Characterization of LP-Z Lipoprotein Particles and Quantification in Subjects with Liver Disease Using a Newly Developed NMR-Based Assay

 , , ,
, , ,
Abstract
:1. Introduction
2. Experimental Section
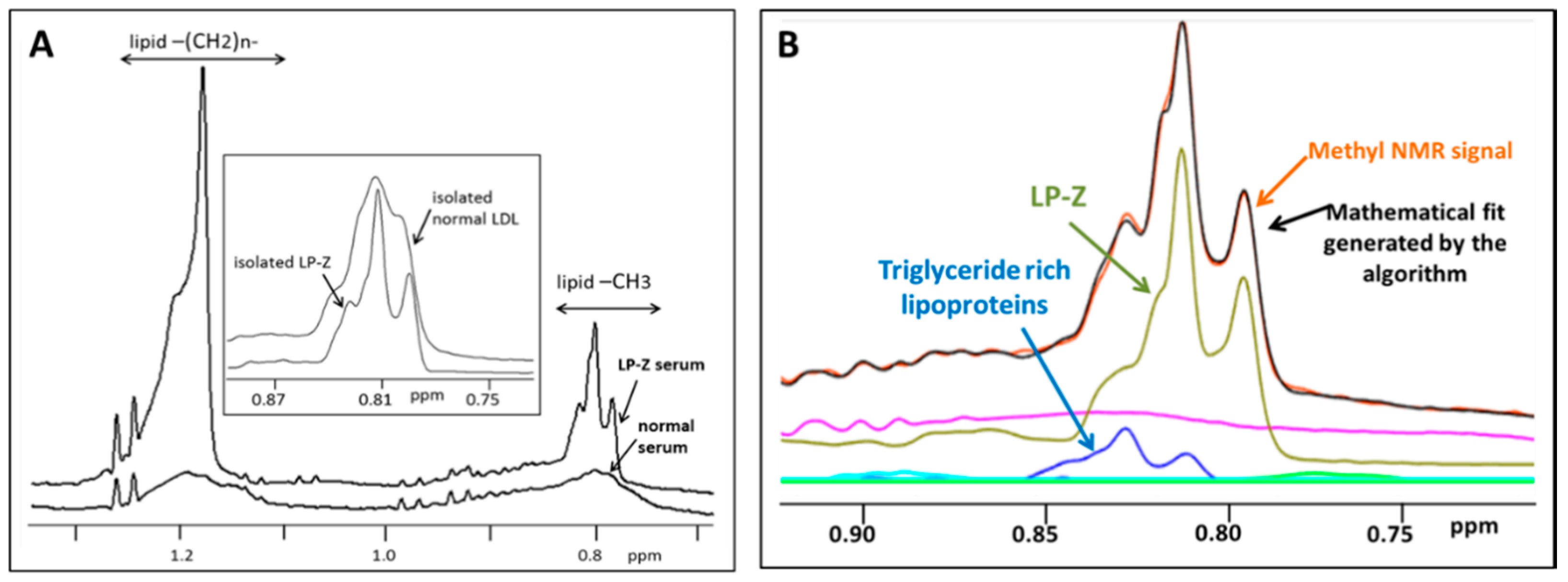
2.1. Development of an Assay for LP-Z Particle Quantification
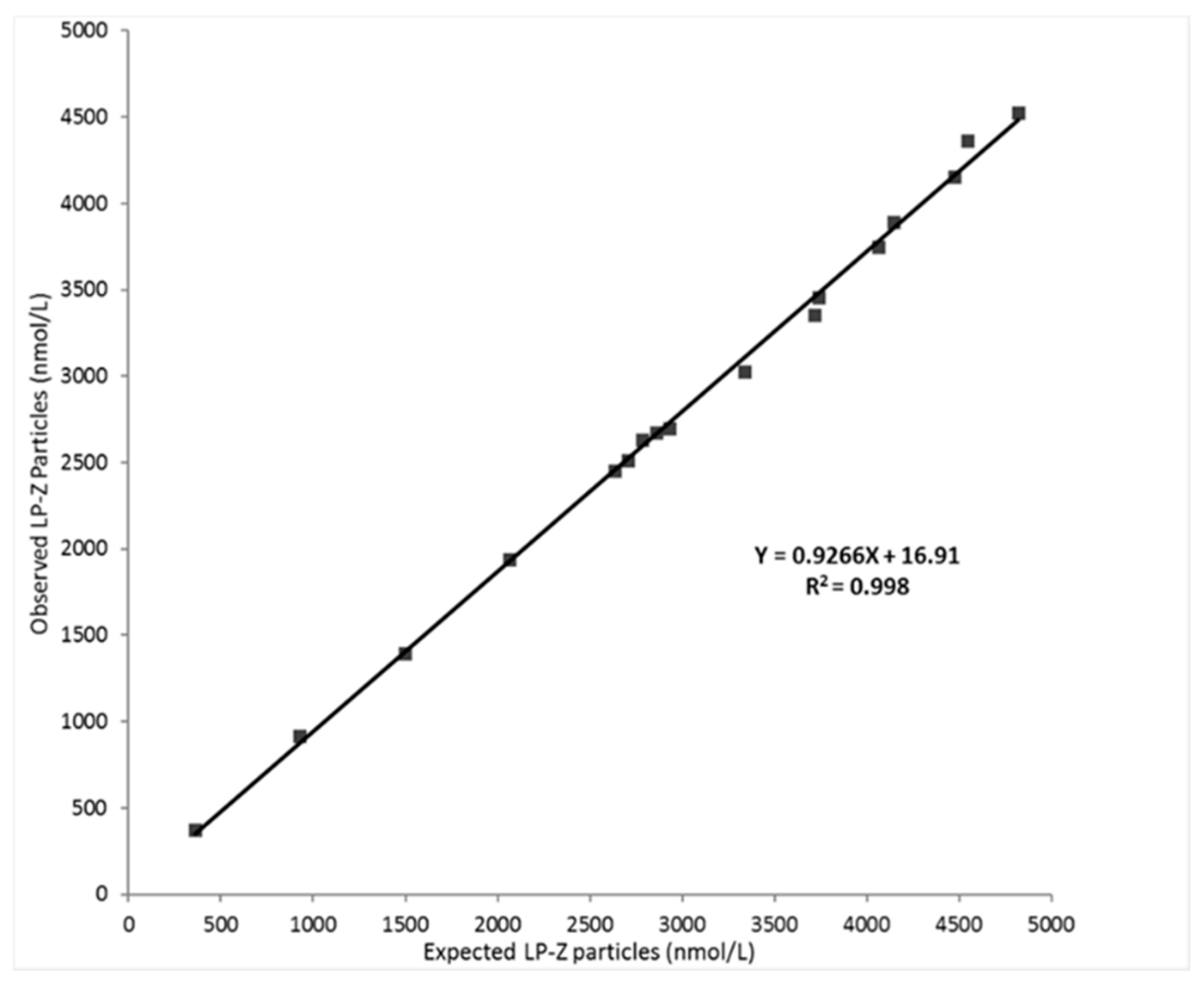
2.2. Evaluation of LP-Z Assay Performance
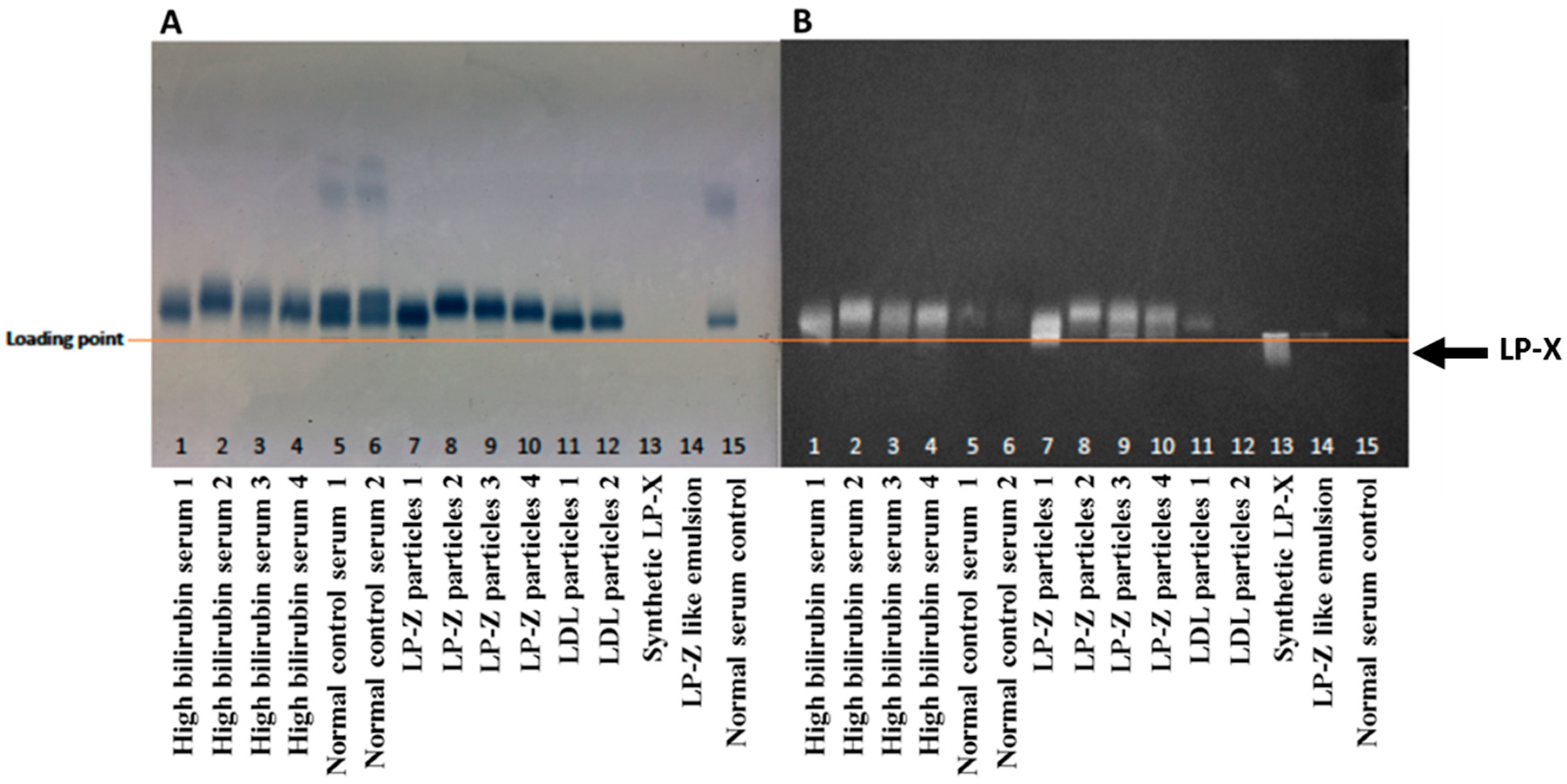
2.3. Isolation of LP-Z Particles from High Bilirubin Samples
2.4. Agarose Gel Electrophoresis and Staining
2.5. Lipid and ApoB Content of Isolated Lipoproteins
2.6. Targeted Lipidomic and Metabolomic Analysis
2.7. Protein Delipidation and Solubilization
2.8. Tryptic Protein Digestion
2.9. Proteomic Analysis
2.10. Polyacrylamide Gel Electrophoresis
2.11. Quantification of LP-Z in Samples from Subjects with Liver Disease or Controls
2.12. Statistical Analysis
3. Results
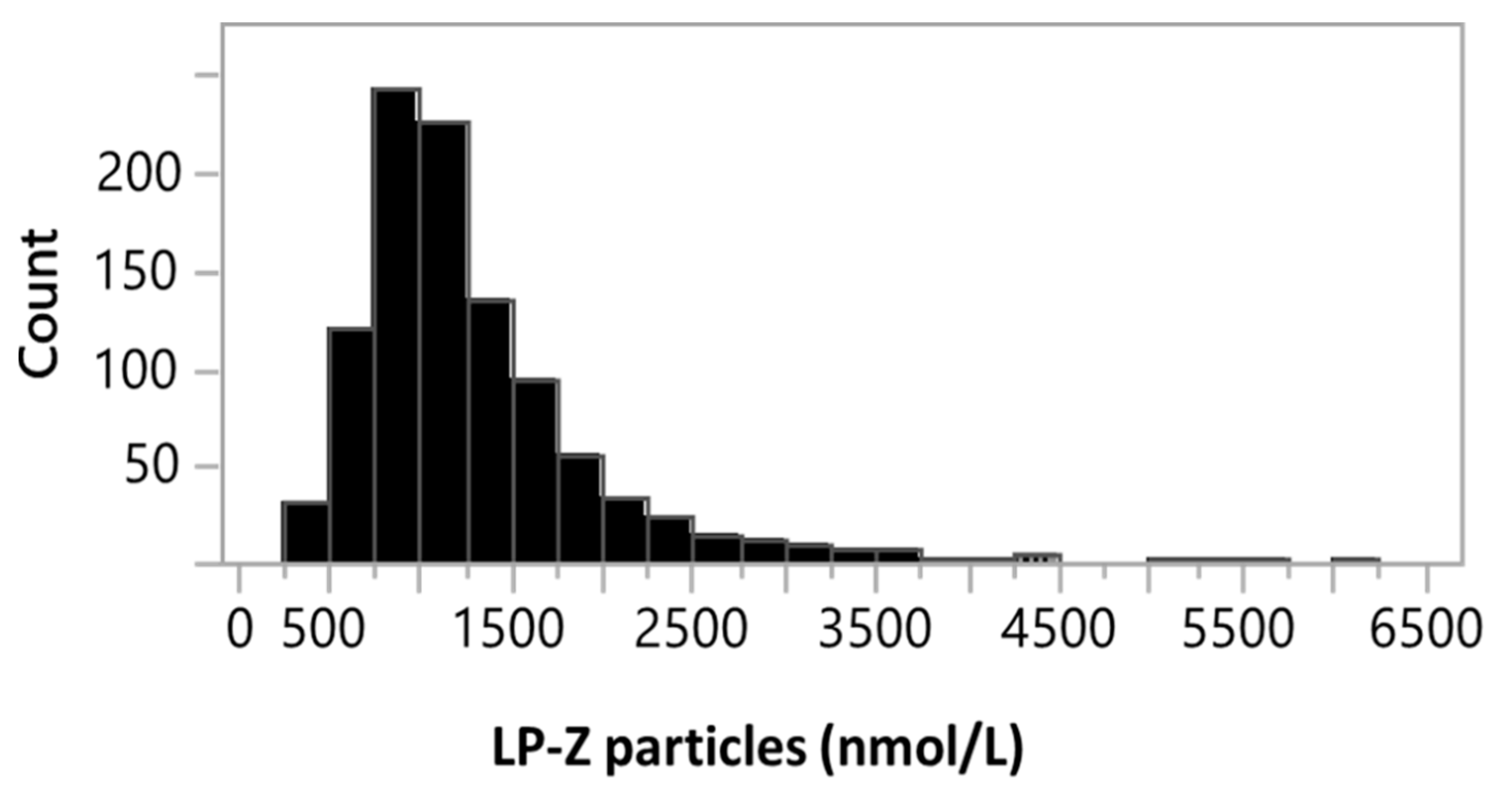
3.1. Analytical Performance of the LP-Z NMR Assay
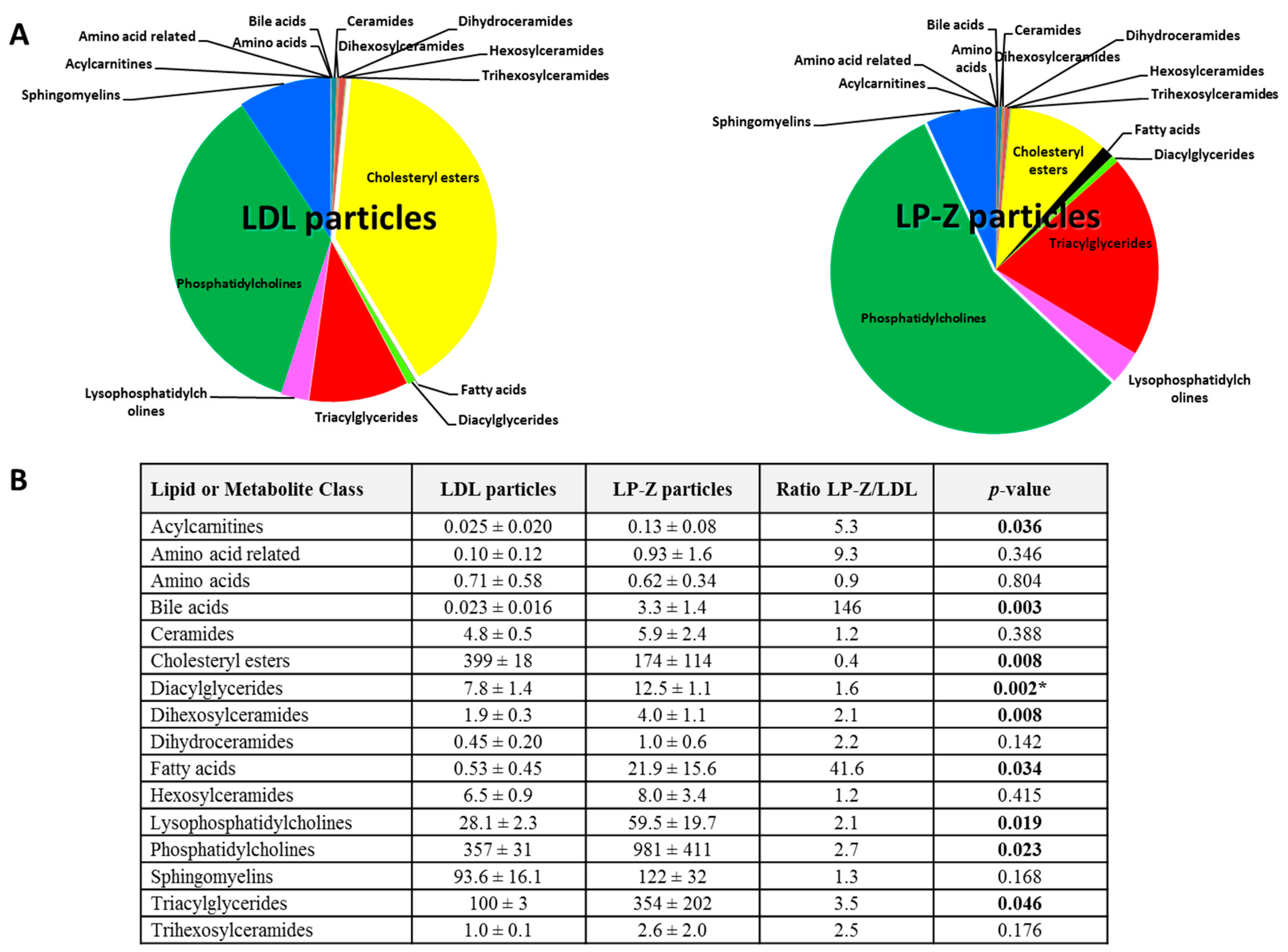
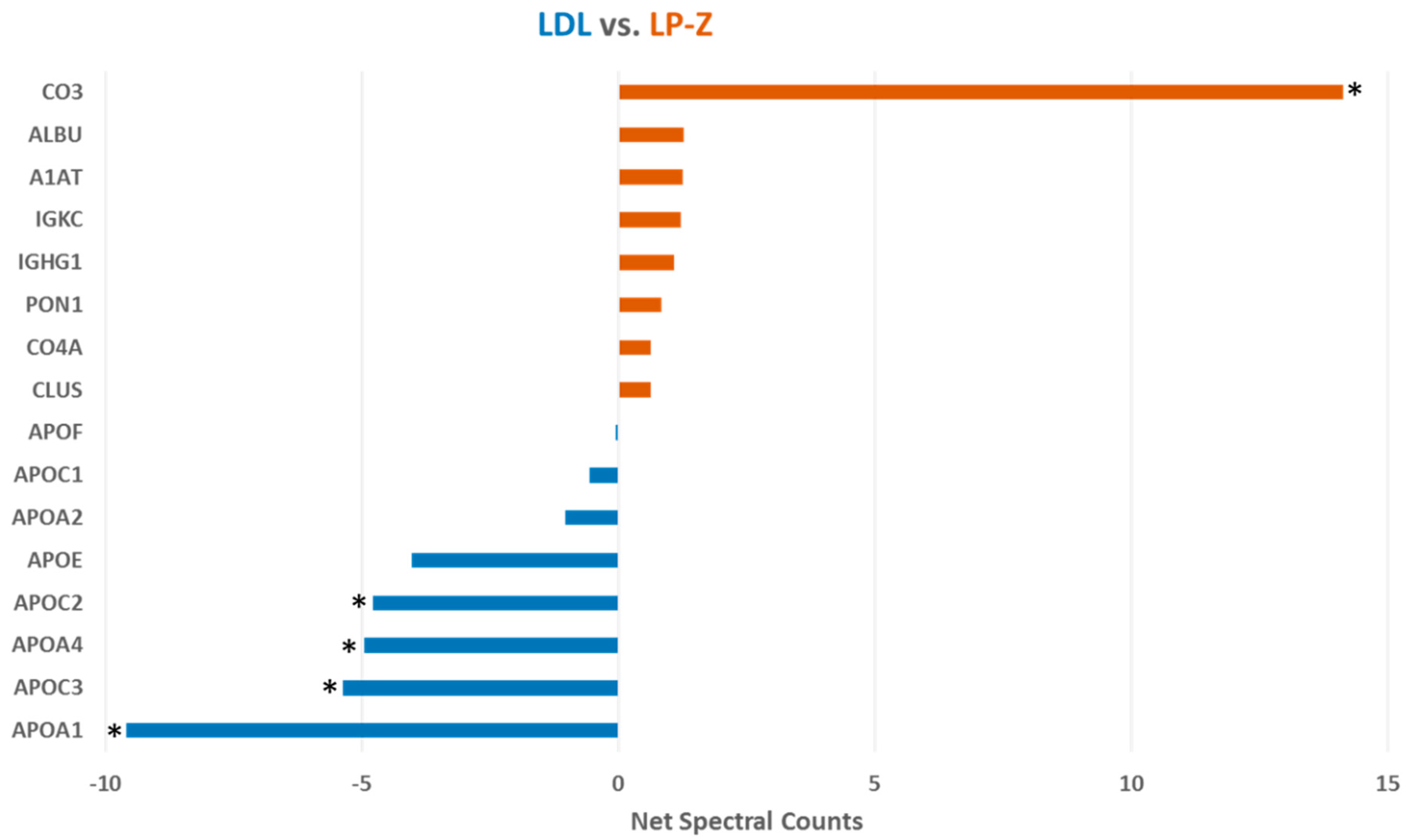
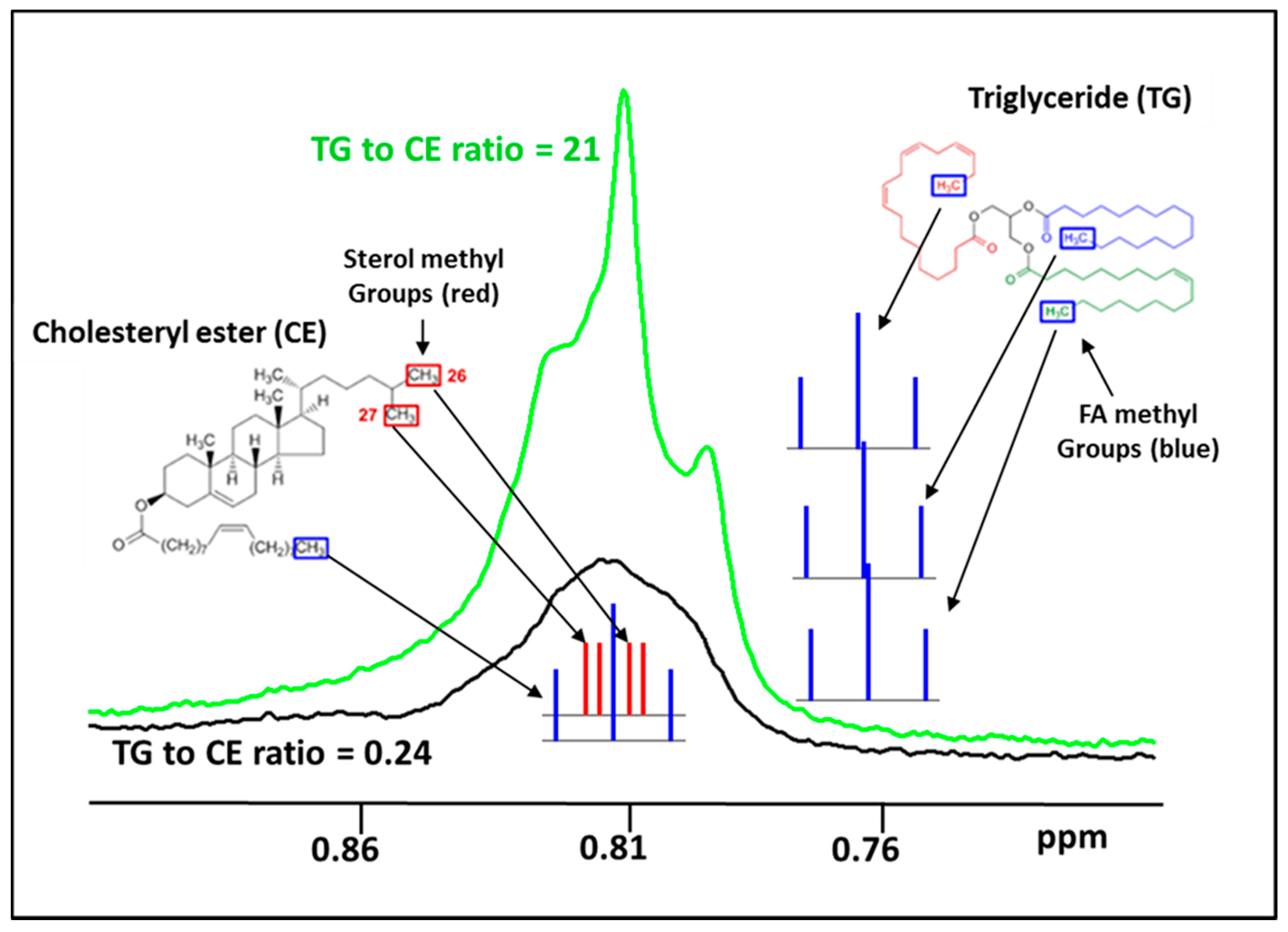
3.2. Characterization of Isolated LP-Z Particles
3.3. Quantification of LPZ Particles in Serum of Subjects with Various Liver Diseases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jeyarajah, E.J.; Cromwell, W.C.; Otvos, J.D. Lipoprotein particle analysis by nuclear magnetic resonance spectroscopy. Clin. Lab. Med. 2006, 26, 847–870. [Google Scholar] [CrossRef] [PubMed]
- Aru, V.; Lam, C.; Khakimov, B.; Hoefsloot, H.C.J.; Zwanenberg, G.; Vendelbo Lind, M.; Schafer, H.; van Duynhoven, J.; Jacobs, D.M.; Smilde, A.K.; et al. Quantification of lipoprotein profiles by nuclear magnetic resonance spectroscopy and multivariate data analysis. Trends Anal. Chem. 2017, 94, 210–219. [Google Scholar] [CrossRef]
