Neuropilin1 Expression Acts as a Prognostic Marker in Stomach Adenocarcinoma by Predicting the Infiltration of Treg Cells and M2 Macrophages


Abstract
:1. Introduction
2. Experimental Section
2.1. Analysis of NRP1 Expression in Various Types of Tumors and Normal Tissues
2.2. Evaluation of the Relationship between NRP1 Expression and Promoter Methylation in Clinical Characteristics
2.3. Evaluation of the Relationship between NRP1 Expression and Patient Survival with Various Tumors
2.4. Evaluation of the Correlation between NRP1 Expression and Immune Cell Infiltration
3. Results
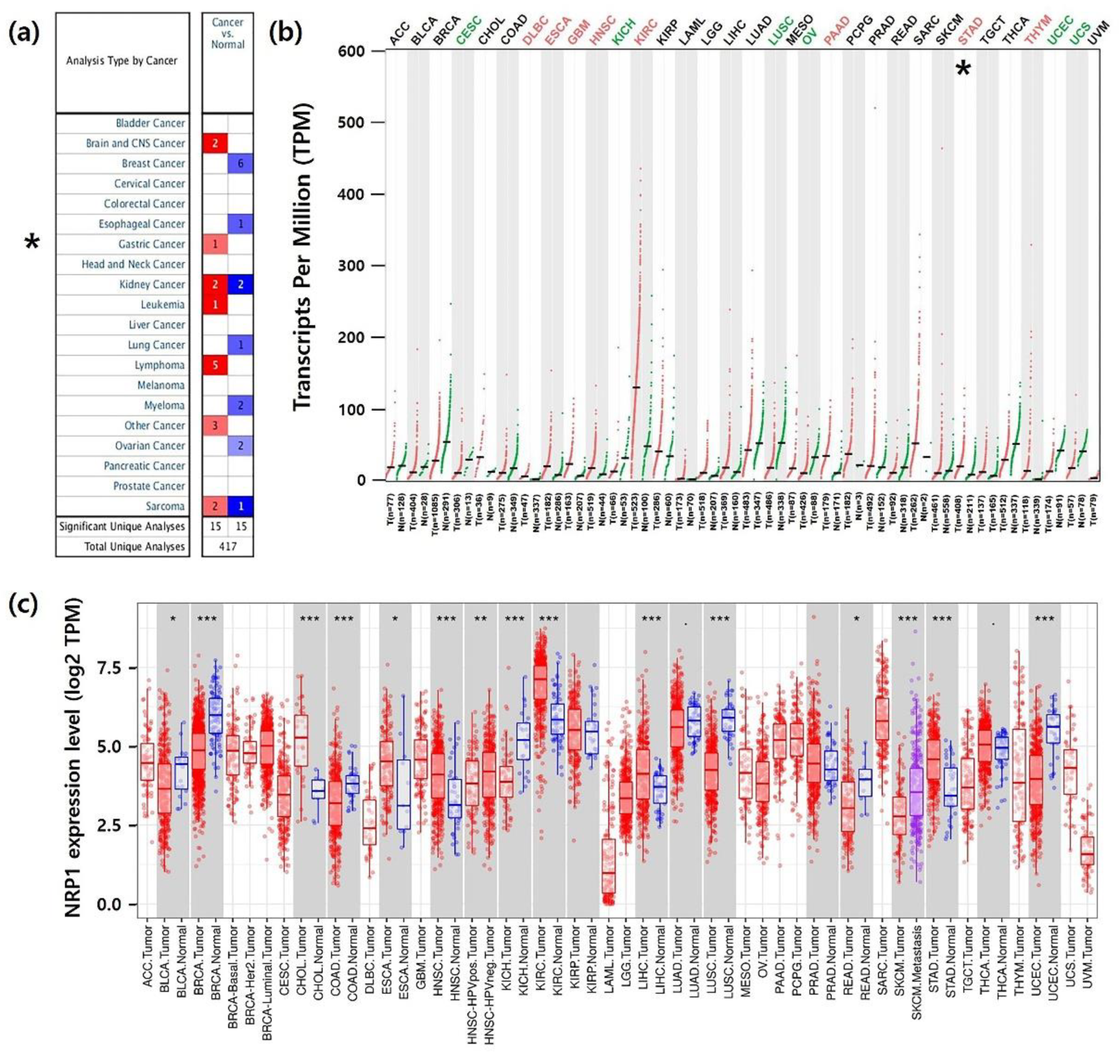
3.1. mRNA Expression Levels of NRP1 in Various Types of Human Cancer
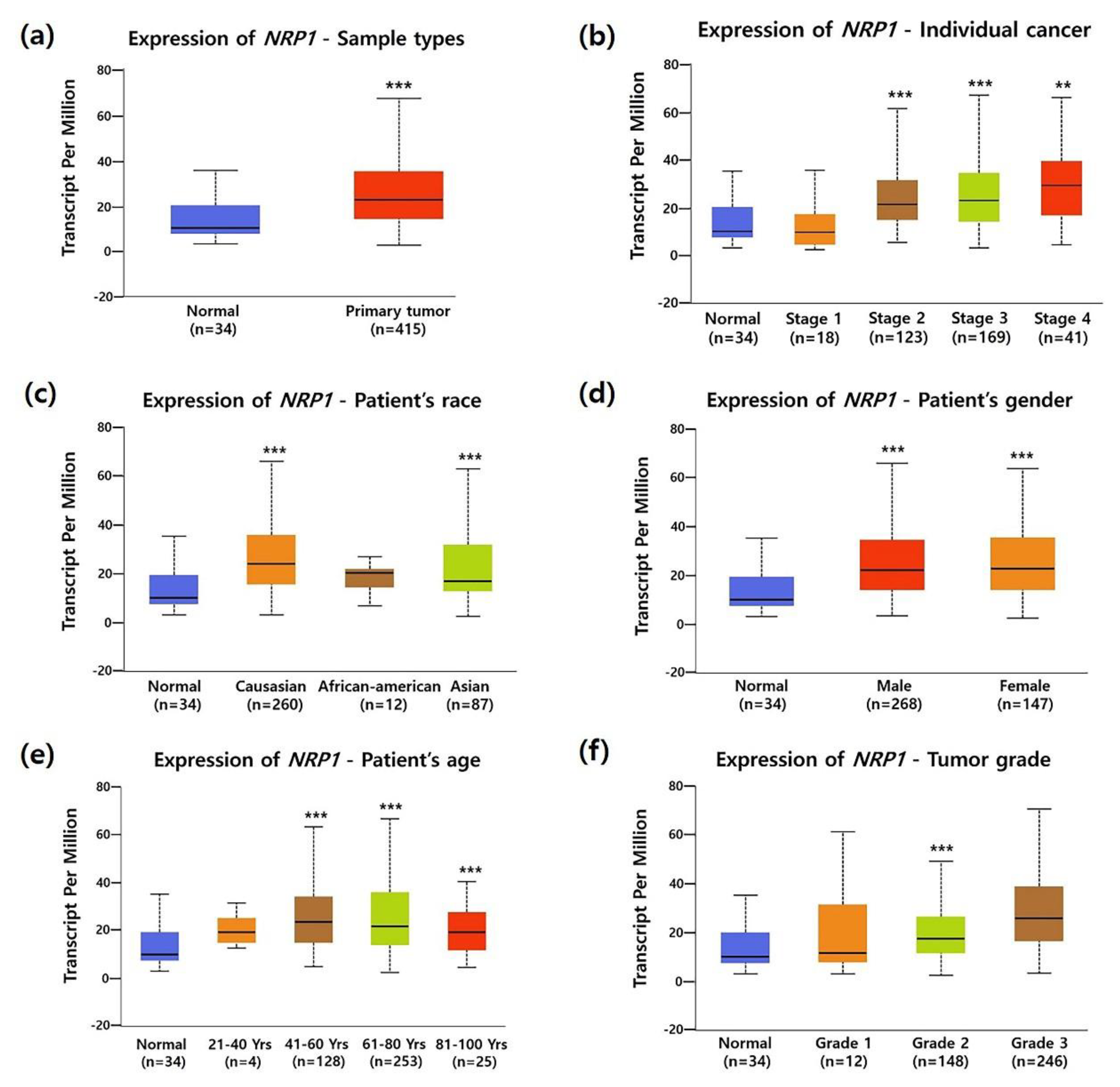
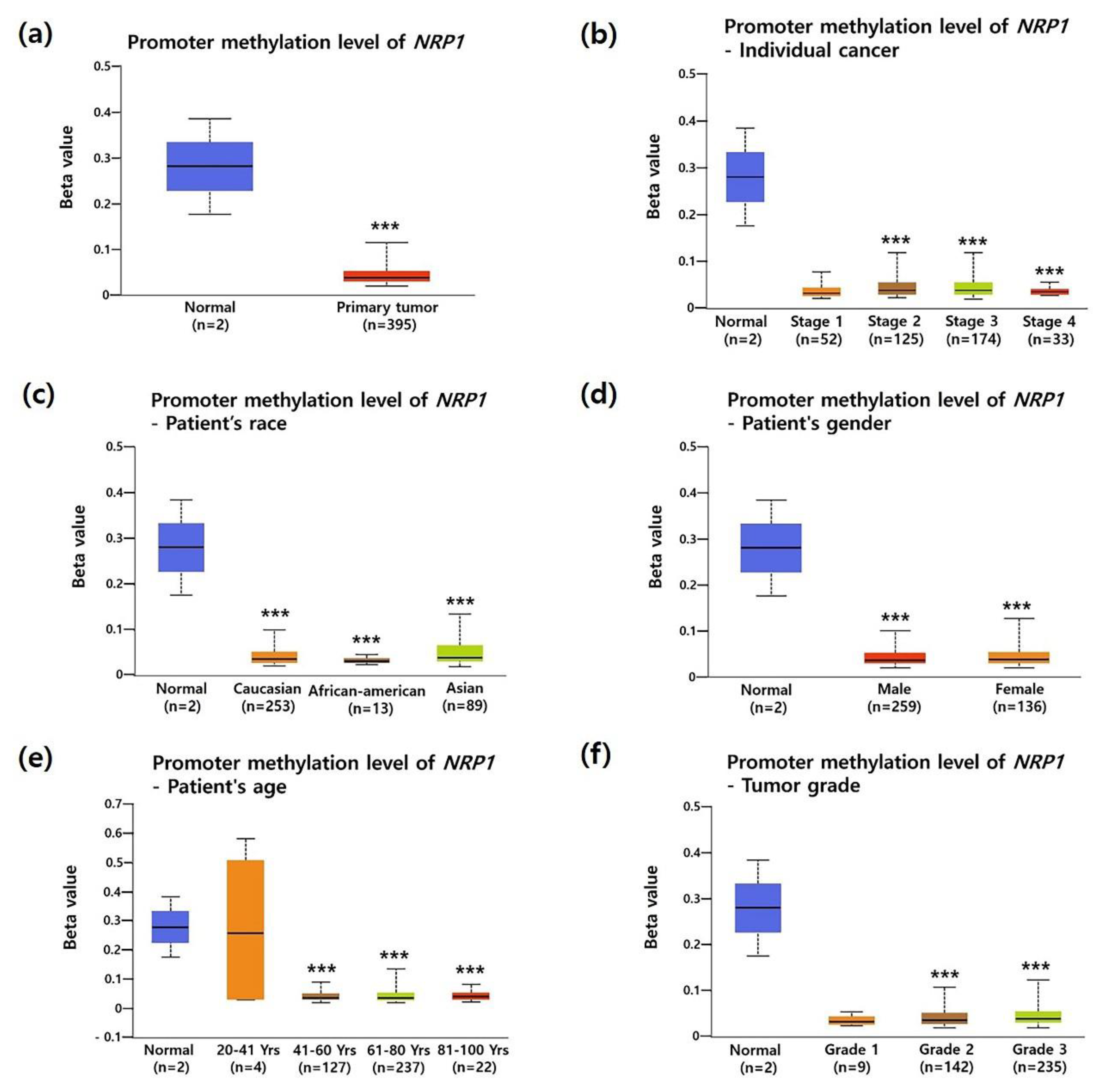
3.2. Association between NRP1 Expression and Clinical Characteristics in STAD Patients
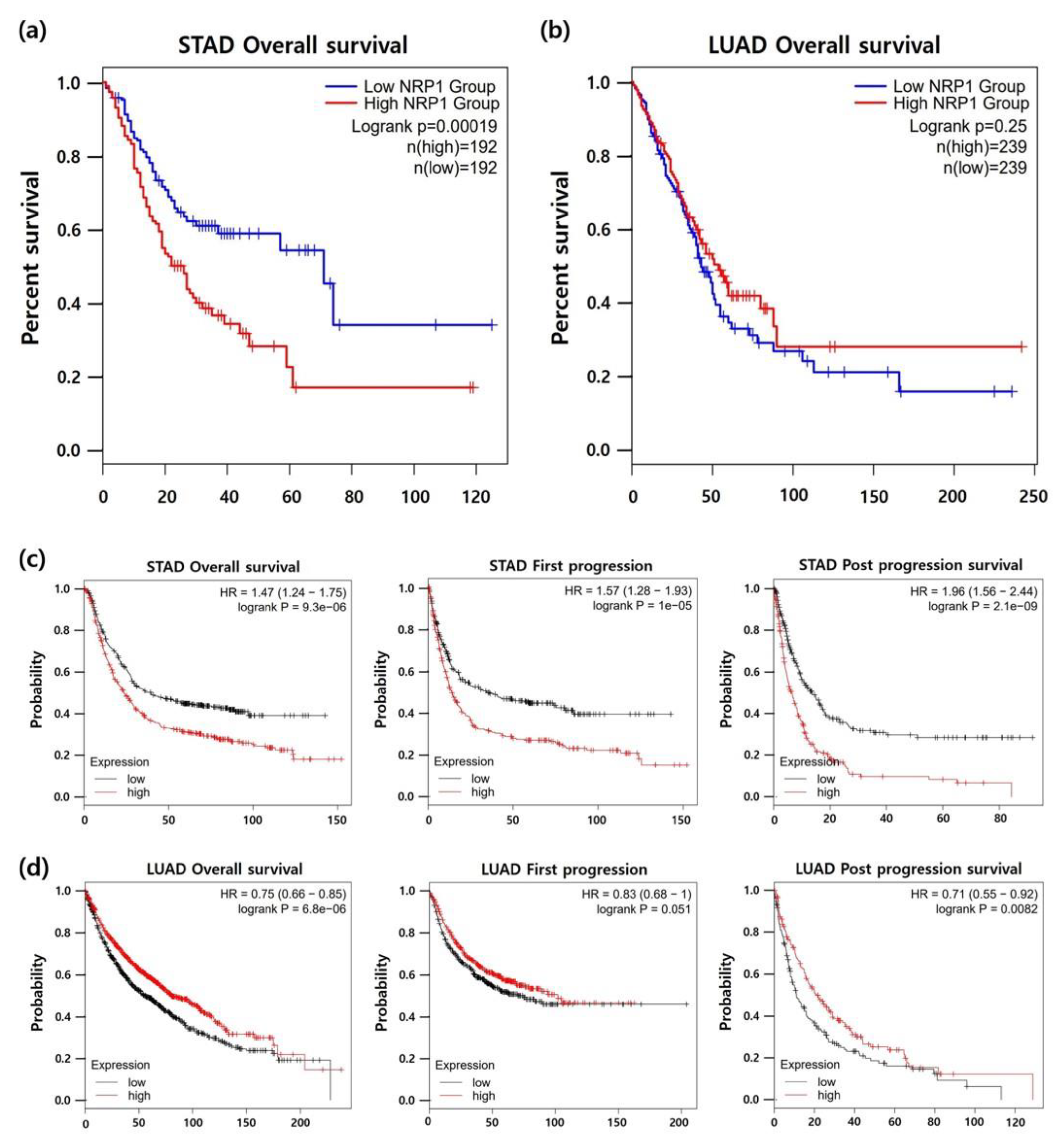
3.3. Association between NRP1 Expression and Prognosis in Patients with STAD
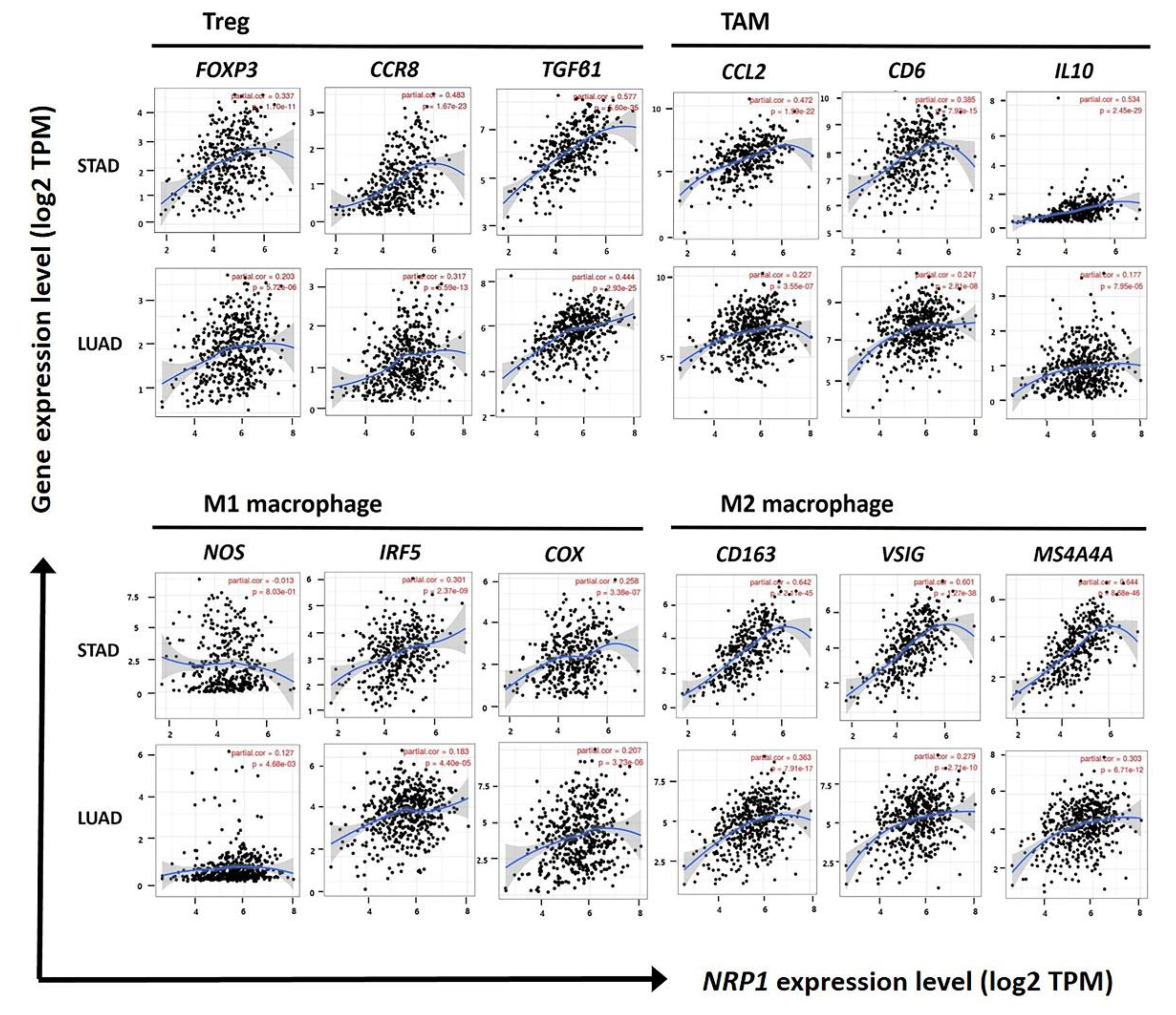
3.4. Correlation of NRP1 Expression with Treg Cells and M2 Macrophages in STAD
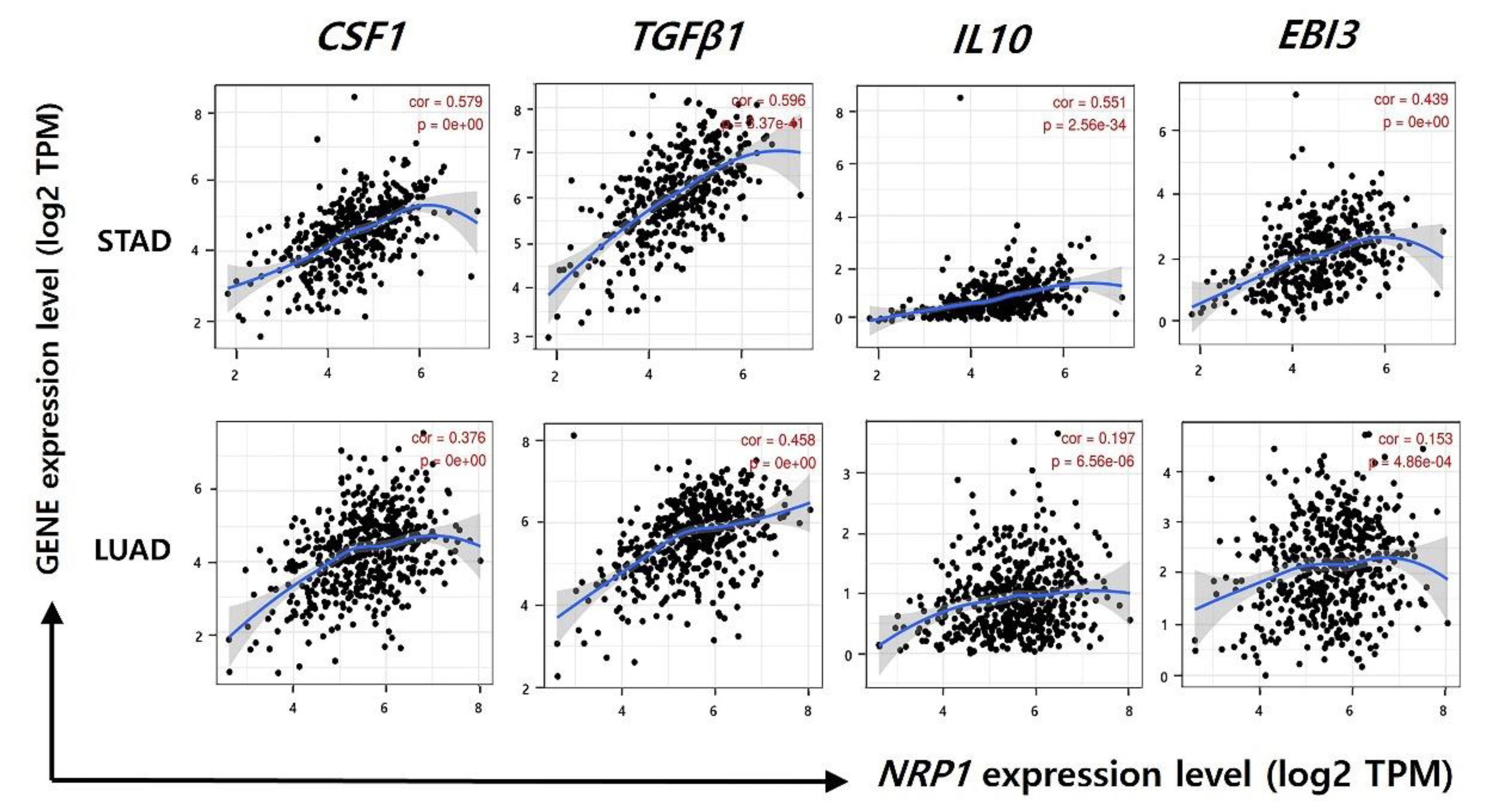
3.5. Correlation between NRP1 Expression and Gene Expression of Immune-Suppressive Cytokines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Abate, D.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdel-Rahman, O.; Abdelalim, A.; Abdoli, A.; Abdollahpour, I.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-Years for 29 Cancer Groups, 1990 to 2017: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Wu, Y.; Yang, J.; Yang, D.; Fang, X. Progress in the treatment of advanced gastric cancer. Tumour Biol. 2017, 39, 4626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chon, S.H.; Berlth, F.; Plum, P.S.; Herbold, T.; Alakus, H.; Kleinert, R.; Moenig, S.P.; Bruns, C.J.; Hoelscher, A.H.; Meyer, H.J. Gastric cancer treatment in the world: Germany. Transl. Gastroenterol. Hepatol. 