Lactate and BDNF: Key Mediators of Exercise Induced Neuroplasticity?


1
Research Group Neuroprotection, German Center for Neurodegenerative Diseases (DZNE), Leipziger Str. 44, 39120 Magdeburg, Germany
2
Medical Faculty, Department of Neurology, Otto-von-Guericke University, Leipziger Str. 44, 39120 Magdeburg, Germany
3
Medical Faculty, Institute of Physiology, Otto-von-Guericke University, Leipziger Str. 44, 39120 Magdeburg, Germany
4
Center for Behavioral Brain Sciences (CBBS), Leipziger Str. 44, 39120 Magdeburg, Germany
*
Author to whom correspondence should be addressed.
J. Clin. Med. 2020, 9(4), 1136; https://doi.org/10.3390/jcm9041136
Submission received: 19 March 2020
/
Revised: 7 April 2020
/
Accepted: 12 April 2020
/
Published: 15 April 2020
(This article belongs to the Special Issue Effects of Lifestyle Factors and Exercise on Brain Plasticity and Underlying Neurobiological Mechanisms)
Abstract
:Accumulating evidence from animal and human studies supports the notion that physical exercise can enhance neuroplasticity and thus reduce the risk of several neurodegenerative diseases (e.g., dementia). However, the underlying neurobiological mechanisms of exercise induced neuroplasticity are still largely unknown. One potential mediator of exercise effects is the neurotrophin BDNF, which enhances neuroplasticity via different pathways (e.g., synaptogenesis, neurogenesis, long-term potentiation). Current research has shown that (i) increased peripheral lactate levels (following high intensity exercise) are associated with increased peripheral BDNF levels, (ii) lactate infusion at rest can increase peripheral and central BDNF levels and (iii) lactate plays a very complex role in the brain’s metabolism. In this review, we summarize the role and relationship of lactate and BDNF in exercise induced neuroplasticity.
1. Introduction
Physical activity and/or exercise (Table 1) is a low-cost intervention in primary and secondary prevention for numerous chronical diseases (e.g., diabetes, cancer, cardiovascular diseases, sarcopenia) [1]. Furthermore, physical activity has several positive effects on brain health, can stimulate neuroplasticity and reduce the risk of neurodegenerative diseases (e.g., dementia) [2,3,4,5,6]. Especially in the context of an unavailable causal pharmacological treatment for dementia, research is currently focusing on modifiable risk and lifestyle factors that can delay the outbreak of the disease or ameliorate associated memory deficits [7,8]. In western societies physical inactivity is one central modifiable risk factor for dementia [9] and thus target for interventions. In the last decades, the impact of exercise on brain health has become a central research project in neuroscience and several lines of evidence suggest that BDNF is one of the essential mediating factors of exercise induced neuroplasticity. However, the underlying neurobiological and molecular mechanisms of exercise induced neuroplasticity are still largely unknown [10]. Interestingly, recent results from animal and human research suggest that lactate might link physical exercise to BDNF-dependent neurobiological pathways [11,12,13,14].
2. BDNF and Neuroplasticity
The protein brain-derived neurotrophic factor [BDNF] belongs to the family of neurotrophins (NT), which in mammals also comprise nerve growth factor [NGF], neurotrophin 3 [NT-3] and NT-4/5. Neurotrophins crucially regulate important neurobiological processes such as neurogenesis, synaptogenesis, growth of dendritic spines, long-term potentiation [LTP] and efficacy of protein synthesis [19,20,21,22,23,24]. BDNF is synthesized in the endoplasmic reticulum (ER) as a pre-cursor protein (calculated molecular weight 28 kDa). After initial cleavage of the pre-sequence, the resulting proBDNF (calculated molecular weight 26 kDa) undergoes further processing (e.g., glycosylation) in the ER and in the Golgi apparatus (reviewed in [25]), including partial endoproteolytic cleavage of the pro-form which eventually yields mature BDNF (mBDNF; commonly termed as BDNF; calculated molecular weight 13.5 kDa). Accordingly, proBDNF and mBDNF are co-stored in vesicles that bud-off from the trans-Golgi network and are transported to the cell membrane where they undergo exocytosis, thus leading to secretion of proBDNF, mBDNF and smaller cleavage products into the extracellular space. These vesicles belong either to the constitutive or the regulated secretory pathway (see below). However, the exact composition of BDNF species in secreted vesicles is yet unknown. Released proBDNF can be converted in the extracellular space by endoproteases to mBDNF [25]. While mBDNF binds selectively to the tyrosin-related kinase receptor B [TrkB], proBDNF binds specifically to the so-called p75 neurotrophin receptor [p75NTR] of target cell membranes. Because of the antagonistic effects of proBDNF/p75 signaling (mediating apoptosis, shrinkage of dendritic spines, and long-term depression [LTD] compared to mBDNF/TrkB signaling pathways, (supporting neuronal survival, synaptogenesis, growth of dendritic spines, and LTP) [26] the proportion of released proBDNF and mBDNF critically determines the direction of neuroplasticity that can be elicited in target neurons (for further information on BDNF synthesis, processing, and expression see [27,28]). Activation of TrkB triggers intracellular signal cascades such as mitogen-activated protein kinase (MAPK), phospholipase C-γ (PLCγ) or phosphatidylinositol-3-kinase (PI3K) pathways [29]. Additionally, TrkB activation can also increase the expression of the peroxisome proliferator-activated receptor γ co-activator α (PGC1α) which in turn increases BDNF expression in neurons via the PGC1α/FNDC5/BDNF pathway [30,31].
As mentioned above, BDNF release can take place via two classes of vesicles. The Golgi-derived vesicles of the constitutive pathway of secretion undergo exocytosis by default when they reach the plasma membrane. In contrast, activity-dependent release of BDNF (e.g., driven by repetitive firing of action potentials) from Golgi-derived vesicles of the regulated pathway of secretion requires sustained intracellular Ca2+ elevations [32]. In the brain, most of the released BDNF is secreted via the regulated pathway [23,25]. Importantly, under physiological conditions, intracellular BDNF protein levels are extraordinarily low in many brain regions. In rodents, highest levels are observed in the hippocampal CA3 and dentate gyrus area, the amygdala, and selected regions of the cerebral cortex, including the visual and somatosensory cortex [27,28,33,34,35]. The low expression level and the largely activity-dependent and locally restricted release give BDNF a strong command in fine-tuning cellular functions with high spatial selectivity [23,25].
In the central nervous system (CNS) only excitatory glutamatergic (but not inhibitory GABAergic) neurons have the capacity to synthesize BDNF. However, GABAergic neurons are dependent on extracellular BDNF for survival and synaptogenesis [24,36]. In addition, microglial cells, T and B lymphocytes, monocytes and skeletal muscle cells can synthesize and release BDNF [27,37,38]. Thrombocytes cannot synthesize, but seem to take up and store BDNF via receptor-mediated endocytosis [39,40,41]. BDNF can pass the blood-brain barrier [42] and about 75% of the peripheral BDNF plasma level originates from the brain [43,44]. This is the reason why blood plasma (and serum) BDNF levels are considered to serve as a proxy for released BDNF in the brain and can thus be used to investigate the effect of lifestyle interventions (e.g., physical exercise, caloric-restricted diet) on BDNF-mediated neuroplasticity in the brain.
BDNF mediates neuroplasticity via different mechanisms and on distinct time scales [27,29]. Thus, BDNF/TrkB signaling induces within seconds to minutes LTP at glutamatergic synaptic connections [45,46] and the growth of new synaptic spines [47,48], thereby enabling the initial formation of new memory traces at existing synapses. On the time scale of hours, BDNF contributes to consolidation of protein synthesis dependent long lasting LTP and memory formation, and on even longer time scales shapes memory engrams by incorporating newborn neurons into neuronal circuits by promoting neuro- and synaptogenesis. In these ways, BDNF signaling crucially contributes to cellular mechanisms of neuronal plasticity that drive formation, consolidation and retrieval of memory.