- Lounila, J.; Ala-Korpela, M.; Jokisaari, J.; Savolainen, M.J.; Kesaniemi, Y.A. Effects of orientational order and particle size on the NMR line positions of lipoproteins. Phys. Rev. Lett. 1994, 72, 4049–4052. [Google Scholar] [CrossRef] [PubMed]
- Matyus, S.P.; Braun, P.J.; Wolak-Dinsmore, J.; Jeyarajah, E.J.; Shalaurova, I.; Xu, Y.; Warner, S.M.; Clement, T.S.; Connelly, M.A.; Fischer, T.J. NMR measurement of LDL particle number using the Vantera Clinical Analyzer. Clin. Biochem. 2014, 47, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makri, A.; Cheung, A.; Sinaii, N.; Remaley, A.T.; Sampson, M.; Keil, M.; Belyavskaya, E.; Lyssikatos, C.; De La Luz Sierra, M.; Stratakis, C.A.; et al. Lipoprotein particles in patients with pediatric Cushing disease and possible cardiovascular risks. Pediatric. Res. 2019, 86, 375–381. [Google Scholar] [CrossRef]
- Kinzer, A.B.; Shamburek, R.D.; Lightbourne, M.; Muniyappa, R.; Brown, R.J. Advanced Lipoprotein Analysis Shows Atherogenic Lipid Profile That Improves After Metreleptin in Patients with Lipodystrophy. J. Endocr. Soc. 2019, 3, 1503–1517. [Google Scholar] [CrossRef]
- Miller, J.P. Dyslipoproteinaemia of liver disease. Bailliere’s Clin. Endocrinol. Metab. 1990, 4, 807–832. [Google Scholar] [CrossRef]
- Seidel, D.; Alaupovic, P.; Furman, R.H. A lipoprotein characterizing obstructive jaundice. I. Method for quantitative separation and identification of lipoproteins in jaundiced subjects. J. Clin. Investig. 1969, 48, 1211–1223. [Google Scholar] [CrossRef] [Green Version]
- Ross, A.; Murphy, G.M.; Wilkinson, P.A.; Mills, G.L.; Sherlock, S. Occurrence of an abnormal lipoprotein in patients with liver disease. Gut 1970, 11, 1035–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepien, K.M.; Divyateja, H.; Ahmed, F.; Prinsloo, P.; Gupta, P. Lipoprotein X in a patient with cholestasis and hypertriglyceridaemia. Ann. Clin. Biochem. 2013, 50, 173–175. [Google Scholar] [CrossRef] [PubMed]
