Fibrosis Distinguishes Critical Limb Ischemia Patients from Claudicants in a Transcriptomic and Histologic Analysis

 ,
,
Abstract
:1. Introduction
2. Experimental Section
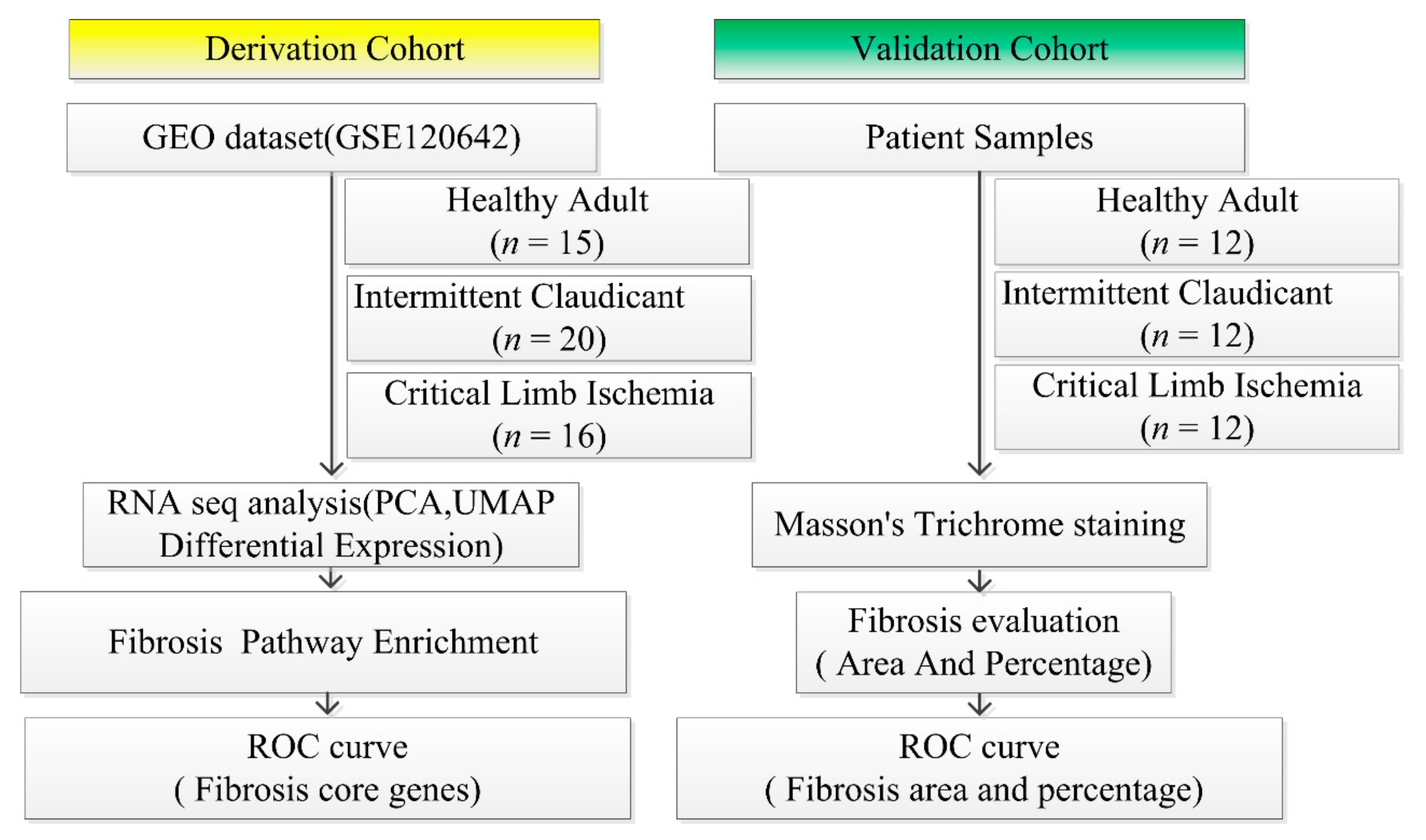
2.1. Study Design
2.2. Retrieval of Derivation Cohort Datasets of Patients from the GEO
2.3. Bioinformatics Analysis
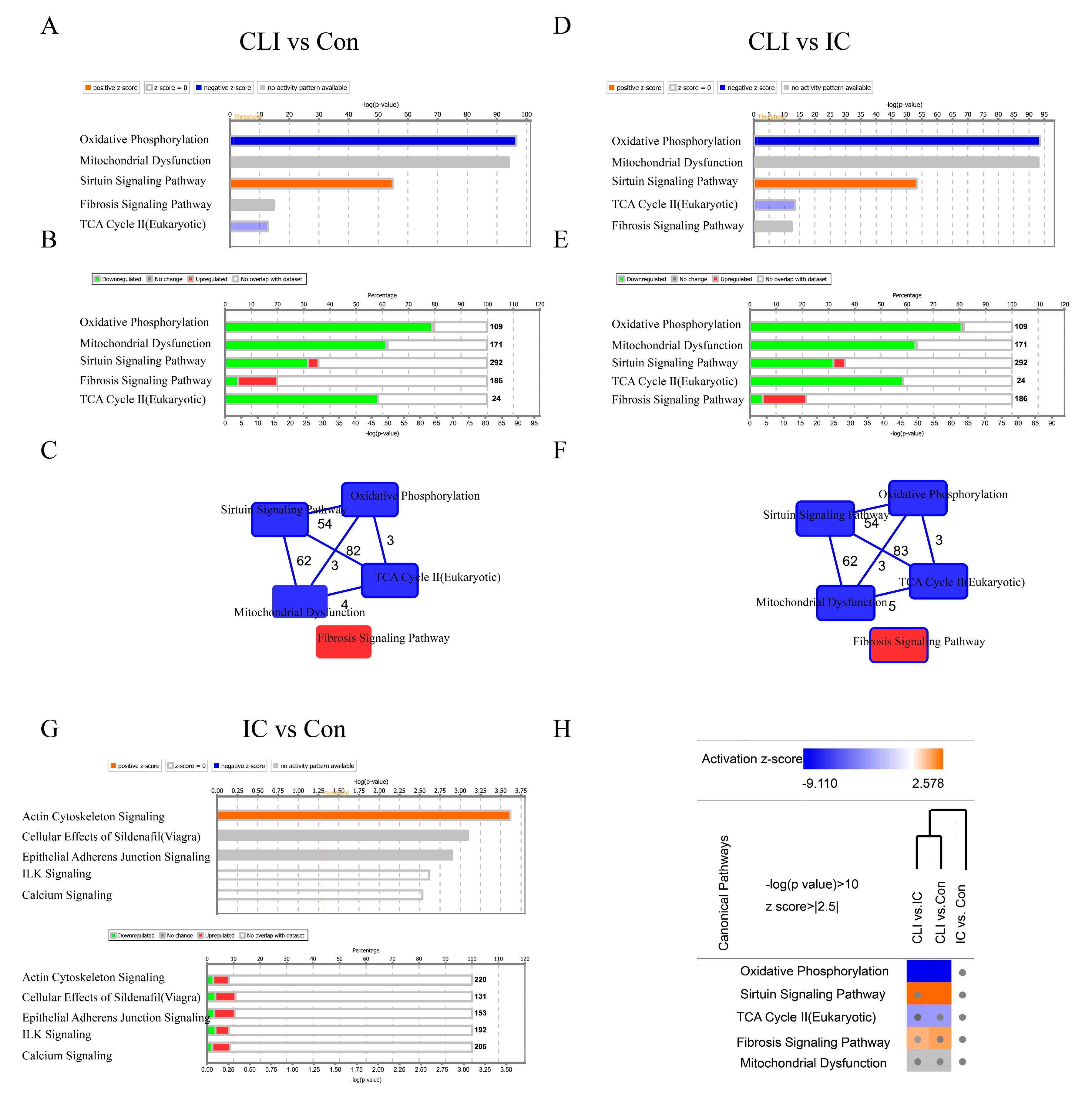
2.4. Pathway Enrichment Analysis
2.5. Fibrosis Core Gene Expression Analysis
2.6. Validation Cohort Data
2.7. Ethical Compliance
2.8. Fibrosis Evaluation in Validation Cohort
2.9. ROC Analysis Both in Derivation and Validation Cohort
2.10. Statistics for Clinical Data
3. Results
3.1. Subjects’ Age and ABI Derivation and Validation Cohort
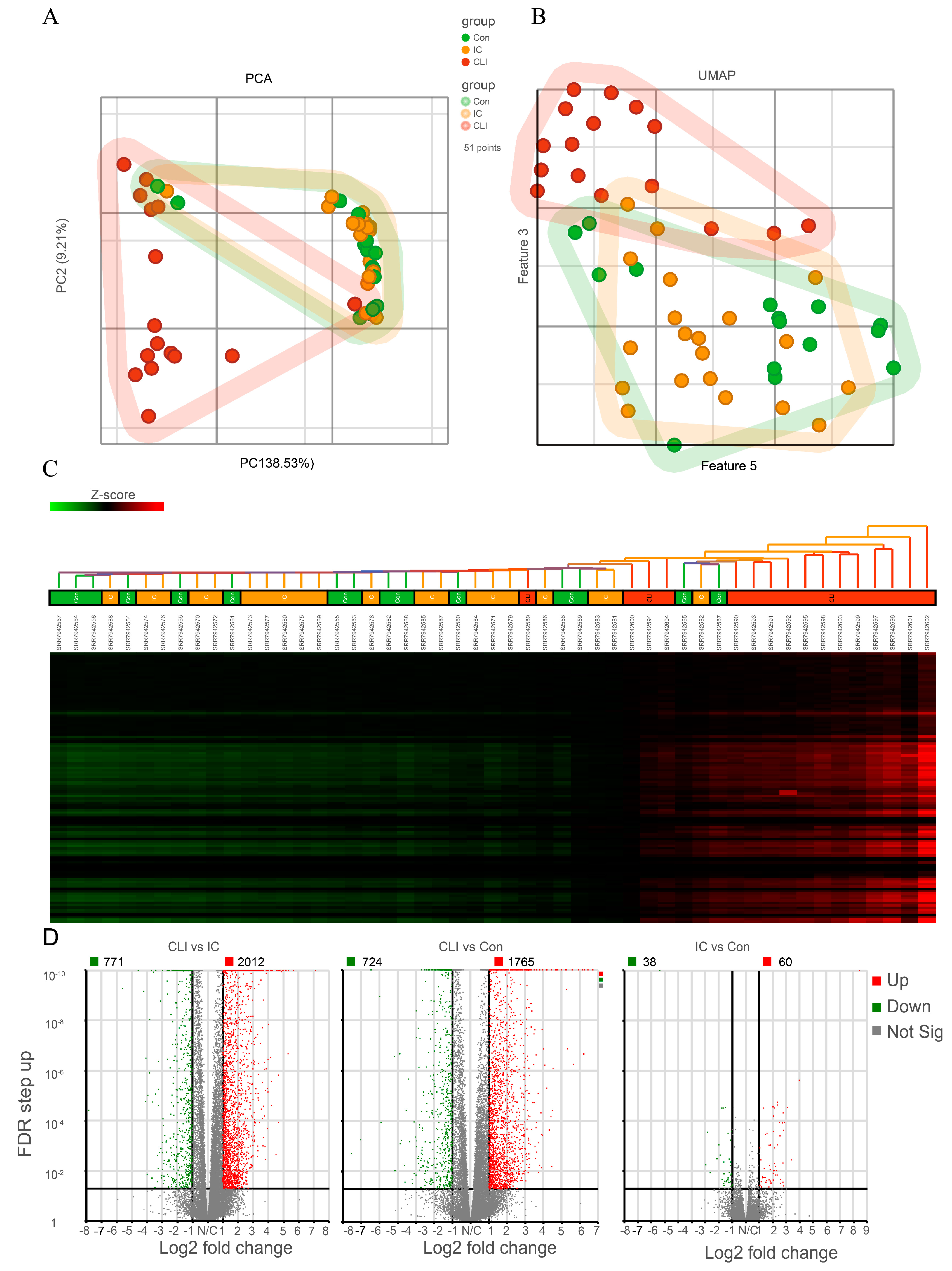
3.2. CLI Patients Have a Unique Gene Expression Profile Compared to Claudicants and Non-PAD Controls Subjects
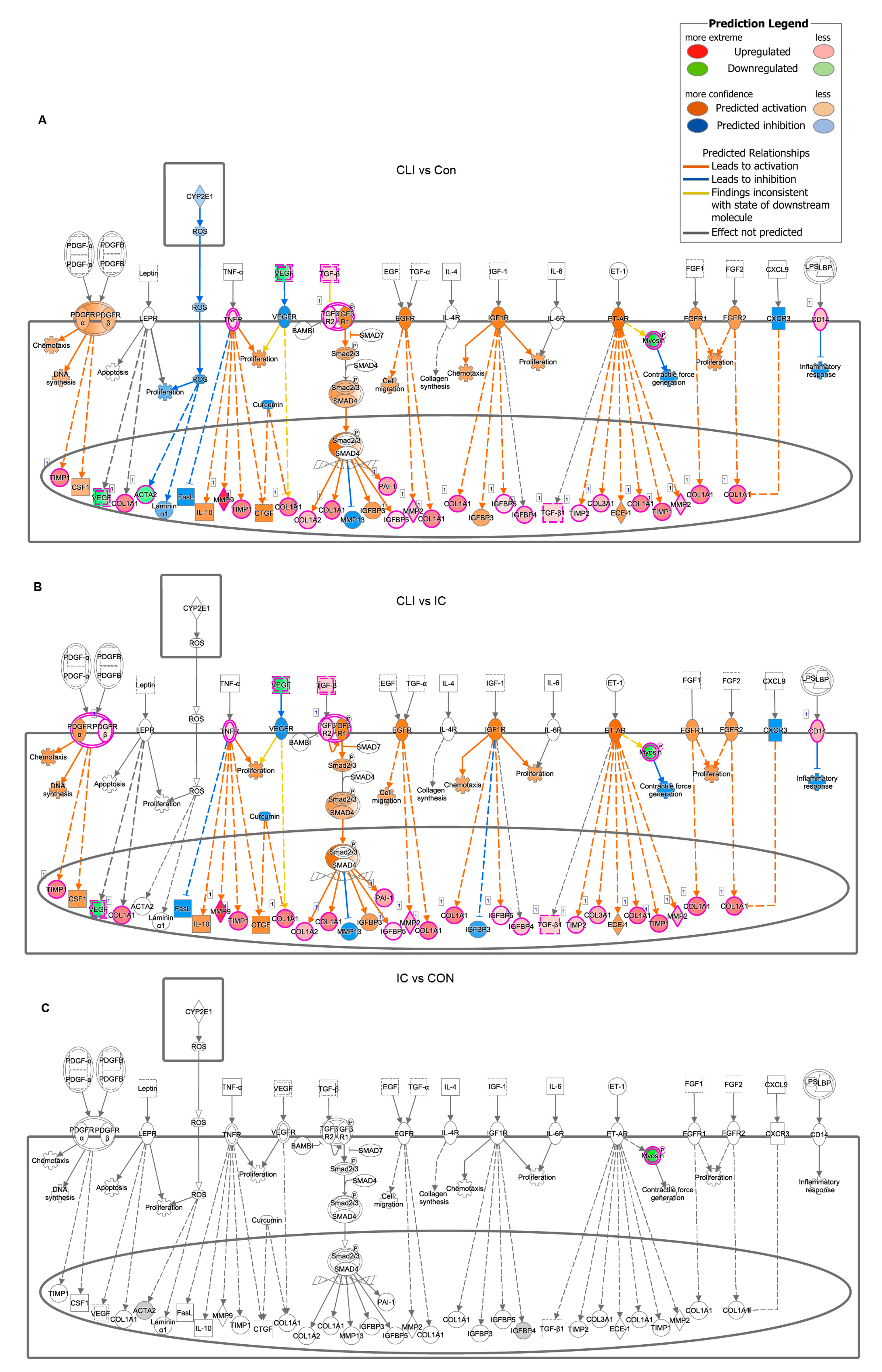
3.3. Differentially Expressed Genes (DEG) and Canonical Pathway Analysis Distinguishes CLI Patients
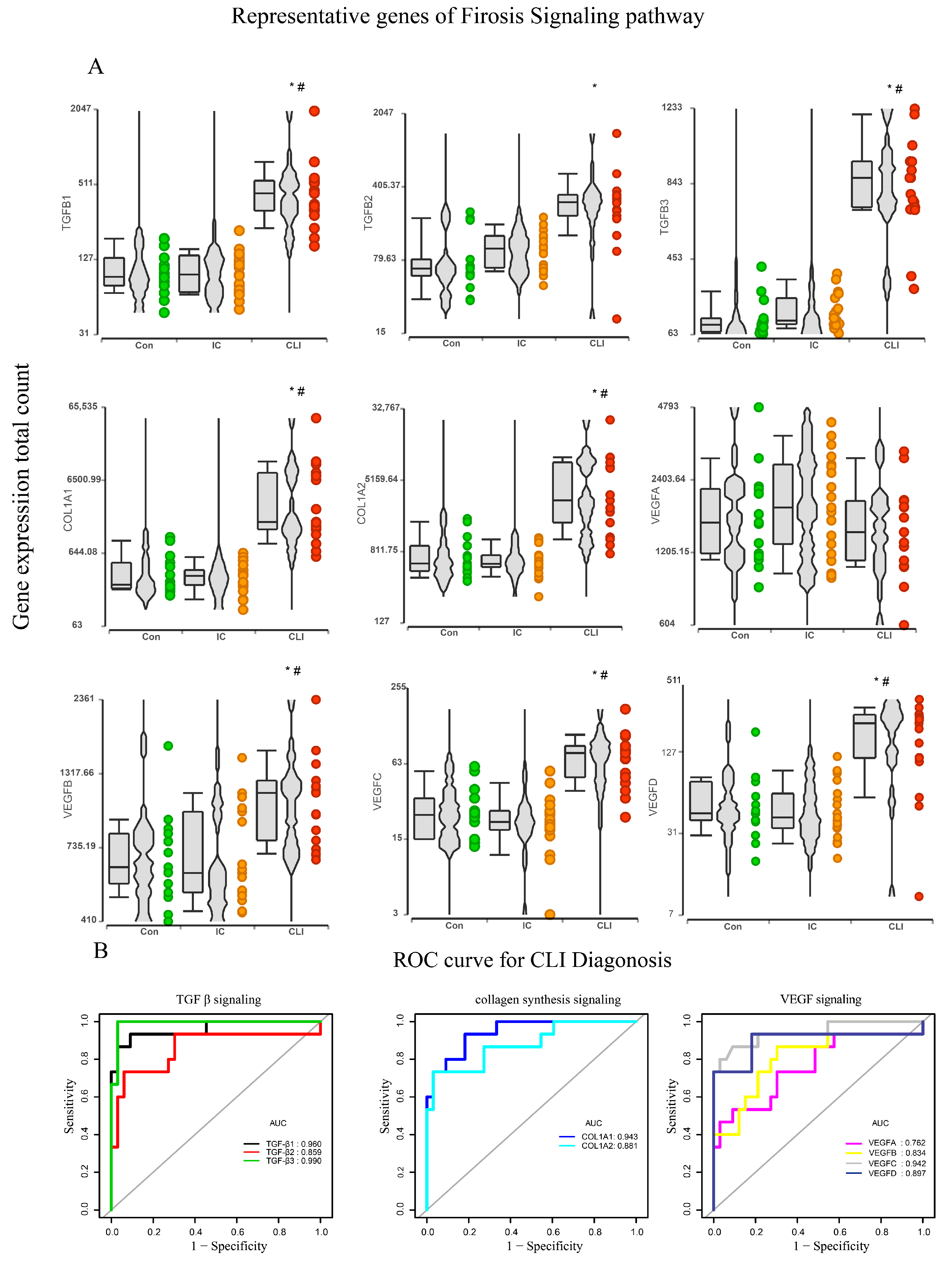
3.4. Fibrosis Pathway Core Genes Expression in Transcriptomics and ROC in the Derivation Cohort
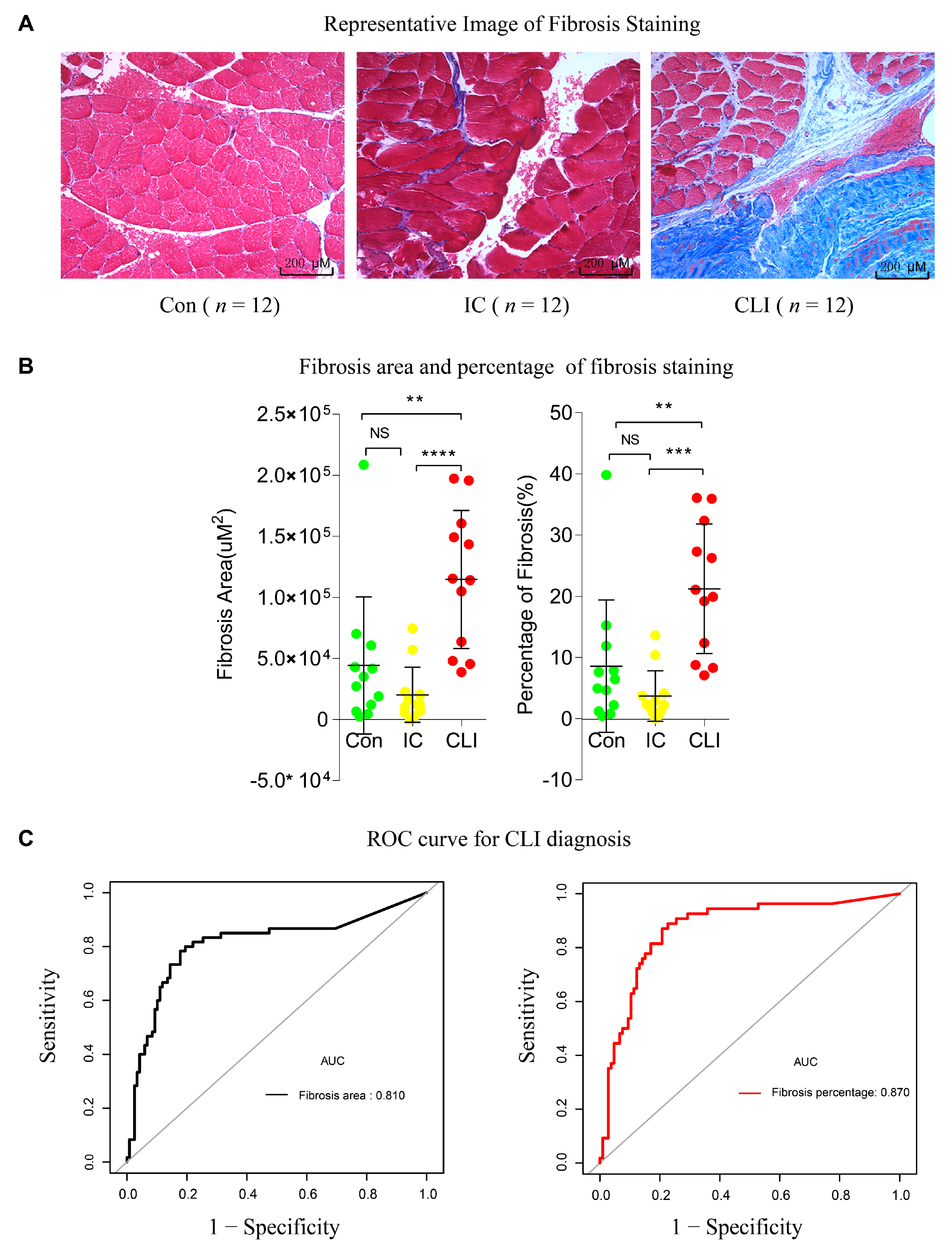
3.5. Fibrosis Quantification and ROC in the Validation Cohort
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
References