2017, 2, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.; Gao, X.; Liu, H.; Jiang, T.; Zhai, X. Decreased expression of SEMA3A is associated with poor prognosis in gastric carcinoma. Int. J. Clin. Exp. Pathol. 2014, 7, 4782–4794. [Google Scholar] [PubMed]
- Ren, J.; Niu, G.; Wang, X.; Song, T.; Hu, Z.; Ke, C. Overexpression of FNDC1 in Gastric Cancer and its Prognostic Significance. J. Cancer 2018, 9, 4586–4595. [Google Scholar] [CrossRef]
- Jimenez-Hernandez, L.E.; Vazquez-Santillan, K.; Castro-Oropeza, R.; Martinez-Ruiz, G.; Munoz-Galindo, L.; Gonzalez-Torres, C.; Cortes-Gonzalez, C.C.; Victoria-Acosta, G.; Melendez-Zajgla, J.; Maldonado, V. NRP1-positive lung cancer cells possess tumor-initiating properties. Oncol. Rep. 2018, 39, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Tse, B.W.C.; Volpert, M.; Ratther, E.; Stylianou, N.; Nouri, M.; McGowan, K.; Lehman, M.L.; McPherson, S.J.; Roshan-Moniri, M.; Butler, M.S.; et al. Neuropilin-1 is upregulated in the adaptive response of prostate tumors to androgen-targeted therapies and is prognostic of metastatic progression and patient mortality. Oncogene 2017, 36, 3417–3427. [Google Scholar] [CrossRef] [Green Version]
- Geretti, E.; Klagsbrun, M. Neuropilins: Novel targets for anti-angiogenesis therapies. Cell Adhes. Migr. 2007, 1, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Barr, M.P.; Gray, S.G.; Gately, K.; Hams, E.; Fallon, P.G.; Davies, A.M.; Richard, D.J.; Pidgeon, G.P.; O’Byrne, K.J. Vascular endothelial growth factor is an autocrine growth factor, signaling through neuropilin-1 in non-small cell lung cancer. Mol. Cancer 2015, 14, 45. [Google Scholar] [CrossRef] [Green Version]