In humans, BDNF has been associated with psychiatric [49] (e.g., schizophrenia, major depressive disorder, anxiety disorders) and neurological diseases (e.g., dementia [50,51], Huntington‘s disease [52]). Reduced BDNF levels have been reported in the hippocampus [53] and in the blood [54,55,56] in patients with mild cognitive impairment (MCI) or Alzheimer‘s disease (AD). In contrast, some studies have found no association of BDNF and AD [57]. Based on a community-based, prospective cohort study with 2131 dementia-free participants Weinstein et al., proposed that higher serum BDNF levels may protect against dementia [58]. However, these cross-sectional data do not allow a causal interpretation. A common (observed in approx. 30% of humans) single nucleotide polymorphism (SNP) in the BDNF gene that leads to a substitution of valine to methionine in the pro-region has been associated with reduced activity-dependent BDNF secretion [59]. Nonetheless, a higher incidence of the above mentioned neurological and neuropsychiatric disorders in carriers of the SNP has thus far remained elusive. This might at least in part result from the physical exercise-dependent regulation of BDNF protein expression and release (see below) that might overcome the decreased basal secretion of Val66Met BDNF [59].
3. Physical Exercise and BDNF
Physical exercise can improve cognition (e.g., executive functions, spatial memory, learning) [60,61,62,63,64] and induce structural and functional brain plasticity [65,66,67,68,69,70,71,72,73,74,75,76]. Epidemiological, observational and intervention studies indicate that exercising can reduce the risk of neurodegenerative diseases (e.g., dementia [2,4,77], multiple sclerosis [78], Parkinson’s disease [79,80]). Proposed mechanisms of exercise induced neuroplasticity are increased expression, secretion, and downstream signaling of neurotrophic factors (e.g., BDNF, VEGF, IGF-1), reduced inflammation [81,82], reduced stress levels [83] and improved cardiovascular (e.g., reduced arterial stiffness, reduced blood pressure) [84,85] and metabolic (e.g., insulin sensitivity) [86,87] parameters. Numerous studies have shown that exercise induced neuroplasticity is associated with BDNF [88,89]. Indeed, at least in rodents, pharmacological blocking of BDNF signaling in the hippocampus attenuates the neuroplastic effects of physical exercise [90].
Neeper et al., were the first to report a positive correlation between physical activity and BDNF mRNA levels in rodents [91]. In animal models, short and long term periods of exercise increase BDNF gene and protein expression in the hippocampus [92,93] and other brain structures (e.g., amygdala [94], cerebellum [92], perirhinal cortex [95]). However, the effect of exercise on BDNF expression is smaller in aged animals compared to young ones [96]. Furthermore, Choi et al., reported that exercise induced adult hippocampal neurogenesis is associated with improved cognition, reduced ß-amyloid in the brain and increased levels of BDNF in an AD mouse model [97]. Similar to rodents, physical activity increases peripheral BDNF levels in healthy humans [98,99], and numerous studies have shown a positive impact of acute exercise on BDNF plasma [43,100,101] and/or serum levels [102,103]. Similarly, also chronic exercise increases BDNF plasma [104,105] and/or serum levels [106]. Several reviews and meta-analyses have investigated the effects of physical exercise on BDNF blood levels [98,99,107,108,109,110,111]. Overall, they strongly suggest that acute [43,112,113] and chronic [99,114,115] exercise can increase peripheral BDNF levels in humans. Regarding acute exercise the duration and intensity of exercise correlate with larger increases in BDNF [112,113]. Additionally, effect sizes in women seem to be significantly smaller after acute exercise [116]. In contrast, no gender effect was reported following chronic exercise [115]. A recent meta-analysis reported that aerobic exercise alone does not increase BDNF in older adults while resistance exercise and combined aerobic/resistance exercise increases peripheral BDNF levels [117].
4. Physical Exercise, Lactate and BDNF
Several trials have used blood lactate for the monitoring of exercise intensity. These studies indicate that higher lactate concentrations are associated with increased BDNF plasma and/or serum levels [102,118,119]. Furthermore, current evidence indicates that high intensity interval training evokes larger BDNF levels compared to moderate and/or intensive continuous exercise [110,112].
For long, lactate was simply considered a waste product of the anaerobic metabolism. Nowadays, however, it is clear that lactate is an important signaling molecule that is involved in several metabolic processes [14,120]. Energy supply for exercise is based mainly on three pathways: (i) ATP-Creatinkinase, (ii) glycolysis and (iii) oxidative phosphorylation [121]. Lactate is produced by glucose oxidation when oxygen uptake is low, and it can buffer acidosis. Accumulated lactate can be transported to the liver (where lactate is synthesized to glucose through gluconeogenesis) or can be directly used as a fuel by muscles, heart and brain [122]. During acute exercise lactate accumulates depending on the intensity and the duration of the exercise. The lactate threshold (also called anaerobic threshold) is defined as the highest level of physical activity that can be achieved without lactate accumulation and is a predictor of an individual’s fitness level. Physical exercise can improve the fitness level and can increase the lactate threshold [123].
Lactate can cross the blood-brain barrier (BBB) [120] reaching neurons via monocarboxylate transporters (MCT) [124,125,126]. MCT 2 is the major transporter in neurons [127] while MCT 4 is only expressed in astrocytes [128]. Astrocytes have complex interactions with neurons. They are involved in the control of cell volumes, energy metabolism and ionic homeostasis [129]. Astrocytes show a glucose gradient with high glucose concentrations close to the BBB and low glucose concentrations close to neurons. This gradient allows a rapid glucose transfer to neurons. Furthermore, astrocytes can store glycogen and support the neuronal energy metabolism [126,130,131].
Additionally, Pellerin et al., proposed an astrocyte-neuronal lactate transport during excitatory neurotransmission [132]. Here lactate is transported from astrocytes to neurons through MCTs where lactate is converted to pyruvate and enters the tricarboxylic acid cycle. Lactate in neurons can origin from astrocyte metabolism or from peripheral muscle activity. Moreover, neurons in vitro prefer lactate instead of glucose [133]. Current research indicates that lactate transport from astrocytes to neurons plays a crucial role for memory formation [134,135,136] and could be a link between exercise and neuroplasticity [120]. Pharmacological inhibition of MCT 2 irreversibly impairs long-term memory [136]. Van de Hall et al. [125] have shown that lactate uptake in the brain increases from 8% at rest up to 20% during exercise. Additionally, Kemppainen et al. [137] reported a higher lactate metabolism in trained healthy adults compared to controls. In rodents a single bout of exercise can induce up-regulation of MCTs [138]. Proia et al., hypothesized that exercise can increase levels of BDNF and of other growth factors such as insulin-like growth factor 1 (IGF-1) [120] and vascular endothelial growth factor (VEGF) [139,140,141].
However, the interaction between lactate and BDNF levels is not yet well resolved. Potential mechanisms that link both molecules could be (i) a lactate regulated increased NMDA-receptor activation and as a consequence increased intracellular calcium levels, (ii) a signaling cascade initiated by lactate binding to different G-protein coupled receptors (GPCR), and (iii) through silent information regulator 1 (SIRT1) activation of the PGC1α/FNDC5/BDNF pathway (Figure 1).
- (i)
- Yang et al., reported that lactate promotes plasticity related gene expression by potentiating NMDA glutamate receptor activity in neurons [142]. Furthermore, lactate increases intracellular NADH and calcium levels. This could be a central mechanism for neuroplasticity induced by lactate from astrocytes. Additionally, increased intracellular calcium following lactate induced enhanced NMDA receptor activity could be a link between exercise and BDNF expression. [25].
- (ii)
- Lactate can bind to GPCR81 (also known as hydroxycarboxylic acid receptor [HCAR1]) on neurons [143] and at the BBB [144]. Lauritzen et al., have shown, that HCAR1 at the BBB is essential for mediating exercise effects on angiogenesis in a mouse model [144]. Furthermore, lactate binding to HCAR1 on neurons inhibits the adenylate cyclase and thus decreases cAMP, resulting in reduced neuronal activity and gene regulation [14,143,145]. Here, lactate could have a metabolic and regulatory function in the control of blood flow and synaptic function [146]. Furthermore, lactate can influence prostaglandin E2 uptake and thus influence vasodilation [147]. The potential negative modulation of BDNF production by lactate through HCAR1 should be examined more closely in the future.
- (iii)
- Lactate can induce the PGC1α/FNDC5/BDNF pathway through SIRT1activation [11]. El Hayek et al., have shown, that voluntary exercise promotes hippocampal BDNF expression and improves memory and learning in a lactate-dependent manner in rodents [11]. Furthermore, they have shown that intraperitoneal lactate infusion in mice induces SIRT1 activity and thus enhances the PGC1α/FNDC5/BDNF pathway which results in improved spatial learning and memory retention.