- Hiatt, W.R.; Fowkes, F.G.; Heizer, G.; Berger, J.S.; Baumgartner, I.; Held, P.; Katona, B.G.; Mahaffey, K.W.; Norgren, L.; Jones, W.S.; et al. Ticagrelor versus Clopidogrel in Symptomatic Peripheral Artery Disease. N. Engl. J. Med. 2017, 376, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart Disease and Stroke Statistics-2019 Update A Report from the American Heart Association. Circulation 2019, 139, E56–E528. [Google Scholar] [CrossRef]
- Davies, M.G. Critical limb ischemia: Introduction. Methodist Debakey Cardiovasc. J. 2012, 8, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, S.; Sud, K.; Shishehbor, M.H. Nationwide Trends of Hospital Admission and Outcomes among Critical Limb Ischemia Patients: From 2003–2011. J. Am. Coll. Cardiol. 2016, 67, 1901–1913. [Google Scholar] [CrossRef] [PubMed]
- Aquino, R.; Johnnides, C.; Makaroun, M.; Whittle, J.C.; Muluk, V.S.; Kelley, M.E.; Muluk, S.C. Natural history of claudication: Long-term serial follow-up study of 1244 claudicants. J. Vasc. Surg. 2001, 34, 962–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzke, S.; Lepantalo, M. Claudication does not always precede critical leg ischemia. Vasc. Med. 2001, 6, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Shishehbor, M.H.; White, C.J.; Gray, B.H.; Menard, M.T.; Lookstein, R.; Rosenfield, K.; Jaff, M.R. Critical Limb Ischemia: An Expert Statement. J. Am. Coll. Cardiol. 2016, 68, 2002–2015. [Google Scholar] [CrossRef]
- Bunte, M.C.; Jacob, J.; Nudelman, B.; Shishehbor, M.H. Validation of the relationship between ankle-brachial and toe-brachial indices and infragenicular arterial patency in critical limb ischemia. Vasc. Med. 2015, 20, 23–29. [Google Scholar] [CrossRef]