- Rizzolio, S.; Rabinowicz, N.; Rainero, E.; Lanzetti, L.; Serini, G.; Norman, J.; Neufeld, G.; Tamagnone, L. Neuropilin-1-dependent regulation of EGF-receptor signaling. Cancer Res. 2012, 72, 5801–5811. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Jiang, X.; Zhang, Q.; Dong, X.; Gao, Y.; He, Y.; Qiao, H.; Xie, F.; Xie, X.; Sun, X. Neuropilin-1 is associated with clinicopathology of gastric cancer and contributes to cell proliferation and migration as multifunctional co-receptors. J. Exp. Clin. Cancer Res. 2016, 35, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzog, B.; Pellet-Many, C.; Britton, G.; Hartzoulakis, B.; Zachary, I.C. VEGF binding to NRP1 is essential for VEGF stimulation of endothelial cell migration, complex formation between NRP1 and VEGFR2, and signaling via FAK Tyr407 phosphorylation. Mol. Biol. Cell 2011, 22, 2766–2776. [Google Scholar] [CrossRef] [PubMed]
- Akagi, M.; Kawaguchi, M.; Liu, W.; McCarty, M.F.; Takeda, A.; Fan, F.; Stoeltzing, O.; Parikh, A.A.; Jung, Y.D.; Bucana, C.D.; et al. Induction of neuropilin-1 and vascular endothelial growth factor by epidermal growth factor in human gastric cancer cells. Br. J. Cancer 2003, 88, 796–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhang, S.; Wang, Q.; Zhang, X. Tumor-recruited M2 macrophages promote gastric and breast cancer metastasis via M2 macrophage-secreted CHI3L1 protein. J. Hematol. Oncol. 2017, 10, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.J.; Wen, X.Y.; Yang, H.K.; Kim, W.H.; Kang, G.H. Prognostic Implication of M2 Macrophages Are Determined by the Proportional Balance of Tumor Associated Macrophages and Tumor Infiltrating Lymphocytes in Microsatellite-Unstable Gastric Carcinoma. PLoS ONE 2015, 10, e0144192. [Google Scholar] [CrossRef] [Green Version]
