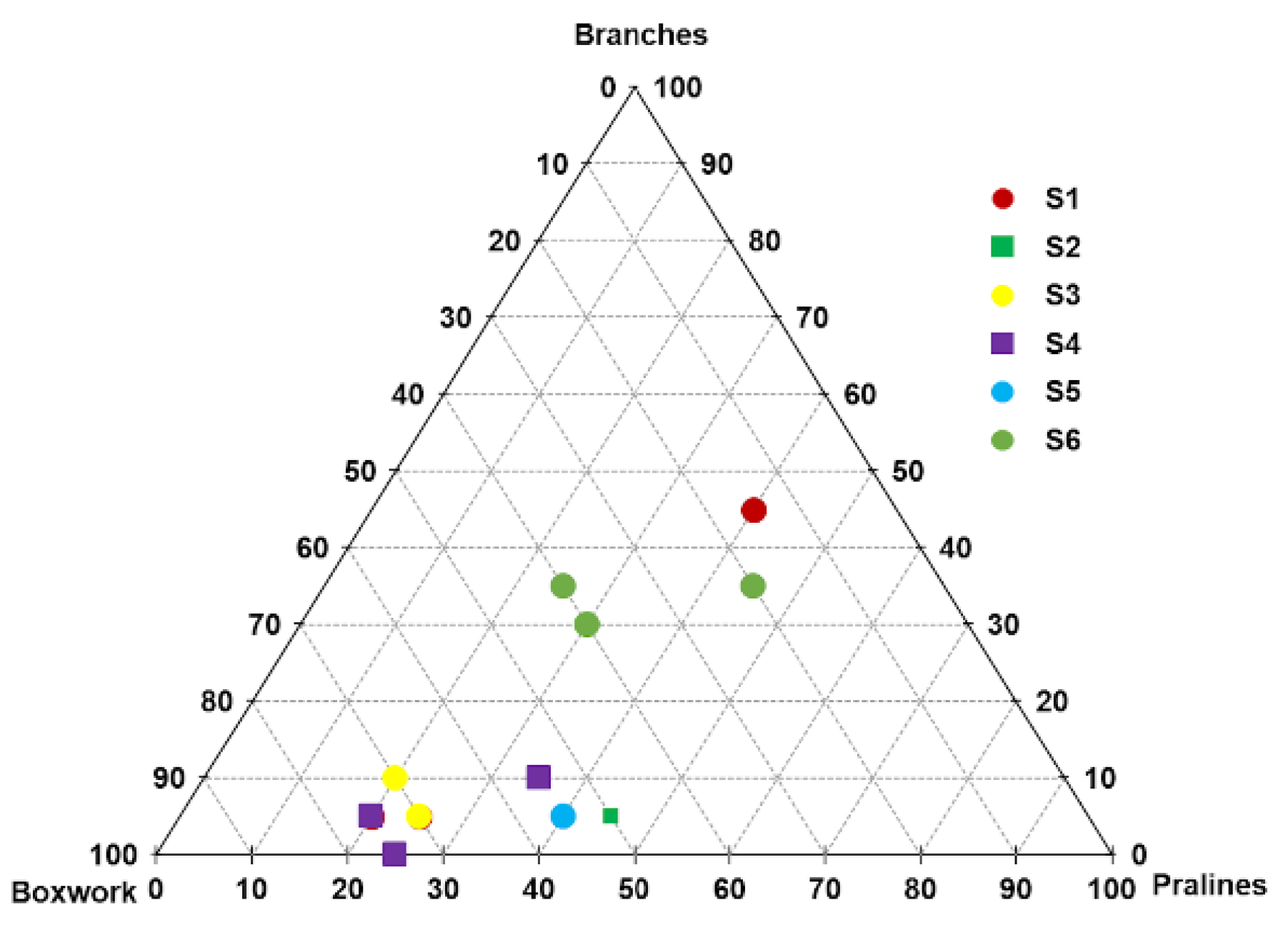
3.2. Analysis of Video Data
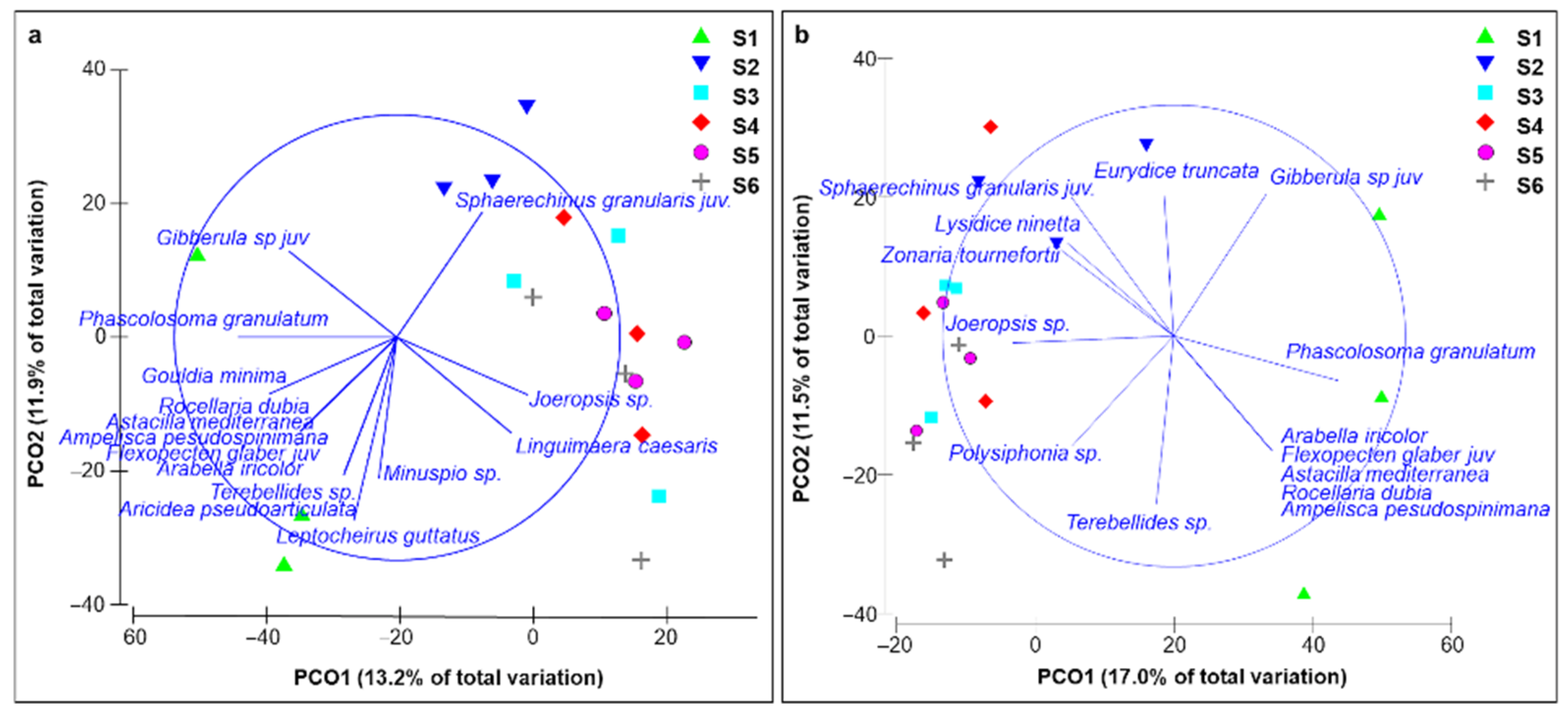
3.3. Benthic Community Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
| 1. | The United Nations officially recognised Turkey renaming as Türkiye after the government formally advocated for the new name. |
References
- Steneck, R.S. The ecology of coralline algae crusts: Convergent patterns and adaptive strategies. Annu. Rev. Ecol. Syst. 1986, 17, 273–303. [Google Scholar] [CrossRef]
- Bressan, G.; Babbini, L.; Ghirardelli, L.; Basso, D. Bio–costruzione e bio–distruzione di Corallinales nel mar Mediterraneo. Biol. Mar. Medit. 2001, 8, 131–174. [Google Scholar]
- Bosence, D.W.J. The occurrence and ecology of recent rhodoliths a review. In Coated Grains; Peryt, T.M., Ed.; Springer: Berlin/Heidelberg, Germany, 1983; pp. 225–242. [Google Scholar]
- Basso, D.; Babbini, L.; Ramos–Esplá, A.A.; Salomidi, M. Mediterranean rhodolith beds. In Rhodolith/Maërl Beds: A Global Perspective; Springer: Cham, Switzerland, 2016; pp. 281–298. [Google Scholar]
- Hall–Spencer, J. Conservation issues relating to maërl beds as habitats for molluscs. J. Conchol. 1998, 2, S271–S286. [Google Scholar]
- Jacquotte, R. Etude des fonds de maërl de Méditerranée. Rec. Trav. St. Mar. ENDOUME 1962, 26, 141–235. [Google Scholar]
- Sciberras, M.; Rizzo, M.; Mifsud, J.R.; Camilleri, K.; Borg, J.A.; Lanfranco, E.; Schembri, P.J. Habitat structure and biological characteristics of a maërl bed off the northeastern coast of the Maltese Islands (central Mediterranean). Mar. Biodivers. 2009, 39, 251–264. [Google Scholar] [CrossRef]
- Basso, D.; Babbini, L.; Kaleb, S.; Bracchi, V.; Falace, A. Monitoring deep Mediterranean rhodolith beds. Aquat. Conserv. 2016, 26, 549–561. [Google Scholar] [CrossRef] [Green Version]
- Pérès, J.; Picard, J.M. Nouveau manuel de bionomie benthique de la mer Méditerranée. Recl. Trav. St. Mar. ENDOUME 1964, 31, 1–131. [Google Scholar]
- Relini, G.; Giaccone, G. Priority habitats according to the SPA/BIO protocol (Barcelona Convention) present in Italy: Identification sheets. Biol. Mar. Medit. 2009, 16, 1–367. [Google Scholar]
- Bosence, D.W.J. Ecological studies on two unattached coralline algae from western Ireland. Paleontol. 1976, 19, 365–395. [Google Scholar]
- Foster, M.S. Rhodoliths: Between rocks and soft places. J. Phycol. 2001, 37, 659–667. [Google Scholar] [CrossRef]
- Ramos–Esplà, A.A.; Luque, A.A. Maërl bed: A fragile oasis of marine life. In The Seas of Spain; Ministero del Medio ambiente y medio rural y marino: Madrid, Spain, 2008; pp. 273–290. [Google Scholar]