- Heimerl, S.; Boettcher, A.; Kaul, H.; Liebisch, G. Lipid profiling of lipoprotein X: Implications for dyslipidemia in cholestasis. Biochim. Et Biophys. Acta 2016, 1861, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S. Biochemistry and clinical relevance of lipoprotein X. Ann. Clin. Lab. Sci. 1984, 14, 371–374. [Google Scholar]
- Seidel, D.; Alaupovic, P.; Furman, R.H.; McConathy, W.J. A lipoprotein characterizing obstructive jaundice. II. Isolation and partial characterization of the protein moieties of low density lipoproteins. J. Clin. Investig. 1970, 49, 2396–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostner, G.M.; Laggner, P.; Prexl, H.J.; Holasek, A. Investigation of the abnormal low-density lipoproteins occurring in patients with obstructive jaundice. Biochem. J. 1976, 157, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Agorastos, J.; Fox, C.; Harry, D.S.; McIntyre, N. Lecithin--cholesterol acyltransferase and the lipoprotein abnormalities of obstructive jaundice. Clin. Sci. Mol. Med. 1978, 54, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Matyus, S.P.; Braun, P.J.; Wolak-Dinsmore, J.; Saenger, A.K.; Jeyarajah, E.J.; Shalaurova, I.; Warner, S.M.; Fischer, T.J.; Connelly, M.A. HDL particle number measured on the Vantera(R), the first clinical NMR analyzer. Clin. Biochem. 2015, 48, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinl, R.E.; Tennant, H.M.; Ricketts, J.C.; Rice, C.R.; Robinson, C.B.; Sandesara, P.B.; Moriarty, P.M.; Sperling, L. Lipoprotein-X disease in the setting of severe cholestatic hepatobiliary autoimmune disease. J. Clin. Lipidol. 2017, 11, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. CLSI Document EP17-A: Protocols for Determination of Limits of Detection and Limits of Quantitation; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2004. [Google Scholar]