Schiffer et al. [12] investigated whether lactate infusion at rest can elevate BDNF blood concentration in young adults. The lactate clamp method is a well-established method to examine (neuro-) physiological effects of lactate without stimulating physical exercise. After infusion of a 4 molar sodium-lactate solution, BDNF serum and lactate levels increase significantly and returned to baseline values at follow-up [12]. Potential mechanisms of BDNF serum increase after lactate infusion could be (i) a lactate driven BDNF expression or (ii) a release of BDNF from platelets (in context of blood gases disturbance).
5. Conclusions
Physical exercise induces numerous metabolic adaptations. However, most studies have investigated the effects of acute and/or chronic exercise on brain plasticity and underlying mechanism with a limited test battery only (e.g., neuropsychological tests, magnetic resonance imaging [MRI] and BDNF). To better understand the neurobiological mechanism induced by exercise, more extensive assessments are necessary. For instance, research studies in rodents and/or humans have shown that exercise can enhance cathepsin B [148], FNDC5/Irisin [31], lactate and BDNF, while the mechanisms interconnecting these observations are yet not understood. Future studies are urgently needed that combine animal and human exercise research with extensive test batteries to foster our understanding on how exercise can induce neuroplasticity. Identifying the underlying molecular mechanisms will help to design (i) more tailored exercise interventions [149] and (ii) pharmacological agents that can mimic the effects of physical exercise [150].
Author Contributions
P.M. drafted the manuscript, which was critically reviewed by Y.D., V.L. and N.G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the EU Joint Program-Neurodegenerative Disease Research (JPND) project CIRCPROT jointly funded by the BMBF and EU Horizon 2020 (project no. 643417, to VL).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AD | Alzheimer’s disease |
| BBB | blood-brain barrier |
| BDNF | brain-derived neurotrophic factor |
| CNS | central nervous system |
| HCAR1 | hydroxycarboxylic acid receptor 1 |
| ER | endoplasmic reticulum |
| FNDC5 | fibronectin type III domain-containing 5 |
| LTD | long-term depression |
| LTP | long-term potentiation |
| GPCR | G protein coupled receptors |
| MAPK | mitogen-activated protein kinase |
| mBDNF | mature brain-derived neurotrophic factor |
| MCI | mild cognitive impairment |
| MCT | monocarboxylate transporter |
| MET | metabolic equivalent of task |
| NGF | nerve growth factor |
| NMDA | N-Methyl-d-aspartate |
| NT-3 | neurotrophin-3 |
| NT-4/5 | neurotrophin-4/5 |
| PGC1α | peroxisome proliferator-activated receptor γ co-activator α |
| PI3K | phosphatidylinositol-3-kinase |
| PLCγ | phospholipase C-γ |
| SIRT1 | silent information regulator 1 |
| SNP | single nucleotide polymorphism |
| TrkB | tyrosine-kinase receptor B |
References
- Warburton, D.E.R.; Nicol, C.W.; Bredin, S.S.D. Health benefits of physical activity: The evidence. CMAJ 2006, 174, 801–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlskog, J.E.; Geda, Y.E.; Graff-Radford, N.R.; Petersen, R.C. Physical exercise as a preventive or disease-modifying treatment of dementia and brain aging. Mayo Clin. Proc. 2011, 86, 876–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deslandes, A.; Moraes, H.; Ferreira, C.; Veiga, H.; Silveira, H.; Mouta, R.; Pompeu, F.A.M.S.; Coutinho, E.S.F.; Laks, J. Exercise and mental health: Many reasons to move. Neuropsychobiology 2009, 59, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Hamer, M.; Chida, Y. Physical activity and risk of neurodegenerative disease: A systematic review of prospective evidence. Psychol. Med. 2009, 39, 3–11. [Google Scholar] [CrossRef]
- Liu-Ambrose, T.; Barha, C.K.; Best, J.R. Physical activity for brain health in older adults. Appl. Physiol. Nutr. Metab. 2018, 43, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Hökelmann, A.; Müller, N.G. The impact of physical activities on age-related brain function and structure and the underlying neural mechanisms. In Physical Activity and Educational Achievement: Insights from Exercise Neuroscience; Meeusen, R., Schaefer, S., Tomporowski, P.D., Bailey, R., Eds.; Routledge Taylor & Francis Group: London, UK, 2018; pp. 164–177. [Google Scholar]
- Kivipelto, M.; Mangialasche, F.; Ngandu, T. Lifestyle interventions to prevent cognitive impairment, dementia and Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 653–666. [Google Scholar] [CrossRef]
- Müller, P.; Fendt, M.; Müller, N.G. Pharmakologische Therapie der Alzheimer-Demenz: Aktueller Stand und Perspektiven. Internist 2019, 60, 761–768. [Google Scholar] [CrossRef]
- Norton, S.; Matthews, F.E.; Barnes, D.E.; Yaffe, K.; Brayne, C. Potential for primary prevention of Alzheimer’s disease: An analysis of population-based data. Lancet Neurol. 2014, 13, 788–794. [Google Scholar] [CrossRef] [Green Version]
- Kempermann, G. Neurodegenerative Erkrankungen und zelluläre Plastizität als sportmedizinische Herausforderung. Dtsch. Z. Sportmed. 2015, 2015, 31–35. [Google Scholar] [CrossRef]
- El Hayek, L.; Khalifeh, M.; Zibara, V.; Abi Assaad, R.; Emmanuel, N.; Karnib, N.; El-Ghandour, R.; Nasrallah, P.; Bilen, M.; Ibrahim, P.; et al. Lactate Mediates the Effects of Exercise on Learning and Memory through SIRT1-Dependent Activation of Hippocampal Brain-Derived Neurotrophic Factor (BDNF). J. Neurosci. 2019, 39, 2369–2382. [Google Scholar] [CrossRef] [Green Version]
- Schiffer, T.; Schulte, S.; Sperlich, B.; Achtzehn, S.; Fricke, H.; Strüder, H.K. Lactate infusion at rest increases BDNF blood concentration in humans. Neurosci. Lett. 2011, 488, 234–237. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.I.; Smith, L.; Northey, J.; Rattray, B.; Cherbuin, N. Towards an understanding of the physical activity-BDNF-cognition triumvirate: A review of associations and dosage. Ageing Res. Rev. 2020, 101044. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P.J.; Allaman, I. Lactate in the brain: From metabolic end-product to signalling molecule. Nat. Rev. Neurosci. 2018, 19, 235–249. [Google Scholar] [CrossRef]
- Caspersen, C.J.; Powell, K.E.; Christenson, G.M. Physical activity, exercise, and physical fitness: Definitions and distinctions for health-related research. Public Health Rep. 1985, 100, 126–131. [Google Scholar] [PubMed]
- Herold, F.; Müller, P.; Gronwald, T.; Müller, N.G. Dose-Response Matters! A Perspective on the Exercise Prescription in Exercise-Cognition Research. Front. Psychol. 2019, 10, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskell, W.L.; Lee, I.-M.; Pate, R.R.; Powell, K.E.; Blair, S.N.; Franklin, B.A.; Macera, C.A.; Heath, G.W.; Thompson, P.D.; Bauman, A. Physical activity and public health: Updated recommendation for adults from the American College of Sports Medicine and the American Heart Association. Circulation 2007, 116, 1081–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, M.E.; Rejeski, W.J.; Blair, S.N.; Duncan, P.W.; Judge, J.O.; King, A.C.; Macera, C.A.; Castaneda-Sceppa, C. Physical activity and public health in older adults: Recommendation from the American College of Sports Medicine and the American Heart Association. Circulation 2007, 116, 1094–1105. [Google Scholar] [CrossRef] [Green Version]
