
The Role of Selected Serpins in Gastrointestinal (GI) Malignancies


1
Department of Biochemical Diagnostics, University Hospital in Bialystok, Waszyngtona 15A St., 15-269 Bialystok, Poland
2
Department of Biochemical Diagnostics, Medical University in Bialystok, Waszyngtona 15A St., 15-269 Bialystok, Poland
3
Department of Neurodegeneration Diagnostics, Medical University in Bialystok, Waszyngtona 15A St., 15-269 Bialystok, Poland
*
Author to whom correspondence should be addressed.
J. Clin. Med. 2022, 11(20), 6225; https://doi.org/10.3390/jcm11206225
Submission received: 6 September 2022
/
Revised: 18 October 2022
/
Accepted: 19 October 2022
/
Published: 21 October 2022
(This article belongs to the Special Issue Colorectal Disorders: Diagnosis and Treatments)
Abstract
:Gastrointestinal (GI) cancers, which are a diverse group of malignant diseases, represent a major healthcare problem around the world. Due to the lack of specific symptoms in the early stages as well as insufficient diagnostic possibilities, these malignancies occupy the leading position in the causes of death worldwide. The currently available tests have too many limitations to be part of routine diagnostics. Therefore, new potential biomarkers that could be used as diagnostic and prognostic factors for these cancers are still being sought. Among the proteins that might fit this role are serpins, which are serine protease inhibitors. Although the serpins themselves have been known for many years, they have recently become the centre of attention for many authors, especially due to the fact that a number of proteins in this family are involved in many stages of neoplasia formation, from angiogenesis through tumour growth to progression. Therefore, the aim of this review is to present the current knowledge about the significance of serpins in GI malignancies, especially their involvement in the development and progression of oesophageal, gastric, pancreatic and colorectal cancers. This review summarises and confirms the important roles of selected serpins in the pathogenesis of various GI cancers and also points to their promising roles as therapeutic targets. However, due to the relatively nonspecific nature of serpins, future research should be carried out to elucidate the mechanisms involved in tumour pathogenesis in more detail.
1. Introduction
1.1. Gastrointestinal (GI) Cancers
The digestive tract consists of many interconnected sections responsible for maintaining the human body in full health [1]. Gastrointestinal (GI) cancers are a diverse group of diseases that include oesophageal cancer (EC), gastric cancer (GC), pancreatic cancer (PC), liver cancer (LC), gallbladder cancer (GBC) and colorectal cancer (CRC) [2]. According to the data presented by the American Cancer Society, in 2018, there were almost 2 million new cases of GI cancers, accounting for over 30% of total cancer deaths, according to estimates [3]. These tumours have been observed to occur in both sexes and represent the most common types of cancer among men and the second most common in women. In most cases, tumours of the GI tract are neoplasms with relatively poor prognoses, which is reflected in the small differences between the numbers of cases and deaths in particular years of follow-up [4]. Although the incidence of some types of GI cancer has decreased, this group of malignancies remains a major public health challenge [5]. The incidence of GI cancers also presents geographical variation. The CRC incidence is higher in Western Europe and North America, while the LC and GC incidences are higher in Asia and Africa. However, PC is more common in the populations of industrialised countries. Thus, in Europe, it is much more frequent in the Nordic countries than in the Mediterranean ones [6,7,8]. The evaluation of the risk factors for GI cancers has shown that more than half of all these cancers are caused by modifiable risk factors, such as high-percentage alcohol consumption and tobacco smoking, whose roles in carcinogenesis have been confirmed for many types of histological malignancies [9,10,11]. Many studies have shown that prevention in the form of regular physical activity, an appropriate diet rich in fibre and protection against viral infections, as well as drug elimination, reduce the risk of GI cancers [11,12].
The clinical symptoms of GI malignancies vary depending on the location as well as the tumour stage and mostly affect fatigue, which is followed by malaise, pain, sleep disturbances, a lack of appetite, dyspnoea and nausea [13,14,15,16]. However, the later a diagnosis is made, the lower the chance of a cure.
The diagnostic process for patients with GI cancers is based on endoscopic evaluation. However, the auxiliary examinations cover the scope of imaging diagnostics as an important tool in both diagnosis and staging. In the diagnostic process for patients with GI neoplasms, apart from imaging methods, it is also recommended to perform laboratory tests. In addition to visualizing the patient’s general condition, they allow the assessment of the classical tumour markers of GI neoplasms [17,18,19]. These include such markers as carcinoembryonic antigen (CEA), tumour antigen 19-9 (CA 19.9), tumour antigen 72.4 (CA 72.4) and tumour antigen 50 (CA 50). Unfortunately, these markers are not useful in the early detection of these tumours due to their poor diagnostic criteria such as low diagnostic sensitivity and specificity [20,21,22,23].
The treatment options for GI cancers primarily depend on the type and stage of the cancer and the patient’s age, general health and preferences. The most common treatments for GI today include chemotherapy with targeted therapy, surgery and radiation therapy. A relatively new concept is neoadjuvant therapy, which can consist of chemotherapy, radiation therapy or hormone therapy [24,25,26,27]. One example of the use of neoadjuvant therapy techniques is rectal cancer. In this case of locally advanced neoplasm, the CXRT (chemo-radiation therapy) is used, which allows usually for the reduction of local recurrence, but usually does not affect distant metastases. This treatment is followed by surgery for 8–12 weeks [28].
The prognosis of patients with GI neoplasms remains unchanged. Although current treatment strategies have developed, patients’ quality of life has only slightly improved. The limited efficacy of therapy in patients with advanced disease reflects an incomplete understanding of the molecular basis of GI carcinogenesis. Currently, the most important prognostic factors for patient survival are the histological degree of the tumour and the tumour stage at diagnosis, including the depth of tumour infiltration and the involvement of regional lymph nodes. In addition to these clinico-pathological parameters, GI cancer biomarkers are being intensively researched and validated. However, the low sensitivity and specificity of the available biomarkers limit their usefulness, especially in early stage screening or in distinguishing between aggressive and benign tumours. For this reason, research is ongoing to find new biomarkers that would allow for early cancer detection and assessment of response to therapy [29,30,31].
1.2. The Structure and Functions of Serpins
The activity of proteases appears to play a key role in the survival of multicellular organisms. These enzymes are responsible for breaking peptide bonds, which in turn leads to irreversible post-translational modifications of proteins [32]. An example of such proteolytic cascades is inflammation and coagulation, which, if inappropriately activated, can lead to clotting disorders and inflammation in the host. Thus, it is not surprising that protease activity is regulated by various inhibitors, including the serine protease inhibitors [33]. Serpins (SERine Proteinase INhibitors) are the largest superfamily of protease inhibitors, representing about 2–10% of circulating plasma proteins [34]. They generally consist of approximately 350–400 amino acid residues, with molecular masses of 40–100 kDa. The core of a serpin’s structure is described as highly conserved, which has been found to be important for its functions [35]. Native serpins present two common features: five-stranded beta-sheets A are located in the middle of the molecule, and a flexible RCL (reactive centre loop) is located on the top of the molecule. The currently used nomenclature for serpins consists of two parts: alphabetic and numerical designations. However, this nomenclature does not refer to any of their evolutionary relations, but rather to the order of their discovery [36]. The comprehensive phylogenetic analysis of the eukaryotic serpin family divided them into 16 clades—from A to P. However, many serpins bear alternative names that were given before the current classification was proposed [37]. Serpins are mainly found as extracellular molecules, although some of them, such as clade B serpins, may act inside cells due to their absence of N-terminal secretory signals [38]. Among the newly characterised functions of these proteins, the regulation of cell proliferation appears to play a major role [39]. Accordingly, several serpins have been found to be overexpressed in tumour cells [40]. Most serpins are responsible for the inhibition of serine proteases, but some of them have additional functions that inhibit some representatives of cysteine proteins, such as caspases as well as cathepsins [41]. Serpins have been also called the “workhorses” of the human body because they play extremely important roles in the regulation of diverse biological activities. Moreover, these proteins are involved in several physiological processes such as blood coagulation, inflammation and immunity [42]. Therefore, serpins have been also classified according to their functions. The inhibitory and non-inhibitory functions of serpins are presented in Figure 1 [43]. Apart from their physiological implications, serpins have also been found to be involved in the pathogenesis of many diseases. Diseases caused by both mutations and dysfunction of the serpins themselves have been termed serpinopathies [44]. Most serpin-related diseases are the result of serpin polymerisation and aggregation, but there are several other types of disease that are caused by their mutations. The most common inherited disease is alpha-1-antitrypsin deficiency [45]. Recent scientific work has defined two potential pathways by which serpins are responsible for tissue damage. The first one concerns the accumulation of polymers, which leads to endoplasmic reticulum stress and inflammation (e.g., liver cirrhosis) [45]. The second mechanism involves the loss of serpin function, which results in uncontrolled activity of proteases. An example is the hyperactivity of elastase resulting from alpha-1-antitrypsin (A1AT) deficiency, leading to emphysema [46].