- Shishehbor, M.H.; Hammad, T.A. Treatment of Infrapopliteal Disease in Critical Limb Ischemia: Beyond Angioplasty. Circ. Cardiovasc. Interv. 2016, 9, e003882. [Google Scholar] [CrossRef] [Green Version]
- Schiano, C.; Costa, V.; Aprile, M.; Grimaldi, V.; Maiello, C.; Esposito, R.; Soricelli, A.; Colantuoni, V.; Donatelli, F.; Ciccodicola, A.; et al. Heart failure: Pilot transcriptomic analysis of cardiac tissue by RNA-sequencing. Cardiol. J. 2017, 24, 539–553. [Google Scholar] [CrossRef] [Green Version]
- Matkovich, S.J. Multiomic approaches to delineate the pathogenesis of cardiac disease. Curr. Opin. Cardiol. 2019, 34. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.L.; Huang, L.J.; Luo, W.J.; Ye, S.; Hu, Q.H. Genomic analysis of lncRNA and mRNA profiles in circulating exosomes of patients with rheumatic heart disease. Biol. Open 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, C.K.; Tian, L.; Belbachir, N.; Wnorowski, A.; Shrestha, R.; Ma, N.; Kitani, T.; Rhee, J.W.; Wu, J.C. Identifying the Transcriptome Signatures of Calcium Channel Blockers in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Circ. Res. 2019, 125, 212–222. [Google Scholar] [CrossRef]