- Ao, J.Y.; Zhu, X.D.; Chai, Z.T.; Cai, H.; Zhang, Y.Y.; Zhang, K.Z.; Kong, L.Q.; Zhang, N.; Ye, B.G.; Ma, D.N.; et al. Colony-Stimulating Factor 1 Receptor Blockade Inhibits Tumor Growth by Altering the Polarization of Tumor-Associated Macrophages in Hepatocellular Carcinoma. Mol. Cancer Ther. 2017, 16, 1544–1554. [Google Scholar] [CrossRef] [Green Version]
- Chanmee, T.; Ontong, P.; Konno, K.; Itano, N. Tumor-associated macrophages as major players in the tumor microenvironment. Cancers 2014, 6, 1670–1690. [Google Scholar] [CrossRef] [Green Version]
- Paolino, M.; Penninger, J.M. The Role of TAM Family Receptors in Immune Cell Function: Implications for Cancer Therapy. Cancers 2016, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Traves, P.G.; Luque, A.; Hortelano, S. Macrophages, inflammation, and tumor suppressors: ARF, a new player in the game. Mediat. Inflamm. 2012, 2012, 568783. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Wu, F.; Zhou, Y.; Bao, Z.; Li, H.; Zheng, P.; Zhao, S. Gastric cancer-derived mesenchymal stromal cells trigger M2 macrophage polarization that promotes metastasis and EMT in gastric cancer. Cell Death Dis. 2019, 10, 918. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.Y.; Lin, Y.C.; Mahalingam, J.; Huang, C.T.; Chen, T.W.; Kang, C.W.; Peng, H.M.; Chu, Y.Y.; Chiang, J.M.; Dutta, A.; et al. Tumor-derived chemokine CCL5 enhances TGF-beta-mediated killing of CD8(+) T cells in colon cancer by T-regulatory cells. Cancer Res. 2012, 72, 1092–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, G.; Jones, E.; Junaid, S.; El-Shanawany, T.; Scurr, M.; Mizen, P.; Kumar, M.; Jones, S.; Rees, B.; Williams, G.; et al. Suppression of tumour-specific CD4(+) T cells by regulatory T cells is associated with progression of human colorectal cancer. Gut 2012, 61, 1163–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wei, F.Q.; Li, W.J.; Wei, J.W.; Zhong, H.; Wen, Y.H.; Lei, W.B.; Chen, L.; Li, H.; Lin, H.Q.; et