2. Materials and Methods
A comprehensive literature search was performed by using the MEDLINE/PubMed database covering the period up to March 2022. The authors used following search strategy: studies were limited to those in the English language, and duplicates and all non-significant manuscripts (i.e., papers that did not concern the selected serpins/cancers and biomarkers) were excluded. Initially, the authors used the keyword “serpins” (68,332 studies). Then, the following keywords were employed: “serpins AND cancer” (8657 studies) and “serpins AND gastrointestinal cancer or GI cancer” (744 studies). Due to the small number of records, the review focused on selected cancers, which most often appeared in the results, taking into account their abbreviations: “serpins AND oesophageal cancer or EC or ESCC” (15,664 studies), “serpins AND gastric cancer or GC” (291 studies), “serpins AND pancreatic cancer or PC” (4530 studies) and “serpins AND colorectal cancer or CRC” (390 studies). The analysis of the obtained results identified the five most common serpins, which are described in this review. Bearing in mind the different names of the serpins, the aliases as well as abbreviations were also included in the key words, e.g., “serpinA1 or A1AT or alpha-1-antitripsin AND CRC or colorectal cancer”. A literature search on the remaining serpins (vaspin, maspin, PAI-1 and PEDF) and tumours (GC, PC and CRC) was conducted following the same pattern.
3. Results
3.1. Serpins in Cancer
Oncogenesis is the process by which normal cells become cancerous. It is characterised by both genetic and cellular changes, as well as abnormal cell division [47]. Under normal conditions, the balance between cells’ proliferation and their apoptosis preserves the integrity of both tissues and organs [48].
Serpins can bind to and interact with numerous molecules, and their function has therefore evolved from simple serine protease inhibitors to biological molecules with complex properties. These proteins are associated with the progression or remission of various cancers, making them valuable in diagnostic and therapeutic applications [49]. However, the role of serpins in cancer, as well as their mechanisms, is subject to much controversy [50]. In the present review, the involvement of these molecules in tumours will be described based on two examples: plasminogen activator inhibitor-1 (PAI-1) and pigment epithelium-derived factor (PEDF).
PAI-1 appears to be the most studied serpin in cancer biology. It has been suggested that it plays a key role in cancer, mainly through its association with urokinase-type plasminogen activator (uPA) and the extracellular matrix (ECM) protein vitronectin [51]. However, the involvement of PAI-1 in human cancer is multifaceted and, through a number of different mechanisms, may affect the maintenance of proliferative signals, angiogenesis, invasion and the formation of distant metastases. A relatively recent discovery is the participation of PAI-1 in cancer-related inflammation [52]. Experimental studies conducted in both animal and human models have shown that inflammatory cytokines such as interleukin-6 (IL-6) or tumour necrosis factor alpha (TNF-alpha) increase the expression of PAI-1 [53]. This suggests that the increase in PAI-1 is a consequence of an inflammatory state. The latest research supports this theory, leading to strong conclusions that this serpin is both a consequence and a cause of inflammation, especially by increasing interleukin-8 (IL-8) production [54]. However, many reports have shown that elevated levels of PAI-1 are observed in inflammatory conditions not associated with cancer, such as sepsis or arthritis [55]. The role of PAI-1 in cancer is presented in Figure 2.
PEDF is a member of the serpin superfamily, which can act directly on tumours by inducing differentiation to a less malignant phenotype as well as promoting the apoptotic death of tumour cells [57]. Moreover, this protein can also inhibit the proliferation of neoplastic cells, and has antimetastatic effects by inhibiting the invasion and migration of neoplastic cells. In this regard, it is worth bearing in mind that many reports show that reduced PEDF levels are associated with tumour progression, and the exogenous administration of PEDF in animal models results in tumour growth inhibition and prolonged survival [58]. The mechanisms underlying the regulation of the behaviour of cancer cells by PEDF are mainly based on the interaction of this serpin with various receptors on the cell surface, which triggers a number of signalling pathways [59].
3.2. The Role of Serpins in GI Cancers
Much evidence proves that several serpins play roles in various cancers [40,60]. Among all the serpins, A1AT, waspin, maspin, PAI-1 and PEDF have been shown to make the greatest contributions in cancer [60,61,62,63]. Therefore, they are described more fully in the present manuscript. The involvement of selected serpins in GI cancers is presented in Table 1.
3.2.1. SERPIN A1 (Serpin Family A Member 1)—A1AT (Alpha-1-Antitrypsin)
A1AT is a plasma glycoprotein in the alpha 1 fraction of globulins, and is considered to be one of the most potent serine protease inhibitors [85]. About 70–80% of this protein is produced and secreted into the bloodstream by liver cells, while the rest is synthesised by monocytes and macrophages in the respiratory system [86]. A1AT is one of the acute phase proteins; hence, its serum concentration fluctuates significantly and may increase rapidly in the course of inflammatory responses [87].
A number of studies have demonstrated that A1AT might be a potential biomarker in several cancers including GI cancers [61,88].
The study performed by Bernacka K et al. evaluated the concentrations of A1AT in the sera of GC patients in comparison to healthy volunteers [89]. The authors observed an increased concentration of A1AT in the blood of patients at the initial stage of the disease, and a correlation between the concentration of this protein and the stage of the disease, which suggests that it may have utility in the early detection of this cancer [89]. Similar results were demonstrated by Yang J et al., who revealed the upregulation of A1AT in the serum samples of GC patients [64]. In addition, the level of A1AT was found to be significantly higher in the gastric juice of GC patients in comparison to healthy controls [64]. The opposite results were obtained by Wu JY et al., who found decreased concentrations of A1AT in GC patients when compared to the control group [65]. Due to the fact that A1AT has an anti-inflammatory effect in vitro, thus contributing to the inhibition of the synthesis of pro-inflammatory cytokines such as IL-8 or TNF-alpha, it is suggested that the decreased level of this serpin may be responsible for the lack of the suppressive effect of these cytokines [65].
The association between the serum levels of A1AT and survival in PC patients was studied by Trichopoulos D et al. The authors indicated that the serum levels of A1AT represented a statistically significant prognostic indicator of PC [66]. Moreover, it was shown that the variable values of the difference in serum A1AT concentration at the time of diagnosis influenced the implications for the difference in survival [66].
The relationship between A1AT and CRC still seems controversial, as many studies show both higher and lower blood levels in CRC patients compared to control groups [67,68,69]. One study that evaluated the significance of A1AT in CRC patients was performed by Jaberie H et al. The authors found that the plasma levels of A1AT were significantly higher in CRC patients compared to those in a control group consisting of healthy volunteers, and those concentrations were found to positively correlate with the tumour stage [67]. These results are in line with previous studies involving the proteomic analysis of serum samples, which showed that A1AT outperformed serum CEA in distinguishing CRC patients from healthy controls [68]. However, it is worth remembering that elevated levels of A1AT in the blood are also observed with both benign neoplasms and other malignant neoplasms.
3.2.2. SERPIN A12 (Serpin Family A Member 12)—Vaspin
Vaspin (visceral adipose tissue-derived serine protease inhibitor—VASP) is a serpin that belongs to the family of adipokines. It is encoded by the OL-64 gene, and the locus is on chromosome 14 (14q32.1). VASP is a protein with a molecular weight of 45.2 kDa and about 40% homology with A1AT [90]. Increased concentrations of VASP have been identified in obesity, and it was also shown that elevated levels are associated with diabetes mellitus, metabolic syndrome, obesity and coronary artery diseases [91]. Interactions between VASP, cell apoptosis and proliferation have been already indicated. These relations, on the one hand, are key processes for homeostasis and, on the other hand, through their dysfunction and dysregulation, may predispose to the development and progression of many diseases, including cancer [92,93]. However, the exact link between VASP and cancer is still not entirely clear. According to current knowledge, it is known that most adipokines are capable of promoting tumour progression mainly by enhancing the proliferation and migration of tumour cells as well as inflammatory pathways [94,95]. Another mechanism is the participation of these proteins in anti-apoptotic pathways, which may consequently lead to the formation of cancer metastases [93,96]. With regard to GI cancers, the role of VASP remains a mystery. Thus far, scientists have studied its significance in relation to hepatocellular carcinoma (HCC) and CRC.