- Barde, Y.A.; Edgar, D.; Thoenen, H. Purification of a new neurotrophic factor from mammalian brain. EMBO J. 1982, 1, 549–553. [Google Scholar] [CrossRef]
- Thoenen, H. Neurotrophins and neuronal plasticity. Science 1995, 270, 593–598. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.-M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- Lewin, G.R.; Barde, Y.A. Physiology of the neurotrophins. Ann. Rev. Neurosci. 1996, 19, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Edelmann, E.; Lessmann, V.; Brigadski, T. Pre-and postsynaptic twists in BDNF secretion and action in synaptic plasticity. Neuropharmacology 2014, 76, 610–627. [Google Scholar] [CrossRef]
- Gottmann, K.; Mittmann, T.; Lessmann, V. BDNF signaling in the formation, maturation and plasticity of glutamatergic and GABAergic synapses. Exp. Brain Res. 2009, 199, 203–234. [Google Scholar] [CrossRef] [PubMed]
- Lessmann, V.; Brigadski, T. Mechanisms, locations, and kinetics of synaptic BDNF secretion: An update. Neurosci. Res. 2009, 65, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1545–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brigadski, T.; Lessmann, V. BDNF: A regulator of learning and memory processes with clinical potential. e Neuroforum 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Sasi, M.; Vignoli, B.; Canossa, M.; Blum, R. Neurobiology of local and intercellular BDNF signaling. Pflug. Arch. 2017, 469, 593–610. [Google Scholar] [CrossRef] [Green Version]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in neuronal development and function. Ann. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [Green Version]
- Jodeiri Farshbaf, M.; Ghaedi, K.; Megraw, T.L.; Curtiss, J.; Shirani Faradonbeh, M.; Vaziri, P.; Nasr-Esfahani, M.H. Does PGC1α/FNDC5/BDNF Elicit the Beneficial Effects of Exercise on Neurodegenerative Disorders? Neuromolecular Med. 2016, 18, 1–15. [Google Scholar] [CrossRef]
- Wrann, C.D.; White, J.P.; Salogiannnis, J.; Laznik-Bogoslavski, D.; Wu, J.; Di, M.; Lin, J.; Greenberg, M.E.; Spiegelman, B.M. Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway. Cell Metab. 2013, 18, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Kolarow, R.; Brigadski, T.; Lessmann, V. Postsynaptic secretion of BDNF and NT-3 from hippocampal neurons depends on calcium calmodulin kinase II signaling and proceeds via delayed fusion pore opening. J. Neurosci. 2007, 27, 10350–10364. [Google Scholar] [CrossRef] [PubMed]
- Conner, J.M.; Lauterborn, J.C.; Yan, Q.; Gall, C.M.; Varon, S. Distribution of Brain-Derived Neurotrophic Factor (BDNF) Protein and mRNA in the Normal Adult Rat CNS: Evidence for Anterograde Axonal Transport. J. Neurosci. 1997, 17, 2295–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauskolb, S.; Zagrebelsky, M.; Dreznjak, A.; Deogracias, R.; Matsumoto, T.; Wiese, S.; Erne, B.; Sendtner, M.; Schaeren-Wiemers, N.; Korte, M.; et al. Global deprivation of brain-derived neurotrophic factor in the CNS reveals an area-specific requirement for dendritic growth. J. Neurosci. 2010, 30, 1739–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leschik, J.; Eckenstaler, R.; Endres, T.; Munsch, T.; Edelmann, E.; Richter, K.; Kobler, O.; Fischer, K.-D.; Zuschratter, W.; Brigadski, T.; et al. Prominent Postsynaptic and Dendritic Exocytosis of Endogenous BDNF Vesicles in BDNF-GFP Knock-in Mice. Mol. Neurobiol. 2019, 56, 6833–6855. [Google Scholar] [CrossRef] [PubMed]
- Kohara, K.; Yasuda, H.; Huang, Y.; Adachi, N.; Sohya, K.; Tsumoto, T. A local reduction in cortical GABAergic synapses after a loss of endogenous brain-derived neurotrophic factor, as revealed by single-cell gene knock-out method. J. Neurosci. 2007, 27, 7234–7244. [Google Scholar] [CrossRef] [PubMed]
- Trang, T.; Beggs, S.; Wan, X.; Salter, M.W. P2 × 4-receptor-mediated synthesis and release of brain-derived neurotrophic factor in microglia is dependent on calcium and p38-mitogen-activated protein kinase activation. J. Neurosci. 2009, 29, 3518–3528. [Google Scholar] [CrossRef]
- Matthews, V.B.; Aström, M.-B.; Chan, M.H.S.; Bruce, C.R.; Krabbe, K.S.; Prelovsek, O.; Akerström, T.; Yfanti, C.; Broholm, C.; Mortensen, O.H.; et al. Brain-derived neurotrophic factor is produced by skeletal muscle cells in response to contraction and enhances fat oxidation via activation of AMP-activated protein kinase. Diabetologia 2009, 52, 1409–1418. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, H.; Altar, C.A.; Chen, R.; Nakamura, T.; Nakahashi, T.; Kambayashi, J.-I.; Sun, B.; Tandon, N.N. Brain-derived neurotrophic factor is stored in human platelets and released by agonist stimulation. Thromb. Haemost. 2002, 87, 728–734. [Google Scholar] [CrossRef] [Green Version]
- Karege, F.; Bondolfi, G.; Gervasoni, N.; Schwald, M.; Aubry, J.-M.; Bertschy, G. Low brain-derived neurotrophic factor (BDNF) levels in serum of depressed patients probably results from lowered platelet BDNF release unrelated to platelet reactivity. Biol. Psychiatry 2005, 57, 1068–1072. [Google Scholar] [CrossRef]
- Chacón-Fernández, P.; Säuberli, K.; Colzani, M.; Moreau, T.; Ghevaert, C.; Barde, Y.-A. Brain-derived Neurotrophic Factor in Megakaryocytes. J. Biol. Chem. 2016, 291, 9872–9881. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Banks, W.A.; Fasold, M.B.; Bluth, J.; Kastin, A.J. Transport of brain-derived neurotrophic factor across the blood-brain barrier. Neuropharmacology 1998, 37, 1553–1561. [Google Scholar] [CrossRef]
- Rasmussen, P.; Brassard, P.; Adser, H.; Pedersen, M.V.; Leick, L.; Hart, E.; Secher, N.H.; Pedersen, B.K.; Pilegaard, H. Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp. Physiol. 2009, 94, 1062–1069. [Google Scholar] [CrossRef] [PubMed]
- Krabbe, K.S.; Nielsen, A.R.; Krogh-Madsen, R.; Plomgaard, P.; Rasmussen, P.; Erikstrup, C.; Fischer, C.P.; Lindegaard, B.; Petersen, A.M.W.; Taudorf, S.; et al. Brain-derived neurotrophic factor (BDNF) and type 2 diabetes. Diabetologia 2007, 50, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Edelmann, E.; Cepeda-Prado, E.; Franck, M.; Lichtenecker, P.; Brigadski, T.; Lessmann, V. Theta Burst Firing Recruits BDNF Release and Signaling in Postsynaptic CA1 Neurons in Spike-Timing-Dependent LTP. Neuron 2015, 86, 1041–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korte, M.; Carroll, P.; Wolf, E.; Brem, G.; Thoenen, H.; Bonhoeffer, T. Hippocampal long-term potentiation is impaired in mice lacking brain-derived neurotrophic factor. Proc. Natl. Acad. Sci. USA 1995, 92, 8856–8860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, J.-I.; Horiike, Y.; Matsuzaki, M.; Miyazaki, T.; Ellis-Davies, G.C.R.; Kasai, H. Protein synthesis and neurotrophin-dependent structural plasticity of single dendritic spines. Science 2008, 319, 1683–1687. [Google Scholar] [CrossRef] [Green Version]