al. A positive-feedback loop between tumour infiltrating activated Treg cells and type 2-skewed macrophages is essential for progression of laryngeal squamous cell carcinoma. Br. J. Cancer 2017, 117, 1631–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Huang, H.; Lu, J.; Bi, P.; Wang, F.; Liu, X.; Zhang, B.; Luo, Y.; Li, X. Tumor cells induced-M2 macrophage favors accumulation of Treg in nasopharyngeal carcinoma. Int. J. Clin. Exp. Pathol. 2017, 10, 8389–8401. [Google Scholar] [PubMed]
- Chaudhary, B.; Elkord, E. Novel expression of Neuropilin 1 on human tumor-infiltrating lymphocytes in colorectal cancer liver metastases. Expert Opin. Ther. Targets 2015, 19, 147–161. [Google Scholar] [CrossRef]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Oncomine Database Home Page. Available online: http://www.oncomine.org/ (accessed on 20 April 2020).
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [Green Version]
- Gene Expression Profiling Interactive Analysis Home Page. Available online: http://gepia.cancer-pku.cn/index.html/ (accessed on 21 April 2020).
- Pan, J.H.; Zhou, H.; Cooper, L.; Huang, J.L.; Zhu, S.B.; Zhao, X.X.; Ding, H.; Pan, Y.L.; Rong, L. LAYN Is a Prognostic Biomarker and Correlated with Immune Infiltrates in Gastric and Colon Cancers. Front. Immunol. 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Tumor IMmune Estimation Resource Home Page. Available online: http://cistrome.org/TIMER/ (accessed on 22 April 2020).
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- UALCAN Database Home Page. Available online: http://ualcan.path.uab.edu/index.html/ (accessed on 23 April 2020).
- Kaplan-Meier Survival Plotter Home Page. Available online: http://kmplot.com/analysis/ (accessed on 23 April 2020).