The study by Pazgan-Simon M et al. investigated the relationship between adipokines and metabolic abnormalities as well as the severity of liver dysfunction. The authors assessed the VASP serum levels in HCC patients in comparison to a control group as well as the association between those concentrations and HCC grade and progression [70]. The authors showed a positive association between increased VASP concentrations and HCC, with upregulated VASP concentrations in the sera of patients with LC when compared to healthy volunteers [70].
Fazeli MS et al. assessed the relation of serum VASP concentrations in patients with CRC [71]. Significantly higher levels of VASP were observed in the research group compared to the healthy controls. This finding suggests that this serpin may also be involved in carcinogenicity [71]. Studies report that VASP, by regulating the PI3K/Akt signalling pathway, has the potential to protect vascular endothelial cells from free fatty acid-induced apoptosis [71,72]. Therefore, it is suggested that VASP may prevent apoptosis, by influencing the above pathway [71,72].
3.2.3. SERPIN B5 (Serpin Family B Member 5)—Maspin
Maspin (mammary serine protease inhibitor—MASP) is a 42 kDa serine protease that belongs to the family of ovalbumin-like serpins called ov-serpins [97]. MASP has been proven to be expressed in the epithelia of many human organs such as the thymus, breast, small intestine and colon [98]. Although this serpin is commonly expressed in the cytoplasm, it can also be detected in the nucleus [99]. It has been proven that this protein plays an extremely important role in both the promotion of tumour cells’ adhesion and the inhibition of their mobility [100]. Although MASP has been characterised as a tumour suppressor, its overexpression is suggested to play a key role in tumour progression as well as enhanced aggressiveness for various cancers such as breast, gastric and pancreatic [101,102]. However, some investigations have reached opposite conclusions for CRC [103]. In the context of GI tumours, this serpin has been studied in patients with ESCC, GC, PC and CRC [104,105,106].
The association between MASP expression and better overall survival in ESCC was studied by Wang et al. The authors revealed that cancer patients who showed low or moderate MASP expression had lower postoperative survival rates than those who showed high MASP expression [73]. This leads to the conclusion that the transient regulation of MASP in the early stage of ESCC development may be a kind of defence mechanism, which prevents further progression to more malignant phenotypes, ultimately inhibiting tumour progression.
Wang MC et al. evaluated the significance of MASP expression in GC progression [74]. The expression of MASP was significantly negatively associated with both the invasive depth and metastasis, but not with tumour size and TNM stage [74]. Decreased MASP expression has been suggested to contribute to GC progression through mechanisms such as reducing cell apoptosis as well as facilitating angiogenesis [74]. Therefore, MASP has the potential to be considered as a marker of the biological behaviour of GC.
The study by Snoeren N et al. aimed to identify genes and proteins in CRC patients with liver metastases that may correlate with early disease recurrence [75]. Based on immunohistochemical analysis, elevated MASP expression was observed in the central tumour cores and also associated with poor histological grade in CRC patients. In addition, MASP was shown to be an independent predictor of the time to recurrence and survival specifically for CRC in stage III of the disease [75].
3.2.4. SERPIN E1 (Serpin Family E Member 1)—PAI-1 (Plasminogen Activator Inhibitor-1)
PAI-1 is a glycoprotein member of the serpin superfamily encoded by SERPINE1 that plays an important role in the plasminogen/plasmin system [107]. It is produced by hepatocytes, endothelial cells, megakaryocytes and platelets [108]. It has been shown that PAI-1 circulates in the blood in three different forms: active, latent inactive, or in the form of complexes with uPA, tPA, or vitronectin [109]. Elevated PAI-1 levels have been shown to be a risk factor for diseases such as thrombosis and atherosclerosis [110]. This molecule plays an extremely important role, among others, in cardiovascular diseases and, as a member of the serpin family, in cell migration [111]. However, there is evidence that, apart from playing important roles in cell adhesion, migration, or invasion, this protein is capable of inducing tumour vascularisation [56]. It has been shown that PAI-1 is highly expressed in many types of neoplastic tissues and consequently contributes to tumour progression by promoting cell proliferation and neoplastic metastasis [112].
The association between invasiveness, migration and prognosis as well as PAI-1 expression in ESCC patients was evaluated by Zhang Y et al. [76]. The authors revealed statistically significant differences in PAI-1 expression between ESCC tissue and normal oesophageal mucosa [76]. Additionally, the expression of the tested protein in patients with lymph node metastases (LNMs) was statistically significantly lower than that in patients without them [76]. The above suggests that PAI-1 may be a new molecular marker for predicting both the progression and prognosis of ESCC. The results show that extensive research on the PAI-1 protein could lead to its application as a therapeutic target [76]. The expression of PAI-1 in patients with ESCC was also studied by Wang D et al. [77]. The authors showed that the overexpression of PAI-1, through interaction with LRP1, could increase both the invasion and migration of ESCC cells, and also promote ESCC progression [77].
Chen H et al. evaluated the expression of the PAI-1 protein in the tissue as well as levels of PAI-1 in the plasma of CRC patients [78]. The protein expression was significantly increased in CRC tissue taken from patients with liver metastases when compared to samples from patients without liver metastases [78]. The authors also showed that the plasma PAI-1 levels were higher in CRC patients with liver metastases in comparison to the control group [78]. Furthermore, those results correlated with the tumour size, liver metastasis and LNM.
The study by Märkl B et al. investigated the association between the tissue level of PAI-1 and aggressive behaviour of colon cancer [79]. It was shown that increased PAI-1 was significantly associated with the occurrence of distant metastases [79]. In addition, multivariate analysis revealed PAI-1 as an independent predictive factor of distant metastases [79].
3.2.5. SERPIN F1 (Serpin Family F Member 1)—PEDF (Pigment Epithelium-Derived Factor)
PEDF is 50-kDa serpin, secreted primarily as a soluble glycoprotein. It has been found to have a broad spectrum of biological roles in many pathologies [113]. It is known as an important antitumorigenic, antiangiogenic and antimetastatic factor in several cancers [114,115]. A number of studies have demonstrated the potent tumour-suppressive effects of PEDF [116,117]. The molecular mechanisms of PEDF’s protection against cancer progression remain of interest to scientists [118]. Some theories suggest that it is related to the inhibition of tumour neovascularisation but also to the fact that PEDF is capable of downregulating vascular endothelial growth factor (VEGF) [119]. However, many studies still show many contradictions regarding the pro- and antitumour significance of this serpin. Previous research suggests that PEDF is involved in the induction of apoptosis through several pathways, suggesting it has potential for use in the treatment of cancer [120].
The significance of PEDF in GC was assessed. The study performed by Zhang Y et al. found that, in the case of GC, tumour growth and angiogenesis were inhibited by the injection of PEDF via the downregulation of both hypoxia-inducible factor 1 alpha (HIF-1 α) and vascular endothelial growth factor (VEGF) [80]. However, this serpin could not directly induce tumour cell apoptosis. The opposite results were obtained by Aksoy EK et al., who demonstrated statistically significantly higher PEDF concentrations in the sera of GI patients compared to a group comprising patients with precancerous lesions (PCLs) [81].
Principe DR et al. showed decreased PEDF expression in human pancreatic tissue samples when compared to a control group comprising healthy volunteers [82]. In human PC, decreased levels of PEDF have been observed, both in the tissue and in the serum. This decrease is potentially related to increased tumour angiogenesis, as well as the occurrence of liver metastases and a poorer prognosis [82].
The expression of PEDF in CRC patients was investigated by Wågsäter D et al. [83]. The level of the PEDF protein was assessed in both the plasma and tumour tissue, as well as healthy tissue, obtained from CRC patients [83]. It was shown that the concentration of PEDF in the plasma in the group of patients with CRC was significantly lower compared to that in the control group. Studies by Ji D et al. have shown that decreased PEDF expression in both sera and CRC tissues correlated with liver metastases [84]. In addition, cohort studies showed that patients with lower levels of PEDF in their CRC tissues had worse overall survival (OS) than patients with high levels of these proteins [84]. This led to the conclusion that PEDF could serve as a prognostic marker as well as a potential therapeutic target in CRC.
