Maternal High Fat Diet and in-Utero Metformin Exposure Significantly Impact upon the Fetal Renal Proteome of Male Mice

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Sex-PCR
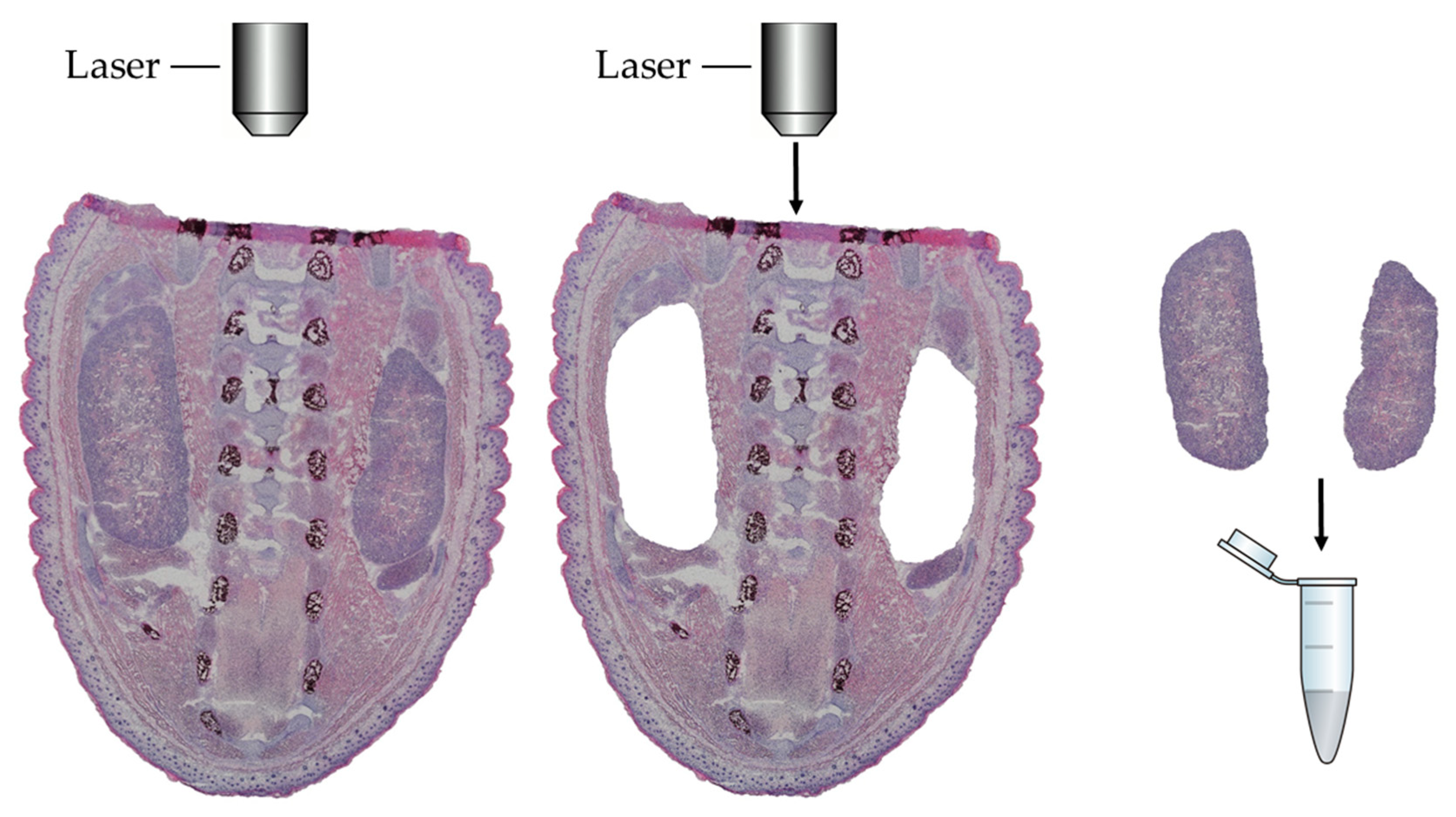
2.3. LCM Procedure and Protein Extraction
2.4. Mass Spectrometry [as Performed by the Proteomics Facility of the Cluster of Excellence—Cellular Stress Responses in Aging-Associated Diseases (CECAD) Cologne]
2.5. Proteomics Data Analysis (as Performed by the Proteomics Facility of the CECAD Cologne)
2.6. Further Data Processing
3. Results
3.1. Animal Phenotype
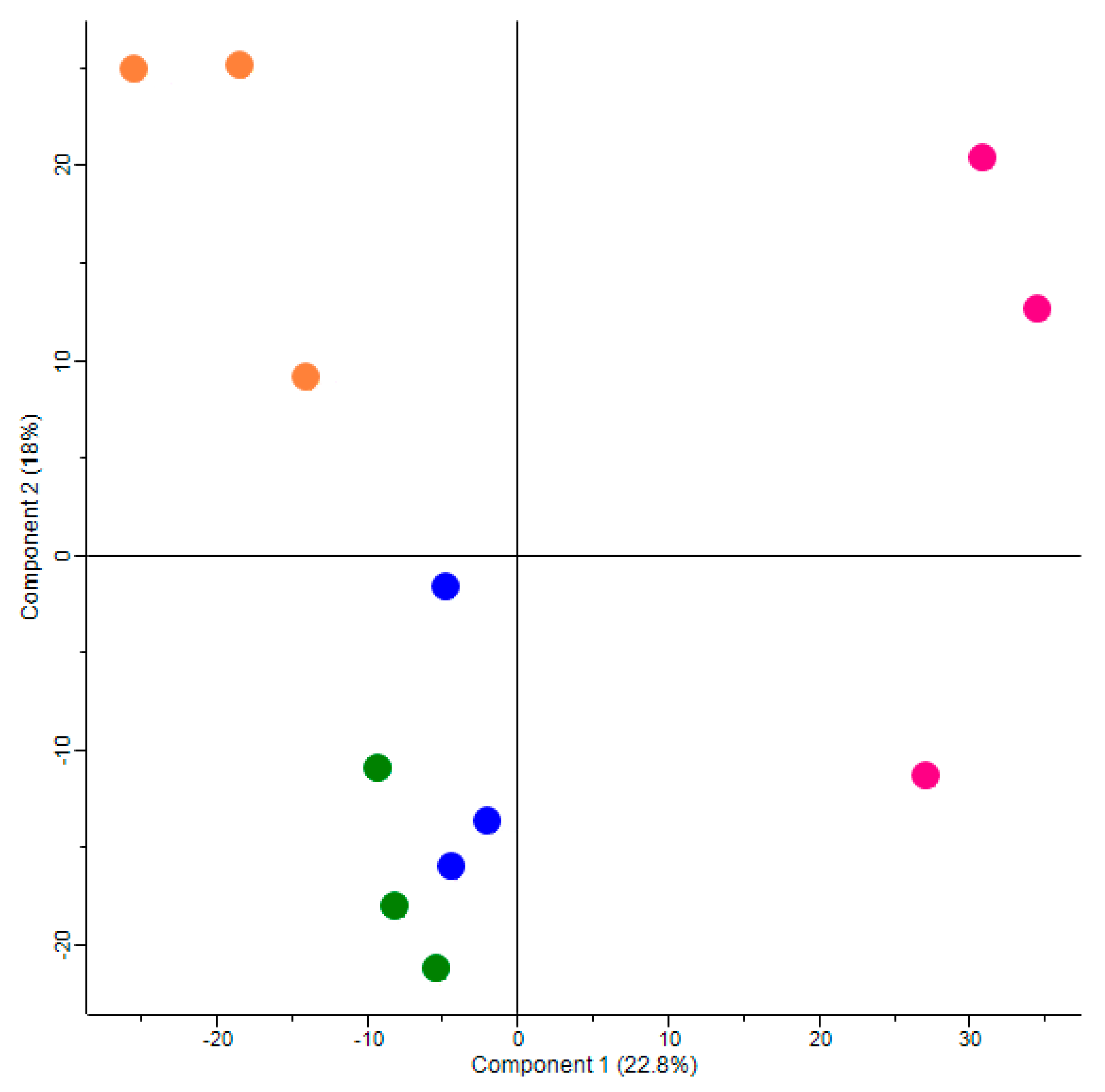
3.2. Principal Component Analysis Iindicates a Strong and Differential Effect of Maternal High Fat Diet and Metformin Treatment on Global Protein Expression in Fetal Kidneys
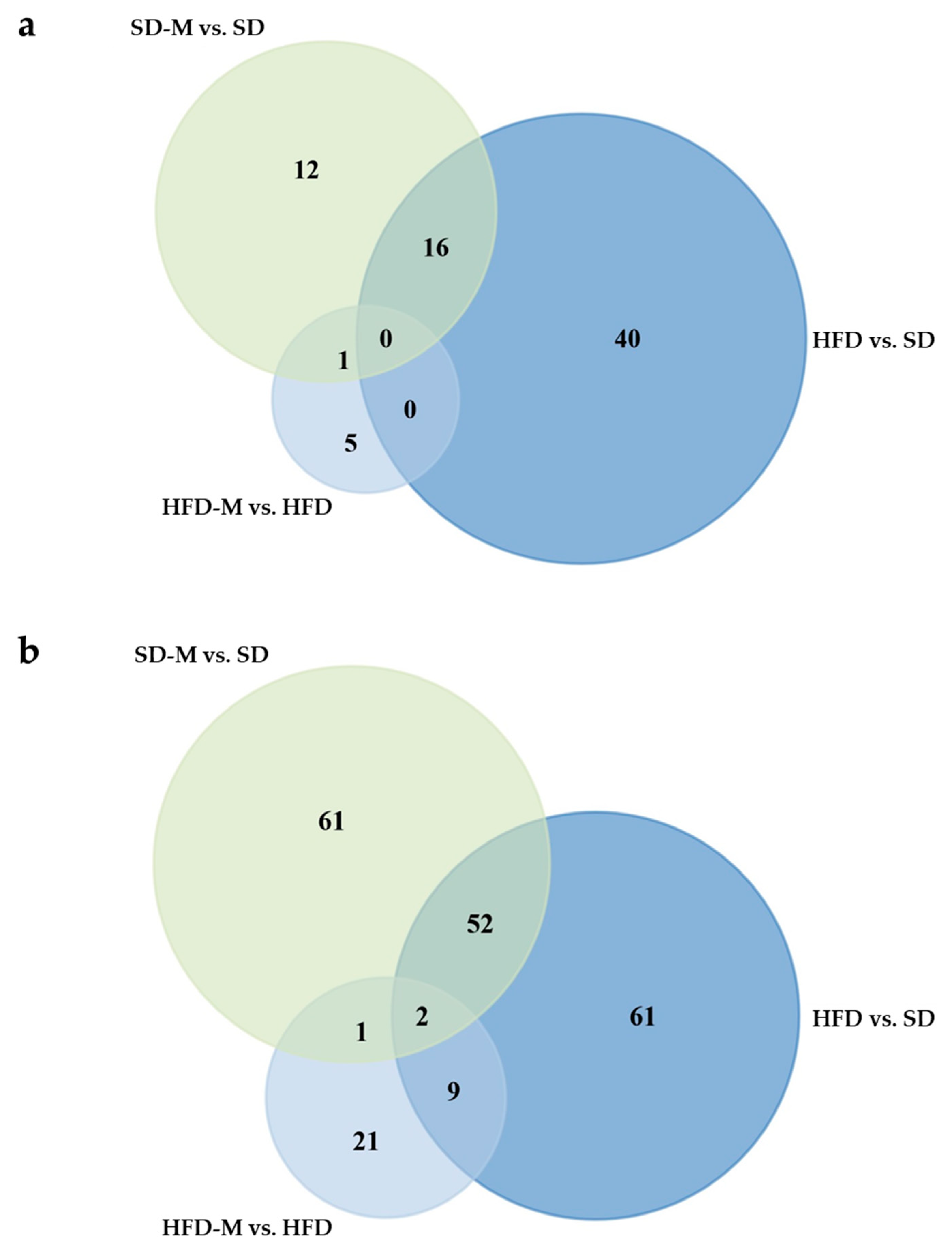
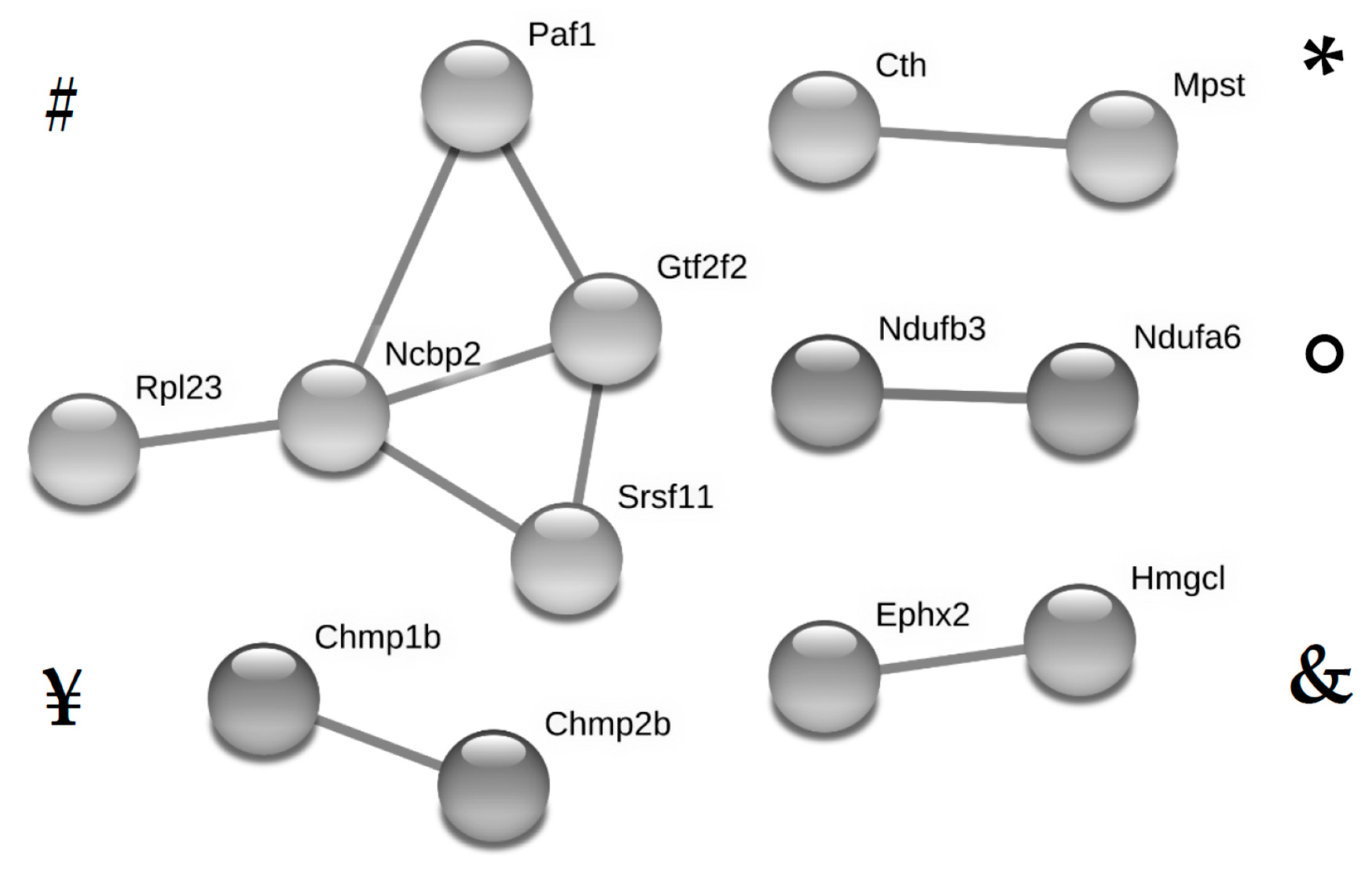
3.3. Overlap Analyses Identify 16 Proteins Which Were Similarly Altered by High Fat Diet and Metformin During Standard Diet, and 11 Proteins Which Were Dysregulated by High Fat diet and Counter-Regulated by Metformin
3.4. High Fat Diet During Mouse Pregnancy Significantly Affects the Levels of 56 Proteins in the Fetal Kidney at Term
3.5. Metformin-Treatment in Healthy Pregnant Mice Significantly Alters the Levels of 29 Proteins in Fetal Kidneys at Term
3.6. Metformin-Treatment in Obese Dams (Group HFD-M) Significantly Alters the Levels of Only Six Proteins in Fetal Kidneys Compared to Group HFD (High Fat Diet Without Metformin)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nüsken, E.; Dötsch, J.; Weber, L.T.; Nüsken, K.D. Developmental Programming of Renal Function and Re-Programming Approaches. Front. Pediat. 2018, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Kanagalingam, M.G.; Forouhi, N.G.; Greer, I.A.; Sattar, N. Changes in booking body mass index over a decade: retrospective analysis from a Glasgow Maternity Hospital. BJOG 2005, 112, 1431–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, L.K.; Prins, J.B.; Chang, A.M.; McIntyre, H.D. The prevalence and impact of overweight and obesity in an Australian obstetric population. Med. J. Aus. 2006, 184, 56–59. [Google Scholar]
- Lisonkova, S.; Muraca, G.M.; Potts, J.; Liauw, J.; Chan, W.S.; Skoll, A.; Lim, K.I. Association Between Prepregnancy Body Mass Index and Severe Maternal Morbidity. JAMA 2017, 318, 1777–1786. [Google Scholar] [CrossRef]
- Kim, S.S.; Zhu, Y.; Grantz, K.L.; Hinkle, S.N.; Chen, Z.; Wallace, M.E.; Smarr, M.M.; Epps, N.M.; Mendola, P. Obstetric and Neonatal Risks Among Obese Women Without Chronic Disease. Obstet. Gynecol. 2016, 128, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Ovesen, P.; Rasmussen, S.; Kesmodel, U. Effect of prepregnancy maternal overweight and obesity on pregnancy outcome. Obstet. Gynecol. 2011, 118, 305–312. [Google Scholar] [CrossRef]
- Zandi-Nejad, K.; Luyckx, V.A.; Brenner, B.M. Adult hypertension and kidney disease: The role of fetal programming. Hypertension 2006, 47, 502–508. [Google Scholar] [CrossRef]