- Ranstam, J.; Cook, J.A. Kaplan-Meier curve. Br. J. Surg. 2017, 104, 442. [Google Scholar] [CrossRef] [PubMed]
- R2: Genomics Analysis and Visualization Platform Home Page. Available online: http://r2.amc.nl/ (accessed on 24 April 2020).
- Holcakova, J. Effect of DNA Methylation on the Development of Cancer. Klin. Onkol. 2018, 31, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hua, R.; Zhang, Y.; Tao, R.; Wang, Q.; Ni, Q. DNA hypomethylation promotes invasion and metastasis of gastric cancer cells by regulating the binding of SP1 to the CDCA3 promoter. J. Cell. Biochem. 2020, 121, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Shi, B.; Fu, Y.; Zhao, S.; Qu, K.; Guo, Q.; Li, K.; She, J. Hypomethylated gene NRP1 is co-expressed with PDGFRB and associated with poor overall survival in gastric cancer patients. Biomed. Pharmacother. 2019, 111, 1334–1341. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, J.; Zhang, L.; Wei, F.; Lian, Y.; Wu, Y.; Gong, Z.; Zhang, S.; Zhou, J.; Cao, K.; et al. Role of tumor microenvironment in tumorigenesis. J. Cancer 2017, 8, 761–773. [Google Scholar] [CrossRef]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Mantovani, A.; Marchesi, F.; Malesci, A.; Laghi, L.; Allavena, P. Tumour-associated macrophages as treatment targets in oncology. Nat. Rev. Clin. Oncol. 2017, 14, 399–416. [Google Scholar] [CrossRef]
- Maimela, N.R.; Liu, S.; Zhang, Y. Fates of CD8+ T cells in Tumor Microenvironment. Comput. Struct. Biotechnol. J. 2019, 17, 1–13. [Google Scholar] [CrossRef]
- Hao, N.B.; Lu, M.H.; Fan, Y.H.; Cao, Y.L.; Zhang, Z.R.; Yang, S.M. Macrophages in tumor microenvironments and the progression of tumors. Clin. Dev. Immunol. 2012, 2012, 948098. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.M.; Recht, L.; Strober, S. The Promise of Targeting Macrophages in Cancer Therapy. Clin. Cancer Res. 2017, 23, 3241–3250. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.Y. Regulatory T cells: Immune suppression and beyond. Cell. Mol. Immunol. 2010, 7, 204–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, H.; Sakaguchi, S. Regulatory T cells in tumor immunity. Int. J. Cancer 2010, 127, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Medina-Echeverz, J.; Fioravanti, J.; Zabala, M.; Ardaiz, N.; Prieto, J.; Berraondo, P. Successful colon cancer eradication after chemoimmunotherapy is associated with profound phenotypic change of intratumoral myeloid cells. J. Immunol. 2011, 186, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Pyonteck, S.M.; Akkari, L.; Schuhmacher, A.J.; Bowman, R.L.; Sevenich, L.; Quail, D.F.; Olson, O.C.; Quick, M.L.; Huse, J.T.; Teijeiro, V.; et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 2013, 19, 1264–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Cheng, Y.; Zhang, G.; Tang, Y.; Dong, Z.; McElwee, K.J.; Li, G. Increased expression of neuropilin 1 in melanoma progression and its prognostic significance in patients with melanoma. Mol. Med. Rep. 2015, 12, 2668–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, T.M.; Chen, Y.L.; Wu, Y.Y.; Yuan, A.; Chao, Y.C.; Chung, Y.C.; Wu, M.H.; Yang, S.C.; Pan, S.H.; Shih, J.Y.; et al. Targeting neuropilin 1 as an antitumor strategy in lung cancer. Clin. Cancer Res. 2007, 13, 4759–4768. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, T.; Kawakami, T.; Abe, Y.; Nishi, M.; Onoda, N.; Miyazaki, N.; Oida, Y.; Yamazaki, H.; Ueyama, Y.; Nakamura, M. The preserved expression of neuropilin (NRP) 1 contributes to a better prognosis in colon cancer. Oncol. Rep. 2006, 15, 369–373. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Lv, Y.; Qian, X.; Wang, S.; Chen, Z.; Jiang, Q.; Cao, C.; Song, Y. Inhibition of HHIP Promoter Methylation Suppresses Human Gastric Cancer Cell Proliferation and Migration. Cell. Physiol. Biochem. 2018, 45, 1840–1850. [Google Scholar] [CrossRef]
- Chen, H.Y.; Zhu, B.H.; Zhang, C.H.; Yang, D.J.; Peng, J.J.; Chen, J.H.; Liu, F.K.; He, Y.L. High CpG island methylator phenotype is associated with lymph node metastasis and prognosis in gastric cancer. Cancer Sci. 2012, 103, 73–79. [Google Scholar] [CrossRef]
- Deng, Y.B.; Nagae, G.; Midorikawa, Y.; Yagi, K.; Tsutsumi, S.; Yamamoto, S.; Hasegawa, K.; Kokudo, N.; Aburatani, H.; Kaneda, A. Identification of genes preferentially methylated in hepatitis C virus-related hepatocellular carcinoma. Cancer Sci. 2010, 101, 1501–1510. [Google Scholar] [CrossRef]
- Vivekanandhan, S.; Mukhopadhyay, D. Genetic status of KRAS influences Transforming Growth Factor-beta (TGF-beta) signaling: An insight into Neuropilin-1 (NRP1) mediated tumorigenesis. Semin. Cancer Biol. 2019, 54, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, A.M. VEGF/Neuropilin Signaling in Cancer Stem Cells. Int. J. Mol. Sci. 2019, 20, 490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziani, G.; Lacal, P.M. Neuropilin-1 as Therapeutic Target for Malignant Melanoma. Front. Oncol. 2015, 5, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhary, B.; Khaled, Y.S.; Ammori, B.J.; Elkord, E. Neuropilin 1: Function and therapeutic potential in cancer. Cancer Immunol. Immunother. 2014, 63, 81–99. [Google Scholar] [CrossRef]