3.3. Serpins as Target for Cancer Therapy and Future Perspectives
Serpins are proteins with a proven role in many processes related to carcinogenesis, such as the proliferation of cancer cells and tumour progression. Moreover, their significance in the remission of selected neoplasms has also been demonstrated, which makes them a potential target for use in monitoring and, above all, in cancer therapy. An example of a serpin that can be used in this way is PAI-1. Although this protein has been shown to both reduce and increase tumour growth, recent studies suggest that blocking PAI-1—by modulating the function of the urokinase activator receptor—reduces cancer cell migration and survival [55,121].
Another anti-cancer serpin is MASP, which increases the sensitivity of cancer cells to apoptotic stimuli. According to PAI-1, MASP inhibits the migration of neoplastic cells. However, while significant experimental data support the role of MASP as a potential tumour suppressor, clinical data on the prognostic implications of its expression have led to conflicting results. This highlights the great need for a better understanding of MASP’s relationships in cancer biology. Moreover, research into recombinant MASP has led to the consideration of the question of whether rMaspin itself may be a viable candidate in the fight against cancer [100,122,123]. In contrast to the intracellular pro-apoptotic activity of MASP, extracellularly, its activity in vitro appears to be cytostatic, as cells have been found to regain both migratory activity and mobility after the discontinuation of rMaspin treatment. Some research assessed the prognostic value of MASP expression following fluarocyl treatment in CRC. These studies showed that low nuclear MASP expression appeared to be an independent predictor of benefits from adjuvant chemotherapy in patients with CRC [124].
PEDF is a multifunctional molecule that is highly antiangiogenic and pro-apoptotic. It also has the ability to inhibit tumour growth and distant metastasis by inducing tumour apoptosis and inhibiting tumour angiogenesis [60]. Recent studies have clearly shown that, in addition to the obvious antitumor activity of exogenous PEDF, changes in endogenous expression are associated with the progression of many malignancies [125]. Immunohistochemical analysis of PEDF expression in human neoplasms revealed that an increase in PEDF expression correlates with a better prognosis, which supports the idea that the determination of PEDF might be a prognostic factor in the treatment of cancer. Therefore, determining the level of PEDF in a tumour or the serum at the time of diagnosis could be an excellent source of information for delineating more or less aggressive treatment protocols to improve the therapeutic outcomes in cancer patients [60,120,126].
In designing serpin therapy, it is critical to consider issues such as protein half-lives, goals and clinical aspects that influence the development of metastatic proteins. Despite promising results, further research into the efficacy and mechanisms of serpin-mediated antitumor activity is needed and warranted in order to develop new alternatives for the treatment of cancer.
4. Conclusions
GI cancers are among the most common cancers in the world. The morbidity and mortality rates are extremely high for both women and men. Therefore, there is an urgent need to conduct further research on early GI tumour biomarkers. One group of proteins that seems to be controversial in terms of showing both pro- and anti-cancer properties is the serpins. This paper summarises the significance of selected serpins in the pathogenesis of GI neoplasms, confirming that selected serpin proteins play extremely important roles in the development and prognosis of GI cancers. The present manuscript demonstrates that, of all the serpins, A1AT may serve as a prognostic biomarker of PC and is observed to be increased in the initial stages of GC. In addition, elevated concentrations of vaspin are observed in the serum of HCC and CRC patients, suggesting that it plays a role in the pathogenesis of those cancers. Moreover, many of the described serpins such as maspin and PEDF are interesting targets for anti-cancer therapies, since lower concentrations and expression of them are associated with worse overall survival and the presence of distant metastases. This review summarises the role of serpins as biomarkers with potential use in GI cancers. Moreover, it has been shown that these proteins in GI cancers are useful not only in the context of screening or diagnosis, but also in prediction and monitoring of those malignancies. In addition, the assessment of diagnostic and predictive factors will most likely change the current stage of GI cancers. However, due to the non-specific nature of serpins and the additional observation of elevated levels of them in non-cancerous diseases, further research should be carried out to determine the most appropriate use of these molecules.
Author Contributions
Both authors produced the idea of the study and coordinated project funding. S.P. drafted the manuscript and B.M. substantively revised it. The visualisation was performed by S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Medical University of Bialystok, Poland. The APC was funded by Medical University of Bialystok, Poland.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
B.M. has received consultation and/or lecture honoraria from Abbott, Wiener, Roche, Cormay and Biameditek.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sensoy, I. A review on the food digestion in the digestive tract and the used in vitro models. Curr. Res. Food Sci. 2021, 4, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Nannini, G.; Meoni, G.; Amedei, A.; Tenori, L. Metabolomics profile in gastrointestinal cancers: Update and future perspectives. World J. Gastroenterol. 2020, 26, 2514–2532. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Lu, L.; Mullins, C.S.; Schafmayer, C.; Zeißig, S.; Linnebacher, M. A global assessment of recent trends in gastrointestinal cancer and lifestyle-associated risk factors. Cancer Commun. 2021, 41, 1137–1151. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Abnet, C.C.; Neale, R.E.; Vignat, J.; Giovannucci, E.L.; McGlynn, K.A.; Bray, F. Global Burden of 5 Major Types of Gastrointestinal Cancer. Gastroenterology 2020, 159, 335–349.e15. [Google Scholar] [CrossRef] [PubMed]
- Dizdar, Ö.; Kılıçkap, S. Global Epidemiology of Gastrointestinal Cancers; Yalcin, S., Philip, P.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–12. [Google Scholar]
- Thrift, A.P. Global burden and epidemiology of Barrett oesophagus and oesophageal cancer. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 432–443. [Google Scholar] [CrossRef]
- Murphy, N.; Jenab, M.; Gunter, M.J. Adiposity and gastrointestinal cancers: Epidemiology, mechanisms and future directions. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 659–670. [Google Scholar] [CrossRef]