- Ryan, T.E.; Yamaguchi, D.J.; Schmidt, C.A.; Zeczycki, T.N.; Shaikh, S.R.; Brophy, P.; Green, T.D.; Tarpey, M.D.; Karnekar, R.; Goldberg, E.J.; et al. Extensive skeletal muscle cell mitochondriopathy distinguishes critical limb ischemia patients from claudicants. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Di, Y.P. Analysis of RNA Sequencing Data Using CLC Genomics Workbench. Methods Mol. Biol. 2020, 2102, 61–113. [Google Scholar] [CrossRef]
- Conesa, A.; Madrigal, P.; Tarazona, S.; Gomez-Cabrero, D.; Cervera, A.; McPherson, A.; Szczesniak, M.W.; Gaffney, D.J.; Elo, L.L.; Zhang, X.; et al. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Zhao, X.; Kwan, J.Y.Y.; Yip, K.; Liu, P.P.; Liu, F.F. Targeting metabolic dysregulation for fibrosis therapy. Nat. Rev. Drug Discov. 2020, 19, 57–75. [Google Scholar] [CrossRef]
- Xu, J.; Cui, X.; Li, J.; Koutakis, P.; Pipinos, I.; Tzeng, E.; Chen, A.; Sachdev, U. Chloroquine improves the response to ischemic muscle injury and increases HMGB1 after arterial ligation. J. Vasc. Surg. 2018, 67, 910–921. [Google Scholar] [CrossRef] [Green Version]
- Schipke, J.; Brandenberger, C.; Rajces, A.; Manninger, M.; Alogna, A.; Post, H.; Muhlfeld, C. Assessment of cardiac fibrosis: A morphometric method comparison for collagen quantification. J. Appl. Physiol. 2017, 122, 1019–1030. [Google Scholar] [CrossRef]
- Available online: https://imagej.nih.gov/ij/ (accessed on 1 October 2020).
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: http://www.R-project.org (accessed on 1 October 2020).