- Wang, X.; Hu, H.; Liu, H. RNA binding protein Lin28B confers gastric cancer cells stemness via directly binding to NRP-1. Biomed. Pharmacother. 2018, 104, 383–389. [Google Scholar] [CrossRef]
- Rizzolio, S.; Cagnoni, G.; Battistini, C.; Bonelli, S.; Isella, C.; Van Ginderachter, J.A.; Bernards, R.; Di Nicolantonio, F.; Giordano, S.; Tamagnone, L. Neuropilin-1 upregulation elicits adaptive resistance to oncogene-targeted therapies. J. Clin. Investig. 2018, 128, 3976–3990. [Google Scholar] [CrossRef]
- Peng, Y.; Liu, Y.M.; Li, L.C.; Wang, L.L.; Wu, X.L. MicroRNA-338 inhibits growth, invasion and metastasis of gastric cancer by targeting NRP1 expression. PLoS ONE 2014, 9, e94422. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Lai, Z.; Li, Y.; Mu, J.; Yang, M.; Xie, J.; Xu, J. Immune signature profiling identified prognostic factors for gastric cancer. Chin. J. Cancer Res. 2019, 31, 463–470. [Google Scholar] [CrossRef]
- Napolitano, V.; Tamagnone, L. Neuropilins Controlling Cancer Therapy Responsiveness. Int. J. Mol. Sci. 2019, 20, 2049. [Google Scholar] [CrossRef] [Green Version]
- Waniczek, D.; Lorenc, Z.; Snietura, M.; Wesecki, M.; Kopec, A.; Muc-Wierzgon, M. Tumor-Associated Macrophages and Regulatory T Cells Infiltration and the Clinical Outcome in Colorectal Cancer. Arch. Immunol. Ther. Exp. 2017, 65, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Ino, Y.; Yamazaki-Itoh, R.; Shimada, K.; Iwasaki, M.; Kosuge, T.; Kanai, Y.; Hiraoka, N. Immune cell infiltration as an indicator of the immune microenvironment of pancreatic cancer. Br. J. Cancer 2013, 108, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Tong, L.; LiuAnwei Liu, F.; Liu, A.; Zeng, S.; Xiong, Q.; Yang, Z.; He, X.; Sun, Y.; Xu, C. Tumorinfiltrating M2 macrophages driven by specific genomic alterations are associated with prognosis in bladder cancer. Oncol. Rep. 2019, 42, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Paluskievicz, C.M.; Cao, X.; Abdi, R.; Zheng, P.; Liu, Y.; Bromberg, J.S. T Regulatory Cells and Priming the Suppressive Tumor Microenvironment. Front. Immunol. 2019, 10, 2453. [Google Scholar] [CrossRef] [PubMed]
- Shitara, K.; Nishikawa, H. Regulatory T cells: A potential target in cancer immunotherapy. Ann. N. Y. Acad. Sci. 2018, 1417, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Jackute, J.; Zemaitis, M.; Pranys, D.; Sitkauskiene, B.; Miliauskas, S.; Vaitkiene, S.; Sakalauskas, R. Distribution of M1 and M2 macrophages in tumor islets and stroma in relation to prognosis of non-small cell lung cancer. BMC Immunol. 2018, 19, 3. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Chikina, M.; Deshpande, R.; Menk, A.V.; Wang, T.; Tabib, T.; Brunazzi, E.A.; Vignali, K.M.; Sun, M.; Stolz, D.B.; et al. Treg Cells Promote the SREBP1-Dependent Metabolic Fitness of Tumor-Promoting Macrophages via Repression of CD8(+) T Cell-Derived Interferon-gamma. Immunity 2019, 51, 381–397.e6. [Google Scholar] [CrossRef]
- Ding, J.; Guo, C.; Hu, P.; Chen, J.; Liu, Q.; Wu, X.; Cao, Y.; Wu, J. CSF1 is involved in breast cancer progression through inducing monocyte differentiation and homing. Int. J. Oncol. 2016, 49, 2064–2074. [Google Scholar] [CrossRef] [Green Version]
- Miyauchi, J.T.; Chen, D.; Choi, M.; Nissen, J.C.; Shroyer, K.R.; Djordevic, S.; Zachary, I.C.; Selwood, D.; Tsirka, S.E. Ablation of Neuropilin 1 from glioma-associated microglia and macrophages slows tumor progression. Oncotarget 2016, 7, 9801–9814. [Google Scholar] [CrossRef] [Green Version]
- Forget, M.A.; Voorhees, J.L.; Cole, S.L.; Dakhlallah, D.; Patterson, I.L.; Gross, A.C.; Moldovan, L.; Mo, X.; Evans, R.; Marsh, C.B.; et al. Macrophage colony-stimulating factor augments Tie2-expressing monocyte differentiation, angiogenic function, and recruitment in a mouse model of breast cancer. PLoS ONE 2014, 9, e98623. [Google Scholar] [CrossRef]
- Zhu, X.D.; Zhang, J.B.; Zhuang, P.Y.; Zhu, H.G.; Zhang, W.; Xiong, Y.Q.; Wu, W.Z.; Wang, L.; Tang, Z.Y.; Sun, H.C. High expression of macrophage colony-stimulating factor in peritumoral liver tissue is associated with poor survival after curative resection of hepatocellular carcinoma. J. Clin. Oncol. 2008, 26, 2707–2716. [Google Scholar] [CrossRef]
- Huang, X.; Zou, Y.; Lian, L.; Wu, X.; He, X.; He, X.; Wu, X.; Huang, Y.; Lan, P. Changes of T cells and cytokines TGF-beta1 and IL-10 in mice during liver metastasis of colon carcinoma: Implications for liver anti-tumor immunity. J. Gastrointest. Surg. 2013, 17, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Collison, L.W.; Workman, C.J.; Kuo, T.T.; Boyd, K.; Wang, Y.; Vignali, K.M.; Cross, R.; Sehy, D.; Blumberg, R.S.; Vignali, D.A. The inhibitory cytokine IL-35 contributes to regulatory T-cell function. Nature 2007, 450, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Mai, J.; Virtue, A.; Yin, Y.; Gong, R.; Sha, X.; Gutchigian, S.; Frisch, A.; Hodge, I.; Jiang, X.; et al. IL-35 is a novel responsive anti-inflammatory cytokine—A new system of categorizing anti-inflammatory cytokines. PLoS ONE 2012, 7, e33628. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Gene Markers | STAD | LUAD | ||||||