- McMenamin, Ú.C.; McCain, S.; Kunzmann, A.T. Do smoking and alcohol behaviours influence GI cancer survival? Best Pract. Res. Clin. Gastroenterol. 2017, 31, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.C.; Meyerhardt, J.A. Obesity and Energy Balance in GI Cancer. J. Clin. Oncol. 2016, 34, 4217–4224. [Google Scholar] [CrossRef]
- D’Elia, L.; Galletti, F.; Strazzullo, P. Dietary salt intake and risk of gastric cancer. Cancer Treat. Res. 2014, 159, 83–95. [Google Scholar]
- Huang, F.L.; Yu, S.J. Esophageal cancer: Risk factors, genetic association, and treatment. Asian J. Surg. 2018, 41, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Yavuzsen, T.; Kazaz, N.; Tanriverdi, Ö.; Akman, T.; Davis, M.P. Symptom Management in Gastrointestinal Cancers; Yalcin, S., Philip, P.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 669–685. [Google Scholar]
- Pullmer, R.; Linden, W.; Rnic, K.; Vodermaier, A. Measuring symptoms in gastrointestinal cancer: A systematic review of assessment instruments. Support Care Cancer 2014, 22, 2941–2955. [Google Scholar] [CrossRef] [PubMed]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric cancer: Descriptive epidemiology, risk factors, screening, and prevention. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuntz, S.; Krieghoff-Henning, E.; Kather, J.N.; Jutzi, T.; Höhn, J.; Kiehl, L.; Hekler, A.; Alwers, E.; von Kalle, C.; Fröhling, S.; et al. Gastrointestinal cancer classification and prognostication from histology using deep learning: Systematic review. Eur. J. Cancer 2021, 155, 200–215. [Google Scholar] [CrossRef]
- Katabathina, V.S.; Menias, C.O.; Khanna, L.; Murphy, L.; Dasyam, A.K.; Lubner, M.G.; Prasad, S.R. Hereditary Gastrointestinal Cancer Syndromes: Role of Imaging in Screening, Diagnosis, and Management. Radiographics 2019, 39, 1280–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaosmanoglu, A.D.; Onur, M.R.; Arellano, R.S. Imaging in Gastrointestinal Cancers; Yalcin, S., Philip, P.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 445–464. [Google Scholar]
- Bouri, S.; Martin, J. Investigation of iron deficiency anaemia. Clin. Med. 2018, 18, 242–244. [Google Scholar] [CrossRef] [Green Version]
- Kuiken, N.S.S.; Rings, E.H.H.M.; Blijlevens, N.M.A.; Tissing, W.J.E. Biomarkers and non-invasive tests for gastrointestinal mucositis. Support Care Cancer 2017, 25, 2933–2941. [Google Scholar] [CrossRef] [Green Version]
- Li, L.S.; Guo, X.Y.; Sun, K. Recent advances in blood-based and artificial intelligence-enhanced approaches for gastrointestinal cancer diagnosis. World J. Gastroenterol. 2021, 27, 5666–5681. [Google Scholar] [CrossRef]
- Grady, W.M.; Yu, M.; Markowitz, S.D. Epigenetic Alterations in the Gastrointestinal Tract: Current and Emerging Use for Biomarkers of Cancer. Gastroenterology 2021, 160, 690–709. [Google Scholar] [CrossRef]
- De Mattos-Arruda, L.; Olmos, D.; Tabernero, J. Prognostic and predictive roles for circulating biomarkers in gastrointestinal cancer. Future Oncol. 2011, 7, 1385–1397. [Google Scholar] [CrossRef]
- Fiorica, F.; Stefanelli, A.; Pascale, G.; Fisichella, R. Elderly gastrointestinal cancer patients and radiochemotherapy: A review. Clin. Ter. 2014, 165, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Ludmir, E.B.; Palta, M.; Willett, C.G.; Czito, B.G. Total neoadjuvant therapy for rectal cancer: An emerging option. Cancer 2017, 123, 1497–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanani, A.; Veen, T.; Søreide, K. Neoadjuvant immunotherapy in primary and metastatic colorectal cancer. Br. J. Surg. 2021, 108, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Nakajima, M. Treatments for esophageal cancer: A review. Gen. Thorac. Cardiovasc. Surg. 2013, 61, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Bordry, N.; Astaras, C.; Ongaro, M.; Goossens, N.; Frossard, J.L.; Koessler, T. Recent advances in gastrointestinal cancers. World J. Gastroenterol. 2021, 27, 4493–4503. [Google Scholar] [CrossRef]
- Patel, S.R.; Reichardt, P. An updated review of the treatment landscape for advanced gastrointestinal stromal tumors. Cancer 2021, 127, 2187–2195. [Google Scholar] [CrossRef] [PubMed]
- Macha, M.A.; Seshacharyulu, P.; Krishn, S.R.; Pai, P.; Rachagani, S.; Jain, M.; Batra, S.K. MicroRNAs (miRNAs) as biomarker(s) for prognosis and diagnosis of gastrointestinal (GI) cancers. Curr. Pharm. Des. 2014, 20, 5287–5297. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.L.; Bhatia, V.; Agarwal, P.; Kumar, A. Gastrointestinal Cancers: Molecular Genetics and Biomarkers. Can. J. Gastroenterol. Hepatol. 2018, 2018, 4513860. [Google Scholar] [CrossRef]
- Kryvalap, Y.; Czyzyk, J. The Role of Proteases and Serpin Protease Inhibitors in β-Cell Biology and Diabetes. Biomolecules 2022, 12, 67. [Google Scholar] [CrossRef]
- Castro, H.C.; Abreu, P.A.; Geraldo, R.B.; Martins, R.C.; dos Santos, R.; Loureiro, N.I.; Cabral, L.M.; Rodrigues, C.R. Looking at the proteases from a simple perspective. J. Mol. Recognit. 2011, 24, 165–181. [Google Scholar] [CrossRef]
- Lucas, A.; Yaron, J.R.; Zhang, L.; Ambadapadi, S. Overview of Serpins and Their Roles in Biological Systems. Methods Mol. Biol. 2018, 1826, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J. Serpins in cartilage and osteoarthritis: What do we know? Biochem. Soc. Trans. 2021, 49, 1013–1026. [Google Scholar] [CrossRef] [PubMed]
- Van Gent, D.; Sharp, P.; Morgan, K.; Kalsheker, N. Serpins: Structure, function and molecular evolution. Int. J. Biochem. Cell Biol. 2003, 35, 1536–1547. [Google Scholar] [CrossRef]
- Law, R.H.; Zhang, Q.; McGowan, S.; Buckle, A.M.; Silverman, G.A.; Wong, W.; Rosado, C.J.; Langendorf, C.G.; Pike, R.N.; Bird, P.I.; et al. An overview of the serpin superfamily. Genome Biol. 2006, 7, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askew, D.J.; Silverman, G.A. Intracellular and extracellular serpins modulate lung disease. J. Perinatol. 2008, 28 (Suppl. 3), S127–S135. [Google Scholar] [CrossRef] [Green Version]
- Lucas, A.; Yaron, J.R.; Zhang, L.; Macaulay, C.; McFadden, G. Serpins: Development for Therapeutic Applications. Methods Mol. Biol. 2018, 1826, 255–265. [Google Scholar] [CrossRef]
- Valiente, M.; Obenauf, A.C.; Jin, X.; Chen, Q.; Zhang, X.H.; Lee, D.J.; Chaft, J.E.; Kris, M.G.; Huse, J.T.; Brogi, E.; et al. Serpins promote cancer cell survival and vascular co-option in brain metastasis. Cell 2014, 156, 1002–1016. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Singh, P.; Azhar, A.; Naseem, A.; Rashid, Q.; Kabir, M.A.; Jairajpuri, M.A. Serpin Inhibition Mechanism: A Delicate Balance between Native Metastable State and Polymerization. J. Amino Acids 2011, 2011, 606797. [Google Scholar] [CrossRef] [Green Version]
- Gatto, M.; Iaccarino, L.; Ghirardello, A.; Bassi, N.; Pontisso, P.; Punzi, L.; Shoenfeld, Y.; Doria, A. Serpins, immunity and autoimmunity: Old molecules, new functions. Clin. Rev. Allergy Immunol. 2013, 45, 267–280. [Google Scholar] [CrossRef]