- Harward, S.C.; Hedrick, N.G.; Hall, C.E.; Parra-Bueno, P.; Milner, T.A.; Pan, E.; Laviv, T.; Hempstead, B.L.; Yasuda, R.; McNamara, J.O. Autocrine BDNF-TrkB signalling within a single dendritic spine. Nature 2016, 538, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [Green Version]
- Holsinger, R.M.D.; Schnarr, J.; Henry, P.; Castelo, V.T.; Fahnestock, M. Quantitation of BDNF mRNA in human parietal cortex by competitive reverse transcription-polymerase chain reaction: Decreased levels in Alzheimer’s disease. Mol. Brain Res. 2000, 76, 347–354. [Google Scholar] [CrossRef]
- Tapia-Arancibia, L.; Aliaga, E.; Silhol, M.; Arancibia, S. New insights into brain BDNF function in normal aging and Alzheimer disease. Brain Res. Rev. 2008, 59, 201–220. [Google Scholar] [CrossRef]
- Zuccato, C.; Cattaneo, E. Role of brain-derived neurotrophic factor in Huntington’s disease. Prog. Neurobiol. 2007, 81, 294–330. [Google Scholar] [CrossRef]
- Peng, S.; Wuu, J.; Mufson, E.J.; Fahnestock, M. Precursor form of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinical stages of Alzheimer’s disease. J. Neurochem. 2005, 93, 1412–1421. [Google Scholar] [CrossRef]
- Laske, C.; Stransky, E.; Leyhe, T.; Eschweiler, G.W.; Maetzler, W.; Wittorf, A.; Soekadar, S.; Richartz, E.; Koehler, N.; Bartels, M.; et al. BDNF serum and CSF concentrations in Alzheimer’s disease, normal pressure hydrocephalus and healthy controls. J. Psychiatr. Res. 2007, 41, 387–394. [Google Scholar] [CrossRef]
- Lee, J.G.; Shin, B.S.; You, Y.S.; Kim, J.E.; Yoon, S.W.; Jeon, D.W.; Baek, J.H.; Park, S.W.; Kim, Y.H. Decreased serum brain-derived neurotrophic factor levels in elderly korean with dementia. Psychiatry Investig. 2009, 6, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Forlenza, O.V.; Diniz, B.S.; Teixeira, A.L.; Ojopi, E.B.; Talib, L.L.; Mendonça, V.A.; Izzo, G.; Gattaz, W.F. Effect of brain-derived neurotrophic factor Val66Met polymorphism and serum levels on the progression of mild cognitive impairment. World J. Biol. Psychiatry 2010, 11, 774–780. [Google Scholar] [CrossRef] [PubMed]
- O’Bryant, S.E.; Hobson, V.L.; Hall, J.R.; Barber, R.C.; Zhang, S.; Johnson, L.; Diaz-Arrastia, R. Serum brain-derived neurotrophic factor levels are specifically associated with memory performance among Alzheimer’s disease cases. Dement. Geriatr. Cogn. Disord. 2011, 31, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, G.; Beiser, A.S.; Choi, S.H.; Preis, S.R.; Chen, T.C.; Vorgas, D.; Au, R.; Pikula, A.; Wolf, P.A.; DeStefano, A.L.; et al. Serum brain-derived neurotrophic factor and the risk for dementia: The Framingham Heart Study. JAMA Neurol. 2014, 71, 55–61. [Google Scholar] [CrossRef]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met Polymorphism Affects Activity-Dependent Secretion of BDNF and Human Memory and Hippocampal Function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Colcombe, S.; Kramer, A.F. Fitness effects on the cognitive function of older adults: A meta-analytic study. Psychol. Sci. 2003, 14, 125–130. [Google Scholar] [CrossRef]
- Dresler, M.; Sandberg, A.; Ohla, K.; Bublitz, C.; Trenado, C.; Mroczko-Wąsowicz, A.; Kühn, S.; Repantis, D. Non-pharmacological cognitive enhancement. Neuropharmacology 2013, 64, 529–543. [Google Scholar] [CrossRef]
- Duzel, E.; van Praag, H.; Sendtner, M. Can physical exercise in old age improve memory and hippocampal function? Brain 2016, 139, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Schwarck, S.; Schmicker, M.; Dordevic, M.; Rehfeld, K.; Müller, N.; Müller, P. Inter-Individual Differences in Cognitive Response to a Single Bout of Physical Exercise-A Randomized Controlled Cross-Over Study. J. Clin. Med. 2019, 8, 1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dordevic, M.; Hökelmann, A.; Müller, P.; Rehfeld, K.; Müller, N.G. Improvements in Orientation and Balancing Abilities in Response to One Month of Intensive Slackline-Training. A Randomized Controlled Feasibility Study. Front. Hum. Neurosci. 2017, 11, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sexton, C.E.; Betts, J.F.; Demnitz, N.; Dawes, H.; Ebmeier, K.P.; Johansen-Berg, H. A systematic review of MRI studies examining the relationship between physical fitness and activity and the white matter of the ageing brain. Neuroimage 2016, 131, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berchicci, M.; Lucci, G.; Di Russo, F. Benefits of physical exercise on the aging brain: The role of the prefrontal cortex. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1337–1341. [Google Scholar] [CrossRef] [Green Version]
- Colcombe, S.J.; Erickson, K.I.; Scalf, P.E.; Kim, J.S.; Prakash, R.; McAuley, E.; Elavsky, S.; Marquez, D.X.; Hu, L.; Kramer, A.F. Aerobic exercise training increases brain volume in aging humans. J. Gerontol. A Biol. Sci. Med. Sci. 2006, 61, 1166–1170. [Google Scholar] [CrossRef] [Green Version]
- Erickson, K.I.; Leckie, R.L.; Weinstein, A.M. Physical activity, fitness, and gray matter volume. Neurobiol. Aging 2014, 35, S20–S28. [Google Scholar] [CrossRef] [Green Version]
- Voelcker-Rehage, C.; Niemann, C. Structural and functional brain changes related to different types of physical activity across the life span. Neurosci. Biobehav. Rev. 2013, 37, 2268–2295. [Google Scholar] [CrossRef]
- Thomas, A.G.; Dennis, A.; Bandettini, P.A.; Johansen-Berg, H. The effects of aerobic activity on brain structure. Front. Psychol. 2012, 3, 86. [Google Scholar] [CrossRef] [Green Version]
- Dordevic, M.; Taubert, M.; Müller, P.; Kaufmann, J.; Hökelmann, A.; Müller, N.G. Brain Gray Matter Volume Is Modulated by Visual Input and Overall Learning Success but Not by Time Spent on Learning a Complex Balancing Task. J. Clin. Med. 2018, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Dordevic, M.; Schrader, R.; Taubert, M.; Müller, P.; Hökelmann, A.; Müller, N.G. Vestibulo-Hippocampal Function Is Enhanced and Brain Structure Altered in Professional Ballet Dancers. Front. Integr. Neurosci. 2018, 12, 50. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, K.; Müller, P.; Aye, N.; Schmicker, M.; Dordevic, M.; Kaufmann, J.; Hökelmann, A.; Müller, N.G. Dancing or Fitness Sport? The Effects of Two Training Programs on Hippocampal Plasticity and Balance Abilities in Healthy Seniors. Front. Hum. Neurosci. 2017, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Dordevic, M.; Taubert, M.; Müller, P.; Riemer, M.; Kaufmann, J.; Hökelmann, A.; Müller, N.G. Which Effects on Neuroanatomy and Path-Integration Survive? Results of a Randomized Controlled Study on Intensive Balance Training. Brain Sci. 2020, 10, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, C.; Zou, L.; Mei, J.; Moore, D.; Herold, F.; Müller, P.; Yu, Q.; Liu, Y.; Lin, J.; Tao, Y.; et al. Tai Chi Training Evokes Significant Changes in Brain White Matter Network in Older Women. Healthcare 2020, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Yue, C.; Zhang, Y.; Jian, M.; Herold, F.; Yu, Q.; Mueller, P.; Lin, J.; Wang, G.; Tao, Y.; Zhang, Z.; et al. Differential Effects of Tai Chi Chuan (Motor-Cognitive Training) and Walking on Brain Networks: A Resting-State fMRI Study in Chinese Women Aged 60. Healthcare 2020, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Müller, P.; Schmicker, M.; Müller, N.G. Präventionsstrategien gegen Demenz. Z. Gerontol. Geriatr. 2017, 50, 89–95. [Google Scholar] [CrossRef]