- Mahdy, M.A.A. Skeletal muscle fibrosis: An overview. Cell Tissue Res. 2019, 375, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Mizzi, A.; Cassar, K.; Bowen, C.; Formosa, C. The progression rate of peripheral arterial disease in patients with intermittent claudication: A systematic review. J. Foot Ankle Res. 2019, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Stather, P.W.; Sylvius, N.; Wild, J.B.; Choke, E.; Sayers, R.D.; Bown, M.J. Differential microRNA expression profiles in peripheral arterial disease. Circ. Cardiovasc. Genet. 2013, 6, 490–497. [Google Scholar] [CrossRef] [Green Version]
- Cooke, J.P.; Wilson, A.M. Biomarkers of peripheral arterial disease. J. Am. Coll. Cardiol. 2010, 55, 2017–2023. [Google Scholar] [CrossRef] [Green Version]
- Baczynska, D.; Michalowska, D.; Barc, P.; Skora, J.; Karczewski, M.; Sadakierska-Chudy, A. The expression profile of angiogenic genes in critical limb ischemia popliteal arteries. J. Physiol. Pharm. 2016, 67, 353–362. [Google Scholar]
- Hamburg, N.M.; Creager, M.A. Pathophysiology of Intermittent Claudication in Peripheral Artery Disease. Circ. J. 2017, 81, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Ha, D.M.; Carpenter, L.C.; Koutakis, P.; Swanson, S.A.; Zhu, Z.; Hanna, M.; DeSpiegelaere, H.K.; Pipinos, I.I.; Casale, G.P. Transforming growth factor-beta 1 produced by vascular smooth muscle cells predicts fibrosis in the gastrocnemius of patients with peripheral artery disease. J. Transl. Med. 2016, 14, 39. [Google Scholar] [CrossRef] [Green Version]