- Sanrattana, W.; Maas, C.; de Maat, S. SERPINs-From Trap to Treatment. Front. Med. 2019, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J.; Lomas, D.A. The molecular aetiology of the serpinopathies. Int. J. Biochem. Cell Biol. 2008, 40, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Gettins, P.G.; Olson, S.T. Inhibitory serpins. New insights into their folding, polymerization, regulation and clearance. Biochem. J. 2016, 473, 2273–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazari, Y.M.; Bashir, A.; Habib, M.; Bashir, S.; Habib, H.; Qasim, M.A.; Shah, N.N.; Haq, E.; Teckman, J.; Fazili, K.M. Alpha-1-antitrypsin deficiency: Genetic variations, clinical manifestations and therapeutic interventions. Mutat. Res. Rev. Mutat. Res. 2017, 773, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M.; Gonzalez, F.J. The Evolution of Carcinogenesis. Toxicol. Sci. 2018, 165, 272–276. [Google Scholar] [CrossRef] [Green Version]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, H.J.; Strange, R.; Schedin, P.J. Apoptosis in the genesis and prevention of cancer. Cancer Epidemiol Biomark. Prev. 1992, 1, 597–602. [Google Scholar]
- Tian, S.; Peng, P.; Li, J.; Deng, H.; Zhan, N.; Zeng, Z.; Dong, W. SERPINH1 regulates EMT and gastric cancer metastasis via the Wnt/β-catenin signaling pathway. Aging 2020, 12, 3574–3593. [Google Scholar] [CrossRef]
- Duffy, M.J.; McGowan, P.M.; Harbeck, N.; Thomssen, C.; Schmitt, M. uPA and PAI-1 as biomarkers in breast cancer: Validated for clinical use in level-of-evidence-1 studies. Breast Cancer Res. 2014, 16, 428. [Google Scholar] [CrossRef] [Green Version]
- Andreasen, P.A. PAI-1—A potential therapeutic target in cancer. Curr. Drug Targets 2007, 8, 1030–1041. [Google Scholar] [CrossRef]
- Katayama, S.; Koyama, K.; Shima, J.; Tonai, K.; Goto, Y.; Koinuma, T.; Nunomiya, S. Thrombomodulin, Plasminogen Activator Inhibitor-1 and Protein C Levels, and Organ Dysfunction in Sepsis. Crit. Care Explor. 2019, 1, e0013. [Google Scholar] [CrossRef]
- Swiatkowska, M.; Szemraj, J.; Cierniewski, C.S. Induction of PAI-1 expression by tumor necrosis factor alpha in endothelial cells is mediated by its responsive element located in the 4G/5G site. FEBS J. 2005, 272, 5821–5831. [Google Scholar] [CrossRef] [PubMed]
- Kubala, M.H.; DeClerck, Y.A. The plasminogen activator inhibitor-1 paradox in cancer: A mechanistic understanding. Cancer Metastasis Rev. 2019, 38, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Placencio, V.R.; DeClerck, Y.A. Plasminogen Activator Inhibitor-1 in Cancer: Rationale and Insight for Future Therapeutic Testing. Cancer Res. 2015, 75, 2969–2974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwaki, T.; Urano, T.; Umemura, K. PAI-1, progress in understanding the clinical problem and its aetiology. Br. J. Haematol. 2012, 157, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Filleur, S.; Nelius, T.; de Riese, W.; Kennedy, R.C. Characterization of PEDF: A multi-functional serpin family protein. J. Cell. Biochem. 2009, 106, 769–775. [Google Scholar] [CrossRef]
- Brook, N.; Brook, E.; Dass, C.R.; Chan, A.; Dharmarajan, A. Pigment Epithelium-Derived Factor and Sex Hormone-Responsive Cancers. Cancers 2020, 12, 3483. [Google Scholar] [CrossRef] [PubMed]
- Becerra, S.P.; Notario, V. The effects of PEDF on cancer biology: Mechanisms of action and therapeutic potential. Nat. Rev. Cancer 2013, 13, 258–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphries, J.M.; Penno, M.A.; Weiland, F.; Klingler-Hoffmann, M.; Zuber, A.; Boussioutas, A.; Ernst, M.; Hoffmann, P. Identification and validation of novel candidate protein biomarkers for the detection of human gastric cancer. Biochim. Biophys. Acta. 2014, 1844, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Kan, J.Y.; Lee, Y.C.; Lin, Y.D.; Ho, W.Y.; Moi, S.H. Effect of Baseline Characteristics and Tumor Burden on Vaspin Expression and Progressive Disease in Operable Colorectal Cancer. Diagnostics 2020, 10, 801. [Google Scholar] [CrossRef] [PubMed]
- Sheng, S. The promise and challenge toward the clinical application of maspin in cancer. Front. Biosci. 2004, 9, 2733–2745. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Xiong, X.; Wang, X.; Guo, B.; He, K.; Huang, C. Identification of peptide regions of SERPINA1 and ENOSF1 and their protein expression as potential serum biomarkers for gastric cancer. Tumour Biol. 2015, 36, 5109–5118. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Cheng, C.C.; Wang, J.Y.; Wu, D.C.; Hsieh, J.S.; Lee, S.C.; Wang, W.M. Discovery of tumor markers for gastric cancer by proteomics. PLoS ONE 2014, 9, e84158. [Google Scholar] [CrossRef] [PubMed]
- Trichopoulos, D.; Tzonou, A.; Kalapothaki, V.; Sparos, L.; Kremastinou, T.; Skoutari, M. Alpha 1-antitrypsin and survival in pancreatic cancer. Int. J. Cancer 1990, 45, 685–686. [Google Scholar] [CrossRef] [PubMed]
- Jaberie, H.; Hosseini, S.V.; Naghibalhossaini, F. Evaluation of Alpha 1-Antitrypsin for the Early Diagnosis of Colorectal Cancer. Pathol. Oncol. Res. 2020, 26, 1165–1173. [Google Scholar] [CrossRef]
- Yin, H.R.; Xie, L.Q.; Xu, Y.; Cai, S.J.; Yao, J.; Yang, P.Y.; Lu, H.J. Direct-S: A directed mass spectrometry method for biomarker verification in native serum. Analyst 2015, 140, 3654–3662. [Google Scholar] [CrossRef] [PubMed]
- Żelechowska, P.; Brzezińska-Błaszczyk, E.; Kusowska, A.; Kozłowska, E. The role of adipokines in the modulation of lymphoid lineage cell development and activity: An overview. Obes. Rev. 2020, 21, e13055. [Google Scholar] [CrossRef] [PubMed]
- Pazgan-Simon, M.; Kukla, M.; Zuwała-Jagiełło, J.; Derra, A.; Bator, M.; Menżyk, T.; Lekstan, A.; Grzebyk, E.; Simon, K. Serum visfatin and vaspin levels in hepatocellular carcinoma (HCC). PLoS ONE 2020, 15, e0227459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazeli, M.S.; Dashti, H.; Akbarzadeh, S.; Assadi, M.; Aminian, A.; Keramati, M.R.; Nabipour, I. Circulating levels of novel adipocytokines in patients with colorectal cancer. Cytokine 2013, 62, 81–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Li, X.; Wu, Y.; Duan, R.; Zhang, J.; Du, F.; Zhang, Q.; Li, Y.; Li, N. Effects of vaspin on pancreatic β cell secretion via PI3K/Akt and NF-κB signaling pathways. PLoS ONE 2017, 12, e0189722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sheng, S.; Zhang, J.; Dzinic, S.; Li, S.; Fang, F.; Wu, N.; Zheng, Q.; Yang, Y. Elevated maspin expression is associated with better overall survival in esophageal squamous cell carcinoma (ESCC). PLoS ONE 2013, 8, e63581, Erratum in PLoS ONE 2014, 9, e104715. [Google Scholar] [CrossRef]
- Wang, M.C.; Yang, Y.M.; Li, X.H.; Dong, F.; Li, Y. Maspin expression and its clinicopathological significance in tumorigenesis and progression of gastric cancer. World J. Gastroenterol. 2004, 10, 634–637. [Google Scholar] [CrossRef]