- Dorans, K.S.; Massa, J.; Chitnis, T.; Ascherio, A.; Munger, K.L. Physical activity and the incidence of multiple sclerosis. Neurology 2016, 87, 1770–1776. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Park, Y.; Huang, X.; Hollenbeck, A.; Blair, A.; Schatzkin, A.; Chen, H. Physical activities and future risk of Parkinson disease. Neurology 2010, 75, 341–348. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.M.; Schwarzschild, M.A.; Hernán, M.A.; Ascherio, A. Physical activity and the risk of Parkinson disease. Neurology 2005, 64, 664–669. [Google Scholar] [CrossRef]
- Ryan, S.M.; Nolan, Y.M. Neuroinflammation negatively affects adult hippocampal neurogenesis and cognition: Can exercise compensate? Neurosci. Biobehav. Rev. 2016, 61, 121–131. [Google Scholar] [CrossRef]
- Packer, N.; Pervaiz, N.; Hoffman-Goetz, L. Does exercise protect from cognitive decline by altering brain cytokine and apoptotic protein levels? A systematic review of the literature. Exerc. Immunol. Rev. 2010, 16, 138–162. [Google Scholar] [PubMed]
- Tsatsoulis, A.; Fountoulakis, S. The protective role of exercise on stress system dysregulation and comorbidities. Ann. N. Y. Acad. Sci. 2006, 1083, 196–213. [Google Scholar] [CrossRef] [PubMed]
- Lessiani, G.; Santilli, F.; Boccatonda, A.; Iodice, P.; Liani, R.; Tripaldi, R.; Saggini, R.; Davì, G. Arterial stiffness and sedentary lifestyle: Role of oxidative stress. Vascul. Pharmacol. 2016, 79, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hillman, C.H.; Weiss, E.P.; Hagberg, J.M.; Hatfield, B.D. The relationship of age and cardiovascular fitness to cognitive and motor processes. Psychophysiology 2002, 39, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, S.; Cholerton, B.; Baker, L.D. Insulin and Alzheimer’s disease: Untangling the web. J. Alzheimers Dis. 2013, 33, S263–S275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cholerton, B.; Baker, L.D.; Craft, S. Insulin resistance and pathological brain ageing. Diabet. Med. 2011, 28, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Stillman, C.M.; Cohen, J.; Lehman, M.E.; Erickson, K.I. Mediators of Physical Activity on Neurocognitive Function: A Review at Multiple Levels of Analysis. Front. Hum. Neurosci. 2016, 10, 626. [Google Scholar] [CrossRef] [Green Version]
- Cotman, C.W.; Berchtold, N.C.; Christie, L.-A. Exercise builds brain health: Key roles of growth factor cascades and inflammation. Trends Neurosci. 2007, 30, 464–472. [Google Scholar] [CrossRef]
- Vaynman, S.; Ying, Z.; Gomez-Pinilla, F. Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. Eur. J. Neurosci. 2004, 20, 2580–2590. [Google Scholar] [CrossRef]
- Neeper, S.A.; Gómez-Pinilla, F.; Choi, J.; Cotman, C. Exercise and brain neurotrophins. Nature 1995, 373, 109. [Google Scholar] [CrossRef]
- Neeper, S.A.; Gómez-Pinilla, F.; Choi, J.; Cotman, C.W. Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res. 1996, 726, 49–56. [Google Scholar] [CrossRef]
- Berchtold, N.C.; Castello, N.; Cotman, C.W. Exercise and time-dependent benefits to learning and memory. Neuroscience 2010, 167, 588–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-F.; Chen, H.-I.; Wu, C.-L.; Kuo, Y.-M.; Yu, L.; Huang, A.-M.; Wu, F.-S.; Chuang, J.-I.; Jen, C.J. Differential effects of treadmill running and wheel running on spatial or aversive learning and memory: Roles of amygdalar brain-derived neurotrophic factor and synaptotagmin I. J. Physiol. 2009, 587, 3221–3231. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.E.; Bucci, D.J. BDNF expression in perirhinal cortex is associated with exercise-induced improvement in object recognition memory. Neurobiol. Learn. Mem. 2010, 94, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Adlard, P.A.; Perreau, V.M.; Cotman, C.W. The exercise-induced expression of BDNF within the hippocampus varies across life-span. Neurobiol. Aging 2005, 26, 511–520. [Google Scholar] [CrossRef]
- Choi, S.H.; Bylykbashi, E.; Chatila, Z.K.; Lee, S.W.; Pulli, B.; Clemenson, G.D.; Kim, E.; Rompala, A.; Oram, M.K.; Asselin, C.; et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 2018, 361. [Google Scholar] [CrossRef] [Green Version]
- Zoladz, J.A.; Pilc, A. The effect of physical activity on the brain derived neurotrophic factor: From animal to human studies. J. Physiol. Pharmacol. 2010, 61, 533–541. [Google Scholar]
- Knaepen, K.; Goekint, M.; Heyman, E.M.; Meeusen, R. Neuroplasticity-exercise-induced response of peripheral brain-derived neurotrophic factor: A systematic review of experimental studies in human subjects. Sports Med. 2010, 40, 765–801. [Google Scholar] [CrossRef]
- Seifert, T.; Brassard, P.; Wissenberg, M.; Rasmussen, P.; Nordby, P.; Stallknecht, B.; Adser, H.; Jakobsen, A.H.; Pilegaard, H.; Nielsen, H.B.; et al. Endurance training enhances BDNF release from the human brain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R372–R377. [Google Scholar] [CrossRef] [Green Version]
- Zoladz, J.A.; Pilc, A.; Majerczak, J.; Grandys, M.; Zapart-Bukowska, J.; Duda, K. Endurance training increases plasma brain-derived neurotrophic factor concentration in young healthy men. J. Physiol. Pharmacol. 2008, 59, 119–132. [Google Scholar]
- Ferris, L.T.; Williams, J.S.; Shen, C.-L. The effect of acute exercise on serum brain-derived neurotrophic factor levels and cognitive function. Med. Sci. Sports Exerc. 2007, 39, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Voss, M.W.; Prakash, R.S.; Basak, C.; Szabo, A.; Chaddock, L.; Kim, J.S.; Heo, S.; Alves, H.; White, S.M.; et al. Exercise training increases size of hippocampus and improves memory. Proc. Natl. Acad. Sci. USA 2011, 108, 3017–3022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, P.; Rehfeld, K.; Schmicker, M.; Hökelmann, A.; Dordevic, M.; Lessmann, V.; Brigadski, T.; Kaufmann, J.; Müller, N.G. Evolution of Neuroplasticity in Response to Physical Activity in Old Age: The Case for Dancing. Front. Aging Neurosci. 2017, 9, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfeld, K.; Lüders, A.; Hökelmann, A.; Lessmann, V.; Kaufmann, J.; Brigadski, T.; Müller, P.; Müller, N.G. Dance training is superior to repetitive physical exercise in inducing brain plasticity in the elderly. PLoS ONE 2018, 13, e0196636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maass, A.; Düzel, S.; Brigadski, T.; Goerke, M.; Becke, A.; Sobieray, U.; Neumann, K.; Lövdén, M.; Lindenberger, U.; Bäckman, L.; et al. Relationships of peripheral IGF-1, VEGF and BDNF levels to exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 2016, 131, 142–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Larsen, K.T.; Ried-Larsen, M.; Møller, N.C.; Andersen, L.B. The effects of physical activity and exercise on brain-derived neurotrophic factor in healthy humans: A review. Scand. J. Med. Sci. Sports 2014, 24, 1–10. [Google Scholar] [CrossRef]