- Charlton, R.A.; Klungsoyr, K.; Neville, A.J.; Jordan, S.; Pierini, A.; de Jong-van den Berg, L.T.; Bos, H.J.; Puccini, A.; Engeland, A.; Gini, R.; et al. Prescribing of Antidiabetic Medicines before, during and after Pregnancy: A Study in Seven European Regions. PLoS ONE 2016, 11, e0155737. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.G.; McGuire, T.M.; Cohen, N.; Little, P.J. The emerging role of metformin in gestational diabetes mellitus. Diabetes Obes. Metab. 2017, 19, 765–772. [Google Scholar] [CrossRef]
- Balani, J.; Hyer, S.; Johnson, A.; Shehata, H. Pregnancy outcomes after metformin treatment for gestational diabetes: a case-control study. Obstet. Med. 2012, 5, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Chiswick, C.; Reynolds, R.M.; Denison, F.; Drake, A.J.; Forbes, S.; Newby, D.E.; Walker, B.R.; Quenby, S.; Wray, S.; Weeks, A.; et al. Effect of metformin on maternal and fetal outcomes in obese pregnant women (EMPOWaR): a randomised, double-blind, placebo-controlled trial. Lancet Diabetes Endocrinol. 2015, 3, 778–786. [Google Scholar] [CrossRef] [Green Version]
- Bolton, S.; Cleary, B.; Walsh, J.; Dempsey, E.; Turner, M.J. Continuation of metformin in the first trimester of women with polycystic ovarian syndrome is not associated with increased perinatal morbidity. Eur. J. Pediatr. 2009, 168, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Charles, B.; Norris, R.; Xiao, X.; Hague, W. Population pharmacokinetics of metformin in late pregnancy. Ther. Drug Monit. 2006, 28, 67–72. [Google Scholar] [CrossRef]
- Vanky, E.; Zahlsen, K.; Spigset, O.; Carlsen, S.M. Placental passage of metformin in women with polycystic ovary syndrome. Fertil Steril. 2005, 83, 1575–1578. [Google Scholar] [CrossRef]
- Ijas, H.; Vaarasmaki, M.; Saarela, T.; Keravuo, R.; Raudaskoski, T. A follow-up of a randomised study of metformin and insulin in gestational diabetes mellitus: growth and development of the children at the age of 18 months. BJOG 2015, 122, 994–1000. [Google Scholar] [CrossRef]
- Battin, M.R.; Obolonkin, V.; Rush, E.; Hague, W.; Coat, S.; Rowan, J. Blood pressure measurement at two years in offspring of women randomized to a trial of metformin for GDM: follow up data from the MiG trial. BMC Pediatr. 2015, 15, 54. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, K.U.; Coresh, J.; Devuyst, O.; Johnson, R.J.; Kottgen, A.; Levey, A.S.; Levin, A. Evolving importance of kidney disease: from subspecialty to global health burden. Lancet 2013, 382, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Kasper, P.; Vohlen, C.; Dinger, K.; Mohr, J.; Hucklenbruch-Rother, E.; Janoschek, R.; Koth, J.; Matthes, J.; Appel, S.; Dötsch, J.; et al. Renal Metabolic Programming Is Linked to the Dynamic Regulation of a Leptin-Klf15 Axis and Akt/AMPKalpha Signaling in Male Offspring of Obese Dams. Endocrinology 2017, 158, 3399–3415. [Google Scholar] [CrossRef] [PubMed]