- Snoeren, N.; Emmink, B.L.; Koerkamp, M.J.; van Hooff, S.R.; Goos, J.A.; van Houdt, W.J.; de Wit, M.; Prins, A.M.; Piersma, S.R.; Pham, T.V.; et al. Maspin is a marker for early recurrence in primary stage III and IV colorectal cancer. Br. J. Cancer 2013, 109, 1636–1647. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.L.; Chen, H.M.; Pu, H.W.; Ma, W.J.; Li, X.M.; Ma, H.; Chen, X. Expression of Bmi-1 and PAI-1 in esophageal squamous cell carcinoma. World J. Gastroenterol. 2014, 20, 5533–5539. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yang, L.Y.; Liu, Z.; Yu, J.; Zhang, M.J.; Zhang, Y.; Cai, Y.; Xu, X.; Hao, J.J.; Wang, M.R. PAI-1 overexpression promotes invasion and migration of esophageal squamous carcinoma cells. Hereditas 2020, 42, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Peng, H.; Liu, W.; Sun, Y.; Su, N.; Tang, W.; Zhang, X.; Wang, J.; Cui, L.; Hu, P.; et al. Silencing of plasminogen activator inhibitor-1 suppresses colorectal cancer progression and liver metastasis. Surgery 2015, 158, 1704–1713. [Google Scholar] [CrossRef] [PubMed]
- Märkl, B.; Renk, I.; Oruzio, D.V.; Jähnig, H.; Schenkirsch, G.; Schöler, C.; Ehret, W.; Arnholdt, H.M.; Anthuber, M.; Spatz, H. Tumour budding, uPA and PAI-1 are associated with aggressive behaviour in colon cancer. J. Surg. Oncol. 2010, 102, 235–241. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, J.; Yang, X.; Shao, C.; Xu, Z.; Cheng, R.; Cai, W.; Ma, J.; Yang, Z.; Gao, G. Pigment epithelium-derived factor inhibits angiogenesis and growth of gastric carcinoma by down-regulation of VEGF. Oncol. Rep. 2011, 26, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, E.K.; Akpınar, M.Y.; Doğan, Ö.; Göktaş, Z.; Sapmaz, F.P.; Şimşek, G.G.; Uzman, M.; Nazlıgül, Y. Clinical Significance of Serum Vascular Endothelial Growth Factor, Pigment Epithelium-Derived Factor, Tumor Necrosis Factor Alpha, and Progranulin Levels in Patients with Gastric Cancer and Gastric Precancerous Lesions. J. Gastrointest. Cancer 2019, 50, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Principe, D.R.; DeCant, B.; Diaz, A.M.; Mangan, R.J.; Hwang, R.; Lowy, A.; Shetuni, B.B.; Sreekumar, B.K.; Chung, C.; Bentrem, D.J.; et al. PEDF inhibits pancreatic tumorigenesis by attenuating the fibro-inflammatory reaction. Oncotarget 2016, 7, 28218–28234. [Google Scholar] [CrossRef] [PubMed]
- Wågsäter, D.; Löfgren, S.; Zar, N.; Hugander, A.; Dimberg, J. Pigment epithelium-derived factor expression in colorectal cancer patients. Cancer Investig. 2010, 28, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Li, M.; Zhan, T.; Yao, Y.; Shen, J.; Tian, H.; Zhang, Z.; Gu, J. Prognostic role of serum AZGP1, PEDF and PRDX2 in colorectal cancer patients. Carcinogenesis 2013, 34, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bteich, M. An overview of albumin and alpha-1-acid glycoprotein main characteristics: Highlighting the roles of amino acids in binding kinetics and molecular interactions. Heliyon 2019, 5, e02879. [Google Scholar] [CrossRef] [PubMed]
- Bianchera, A.; Alomari, E.; Bruno, S. Augmentation Therapy with Alpha-1 Antitrypsin: Present and Future of Production, Formulation, and Delivery. Curr. Med. Chem. 2022, 29, 385–410. [Google Scholar] [CrossRef] [PubMed]
- Schrödl, W.; Büchler, R.; Wendler, S.; Reinhold, P.; Muckova, P.; Reindl, J.; Rhode, H. Acute phase proteins as promising biomarkers: Perspectives and limitations for human and veterinary medicine. Proteom. Clin. Appl. 2016, 10, 1077–1092. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.Q.; Zhao, C.; Cai, S.J.; Xu, Y.; Huang, L.Y.; Bian, J.S.; Shen, C.P.; Lu, H.J.; Yang, P.Y. Novel proteomic strategy reveal combined alpha1 antitrypsin and cathepsin D as biomarkers for colorectal cancer early screening. J. Proteome Res. 2010, 9, 4701–4709. [Google Scholar] [CrossRef]
- Bernacka, K.; Kuryliszyn-Moskal, A.; Sierakowski, S. The levels of alpha 1-antitrypsin and alpha 1-antichymotrypsin in the sera of patients with gastrointestinal cancers during diagnosis. Cancer 1988, 62, 1188–1193. [Google Scholar] [CrossRef]
- Heiker, J.T.; Klöting, N.; Kovacs, P.; Kuettner, E.B.; Sträter, N.; Schultz, S.; Kern, M.; Stumvoll, M.; Blüher, M.; Beck-Sickinger, A.G. Vaspin inhibits kallikrein 7 by serpin mechanism. Cell. Mol. Life Sci. 2013, 70, 2569–2583. [Google Scholar] [CrossRef] [Green Version]
- Dimova, R.; Tankova, T. The role of vaspin in the development of metabolic and glucose tolerance disorders and atherosclerosis. Biomed. Res. Int. 2015, 2015, 823481. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Z.; Guan, Z. Vaspin deficiency failed to promote the proliferation of BMSCs in osteoarthritis. Int. J. Rheum. Dis. 2021, 24, 90–95. [Google Scholar] [CrossRef]
- Pich, K.; Respekta, N.; Dawid, M.; Mlyczynska, E.; Kurowska, P.; Rak, A. New insights into cell apoptosis and proliferation: The potential role of vaspin. J. Physiol. Pharmacol. 2021, 72, 831–844. [Google Scholar] [CrossRef]
- Christodoulatos, G.S.; Spyrou, N.; Kadillari, J.; Psallida, S.; Dalamaga, M. The Role of Adipokines in Breast Cancer: Current Evidence and Perspectives. Curr. Obes. Rep. 2019, 8, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Perego, S.; Sansoni, V.; Ziemann, E.; Lombardi, G. Another Weapon against Cancer and Metastasis: Physical-Activity-Dependent Effects on Adiposity and Adipokines. Int. J. Mol. Sci. 2021, 22, 2005. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Jiang, Y.; Shan, P.F.; Shen, J.; Liang, Q.H.; Cui, R.R.; Liu, Y.; Liu, G.Y.; Wu, S.S.; Lu, Q.; et al. Vaspin attenuates the apoptosis of human osteoblasts through ERK signaling pathway. Amino Acids 2013, 44, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Khalkhali-Ellis, Z. Maspin: The new frontier. Clin. Cancer Res. 2006, 12, 7279–7283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockett, J.; Yin, S.; Li, X.; Meng, Y.; Sheng, S. Tumor suppressive maspin and epithelial homeostasis. J. Cell. Biochem. 2006, 97, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Sheng, S. A role of novel serpin maspin in tumor progression: The divergence revealed through efforts to converge. J. Cell. Physiol. 2006, 209, 631–635. [Google Scholar] [CrossRef]
- Bodenstine, T.M.; Seftor, R.E.; Khalkhali-Ellis, Z.; Seftor, E.A.; Pemberton, P.A.; Hendrix, M.J. Maspin: Molecular mechanisms and therapeutic implications. Cancer Metastasis Rev. 2012, 31, 529–551. [Google Scholar] [CrossRef]
- Zhang, M. Multiple functions of maspin in tumor progression and mouse development. Front. Biosci. 2004, 9, 2218–2226. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Zheng, H.; Tsuneyama, K.; Takahashi, H.; Nomoto, K.; Xu, H.; Takano, Y. Paradoxical expression of maspin in gastric carcinomas: Correlation with carcinogenesis and progression. Hum. Pathol. 2007, 38, 1248–1255. [Google Scholar] [CrossRef]
- Gurzu, S.; Jung, I. Subcellular Expression of Maspin in Colorectal Cancer: Friend or Foe. Cancers 2021, 13, 366. [Google Scholar] [CrossRef]