- Clinical and Laboratory Standards Institute. CLSI Document EP6-A: Evaluation of the Linearity of Quantitative Measurement Procedures: A Statistical Approach; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2003. [Google Scholar]
- Clinical and Laboratory Standards Institute. CLSI document EP5-A2: Evaluation of Precision Performance of Quantitative Measurements Method; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2004. [Google Scholar]
- Melchior, J.T.; Street, S.E.; Andraski, A.B.; Furtado, J.D.; Sacks, F.M.; Shute, R.L.; Greve, E.I.; Swertfeger, D.K.; Li, H.; Shah, A.S.; et al. Apolipoprotein A-II alters the proteome of human lipoproteins and enhances cholesterol efflux from ABCA1. J. Lipid Res. 2017, 58, 1374–1385. [Google Scholar] [CrossRef] [Green Version]
- Shireman, R.; Kilgore, L.L.; Fisher, W.R. Solubilization of apolipoprotein B and its specific binding by the cellular receptor for low density lipoprotein. Proc. Natl. Acad. Sci. USA 1977, 74, 5150–5154. [Google Scholar] [CrossRef] [Green Version]
- Furtado, J.D.; Yamamoto, R.; Melchior, J.T.; Andraski, A.B.; Gamez-Guerrero, M.; Mulcahy, P.; He, Z.; Cai, T.; Davidson, W.S.; Sacks, F.M. Distinct Proteomic Signatures in 16 HDL (High-Density Lipoprotein) Subspecies. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2827–2842. [Google Scholar] [CrossRef] [Green Version]
- Dullaart, R.P.; Gruppen, E.G.; Connelly, M.A.; Otvos, J.D.; Lefrandt, J.D. GlycA, a biomarker of inflammatory glycoproteins, is more closely related to the leptin/adiponectin ratio than to glucose tolerance status. Clin. Biochem. 2015, 48, 811–814. [Google Scholar] [CrossRef]
- Sabesin, S.M.; Hawkins, H.L.; Kuiken, L.; Ragland, J.B. Abnormal plasma lipoproteins and lecithin-cholesterol acyltransferase deficiency in alcoholic liver disease. Gastroenterology 1977, 72, 510–518. [Google Scholar] [CrossRef]
- Weidman, S.W.; Ragland, J.B.; Sabesin, S.M. Plasma lipoprotein composition in alcoholic hepatitis: Accumulation of apolipoprotein E-rich high density lipoprotein and preferential reappearance of “light”-HDL during partial recovery. J. Lipid Res. 1982, 23, 556–569. [Google Scholar]