- Janoschek, R.; Bae-Gartz, I.; Vohlen, C.; Alcazar, M.A.; Dinger, K.; Appel, S.; Dötsch, J.; Hucklenbruch-Rother, E. Dietary intervention in obese dams protects male offspring from WAT induction of TRPV4, adiposity, and hyperinsulinemia. Obesity 2016, 24, 1266–1273. [Google Scholar] [CrossRef]
- Appel, S.; Schulze-Edinghausen, M.; Kretschmer, T.; Storck, S.; Janoschek, R.; Bae-Gartz, I.; Handwerk, M.; Wohlfarth, M.; Nüsken, K.D.; Hucklenbruch-Rother, E.; et al. Maternal obesity attenuates predelivery inflammatory reaction in C57BL/6N mice. J. Reprod. Immunol. 2017, 122, 10–13. [Google Scholar] [CrossRef]
- Appel, S.; Grothe, J.; Storck, S.; Janoschek, R.; Bae-Gartz, I.; Wohlfarth, M.; Handwerk, M.; Hucklenbruch-Rother, E.; Gellhaus, A.; Dötsch, J. A Potential Role for GSK3beta in Glucose-Driven Intrauterine Catch-Up Growth in Maternal Obesity. Endocrinology 2019, 160, 377–386. [Google Scholar] [CrossRef]
- Hurley, J.H.; Hanson, P.I. Membrane budding and scission by the ESCRT machinery: it’s all in the neck. Nat. Rev. Mol. Cell Biol. 2010, 11, 556–566. [Google Scholar] [CrossRef]
- Szeto, H.H.; Liu, S.; Soong, Y.; Alam, N.; Prusky, G.T.; Seshan, S.V. Protection of mitochondria prevents high-fat diet-induced glomerulopathy and proximal tubular injury. Kidney Int. 2016, 90, 997–1011. [Google Scholar] [CrossRef]
- Fan, F.; Roman, R.J. Effect of Cytochrome P450 Metabolites of Arachidonic Acid in Nephrology. J. Am. Soc. Nephrol. 2017, 28, 2845–2855. [Google Scholar] [CrossRef] [PubMed]
- Kopkan, L.; Huskova, Z.; Sporkova, A.; Varcabova, S.; Honetschlagerova, Z.; Hwang, S.H.; Tsai, H.J.; Hammock, B.D.; Imig, J.D.; Kramer, H.J.; et al. Soluble epoxide hydrolase inhibition exhibits antihypertensive actions independently of nitric oxide in mice with renovascular hypertension. Kidney Blood Press. Res. 2012, 35, 595–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuey, M.M.; Billings, F.T.t.; Wei, S.; Milne, G.L.; Nian, H.; Yu, C.; Brown, N.J. Association of gain-of-function EPHX2 polymorphism Lys55Arg with acute kidney injury following cardiac surgery. PLoS ONE 2017, 12, e0175292. [Google Scholar] [CrossRef]
- Chen, G.; Xu, R.; Wang, Y.; Wang, P.; Zhao, G.; Xu, X.; Gruzdev, A.; Zeldin, D.C.; Wang, D.W. Genetic disruption of soluble epoxide hydrolase is protective against streptozotocin-induced diabetic nephropathy. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E563–E575. [Google Scholar] [CrossRef] [Green Version]
- Kasinath, B.S.; Feliers, D.; Lee, H.J. Hydrogen sulfide as a regulatory factor in kidney health and disease. Biochem. Pharmacol. 2018, 149, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Li, H.; Untereiner, A.; Wu, L.; Yang, G.; Austin, R.C.; Dickhout, J.G.; Lhotak, S.; Meng, Q.H.; Wang, R. Decreased endogenous production of hydrogen sulfide accelerates atherosclerosis. Circulation 2013, 127, 2523–2534. [Google Scholar] [CrossRef]