- Casanegra, A.I.; Stoner, J.A.; Tafur, A.J.; Pereira, H.A.; Rathbun, S.W.; Gardner, A.W. Differences in galectin-3, a biomarker of fibrosis, between participants with peripheral artery disease and participants with normal ankle-brachial index. Vasc. Med. 2016, 21, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Pizzimenti, M.; Riou, M.; Charles, A.L.; Talha, S.; Meyer, A.; Andres, E.; Chakfé, N.; Lejay, A.; Geny, B. The Rise of Mitochondria in Peripheral Arterial Disease Physiopathology: Experimental and Clinical Data. J. Clin. Med. 2019, 8, 2125. [Google Scholar] [CrossRef] [Green Version]
- Rodenburg, R.J. Biochemical diagnosis of mitochondrial disorders. J. Inherit. Metab. Dis. 2011, 34, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannetti, E.F.; Prigione, A.; Smeitink, J.A.M.; Koopman, W.J.H.; Beyrath, J.; Renkema, H. Live-Imaging Readouts and Cell Models for Phenotypic Profiling of Mitochondrial Function. Front. Genet. 2019, 10, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conte, M.S.; Bradbury, A.W.; Kolh, P.; White, J.V.; Dick, F.; Fitridge, R.; Mills, J.L.; Ricco, J.B.; Suresh, K.R.; Murad, M.H.; et al. Global vascular guidelines on the management of chronic limb-threatening ischemia. J. Vasc. Surg. 2019, 69, 3S–125S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herraiz-Adillo, A.; Martinez-Vizcaino, V.; Cavero-Redondo, I.; Alvarez-Bueno, C.; Garrido-Miguel, M.; Notario-Pacheco, B. Diagnostic Accuracy Study of an Oscillometric Ankle-Brachial Index in Peripheral Arterial Disease: The Influence of Oscillometric Errors and Calcified Legs. PLoS ONE 2016, 11, e0167408. [Google Scholar] [CrossRef]