- Coelho, F.G.D.M.; Gobbi, S.; Andreatto, C.A.A.; Corazza, D.I.; Pedroso, R.V.; Santos-Galduróz, R.F. Physical exercise modulates peripheral levels of brain-derived neurotrophic factor (BDNF): A systematic review of experimental studies in the elderly. Arch. Gerontol. Geriatr. 2013, 56, 10–15. [Google Scholar] [CrossRef]
- Enette, L.; Vogel, T.; Fanon, J.L.; Lang, P.O. Effect of Interval and Continuous Aerobic Training on Basal Serum and Plasma Brain-Derived Neurotrophic Factor Values in Seniors: A Systematic Review of Intervention Studies. Rejuvenation Res. 2017, 20, 473–483. [Google Scholar] [CrossRef]
- Jiménez-Maldonado, A.; Rentería, I.; García-Suárez, P.C.; Moncada-Jiménez, J.; Freire-Royes, L.F. The Impact of High-Intensity Interval Training on Brain Derived Neurotrophic Factor in Brain: A Mini-Review. Front. Neurosci. 2018, 12, 839. [Google Scholar] [CrossRef]
- Zimmer, P.; Oberste, M.; Bloch, W. Einfluss von Sport auf das zentrale Nervensystem-Molekulare und zelluläre Wirkmechanismen. Dtsch. Z. Sportmed. 2015, 2015, 42–49. [Google Scholar] [CrossRef]
- Saucedo Marquez, C.M.; Vanaudenaerde, B.; Troosters, T.; Wenderoth, N. High-intensity interval training evokes larger serum BDNF levels compared with intense continuous exercise. J. Appl. Physiol. 2015, 119, 1363–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinoff, A.; Herrmann, N.; Swardfager, W.; Lanctôt, K.L. The effect of acute exercise on blood concentrations of brain-derived neurotrophic factor in healthy adults: A meta-analysis. Eur. J. Neurosci. 2017, 46, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Heisz, J.J.; Clark, I.B.; Bonin, K.; Paolucci, E.M.; Michalski, B.; Becker, S.; Fahnestock, M. The Effects of Physical Exercise and Cognitive Training on Memory and Neurotrophic Factors. J. Cogn. Neurosci. 2017, 29, 1895–1907. [Google Scholar] [CrossRef] [PubMed]
- Dinoff, A.; Herrmann, N.; Swardfager, W.; Liu, C.S.; Sherman, C.; Chan, S.; Lanctôt, K.L. The Effect of Exercise Training on Resting Concentrations of Peripheral Brain-Derived Neurotrophic Factor (BDNF): A Meta-Analysis. PLoS ONE 2016, 11, e0163037. [Google Scholar] [CrossRef]
- Szuhany, K.L.; Bugatti, M.; Otto, M.W. A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J. Psychiatr. Res. 2015, 60, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Marinus, N.; Hansen, D.; Feys, P.; Meesen, R.; Timmermans, A.; Spildooren, J. The Impact of Different Types of Exercise Training on Peripheral Blood Brain-Derived Neurotrophic Factor Concentrations in Older Adults: A Meta-Analysis. Sports Med. 2019, 49, 1529–1546. [Google Scholar] [CrossRef]
- Gold, S.M.; Schulz, K.-H.; Hartmann, S.; Mladek, M.; Lang, U.E.; Hellweg, R.; Reer, R.; Braumann, K.-M.; Heesen, C. Basal serum levels and reactivity of nerve growth factor and brain-derived neurotrophic factor to standardized acute exercise in multiple sclerosis and controls. J. Neuroimmunol. 2003, 138, 99–105. [Google Scholar] [CrossRef]
- Rojas Vega, S.; Strüder, H.K.; Vera Wahrmann, B.; Schmidt, A.; Bloch, W.; Hollmann, W. Acute BDNF and cortisol response to low intensity exercise and following ramp incremental exercise to exhaustion in humans. Brain Res. 2006, 1121, 59–65. [Google Scholar] [CrossRef]
- Proia, P.; Di Liegro, C.M.; Schiera, G.; Fricano, A.; Di Liegro, I. Lactate as a Metabolite and a Regulator in the Central Nervous System. Int. J. Mol. Sci. 2016, 17, 1450. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Brown, A.M.; Frontera, W.R. Principles of exercise physiology: Responses to acute exercise and long-term adaptations to training. PM R 2012, 4, 797–804. [Google Scholar] [CrossRef]
- Adeva-Andany, M.; López-Ojén, M.; Funcasta-Calderón, R.; Ameneiros-Rodríguez, E.; Donapetry-García, C.; Vila-Altesor, M.; Rodríguez-Seijas, J. Comprehensive review on lactate metabolism in human health. Mitochondrion 2014, 17, 76–100. [Google Scholar] [CrossRef]
- Brooks, G.A. Anaerobic threshold: Review of the concept and directions for future research. Med. Sci. Sports Exerc. 1985, 17, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Bergersen, L.H. Is lactate food for neurons? Comparison of monocarboxylate transporter subtypes in brain and muscle. Neuroscience 2007, 145, 11–19. [Google Scholar] [CrossRef] [PubMed]
- van Hall, G.; Strømstad, M.; Rasmussen, P.; Jans, O.; Zaar, M.; Gam, C.; Quistorff, B.; Secher, N.H.; Nielsen, H.B. Blood lactate is an important energy source for the human brain. J. Cereb. Blood Flow Metab. 2009, 29, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Camandola, S.; Mattson, M.P. Brain metabolism in health, aging, and neurodegeneration. EMBO J. 2017, 36, 1474–1492. [Google Scholar] [CrossRef]
- Pierre, K.; Magistretti, P.J.; Pellerin, L. MCT2 is a major neuronal monocarboxylate transporter in the adult mouse brain. J. Cereb. Blood Flow Metab. 2002, 22, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Pellerin, L.; Bergersen, L.H.; Halestrap, A.P.; Pierre, K. Cellular and subcellular distribution of monocarboxylate transporters in cultured brain cells and in the adult brain. J. Neurosci. Res. 2005, 79, 55–64. [Google Scholar] [CrossRef]
- Pannasch, U.; Rouach, N. Emerging role for astroglial networks in information processing: From synapse to behavior. Trends Neurosci. 2013, 36, 405–417. [Google Scholar] [CrossRef]
- Dienel, G.A. The metabolic trinity, glucose-glycogen-lactate, links astrocytes and neurons in brain energetics, signaling, memory, and gene expression. Neurosci. Lett. 2017, 637, 18–25. [Google Scholar] [CrossRef]
- Dienel, G.A.; Cruz, N.F. Contributions of glycogen to astrocytic energetics during brain activation. Metab. Brain Dis. 2015, 30, 281–298. [Google Scholar] [CrossRef] [Green Version]
- Pellerin, L.; Magistretti, P.J. Glutamate uptake into astrocytes stimulates aerobic glycolysis: A mechanism coupling neuronal activity to glucose utilization. Proc. Natl. Acad. Sci. USA 1994, 91, 10625–10629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouzier-Sore, A.-K.; Voisin, P.; Bouchaud, V.; Bezancon, E.; Franconi, J.-M.; Pellerin, L. Competition between glucose and lactate as oxidative energy substrates in both neurons and astrocytes: A comparative NMR study. Eur. J. Neurosci. 2006, 24, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, M.Q.; Gao, V.; Alberini, C.M. The Role of Lactate-Mediated Metabolic Coupling between Astrocytes and Neurons in Long-Term Memory Formation. Front. Integr. Neurosci. 2016, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Newman, L.A.; Korol, D.L.; Gold, P.E. Lactate produced by glycogenolysis in astrocytes regulates memory processing. PLoS ONE 2011, 6, e28427. [Google Scholar] [CrossRef]