- Rizou, M.; Frangou, E.A.; Marineli, F.; Prakoura, N.; Zoidakis, J.; Gakiopoulou, H.; Liapis, G.; Kavvadas, P.; Chatziantoniou, C.; Makridakis, M.; et al. The family of 14-3-3 proteins and specifically 14-3-3sigma are up-regulated during the development of renal pathologies. J. Cell. Mol. Med. 2018, 22, 4139–4149. [Google Scholar] [CrossRef]
- Evren, S.; Dermen, A.; Lockwood, G.; Fleshner, N.; Sweet, J. mTOR-RAPTOR and 14-3-3sigma immunohistochemical expression in high grade prostatic intraepithelial neoplasia and prostatic adenocarcinomas: a tissue microarray study. J. Clin. Pathol. 2011, 64, 683–688. [Google Scholar] [CrossRef]
- Kim, S.; Wong, P.; Coulombe, P.A. A keratin cytoskeletal protein regulates protein synthesis and epithelial cell growth. Nature 2006, 441, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Emoto, M.; Inaba, M. Fetuin-A: a multifunctional protein. Recent Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 124–146. [Google Scholar] [CrossRef] [PubMed]
- Grewal, J.S.; Tsai, J.Y.; Khan, S.R. Oxalate-inducible AMBP gene and its regulatory mechanism in renal tubular epithelial cells. Biochem. J. 2005, 387, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Alper, S.L. Familial renal tubular acidosis. J. Nephrol. 2010, 23 (Suppl. 16), S57–S76. [Google Scholar] [PubMed]
- Trepiccione, F.; Gerber, S.D.; Grahammer, F.; Lopez-Cayuqueo, K.I.; Baudrie, V.; Paunescu, T.G.; Capen, D.E.; Picard, N.; Alexander, R.T.; Huber, T.B.; et al. Renal Atp6ap2/(Pro)renin Receptor Is Required for Normal Vacuolar H+-ATPase Function but Not for the Renin-Angiotensin System. J. Am. Soc. Nephrol. 2016, 27, 3320–3330. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, F.; Sun, Y.; Lei, L.; Zhou, H.; Lei, T.; Xia, Y.; Verkman, A.S.; Yang, B. Aquaporin-1 retards renal cyst development in polycystic kidney disease by inhibition of Wnt signaling. FASEB J. 2015, 29, 1551–1563. [Google Scholar] [CrossRef] [Green Version]
- Toussaint, J.; Raval, C.B.; Nguyen, T.; Fadaifard, H.; Joshi, S.; Wolberg, G.; Quarfordt, S.; Jan, K.M.; Rumschitzki, D.S. Chronic hypertension increases aortic endothelial hydraulic conductivity by upregulating endothelial aquaporin-1 expression. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H1063–H1073. [Google Scholar] [CrossRef]
- Woeller, C.F.; O’Loughlin, C.W.; Pollock, S.J.; Thatcher, T.H.; Feldon, S.E.; Phipps, R.P. Thy1 (CD90) controls adipogenesis by regulating activity of the Src family kinase, Fyn. FASEB J. 2015, 29, 920–931. [Google Scholar] [CrossRef]
- Zhang, Q.; Luan, H.; Wang, L.; He, F.; Zhou, H.; Xu, X.; Li, X.; Xu, Q.; Niki, T.; Hirashima, M.; et al. Galectin-9 ameliorates anti-GBM glomerulonephritis by inhibiting Th1 and Th17 immune responses in mice. Am. J. Physiol. Renal Physiol. 2014, 306, F822–F832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurose, Y.; Wada, J.; Kanzaki, M.; Teshigawara, S.; Nakatsuka, A.; Murakami, K.; Inoue, K.; Terami, T.; Katayama, A.; Watanabe, M.; et al. Serum galectin-9 levels are elevated in the patients with type 2 diabetes and chronic kidney disease. BMC Nephrol. 2013, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.; Wada, J.; Eguchi, J.; Hashimoto, I.; Okada, T.; Yasuhara, A.; Shikata, K.; Kanwar, Y.S.; Makino, H. Galectin-9 inhibits glomerular hypertrophy in db/db diabetic mice via cell-cycle-dependent mechanisms. J. Am. Soc. Nephrol. 2005, 16, 3222–3234. [Google Scholar] [CrossRef] [PubMed]