- Sukul, D.; Grey, S.F.; Henke, P.K.; Gurm, H.S.; Grossman, P.M. Heterogeneity of Ankle-Brachial Indices in Patients Undergoing Revascularization for Critical Limb Ischemia. JACC Cardiovasc. Interv. 2017, 10, 2307–2316. [Google Scholar] [CrossRef]
- Baues, M.; Dasgupta, A.; Ehling, J.; Prakash, J.; Boor, P.; Tacke, F.; Kiessling, F.; Lammers, T. Fibrosis imaging: Current concepts and future directions. Adv. Drug Deliv. Rev. 2017, 121, 9–26. [Google Scholar] [CrossRef]
- Cummings, B.B.; Marshall, J.L.; Tukiainen, T.; Lek, M.; Donkervoort, S.; Foley, A.R.; Bolduc, V.; Waddell, L.B.; Sandaradura, S.A.; O’Grady, G.L.; et al. Improving genetic diagnosis in Mendelian disease with transcriptome sequencing. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Frisk, C.; Eriksson, M.J.; Walentinsson, A.; Corbascio, M.; Hage, C.; Kumar, C.; Asp, M.; Lundeberg, J.; Maret, E.; et al. Transcriptomics of cardiac biopsies reveals differences in patients with or without diagnostic parameters for heart failure with preserved ejection fraction. Sci. Rep. 2019, 9, 3179. [Google Scholar] [CrossRef] [Green Version]
- Ching, T.; Huang, S.; Garmire, L.X. Power analysis and sample size estimation for RNA-Seq differential expression. RNA 2014, 20, 1684–1696. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://doi.org/10.5061/dryad.rr4xgxd67 (accessed on 1 October 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-PAD Controls | Intermittent Claudicant | Critical Limb Ischemia | p Value | |