- Kemppainen, J.; Aalto, S.; Fujimoto, T.; Kalliokoski, K.K.; Långsjö, J.; Oikonen, V.; Rinne, J.; Nuutila, P.; Knuuti, J. High intensity exercise decreases global brain glucose uptake in humans. J. Physiol. 2005, 568, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Takimoto, M.; Hamada, T. Acute exercise increases brain region-specific expression of MCT1, MCT2, MCT4, GLUT1, and COX IV proteins. J. Appl. Physiol. 2014, 116, 1238–1250. [Google Scholar] [CrossRef] [Green Version]
- Lista, I.; Sorrentino, G. Biological mechanisms of physical activity in preventing cognitive decline. Cell. Mol. Neurobiol. 2010, 30, 493–503. [Google Scholar] [CrossRef]
- Schwarz, A.J.; Brasel, J.A.; Hintz, R.L.; Mohan, S.; Cooper, D.M. Acute effect of brief low-and high-intensity exercise on circulating insulin-like growth factor (IGF) I, II, and IGF-binding protein-3 and its proteolysis in young healthy men. J. Clin. Endocrinol. Metab. 1996, 81, 3492–3497. [Google Scholar] [CrossRef] [Green Version]
- Skriver, K.; Roig, M.; Lundbye-Jensen, J.; Pingel, J.; Helge, J.W.; Kiens, B.; Nielsen, J.B. Acute exercise improves motor memory: Exploring potential biomarkers. Neurobiol. Learn. Mem. 2014, 116, 46–58. [Google Scholar] [CrossRef]
- Yang, J.; Ruchti, E.; Petit, J.-M.; Jourdain, P.; Grenningloh, G.; Allaman, I.; Magistretti, P.J. Lactate promotes plasticity gene expression by potentiating NMDA signaling in neurons. Proc. Natl. Acad. Sci. USA 2014, 111, 12228–12233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morland, C.; Lauritzen, K.H.; Puchades, M.; Holm-Hansen, S.; Andersson, K.; Gjedde, A.; Attramadal, H.; Storm-Mathisen, J.; Bergersen, L.H. The lactate receptor, G-protein-coupled receptor 81/hydroxycarboxylic acid receptor 1: Expression and action in brain. J. Neurosci. Res. 2015, 93, 1045–1055. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, K.H.; Morland, C.; Puchades, M.; Holm-Hansen, S.; Hagelin, E.M.; Lauritzen, F.; Attramadal, H.; Storm-Mathisen, J.; Gjedde, A.; Bergersen, L.H. Lactate receptor sites link neurotransmission, neurovascular coupling, and brain energy metabolism. Cereb. Cortex 2014, 24, 2784–2795. [Google Scholar] [CrossRef] [PubMed]
- Bozzo, L.; Puyal, J.; Chatton, J.-Y. Lactate modulates the activity of primary cortical neurons through a receptor-mediated pathway. PLoS ONE 2013, 8, e71721. [Google Scholar] [CrossRef]
- Barros, L.F. Metabolic signaling by lactate in the brain. Trends Neurosci. 2013, 36, 396–404. [Google Scholar] [CrossRef]
- Gordon, G.R.J.; Choi, H.B.; Rungta, R.L.; Ellis-Davies, G.C.R.; MacVicar, B.A. Brain metabolism dictates the polarity of astrocyte control over arterioles. Nature 2008, 456, 745–749. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.Y.; Becke, A.; Berron, D.; Becker, B.; Sah, N.; Benoni, G.; Janke, E.; Lubejko, S.T.; Greig, N.H.; Mattison, J.A.; et al. Running-Induced Systemic Cathepsin B Secretion Is Associated with Memory Function. Cell Metab. 2016, 24, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Müllers, P.; Taubert, M.; Müller, N.G. Physical Exercise as Personalized Medicine for Dementia Prevention? Front. Physiol. 2019, 10, 672. [Google Scholar] [CrossRef]
- Guerrieri, D.; Moon, H.Y.; van Praag, H. Exercise in a Pill: The Latest on Exercise-Mimetics. Brain Plast. 2017, 2, 153–169. [Google Scholar] [CrossRef] [Green Version]
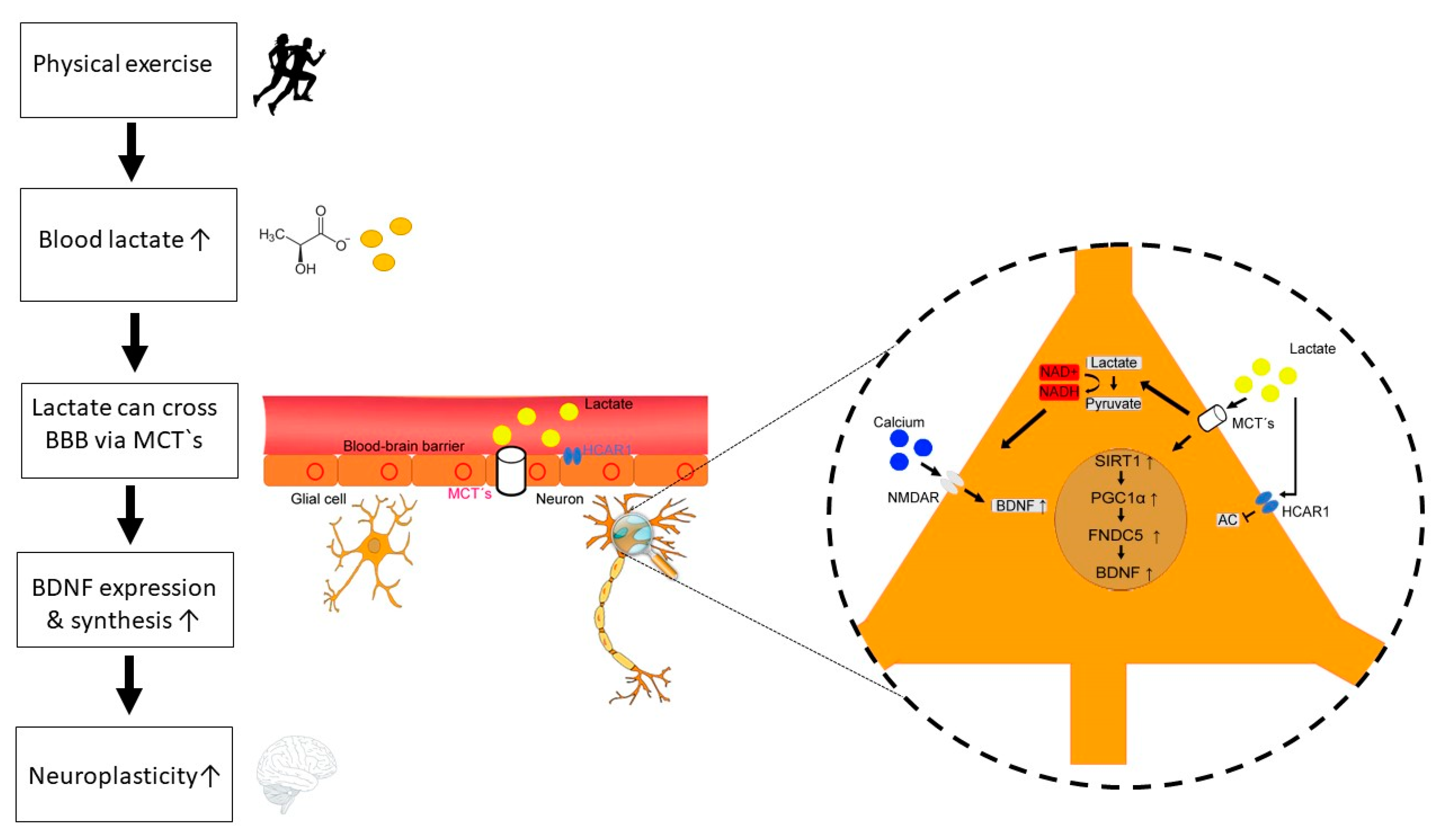
Figure 1.
Potential mechanisms of lactate-BDNF interaction following physical exercise. Physical exercise is associated with intensity-dependent increase of lactate levels. Lactate can cross the blood-brain barrier (BBB) via different monocarboxylate transporters (MCT‘s). Furthermore, lactate binding to the hydroxycarboxylic acid receptor (HCAR1) at the BBB can induce angiogenesis. In neurons, lactate exerts several neurotrophic and metabolic effects through transmembrane transport via MCT’s and direct binding to HCAR1. Firstly, lactate binding to HCAR1 on neurons inhibits the adenylate cyclase (AC) and thus decreases cAMP, resulting reduced BDNF expression and regulatory function in the control of blood flow, and synaptic functions. Secondly, lactate can induce the PGC1α/FNDC5/BDNF pathway through SIRT1 activation. Thirdly, lactate increases intracellular NADH, resulting in enhanced calcium levels and BDNF gene expression. Released BDNF can then enhance neuroplasticity via different neurobiological mechanisms (e.g., neurogenesis, synaptogenesis, growth of dendritic spines, long-term potentiation [LTP]).
Figure 1.
Potential mechanisms of lactate-BDNF interaction following physical exercise. Physical exercise is associated with intensity-dependent increase of lactate levels. Lactate can cross the blood-brain barrier (BBB) via different monocarboxylate transporters (MCT‘s). Furthermore, lactate binding to the hydroxycarboxylic acid receptor (HCAR1) at the BBB can induce angiogenesis. In neurons, lactate exerts several neurotrophic and metabolic effects through transmembrane transport via MCT’s and direct binding to HCAR1. Firstly, lactate binding to HCAR1 on neurons inhibits the adenylate cyclase (AC) and thus decreases cAMP, resulting reduced BDNF expression and regulatory function in the control of blood flow, and synaptic functions. Secondly, lactate can induce the PGC1α/FNDC5/BDNF pathway through SIRT1 activation. Thirdly, lactate increases intracellular NADH, resulting in enhanced calcium levels and BDNF gene expression. Released BDNF can then enhance neuroplasticity via different neurobiological mechanisms (e.g., neurogenesis, synaptogenesis, growth of dendritic spines, long-term potentiation [LTP]).


{kind=link}
Table 1.
Physical activity and physical exercise.
| Physical Activity and Physical Exercise-Where is the Difference? |
|---|
| “Physical activity”’ is defined as any muscle-induced bodily movement which increases energy expenditure above ~1.0/1.5 metabolic equivalent of task (MET, 1 MET = 1 kcal (4184 kJ) × kg−1 × h−1) whereby “physical exercise” is a specific, planned and structured form of physical activities [15,16]. Additionally, physical exercise can be divided into acute physical exercise (single bout) and chronic physical exercises (repeated single bouts). Current guidelines recommend a minimum of 150 min moderate-intensity or 75 min vigorous-intensity aerobic activity and strength training per week [17,18]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Müller, P.; Duderstadt, Y.; Lessmann, V.; Müller, N.G. Lactate and BDNF: Key Mediators of Exercise Induced Neuroplasticity? J. Clin. Med. 2020, 9, 1136. https://doi.org/10.3390/jcm9041136
AMA Style
Müller P, Duderstadt Y, Lessmann V, Müller NG. Lactate and BDNF: Key Mediators of Exercise Induced Neuroplasticity? Journal of Clinical Medicine. 2020; 9(4):1136. https://doi.org/10.3390/jcm9041136
Chicago/Turabian StyleMüller, Patrick, Yves Duderstadt, Volkmar Lessmann, and Notger G. Müller. 2020. "Lactate and BDNF: Key Mediators of Exercise Induced Neuroplasticity?" Journal of Clinical Medicine 9, no. 4: 1136. https://doi.org/10.3390/jcm9041136
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.