- Kvirkvelia, N.; McMenamin, M.; Warren, M.; Jadeja, R.N.; Kodeboyina, S.K.; Sharma, A.; Zhi, W.; O’Connor, P.M.; Raju, R.; Lucas, R.; et al. Kidney-targeted inhibition of protein kinase C-alpha ameliorates nephrotoxic nephritis with restoration of mitochondrial dysfunction. Kidney Int. 2018, 94, 280–291. [Google Scholar] [CrossRef]
- Koya, D. Dual protein kinase C alpha and beta inhibitors and diabetic kidney disease: a revisited therapeutic target for future clinical trials. J. Diabetes Investig. 2014, 5, 147–148. [Google Scholar] [CrossRef]
- Serlachius, E.; Svennilson, J.; Schalling, M.; Aperia, A. Protein kinase C in the developing kidney: isoform expression and effects of ceramide and PKC inhibitors. Kidney Int. 1997, 52, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Kishino, M.; Yukawa, K.; Hoshino, K.; Kimura, A.; Shirasawa, N.; Otani, H.; Tanaka, T.; Owada-Makabe, K.; Tsubota, Y.; Maeda, M.; et al. Deletion of the kinase domain in death-associated protein kinase attenuates tubular cell apoptosis in renal ischemia-reperfusion injury. J. Am. Soc. Nephrol. 2004, 15, 1826–1834. [Google Scholar] [CrossRef] [PubMed]
- Loayza-Puch, F.; Rooijers, K.; Buil, L.C.; Zijlstra, J.; Oude Vrielink, J.F.; Lopes, R.; Ugalde, A.P.; van Breugel, P.; Hofland, I.; Wesseling, J.; et al. Tumour-specific proline vulnerability uncovered by differential ribosome codon reading. Nature 2016, 530, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Arystarkhova, E.; Ralph, D.L.; Liu, Y.B.; Bouley, R.; McDonough, A.A.; Sweadner, K.J. Paradoxical activation of the sodium chloride cotransporter (NCC) without hypertension in kidney deficient in a regulatory subunit of Na,K-ATPase, FXYD2. Physiol. Rep. 2014, 2. [Google Scholar] [CrossRef]
- Arystarkhova, E. Beneficial Renal and Pancreatic Phenotypes in a Mouse Deficient in FXYD2 Regulatory Subunit of Na,K-ATPase. Front. Physiol. 2016, 7, 88. [Google Scholar] [CrossRef]
- de Baaij, J.H.; Dorresteijn, E.M.; Hennekam, E.A.; Kamsteeg, E.J.; Meijer, R.; Dahan, K.; Muller, M.; van den Dorpel, M.A.; Bindels, R.J.; Hoenderop, J.G.; et al. Recurrent FXYD2 p.Gly41Arg mutation in patients with isolated dominant hypomagnesaemia. Nephrol. Dial. Transplant. 2015, 30, 952–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairo, E.R.; Swarts, H.G.; Wilmer, M.J.; Willems, P.H.; Levtchenko, E.N.; De Pont, J.J.; Koenderink, J.B. FXYD2 and Na,K-ATPase expression in isolated human proximal tubular cells: disturbed upregulation on renal hypomagnesemia? J. Membr. Biol. 2009, 231, 117–124. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