- Hall–Spencer, J.M.; Grall, J.; Moore, P.G.; Atkinson, R.J.A. Bivalve fishing and maërl–bed conservation in France and the UK–retrospect and prospect. Aquat. Conserv. 2003, 13, S33–S41. [Google Scholar] [CrossRef]
- Massutì, E.; Renones, O.; Carbonell, A.; Oliver, P. Demersal fish communities exploited on the continental shelf and slope off Majorca (Balearic Islands, NW Mediterranean). Vie Milieu 1996, 46, 45–55. [Google Scholar]
- Foster, M.; Filho, S.; Amado, G.M.; Kamenos, N.A.; Riosmena–Rodriguez, R.; Steller, D.L. Rhodoliths and rhodolith beds. In Research and Discoveries: The Revolution of Science Through SCUBA; Lang, M.A., Marinelli, Roberta, L., Roberts, Susan, J., Taylor, P.R., Eds.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2013; Smithsonian contributions to the marine sciences. [Google Scholar]
- Blake, C.; Maggs, C.A. Comparative growth rates and internal banding periodicity of maërl species (Corallinales, Rhodophyta) from northern Europe. Phycologia 2003, 42, 606–612. [Google Scholar] [CrossRef]
- Martin, C.S.; Giannoulaki, M.; De Leo, F.; Scardi, M.; Salomidi, M.; Knitweiss, L.; Pace, M.L.; Garofalo, G.; Gristina, M.; Ballesteros, E.; et al. Coralligenous and maërl habitats: Predictive modelling to identify their spatial distributions across the Mediterranean Sea. Sci. Rep. 2014, 4, 5073. [Google Scholar] [CrossRef] [Green Version]
- Barbera, C.; Bordehore, C.; Borg, J.A.; Glémarec, M.; Grall, J.; Hall–Spencer, J.M.; De la Huz, C.H.; Lanfranco, E.; Lastra, M.; Moore, P.G.; et al. Conservation and management of northeast Atlantic and Mediterranean maërl beds. Aquat. Conserv. 2003, 13, S65–S76. [Google Scholar] [CrossRef]
- Borg, J.A.; Howege, H.M.; Lanfranco, E.; Micallef, S.A.; Mifsud, C.; Schembri, P.J. The macrobenthic species of the infralittoral to circalittoral transition zone off the northeastern coast of Malta (Central Mediterranean). Xjenza 1998, 3, 16–24. [Google Scholar]
- Consoli, P.; Esposito, V.; Falautano, M.; Battaglia, P.; Castriota, L.; Romeo, T.; Sinopoli, M.; Vivona, P.; Andaloro, F. The impact of fisheries on vulnerable habitats: The case of trawling on circa–littoral grounds in the Strait of Sicily (central Mediterranean Sea). Mar. Biol. Res. 2017, 13, 1084–1094. [Google Scholar] [CrossRef]
- Farriols, M.T.; Irlinger, C.; Ordines, F.; Palomino, D.; Marco–Herrero, E.; Soto–Navarro, J.; Jordà, G.; Mallol Martínez, S.; Díaz, D.; Carreño, N.; et al. Recovery Signals of Rhodoliths Beds since Bottom Trawling Ban in the SCI Menorca Channel (Western Mediterranean). Diversity 2022, 14, 20. [Google Scholar] [CrossRef]