- Meng, H.; Guan, X.; Guo, H.; Xiong, G.; Yang, K.; Wang, K.; Bai, Y. Association between SNPs in Serpin gene family and risk of esophageal squamous cell carcinoma. Tumour Biol. 2015, 36, 6231–6238. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chang, L.L. Maspin suppresses cell invasion and migration in gastric cancer through inhibiting EMT and angiogenesis via ITGB1/FAK pathway. Hum. Cell 2020, 33, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Banias, L.; Jung, I.; Bara, T.; Fulop, Z.; Simu, P.; Simu, I.; Satala, C.; Gurzu, S. Immunohistochemical-Based molecular subtyping of colorectal carcinoma using maspin and markers of epithelial-mesenchymal transition. Oncol. Lett. 2020, 19, 1487–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlov, M.; Ćelap, I. Plasminogen activator inhibitor 1 in acute coronary syndromes. Clin. Chim. Acta. 2019, 491, 52–58. [Google Scholar] [CrossRef] [PubMed]
- De Faria, C.A.; Zanette, D.L.; Silva, W.A., Jr.; Ribeiro-Paes, J.T. PAI-1 inhibition by simvastatin as a positive adjuvant in cell therapy. Mol. Biol. Rep. 2019, 46, 1511–1517. [Google Scholar] [CrossRef]
- Cesari, M.; Pahor, M.; Incalzi, R.A. Plasminogen activator inhibitor-1 (PAI-1): A key factor linking fibrinolysis and age-related subclinical and clinical conditions. Cardiovasc. Ther. 2010, 28, e72–e91. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, D.E. PAI-1 and atherothrombosis. J. Thromb. Haemost. 2005, 3, 1879–1883. [Google Scholar] [CrossRef]
- Sakamoto, H.; Koma, Y.I.; Higashino, N.; Kodama, T.; Tanigawa, K.; Shimizu, M.; Fujikawa, M.; Nishio, M.; Shigeoka, M.; Kakeji, Y.; et al. PAI-1 derived from cancer-associated fibroblasts in esophageal squamous cell carcinoma promotes the invasion of cancer cells and the migration of macrophages. Lab. Investig. 2021, 101, 353–368. [Google Scholar] [CrossRef]
- Hogan, N.M.; Joyce, M.R.; Murphy, J.M.; Barry, F.P.; O’Brien, T.; Kerin, M.J.; Dwyer, R.M. Impact of mesenchymal stem cell secreted PAI-1 on colon cancer cell migration and proliferation. Biochem. Biophys. Res. Commun. 2013, 435, 574–579. [Google Scholar] [CrossRef]
- Tombran-Tink, J. The neuroprotective and angiogenesis inhibitory serpin, PEDF: New insights into phylogeny, function, and signaling. Front. Biosci. 2005, 10, 2131–2149. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, T.; Yamagishi, S.I.; Sata, M. Structure-function relationships of PEDF. Curr. Mol. Med. 2010, 10, 302–311. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Cheng, R.; Benyajati, S.; Ma, J.X. PEDF and its roles in physiological and pathological conditions: Implication in diabetic and hypoxia-induced angiogenic diseases. Clin. Sci. 2015, 128, 805–823. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Huang, Z.; Zhu, L.; Yu, X.; Gao, T.; Feng, J.; Hong, H.; Yin, H.; Zhou, T.; Qi, W.; et al. The contrary intracellular and extracellular functions of PEDF in HCC development. Cell Death Dis. 2019, 10, 742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Jin, Q.; Zeng, J.; Yu, T.; Chen, Y.; Li, S.; Gong, D.; He, L.; Tan, X.; Yang, L.; et al. Combined Tumor- and Neovascular-“Dual Targeting” Gene/Chemo-Therapy Suppresses Tumor Growth and Angiogenesis. ACS Appl. Mater. Interfaces 2016, 8, 25753–25769. [Google Scholar] [CrossRef]
- Wu, Q.J.; Gong, C.Y.; Luo, S.T.; Zhang, D.M.; Zhang, S.; Shi, H.S.; Lu, L.; Yan, H.X.; He, S.S.; Li, D.D.; et al. AAV-Mediated human PEDF inhibits tumor growth and metastasis in murine colorectal peritoneal carcinomatosis model. BMC Cancer 2012, 12, 129. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.X.; Wang, J.J.; Gao, G.; Parke, K.; Ma, J.X. Pigment epithelium-derived factor downregulates vascular endothelial growth factor (VEGF) expression and inhibits VEGF-VEGF receptor 2 binding in diabetic retinopathy. J. Mol. Endocrinol. 2006, 37, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Zeng, J.; Huang, H.; Ma, C.; Wang, L.; Wang, F.; Liao, X.; Song, X. Cancer-Targeted PEDF-DNA therapy for metastatic colorectal cancer. Int. J. Pharm. 2020, 576, 118999. [Google Scholar] [CrossRef]
- Noh, H.; Hong, S.; Huang, S. Role of urokinase receptor in tumor progression and development. Theranostics 2013, 3, 487–495. [Google Scholar] [CrossRef]
- Berardi, R.; Morgese, F.; Onofri, A.; Mazzanti, P.; Pistelli, M.; Ballatore, Z.; Savini, A.; De Lisa, M.; Caramanti, M.; Rinaldi, S.; et al. Role of maspin in cancer. Clin. Transl. Med. 2013, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Bass, R.; Fernández, A.M.; Ellis, V. Maspin inhibits cell migration in the absence of protease inhibitory activity. J. Biol. Chem. 2002, 277, 46845–46848. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, Z.; Jung, I.; Szalman, K.; Banias, L.; Bara, T.J.; Gurzu, S. Interaction of arylsulfatases A and B with maspin: A possible explanation for dysregulation of tumor cell metabolism and invasive potential of colorectal cancer. World J. Clin. Cases 2019, 7, 3990–4003. [Google Scholar] [CrossRef] [PubMed]
- Broadhead, M.L.; Dass, C.R.; Choong, P.F. In vitro and in vivo biological activity of PEDF against a range of tumors. Expert Opin. Ther. Targets 2009, 13, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Broadhead, M.L.; Dass, C.R.; Choong, P.F. Cancer cell apoptotic pathways mediated by PEDF: Prospects for therapy. Trends Mol. Med. 2009, 15, 461–467. [Google Scholar] [CrossRef] [PubMed]
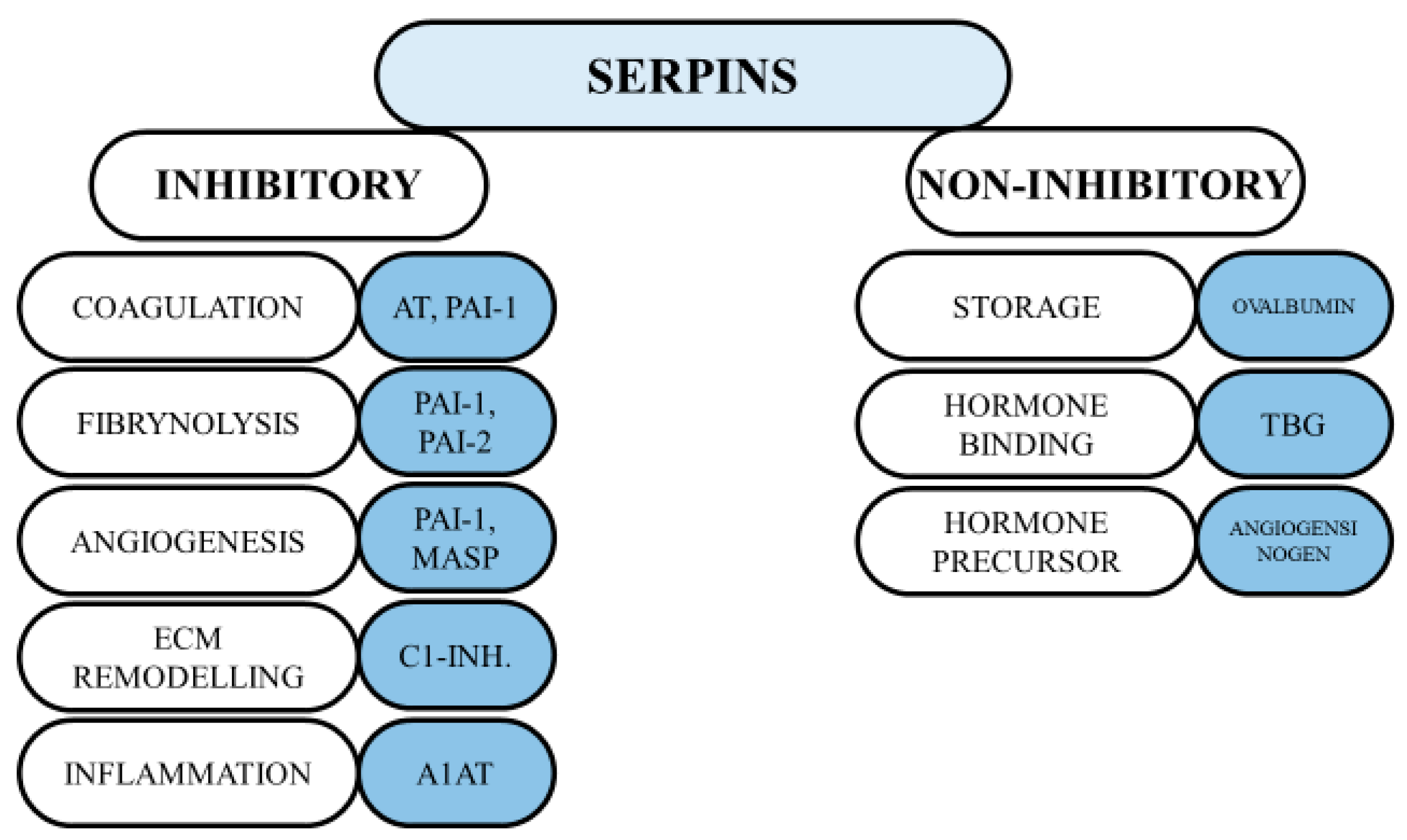
Figure 1.
The inhibitory and non-inhibitory function of serpins [43]. A1AT: alpha-1-antitripsin; AT: antithrombin; PAI-1: plasminogen activator inhibitor-1; PAI-2: plasminogen activator inhibitor-2; MASP: maspin; TBG: thyroxine binding globulin.
Figure 1.
The inhibitory and non-inhibitory function of serpins [43]. A1AT: alpha-1-antitripsin; AT: antithrombin; PAI-1: plasminogen activator inhibitor-1; PAI-2: plasminogen activator inhibitor-2; MASP: maspin; TBG: thyroxine binding globulin.

Figure 2.
The role of PAI-1 in cancer [56].
Figure 2.
The role of PAI-1 in cancer [56].


{kind=link}
{kind=link}
Table 1.
The roles of selected serpins in GI cancers.
| Serpin | Alternative Name | GI Cancer | Significance | References |
|---|---|---|---|---|
| SERPINA1 | A1AT | GC |
| [64,65,66,67,68,69] |
| PC |
| |||
| CRC |
| |||
| SERPINA12 | Vaspin | HCC |
| [70,71,72] |
| CRC |
| |||
| SERPINB5 | Maspin | ESCC |
| [73,74,75] |
| GC |
| |||
| CRC |
| |||
| SERPINE1 | PAI-1 | ESCC |
| [76,77,78,79] |
| CRC |
| |||
| SERPINF1 | PEDF | GC |
| [80,81,82,83,84] |
| PC |
| |||
| CRC |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pączek, S.; Mroczko, B. The Role of Selected Serpins in Gastrointestinal (GI) Malignancies. J. Clin. Med. 2022, 11, 6225. https://doi.org/10.3390/jcm11206225
AMA Style
Pączek S, Mroczko B. The Role of Selected Serpins in Gastrointestinal (GI) Malignancies. Journal of Clinical Medicine. 2022; 11(20):6225. https://doi.org/10.3390/jcm11206225
Chicago/Turabian StylePączek, Sara, and Barbara Mroczko. 2022. "The Role of Selected Serpins in Gastrointestinal (GI) Malignancies" Journal of Clinical Medicine 11, no. 20: 6225. https://doi.org/10.3390/jcm11206225
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.