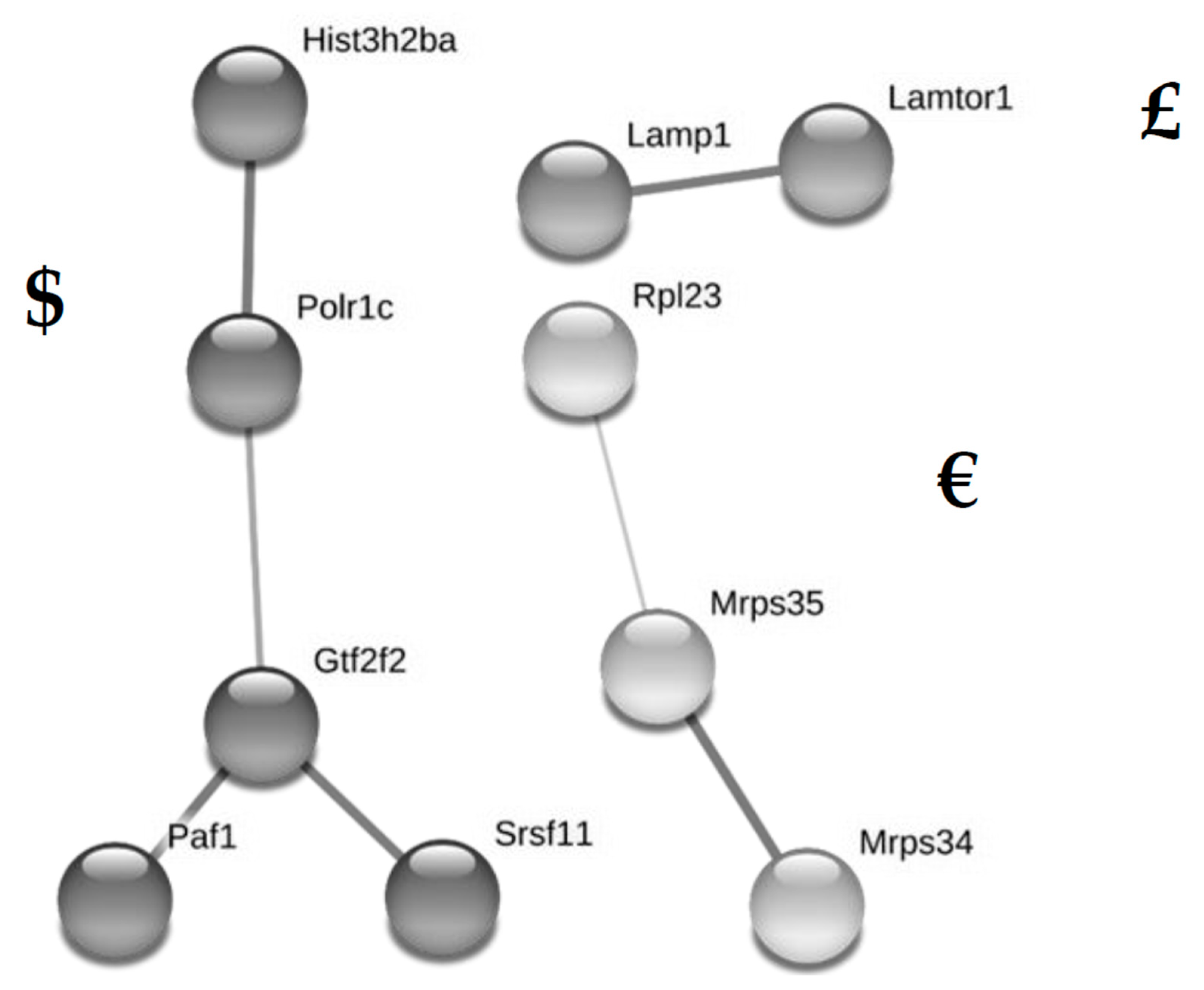{kind=link}
| Protein ID | Protein Name | q-value | Fold Change | Valid Values | Gene Name | Cluster | Functional Enrichments | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SD | HFD | ||||||||||
| O70456 | 14-3-3 protein sigma | <0.05 | 104.8 | 0 | 3 | Sfn | |||||
| P62830 | 60S ribosomal protein L23 | <0.001 | 65.0 | 0 | 3 | Rpl23 | # | ||||
| P43274 | Histone H1.4 | <0.05 | 49.7 | 0 | 3 | Hist1h1e | |||||
| P29699 | Alpha-2-HS-glycoprotein | <0.05 | 35.4 | 0 | 3 | Ahsg | |||||
| Q07456 | Protein AMBP | <0.05 | 32.1 | 0 | 3 | Ambp | |||||
| E9Q8N1 | Titin | <0.05 | 22.8 | 0 | 3 | Ttn | |||||
| O55125 | Protein NipSnap homolog 1 | <0.05 | 16.5 | 0 | 3 | Nipsnap1 | |||||
| Q9CR51 | V-type proton ATPase subunit G 1 | <0.05 | 14.3 | 0 | 3 | Atp6v1g1 | |||||
| Q9D2U9 | Histone H2B type 3-A | <0.001 | 12.0 | 0 | 3 | Hist3h2ba | |||||
| Q02013 | Aquaporin-1 | <0.05 | 10.1 | 0 | 3 | Aqp1 | |||||
| A0A1L1SUX8 | Thy-1 membrane glycoprotein | <0.05 | 10.0 | 0 | 3 | Thy1 | |||||
| B1AQR8 | Galectin | <0.05 | 9.2 | 0 | 3 | Lgals9 | |||||
| Q9EP89 | Serine beta-lactamase-like protein LACTB, mitochondrial | <0.05 | 9.1 | 0 | 3 | Lactb | |||||
| P61028 | Ras-related protein Rab-8B | <0.05 | 9.0 | 0 | 3 | Rab8b | |||||
| Q8BJ64 | Choline dehydrogenase, mitochondrial | <0.05 | 8.4 | 0 | 3 | Chdh | |||||
| Q6P069 | Sorcin | <0.05 | 8.0 | 0 | 3 | Sri | |||||
| Q9D706 | RNA polymerase II-associated protein 3 | <0.05 | 7.2 | 0 | 3 | Rpap3 | |||||
| Q3UFY7 | 7-methylguanosine phosphate-specific 5-nucleotidase | <0.05 | 6.8 | 0 | 3 | Nt5c3b | |||||
| P34914 | Bifunctional epoxide hydrolase 2 | <0.05 | 6.2 | 0 | 3 | Ephx2 | & | ||||
| Q3UW66 | Sulfurtransferase | <0.05 | 6.1 | 0 | 3 | Mpst | * | ||||
| Q3ULB1 | Testin | <0.05 | 6.0 | 0 | 3 | Tes | |||||
| Q8BJF9 | Charged multivesicular body protein 2b | <0.05 | 5.8 | 0 | 3 | Chmp2b | ¥ | ||||
| J3QJX3 | Protein sel-1 homolog 1 | <0.05 | 5.6 | 0 | 3 | Sel1l | |||||
| Q3TE40 | Replication protein A 32 kDa subunit | <0.001 | 5.4 | 0 | 3 | Rpa2 | |||||
| Q8K2T8 | RNA polymerase II-associated factor 1 homolog | <0.05 | 5.0 | 0 | 3 | Paf1 | # | ||||
| D3Z3D2 | Nuclear cap-binding protein subunit 2 | <0.05 | 4.9 | 0 | 3 | Ncbp2 | # | ||||
| Q99LU0 | Charged multivesicular body protein 1b-1 | <0.05 | 4.6 | 0 | 3 | Chmp1b1 | ¥ | ||||
| Q9CZR8 | Elongation factor Ts, mitochondrial | <0.05 | 4.6 | 0 | 3 | Tsfm | |||||
| Q8R409 | Protein HEXIM1 | <0.05 | 4.4 | 0 | 3 | Hexim1 | |||||
| Q8BGY7 | Protein FAM210A | <0.05 | 4.3 | 0 | 3 | Fam210a | |||||
| O88665 | Bromodomain-containing protein 7 | <0.001 | 4.3 | 0 | 3 | Brd7 | |||||
| Q3UIX4 | N/A | <0.05 | 4.1 | 0 | 3 | Srsf11 | # | ||||
| E9QMC1 | Cingulin | <0.05 | 4.0 | 0 | 3 | Cgn | |||||
| A0A0R4J1N9 | Transcription factor A, mitochondrial | <0.05 | 4.0 | 0 | 3 | Tfam | |||||
| Q9JIK9 | 28S ribosomal protein S34, mitochondrial | <0.05 | 3.9 | 0 | 3 | Mrps34 | |||||