- Harry, D.S.; Day, R.C.; Owen, J.S.; Agorastos, J.; Foo, A.Y.; McIntyre, N. Plasma lecithin:cholesterol acyltransferase activity and the lipoprotein abnormalities of liver disease. Scand. J. Clin. Lab. Investig. Suppl. 1978, 150, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Nass, K.J.; van den Berg, E.H.; Gruppen, E.G.; Dullaart, R.P.F. Plasma lecithin:cholesterol acyltransferase and phospholipid transfer protein activity independently associate with nonalcoholic fatty liver disease. Eur. J. Clin. Investig. 2018, 48, e12988. [Google Scholar] [CrossRef] [PubMed]
- Trieb, M.; Horvath, A.; Birner-Gruenberger, R.; Spindelboeck, W.; Stadlbauer, V.; Taschler, U.; Curcic, S.; Stauber, R.E.; Holzer, M.; Pasterk, L.; et al. Liver disease alters high-density lipoprotein composition, metabolism and function. Biochim. Et Biophys. Acta 2016, 1861, 630–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Zychlinski, A.; Williams, M.; McCormick, S.; Kleffmann, T. Absolute quantification of apolipoproteins and associated proteins on human plasma lipoproteins. J. Proteom. 2014, 106, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.Y.; Chen, S.F.; Lai, M.D.; Chang, T.T.; Chen, T.L.; Li, P.Y.; Shieh, D.B.; Young, K.C. Comparative proteomic profiling of plasma very-low-density and low-density lipoproteins. Clin. Chim. Acta Int. J. Clin. Chem. 2010, 411, 336–344. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Nieddu, G.; Zinellu, E.; De Muro, P.; Piredda, F.; Guarino, A.; Spirito, R.; Carta, F.; Turrini, F.; Formato, M. Proteomic analysis of plasma-purified VLDL, LDL, and HDL fractions from atherosclerotic patients undergoing carotid endarterectomy: Identification of serum amyloid A as a potential marker. Oxid. Med. Cell. Longev. 2013, 2013, 385214. [Google Scholar] [CrossRef]