- United Nations Environmental Programme. Mediterranean Action Plan. In Action Plan for the Conservation of the Coralligenous and Other Calcareous Bio-Concretions in the Mediterranean Sea; European Environment Agency: Athens, Greece, 2017. [Google Scholar]
- European Parliament. Council directive 1992/43/EC Conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, L206, 7–50. [Google Scholar]
- European Parliament. Council Regulation No. 1967/2006 concerning management measures for the sustainable exploitation of fishery resources in the Mediterranean Sea, amending Regulation (EEC) No 2847/93 and repealing Regulation (EC) No 1626/94. Off. J. Eur. Union 2006, L409, 11. [Google Scholar]
- European Parliament. Council of the European Union Directive 2008/56/EC of the European Parliament and of the Council of 17 June 2008 establishing a framework for Community action in the field of marine environmental policy (Marine Strategy Framework Directive). Off. J. Eur. Union 2008, L164, 19. [Google Scholar]
- Bracchi, V.A.; Angeletti, L.; Marchese, F.; Taviani, M.; Cardone, F.; Hajdas, I.; Grande, V.; Prampolini, M.; Caragnano, A.; Corselli, C.; et al. A resilient deep–water rhodolith bed off the Egadi Archipelago (Mediterranean Sea) and its actual paleontological significance. Alp. Mediterr. Quat. 2019, 32, 1–20. [Google Scholar]
- Deidun, A.; Marrone, A.; Gauci, A.; Galdies, J.; Lorenti, M.; Mangano, M.C.; Cutajar, K.; Mirto, S.; Sarà, G. Structure and biodiversity of a Maltese maërl bed: New insight into the associated assemblage 24 years after the first investigation. Reg. Stud. Mar. Sci. 2022, 52, 102262. [Google Scholar] [CrossRef]
- Rendina, F.; Kaleb, S.; Caragnano, A.; Ferrigno, F.; Apolloni, L.; Donnarumma, L.; Russo, G.F.; Sandulli, R.; Roviello, V.; Falace, A. Distribution and Characterization of Deep Rhodolith Beds off the Campania Coast (SW Italy, Mediterranean Sea). Plants 2020, 9, 985. [Google Scholar] [CrossRef]
- Innangi, S.; Tonielli, R.; Romagnoli, C.; Budillon, F.; Di Martino, G.; Innangi, M.; Laterza, R.; Le Bas, T.; Lo Iacono, C. Seabed mapping in the Pelagie Islands marine protected area (Sicily Channel, southern Mediterranean) using Remote Sensing Object Based Image Analysis (RSOBIA). Mar. Geophys. Res. 2019, 40, 333–355. [Google Scholar] [CrossRef] [Green Version]