| A0A1L1SSA8 | Transmembrane protein 205 | <0.05 | 3.9 | 0 | 3 | Tmem205 | |||||
| Q99LS3 | Phosphoserine phosphatase | <0.01 | 3.9 | 0 | 3 | Psph | |||||
| Q8BHS6 | Armadillo repeat-containing X-linked protein 3 | <0.05 | 3.8 | 0 | 3 | Armcx3 | |||||
| Q8R0A0 | General transcription factor IIF subunit 2 | <0.05 | 3.6 | 0 | 3 | Gtf2f2 | # | ||||
| O89051 | Integral membrane protein 2B | <0.05 | 3.4 | 0 | 3 | Itm2b | |||||
| Q8VCN5 | Cystathionine gamma-lyase | <0.05 | 3.3 | 0 | 3 | Cth | * | ||||
| O35682 | Myeloid-associated differentiation marker | <0.05 | 3.3 | 0 | 3 | Myadm | |||||
| Q3UI43 | BRISC and BRCA1-A complex member 1 | <0.05 | 3.2 | 0 | 3 | Babam1 | |||||
| P38060 | Hydroxymethylglutaryl-CoA lyase, mitochondrial | <0.001 | 3.1 | 0 | 2 | Hmgcl | & | ||||
| O55106 | Striatin | <0.05 | 3.0 | 0 | 3 | Strn | |||||
| A0A140LIQ1 | Transforming acidic coiled-coil-containing protein 2 | <0.05 | 2.9 | 0 | 3 | Tacc2 | |||||
| Q3UE37 | Ubiquitin-conjugating enzyme E2 Z | <0.05 | 2.4 | 0 | 3 | Ube2z | |||||
| Q9WVL0 | Maleylacetoacetate isomerase | <0.05 | 1.8 | 0 | 3 | Gstz1 | |||||
| Q9ESU7 | Amino acid transporter | <0.05 | −1.5 | 3 | 3 | Slc1a5 | |||||
| E9PZY8 | Protein virilizer homolog | <0.05 | −2.1 | 3 | 0 | Virma | |||||
| Q4VA93 | Protein kinase C | <0.05 | −3.0 | 3 | 0 | Prkca | |||||
| Q9CQZ6 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 3 | <0.001 | −9.6 | 3 | 0 | Ndufb3 | ° | ||||
| Q9CQZ5 | NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 6 | <0.05 | −10.5 | 3 | 0 | Ndufa6 | ° | ||||
| D3YZN4 | Paraplegin | <0.001 | −12.3 | 3 | 0 | Spg7 | |||||
| E9QAE4 | Histone-lysine N-methyltransferase | <0.05 | −17.8 | 3 | 0 | Nsd1 | |||||
| Q80YE7 | Death-associated protein kinase 1 | <0.001 | −48.6 | 3 | 0 | Dapk1 | |||||
| Protein ID | Protein Name | q-value | Fold Change | Valid Values | Gene Name | Cluster | |
|---|---|---|---|---|---|---|---|
| SD | SD-M | ||||||
| P62830 | 60S ribosomal protein L23 | <0.05 | 106.8 | 0 | 3 | Rpl23 | € |
| Q07456 | Protein AMBP | <0.05 | 31.4 | 0 | 3 | Ambp | |
| O55125 | Protein NipSnap homolog 1 | <0.001 | 16.6 | 0 | 3 | Nipsnap1 | |
| P11438 | Lysosome-associated membrane glycoprotein 1 | <0.05 | 15.5 | 0 | 3 | Lamp1 | £ |
| Q9D2U9 | Histone H2B type 3-A | <0.05 | 13.6 | 0 | 3 | Hist3h2ba | $ |
| G3UXX5 | DNA-directed RNA polymerases I and III subunit RPAC1 | <0.05 | 11.3 | 0 | 3 | Polr1c | $ |
| Q9EP89 | Serine beta-lactamase-like protein LACTB, mitochondrial | <0.05 | 9.0 | 0 | 3 | Lactb | |
| Q9D706 | RNA polymerase II-associated protein 3 | <0.05 | 8.2 | 0 | 3 | Rpap3 | |
| Q8K2T8 | RNA polymerase II-associated factor 1 homolog | <0.05 | 7.9 | 0 | 3 | Paf1 | $ |
| Q3ULB1 | Testin | <0.05 | 6.1 | 0 | 3 | Tes | |
| O88665 | Bromodomain-containing protein 7 | <0.05 | 5.3 | 0 | 3 | Brd7 | |
| Q9JIK9 | 28S ribosomal protein S34, mitochondrial | <0.05 | 5.1 | 0 | 3 | Mrps34 | € |
| P11157 | Ribonucleoside-diphosphate reductase subunit M2 | <0.05 | 5.0 | 0 | 3 | Rrm2 | |