- Potter, B.J.; Trueman, A.M.; Jones, E.A. Serum complement in chronic liver disease. Gut 1973, 14, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Schaap, F.G.; Trauner, M.; Jansen, P.L. Bile acid receptors as targets for drug development. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 55–67. [Google Scholar] [CrossRef]
- de Aguiar Vallim, T.Q.; Tarling, E.J.; Edwards, P.A. Pleiotropic roles of bile acids in metabolism. Cell Metab. 2013, 17, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Pircher, P.C.; Schulman, I.G.; Westin, S.K. Regulation of complement C3 expression by the bile acid receptor FXR. J. Biol. Chem. 2005, 280, 7427–7434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claudel, T.; Sturm, E.; Duez, H.; Torra, I.P.; Sirvent, A.; Kosykh, V.; Fruchart, J.C.; Dallongeville, J.; Hum, D.W.; Kuipers, F.; et al. Bile acid-activated nuclear receptor FXR suppresses apolipoprotein A-I transcription via a negative FXR response element. J. Clin. Investig. 2002, 109, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Claudel, T.; Inoue, Y.; Barbier, O.; Duran-Sandoval, D.; Kosykh, V.; Fruchart, J.; Fruchart, J.C.; Gonzalez, F.J.; Staels, B. Farnesoid X receptor agonists suppress hepatic apolipoprotein CIII expression. Gastroenterology 2003, 125, 544–555. [Google Scholar] [CrossRef]
- Perez-Matos, M.C.; Sandhu, B.; Bonder, A.; Jiang, Z.G. Lipoprotein metabolism in liver diseases. Curr. Opin. Lipidol. 2019, 30, 30–36. [Google Scholar] [CrossRef]
- Tapper, E.B.; Parikh, N.D. Mortality due to cirrhosis and liver cancer in the United States, 1999–2016: Observational study. Bmj 2018, 362, k2817. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Related Parameters | Normal Control 1 | Normal Control 2 | High Bilirubin 1 | High Bilirubin 2 | High Bilirubin 3 | High Bilirubin 4 |
|---|---|---|---|---|---|---|
| Bilirubin (mg/dL) | ND | ND | 22.2 | 13.4 | 18.9 | 24.6 |