- Ministero dell’Ambiente e della Tutela del Territorio e del Mare. Scheda metodologica modulo 8 habitat fondi a Maërl/Rodoliti. In Programmi di Monitoraggio per la Strategia Marina Art. 11, D.lgs. 190/2010; U.O. Marine Strategy: Rome, Italy, 2016. [Google Scholar]
- Steller, D.L.; Riosmena–Rodriguez, R.; Foster, M.S.; Roberts, C.A. Rhodolith bed diversity in the Gulf of California: The importance of rhodolith structure and consequences of disturbance. Aquat. Conserv. 2003, 13, S5–S20. [Google Scholar] [CrossRef]
- Bahia, R.G.; Abrantes, D.P.; Brasileiro, P.S.; Pereira–Filho, G.H.; Amado–Filho, G.M. Rhodolith bed structure along a depth gradient on the northern coast of Bahia State, Brazil. Braz. J. Oceanog. 2010, 58, 323–337. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER–E: Plymouth, UK, 2008. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER–E: Plymouth, UK, 2006. [Google Scholar]
- Di Lorenzo, M.; Sinerchia, M.; Colloca, F. The North sector of the Strait of Sicily: A priority area for conservation in the Mediterranean Sea. Hydrobiologia 2018, 821, 235–253. [Google Scholar] [CrossRef]
- Garofalo, G.; Fortibuoni, T.; Gristina, M.; Sinopoli, M.; Fiorentino, F. Persistence and co-occurrence of demersal nurseries in the Strait of Sicily (central Mediterranean): Implications for fishery management. J. Sea Res. 2011, 66, 29–38. [Google Scholar] [CrossRef]
- Clusa, M.; Carreras, C.; Pascual, M.; Gaughran, S.J.; Piovano, S.; Giacoma, C.; Fernández, G.; Levy, Y.; Tomás, J.; Raga, J.A.; et al. Fine-scale distribution of juvenile Atlantic and Mediterranean loggerhead turtles (Caretta caretta) in the Mediterranean Sea. Mar. Biol. 2014, 161, 509–519. [Google Scholar]
- Bracchi, V.A.; Caronni, S.; Meroni, A.N.; Burguett, E.G.; Atzori, F.; Cadoni, N.; Marchese, F.; Basso, D. Morphostructural Characterization of the Heterogeneous Rhodolith Bed at the Marine Protected Area “Capo Carbonara” (Italy) and Hydrodynamics. Diversity 2022, 14, 51. [Google Scholar] [CrossRef]
- Chimienti, G.; Rizzo, L.; Kaleb, S.; Falace, A.; Fraschetti, S.; De Giosa, F.; Tursi, A.; Barbone, E.; Ungaro, N.; Mastrototaro, F. Rhodolith Beds Heterogeneity along the Apulian Continental Shelf (Mediterranean Sea). J. Mar. Sci. Eng. 2020, 8, 813. [Google Scholar] [CrossRef]
- Basso, D. Deep rhodolith distribution in the Pontian Islands, Italy: A model for the paleoecology of a temperate sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1998, 137, 173–187. [Google Scholar] [CrossRef]