|---|---|---|---|---|
| Derivation cohort | n = 32 | n = 27 | n = 19 | |
| Age (Mean ± SD) | 61(7.3) | 61(7.5) | 64(10) | 0.18 |
| ABI (Mean ± SD) | 0.65(0.21) | 0.35(0.30) | 0.0007 | |
| Validation cohort | n = 12 | n = 12 | n = 12 | |
| Age (Mean ± SD) | 66.75(4.93) | 68.55(8.68) | 63.58(5.05) | 0.01 |
| ABI (Mean ± SD) | 1.10(0.10) | 0.62(0.13) | 0.31(0.24) | 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cong, G.; Cui, X.; Ferrari, R.; Pipinos, I.I.; Casale, G.P.; Chattopadhyay, A.; Sachdev, U. Fibrosis Distinguishes Critical Limb Ischemia Patients from Claudicants in a Transcriptomic and Histologic Analysis. J. Clin. Med. 2020, 9, 3974. https://doi.org/10.3390/jcm9123974
Cong G, Cui X, Ferrari R, Pipinos II, Casale GP, Chattopadhyay A, Sachdev U. Fibrosis Distinguishes Critical Limb Ischemia Patients from Claudicants in a Transcriptomic and Histologic Analysis. Journal of Clinical Medicine. 2020; 9(12):3974. https://doi.org/10.3390/jcm9123974
Chicago/Turabian StyleCong, Guangzhi, Xiangdong Cui, Ricardo Ferrari, Iraklis I. Pipinos, George P. Casale, Ansuman Chattopadhyay, and Ulka Sachdev. 2020. "Fibrosis Distinguishes Critical Limb Ischemia Patients from Claudicants in a Transcriptomic and Histologic Analysis" Journal of Clinical Medicine 9, no. 12: 3974. https://doi.org/10.3390/jcm9123974