| TG (mg/dL) | 48.0 | 59.9 | 145.9 | 347.8 | 443.8 | 771.2 |
| TC (mg/dL) | 121.3 | 189.3 | 90.7 | 194.8 | 265.6 | 343.5 |
| LDL-C (mg/dL) | 77.1 | 82.9 | 80.1 | 169.3 | 98.7 | 84.3 |
| HDL-C (mg/dL) | 38.4 | 102.3 | 6.4 | 4.9 | 7.7 | 6.7 |
| Apo A-I (mg/dL) | 102.0 | 169.4 | 19.7 | 14.7 | 23.1 | 21.6 |
| Apo B (mg/dL) | 60.8 | 57.1 | 175.0 | 364.0 | 368.3 | 304.1 |
| TRL-P (nmol/L) | 26.8 | 18.6 | 16.3 | 81.5 | 985.8 | 651.8 |
| VL-TRL-P (nmol/L) | 0.1 | 0.0 | 0.0 | 0.0 | 0.0 | 3.5 |
| L-TRL-P (nmol/L) | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 6.6 |
| M-TRL-P (nmol/L) | 4.5 | 1.5 | 16.3 | 81.5 | 63.0 | 369.2 |
| S-TRL-P (nmol/L) | 22.2 | 17.1 | 0.0 | 0.0 | 0.0 | 0.0 |
| VS-TRL-P (nmol/L) | 0.0 | 0.0 | 0.0 | 0.0 | 920.6 | 272.5 |
| LDL-P (nmol/L) | 1112 | 1170 | 3427 | 7026 | 5693 | 4531 |
| L-LDL-P (nmol/L) | 220.9 | 0.0 | 0.0 | 0.0 | 0.0 | 123.8 |
| M-LDL-P (nmol/L) | 695.3 | 429.3 | 0.0 | 0.0 | 0.0 | 0.0 |
| S-LDL-P (nmol/L) | 195.6 | 740.2 | 458.8 | 1085.2 | 0.0 | 0.0 |
| LP-Z (nmol/L) | 0.0 | 0.0 | 2968 | 5941 | 5693 | 4407 |
| HDL-P (µmol/L) | 19.4 | 20.8 | 3.8 | 4.9 | 7.7 | 6.3 |
| L-HDL-P (µmol/L) | 0.9 | 7.2 | 0.3 | 0.0 | 0.0 | 0.0 |
| M-HDL-P (µmol/L) | 4.1 | 6.2 | 0.0 | 0.0 | 0.0 | 0.0 |
| S-HDL-P (µmol/L) | 14.4 | 7.4 | 3.6 | 4.9 | 7.7 | 6.3 |
| TRLZ (nm) | 36.4 | 37.7 | 42.0 | 40.3 | 50.1 | 51.0 |
| LDLZ (nm) | 21.1 | 20.1 | 19.0 | 19.0 | 19.9 | 22.4 |
| HDLZ (nm) | 8.8 | 10.6 | 8.3 | 7.4 | 7.4 | 7.5 |
| Mass % Composition | Normal Control 1 | Normal Control 2 | High Bilirubin 1 | High Bilirubin 2 | High Bilirubin 3 | High Bilirubin 4 |
|---|---|---|---|---|---|---|
| Protein | 21.9 | 20.5 | 21.4 | 22.6 | 23.9 | 22.2 |
| Phospholipids | 20.5 | 21.3 | 26.6 | 23.3 | 23.9 | 26.7 |
| Triglycerides (TG) | 11.4 | 15.8 | 29.2 | 44.1 | 32.7 | 34.6 |
| Free cholesterol (FC) | 7.4 | 5.4 | 8.6 | 6.2 | 8.6 | 9.2 |