- Teichert, S.; Woelkerling, W.; Rüggeberg, A.; Wisshak, M.; Piepenburg, D.; Meyerhöfer, M.; Form, A.; Büdenbender, J.; Freiwald, A. Rhodolith beds (Corallinales, Rhodophyta) and their physical and biological environment at 80°31′N in Nordkappbukta (Nordaustlandet, Svalbard Archipelago, Norway). Phycologia 2012, 5, 371–390. [Google Scholar] [CrossRef]
- Wilson, S.; Blake, C.; Berges, J.A.; Maggs, C.A. Environmental tolerances of free–living coralline algae (maërl): Implications for European marine conservation. Biol. Conserv. 2004, 120, 279–289. [Google Scholar] [CrossRef]
- Basso, D. Phytobenthic communities in the circalittoral soft bottoms of the Tyrrhenian sea (Mediterranean). Atti 10 Congr. Soc. Ital. Oceanol. Limnol. 1992, 563–573. [Google Scholar]
- Di Geronimo, I.; Giaccone, G. Le alghe calcaree nel Detritico Costiero di Lampedusa (isole Pelagie). Boll. Accad. Gioenia Sci. Nat. Catania 1994, 27, 5–25. [Google Scholar]
- Mannino, A.M.; Castriota, L.; Beltrano, A.M.; Sunseri, G. The epiflora of a rhodolith bed from the Island of Ustica (Southern Tyrrhenian Sea). Flora Mediterr. 2002, 12, 11–28. [Google Scholar]
- Nelson, W.A. Calcified macroalgae–critical to coastal ecosystems and vulnerable to change: A review. Mar. Freshw. Res. 2009, 60, 787–801. [Google Scholar] [CrossRef]
- Steller, D.L.; Foster, M.S. Environmental factors influencing distribution and morphology of rhodoliths in Bahía Concepción, BCS, México. J. Exp. Mar. Biol Ecol. 1995, 194, 201–212. [Google Scholar] [CrossRef]
- Brenchley, G.A. Disturbance and community structure: An experimental approach. J. Mar. Res. 1981, 39, 767–790. [Google Scholar]
- Sousa, W.P. The Role of Disturbance in Natural Communities. Annu. Rev. Ecol. Evol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Bordehore, C.; Ramos–Esplá, A.A.; Riosmena–Rodríguez, R. Comparative study of two maërl beds with different otter trawling history, southeast Iberian Peninsula. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, S43–S54. [Google Scholar] [CrossRef]
- Harvey, A.S.; Bird, F.L. Community structure of a rhodolith bed from cold–temperate waters (southern Australia). Aust. J. Bot. 2008, 56, 437–450. [Google Scholar] [CrossRef]
- Grall, J.; Hall–Spencer, J.M. Problems facing maërl conservation in Brittany. Aquat. Conserv. 2003, 13, 55–64. [Google Scholar] [CrossRef]
- Horta, P.A.; Riul, P.; Amado Filho, G.M.; Gurgel, C.F.D.; Berchez, F.; Nunes, J.M.D.C.; Scherner, F.; Pereira, S.; Lotufo, T.; Peres, L.; et al. Rhodoliths in Brazil: Current knowledge and potential impacts of climate change. Braz. J. Oceanogr. 2016, 64, 117–136. [Google Scholar] [CrossRef] [Green Version]