| Q3UIX4 | Serine/arginine-rich-splicing factor 11 | <0.05 | 5.0 | 0 | 3 | Srsf11 | $ |
| A0A0R4J0L6 | 28S ribosomal protein S35, mitochondrial | <0.05 | 5.0 | 0 | 3 | Mrps35 | € |
| Q9DBX2 | Phosducin-like protein | <0.01 | 4.8 | 0 | 3 | Pdcl | |
| H7BX44 | Non-specific serine/threonine protein kinase | <0.05 | 4.7 | 0 | 3 | Cdc42bpa | |
| Q8R0A0 | General transcription factor IIF subunit 2 | <0.05 | 4.2 | 0 | 3 | Gtf2f2 | $ |
| F8WIK0 | Anamorsin | <0.05 | 4.2 | 0 | 3 | Ciapin1 | |
| E9QMC1 | Cingulin | <0.05 | 3.7 | 0 | 3 | Cgn | |
| O55106 | Striatin | <0.01 | 3.5 | 0 | 3 | Strn | |
| O89051 | Integral membrane protein 2B | <0.001 | 3.4 | 0 | 3 | Itm2b | |
| Q8N7N5 | DDB1- and CUL4-associated factor 8 | <0.05 | 2.8 | 0 | 3 | Dcaf8 | |
| Q3UH93 | Plexin-D1 | <0.05 | 2.7 | 0 | 3 | Plxnd1 | |
| A0A0R4J0G4 | Ran-binding protein 10 | <0.05 | 2.6 | 0 | 3 | Ranbp10 | |
| P52483 | Ubiquitin-conjugating enzyme E2 E3 | <0.05 | 2.3 | 0 | 3 | Ube2e3 | |
| G3UZ63 | Kelch domain-containing protein 4 | <0.05 | 2.3 | 0 | 3 | Klhdc4 | |
| A0A0A6YX02 | Ragulator complex protein LAMTOR1 | <0.05 | −10.3 | 3 | 0 | Lamtor1 | £ |
| Q80YE7 | Death-associated protein kinase 1 | <0.05 | −48.7 | 3 | 0 | Dapk1 | |
| Protein ID | Protein Name | q-value | Fold Change | Valid Values | Gene Name | |
|---|---|---|---|---|---|---|
| HFD | HFD-M | |||||
| A0A0R4J0L6 | 28S ribosomal protein S35, mitochondrial | <0.05 | 8.6 | 0 | 3 | Mrps35 |
| Q8K2C9 | Very-long-chain (3R)-3-hydroxyacyl-CoA dehydratase 3 | <0.05 | 7.4 | 0 | 3 | Hacd3 |
| E9Q784 | Zinc finger CCCH domain-containing protein 13 | <0.001 | 5.7 | 0 | 3 | Zc3h13 |
| Q9Z1J3 | Cysteine desulfurase, mitochondrial | <0.05 | −3.0 | 3 | 0 | Nfs1 |
| Q922W5 | Pyrroline-5-carboxylate reductase 1, mitochondrial | <0.05 | −10.1 | 3 | 0 | Pycr1 |
| Q04646 | Sodium/potassium-transporting ATPase subunit gamma | <0.05 | −19.9 | 3 | 0 | Fxyd2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nüsken, E.; Turnwald, E.-M.; Fink, G.; Voggel, J.; Yosy, C.; Kretschmer, T.; Handwerk, M.; Wohlfarth, M.; Weber, L.T.; Hucklenbruch-Rother, E.; et al. Maternal High Fat Diet and in-Utero Metformin Exposure Significantly Impact upon the Fetal Renal Proteome of Male Mice. J. Clin. Med. 2019, 8, 663. https://doi.org/10.3390/jcm8050663
Nüsken E, Turnwald E-M, Fink G, Voggel J, Yosy C, Kretschmer T, Handwerk M, Wohlfarth M, Weber LT, Hucklenbruch-Rother E, et al. Maternal High Fat Diet and in-Utero Metformin Exposure Significantly Impact upon the Fetal Renal Proteome of Male Mice. Journal of Clinical Medicine. 2019; 8(5):663. https://doi.org/10.3390/jcm8050663
Chicago/Turabian StyleNüsken, Eva, Eva-Maria Turnwald, Gregor Fink, Jenny Voggel, Christopher Yosy, Tobias Kretschmer, Marion Handwerk, Maria Wohlfarth, Lutz T. Weber, Eva Hucklenbruch-Rother, and et al. 2019. "Maternal High Fat Diet and in-Utero Metformin Exposure Significantly Impact upon the Fetal Renal Proteome of Male Mice" Journal of Clinical Medicine 8, no. 5: 663. https://doi.org/10.3390/jcm8050663