|---|---|---|---|---|---|---|---|---|---|
| None | Purity | None | Purity | ||||||
| Cor | P | Cor | P | Cor | P | Cor | P | ||
| T cell (general) | CD3D | 0.306 | *** | 0.273 | *** | 0.143 | * | 0.101 | 0.024 |
| CD3E | 0.333 | *** | 0.39 | *** | 0.155 | ** | 0.117 | * | |
| CD2 | 0.382 | *** | 0.36 | *** | 0.168 | ** | 0.127 | * | |
| Treg cell | FOXP3 | 0.363 | *** | 0.336 | *** | 0.233 | *** | 0.203 | *** |
| CCR8 | 0.495 | *** | 0.482 | *** | 0.331 | *** | 0.317 | *** | |
| STAT5B | 0.579 | 0 | 0.574 | *** | 0.364 | *** | 0.358 | *** | |
| TGFβ (TGFB1) | 0.596 | *** | 0.576 | *** | 0.458 | 0 | 0.444 | *** | |
| TAM | CCL2 | 0.501 | 0 | 0.472 | *** | 0.247 | *** | 0.227 | *** |
| CD68 | 0.414 | 0 | 0.384 | *** | 0.256 | *** | 0.247 | ** | |
| IL10 | 0.551 | *** | 0.534 | *** | 0.197 | *** | 0.177 | * | |
| M1 Macrophage | INOS(NOS2) | −0.001 | 0.976 | −0.013 | 0.803 | 0.151 | ** | 0.127 | * |
| IRF5 | 0.313 | *** | 0.301 | *** | 0.206 | *** | 0.183 | *** | |
| COX2 (PTGS2) | 0.279 | *** | 0.258 | *** | 0.192 | *** | 0.207 | ** | |
| M2 Macrophage | CD163 | 0.662 | 0 | 0.642 | *** | 0.365 | *** | 0.363 | *** |
| VSIG4 | 0.607 | 0 | 0.601 | *** | 0.290 | *** | 0.279 | *** | |
| MS4A4A | 0.656 | 0 | 0.644 | *** | 0.309 | *** | 0.303 | *** | |
| CD8+ T cell | CD8A | 0.365 | *** | 0.339 | *** | 0.156 | ** | 0.129 | * |
| CD8B | 0.220 | *** | 0.202 | *** | 0.077 | 0.081 | 0.049 | 0.280 | |
| Neutrophil | CD66b(CEACAM8) | 0.102 | 0.036 | 0.115 | 0.025 | 0.193 | *** | 0.183 | *** |
| CD11b (ITGAM) | 0.614 | 0 | 0.598 | *** | 0.367 | *** | 0.357 | *** | |
| CCR7 | 0.427 | 0 | 0.403 | *** | 0.157 | ** | 0.119 | * | |
| NK cell | KIR2DL1 | 0.250 | *** | 0.256 | *** | 0.015 | 0.725 | −0.003 | 0.939 |
| KIR2DL3 | 0.180 | *** | 0.169 | *** | 0.089 | 0.043 | 0.069 | 0.123 | |
| KIR2DL4 | 0.113 | 0.02 | 0.086 | 0.091 | 0.068 | 0.122 | 0.044 | 0.324 | |
| KIR3DL1 | 0.219 | *** | 0.210 | *** | 0.021 | 0.629 | −0.011 | 0.810 | |
| KIR3DL2 | 0.204 | *** | 0.202 | *** | 0.122 | 0.005 | 0.100 | 0.026 | |
| KIR3DL3 | −0.001 | 0.991 | 0.022 | 0.667 | 0.064 | 0.145 | 0.042 | 0.352 | |
| KIR2DS4 | 0.162 | *** | 0.161 | * | 0.093 | 0.034 | 0.065 | 0.151 | |
| B cell | CD19 | 0.259 | *** | 0.230 | *** | −0.008 | 0.850 | −0.057 | 0.203 |
| CD79A | 0.263 | *** | 0.219 | *** | 0.040 | 0.369 | 0.004 | 0.925 | |
| Monocyte | CD86 | 0.575 | *** | 0.563 | *** | 0.340 | *** | 0.332 | *** |
| CD115 (CSF1R) | 0.664 | 0 | 0.647 | *** | 0.374 | *** | 0.357 | *** | |
| T cell exhaustion | PD1 (PDCD1) | 0.266 | *** | 0.245 | *** | 0.157 | ** | 0.127 | * |
| CTLA4 | 0.289 | *** | 0.265 | *** | 0.186 | *** | 0.152 | ** | |
| LAG3 | 0.249 | *** | 0.218 | *** | 0.076 | 0.086 | 0.043 | 0.343 | |
| TIM3 (HAVCR2) | 0.569 | 0 | 0.553 | *** | 0.309 | *** | 0.296 | *** | |
| Cell Type | Gene Markers | STAD | LUAD | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Tumor | Normal | Tumor | Normal | ||||||
| R | P | R | P | R | P | R | P | ||
| Treg | FOXP3 | 0.35 | *** | −0.36 | 0.03 | 0.21 | *** | 0.31 | 0.019 |
| CCR8 | 0.5 | *** | −0.22 | 0.19 | 0.33 | *** | 0.25 | 0.059 | |
| STAT5B | 0.62 | *** | 0.87 | *** | 0.37 | *** | 0.84 | *** | |
| TGFβ (TGFB1) | 0.59 | *** | 0.17 | 0.33 | 0.43 | *** | 0.43 | ** | |
| TAM | CCL2 | 0.48 | *** | 0.41 | 0.013 | 0.23 | *** | 0.12 | 0.36 |
| CD68 | 0.46 | *** | −0.32 | 0.057 | 0.29 | *** | 0.11 | 0.42 | |
| IL10 | 0.58 | *** | 0.2 | 0.25 | 0.21 | *** | 0.024 | 0.86 | |
| M1 Macrophage | INOS (NOS2) | 0.017 | 0.74 | 0.28 | 0.1 | 0.19 | *** | 0.62 | *** |
| IRF5 | 0.59 | *** | 0.67 | *** | 0.29 | *** | −0.19 | 0.15 | |
| COX2 (PTGS2) | 0.33 | *** | 0.77 | *** | 0.2 | *** | 0.21 | 0.11 | |
| M2 Macrophage | CD163 | 0.59 | *** | 0.67 | *** | 0.29 | *** | −0.19 | 0.15 |
| VSIG4 | 0.6 | *** | 0.64 | *** | 0.28 | *** | −0.2 | 0.12 | |
| MS4A4A | 0.64 | *** | 0.61 | *** | 0.31 | *** | −0.22 | 0.099 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.Y.; Gil, M.; Kim, K.E. Neuropilin1 Expression Acts as a Prognostic Marker in Stomach Adenocarcinoma by Predicting the Infiltration of Treg Cells and M2 Macrophages. J. Clin. Med. 2020, 9, 1430. https://doi.org/10.3390/jcm9051430
Kang JY, Gil M, Kim KE. Neuropilin1 Expression Acts as a Prognostic Marker in Stomach Adenocarcinoma by Predicting the Infiltration of Treg Cells and M2 Macrophages. Journal of Clinical Medicine. 2020; 9(5):1430. https://doi.org/10.3390/jcm9051430
Chicago/Turabian StyleKang, Ji Young, Minchan Gil, and Kyung Eun Kim. 2020. "Neuropilin1 Expression Acts as a Prognostic Marker in Stomach Adenocarcinoma by Predicting the Infiltration of Treg Cells and M2 Macrophages" Journal of Clinical Medicine 9, no. 5: 1430. https://doi.org/10.3390/jcm9051430