- Peña, V.; Bárbara, I.; Grall, J.; Maggs, C.A.; Hall–Spencer, J.M. The diversity of seaweeds on maërl in the NE Atlantic. Mar. Biodivers. 2014, 44, 533–551. [Google Scholar] [CrossRef]
- Riosmena–Rodríguez, R.; Nelson, W.; Aguirre, J. Rhodolith/maërl beds: A global perspective. In Coastal Research Library; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Scherner, F.; Riul, P.; Bastos, E.; Bouzon, Z.L.; Pagliosa, P.R.; Blankensteyn, A.; Oliveira, E.C.; Horta, P.A. Herbivory in a rhodolith bed: A structuring factor? Panam. J. Aquat. Sci. 2010, 5, 358–366. [Google Scholar]
- Picard, J. Recherches qualitatives sur les biocoenoses marines des substrats meubles dragables de la Région Marseillaise. Recl. Trav. Stn. Mar. D’Endoume 1965, 52, 1–160. [Google Scholar]
- Bakir, K.; Sezgin, M.; Katagan, T. Alien amphipods on the Turkish coasts. Zool. Baetica 2010, 21, 191–196. [Google Scholar]
- Ben Souissi, J.; Kahri, C.; Ben Salem, M.; Zaouali, J. Les espèces non indigènes du macrobenthos des lagunes du sud–est tunisien: Point sur la situation. Rapp. Comm. Int. Pour L’exploration Sci. Mer Méditerranée 2010, 39, 449. [Google Scholar]
- Krapp–Schickel, T. Linguimaera Pirlot, 1936 (Crustacea, Amphipoda, Melitidae), a valid genus. Mem. Mus. Vic. 2003, 60, 257–283. [Google Scholar] [CrossRef] [Green Version]
- Kocatas, A.; Katagan, T.; Benli, H.A. Contribution to the knowledge of the Crustacean fauna of Cyprus. Isr. J. Zool. 2001, 47, 147–160. [Google Scholar] [CrossRef]
- Ortiz, M.; Petrescu, I. The marine Amphipoda (Crustacea: Gammaridae) of the Republic of Libya Southern Mediterranean. Trav. Muséum Natl. D’histoire Nat. Grigore Antipa 2007, 50, 11–23. [Google Scholar]
- Tsiamis, K.; Palialexis, A.; Stefanova, K.; Gladan, Ž.N.; Skejić, S.; Despalatović, M.; Cvitković, I.; Dragičević, B.; Dulčić, J.; Vidjak, O.; et al. Non–indigenous species refined national baseline inventories: A synthesis in the context of the European Union’s Marine Strategy Framework Directive. Mar. Pollut. Bull. 2019, 145, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Lejeusne, C.; Chevaldonné, P.; Pergent–Martini, C.; Boudouresque, C.F.; Pérez, T. Climate change effects on a miniature ocean: The highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 2010, 25, 250–260. [Google Scholar] [CrossRef]
- Castriota, L.; Agamennone, F.; Sunseri, G. The mollusc community associated with maërl beds of Ustica Island (Tyrrhenian Sea). Cah. Biol. Mar. 2005, 46, 289–297. [Google Scholar]