| Cholesteryl esters (CE) | 38.8 | 37.0 | 14.2 | 3.8 | 10.8 | 7.4 |
| FC/CE ratio | 0.2 | 0.1 | 0.6 | 1.6 | 0.8 | 1.2 |
| TG/CE ratio | 0.3 | 0.4 | 2.1 | 11.6 | 3.0 | 4.7 |
| Subjects (# of Samples Tested) | TG (mg/dL) | TC (mg/dL) | LP-Z (nmol/L) |
|---|---|---|---|
| Normal healthy adults (769) | 148 ± 94 | 186 ± 36 | 0.0 |
| Primary biliary cholangitis (PBC) (11) | 113 ± 30 | 173 ± 21 | 0.0 |
| Primary sclerosing cholangitis (PSC) (11) | 131 ± 61 | 179 ± 48 | 0.0 |
| Autoimmune hepatitis (AIH) (19) | 116 ± 64 | 170 ± 48 | 0.0 |
| Alcoholic liver disease (ALD) (47) | 99 ± 72 | 133 ± 46 | 1550 ± 1257 * |
| T2DM and metabolic syndrome (138) | 172 ± 124 | 192 ± 31 | 0.0 |
| Hypertriglyceridemia (≥500 mg/dL TG) (1970) | 730 ± 266 | 273 ± 58 | 1690 ± 772 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedi, S.; Garcia, E.; Jeyarajah, E.J.; Shalaurova, I.; Perez-Matos, M.C.; Jiang, Z.G.; Dullaart, R.P.F.; Matyus, S.P.; Kirk, W.J.; Otvos, J.D.; et al. Characterization of LP-Z Lipoprotein Particles and Quantification in Subjects with Liver Disease Using a Newly Developed NMR-Based Assay. J. Clin. Med. 2020, 9, 2915. https://doi.org/10.3390/jcm9092915
Bedi S, Garcia E, Jeyarajah EJ, Shalaurova I, Perez-Matos MC, Jiang ZG, Dullaart RPF, Matyus SP, Kirk WJ, Otvos JD, et al. Characterization of LP-Z Lipoprotein Particles and Quantification in Subjects with Liver Disease Using a Newly Developed NMR-Based Assay. Journal of Clinical Medicine. 2020; 9(9):2915. https://doi.org/10.3390/jcm9092915
Chicago/Turabian StyleBedi, Shimpi, Erwin Garcia, Elias J. Jeyarajah, Irina Shalaurova, Maria Camila Perez-Matos, Z. Gordon Jiang, Robin P. F. Dullaart, Steven P. Matyus, William J. Kirk, James D. Otvos, and et al. 2020. "Characterization of LP-Z Lipoprotein Particles and Quantification in Subjects with Liver Disease Using a Newly Developed NMR-Based Assay" Journal of Clinical Medicine 9, no. 9: 2915. https://doi.org/10.3390/jcm9092915