- De Grave, S. The influence of sedimentary heterogeneity on within maërl bed differences in infaunal crustacean community. Estuar. Coast. Shelf Sci. 1999, 49, 153–163. [Google Scholar] [CrossRef]
- De Juan, S.; Lo Iacono, C.; Demestre, M. Benthic habitat characterisation of soft–bottom continental shelves: Integration of acoustic surveys, benthic samples and trawling disturbance intensity. Estuar. Coast. Shelf Sci. 2013, 117, 199–209. [Google Scholar] [CrossRef]
- Riera, R.; Delgado, J.D.; Rodrıguez, M.; Monterroso, O.; Ramos, E. Macrofaunal communities of threatened subtidal maërl seabeds on Tenerife (Canary Islands, north–east Atlantic Ocean) in summer. Acta Oceanol. 2012, 31, 98–105. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | S | N * | D * | J’ * | H’ * | D * |
|---|---|---|---|---|---|---|
| S1 | 29 | 42 | 0.96 | 0.93 | 3.10 | 7.22 |
| S2 | 53 | 89 | 0.98 | 0.94 | 3.66 | 10.69 |
| S3 | 100 | 190 | 0.98 | 0.93 | 4.13 | 15.82 |
| S4 | 83 | 170 | 0.98 | 0.93 | 3.97 | 13.44 |
| S5 | 99 | 198 | 0.97 | 0.90 | 3.95 | 14.75 |
| S6 | 90 | 186 | 0.98 | 0.91 | 3.90 | 13.40 |
| Trophic Group | N Species | N Species % | N Individuals | N Individuals % |
|---|---|---|---|---|
| Grazer | 9 | 5.3 | 48 | 5.8 |
| Suspension feeder | 34 | 20.0 | 118 | 14.2 |
| Deposit feeder | 51 | 30.0 | 262 | 31.6 |
| Deposit/suspension feeder | 5 | 2.9 | 78 | 9.4 |
| Omnivorous | 12 | 7.1 | 88 | 10.6 |
| Predator | 43 | 25.3 | 188 | 22.6 |
| unknown | 15 | 8.8 | 47 | 5.7 |
| other | 1 | 0.6 | 1 | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maggio, T.; Perzia, P.; Pazzini, A.; Campagnuolo, S.; Falautano, M.; Mannino, A.M.; Allegra, A.; Castriota, L. Sneaking into a Hotspot of Biodiversity: Coverage and Integrity of a Rhodolith Bed in the Strait of Sicily (Central Mediterranean Sea). J. Mar. Sci. Eng. 2022, 10, 1808. https://doi.org/10.3390/jmse10121808
Maggio T, Perzia P, Pazzini A, Campagnuolo S, Falautano M, Mannino AM, Allegra A, Castriota L. Sneaking into a Hotspot of Biodiversity: Coverage and Integrity of a Rhodolith Bed in the Strait of Sicily (Central Mediterranean Sea). Journal of Marine Science and Engineering. 2022; 10(12):1808. https://doi.org/10.3390/jmse10121808
Chicago/Turabian StyleMaggio, Teresa, Patrizia Perzia, Alfredo Pazzini, Silvana Campagnuolo, Manuela Falautano, Anna Maria Mannino, Alessandro Allegra, and Luca Castriota. 2022. "Sneaking into a Hotspot of Biodiversity: Coverage and Integrity of a Rhodolith Bed in the Strait of Sicily (Central Mediterranean Sea)" Journal of Marine Science and Engineering 10, no. 12: 1808. https://doi.org/10.3390/jmse10121808