
Angiogenesis and Multiple Sclerosis Pathogenesis: A Glance at New Pharmaceutical Approaches

 ,
,  and
and 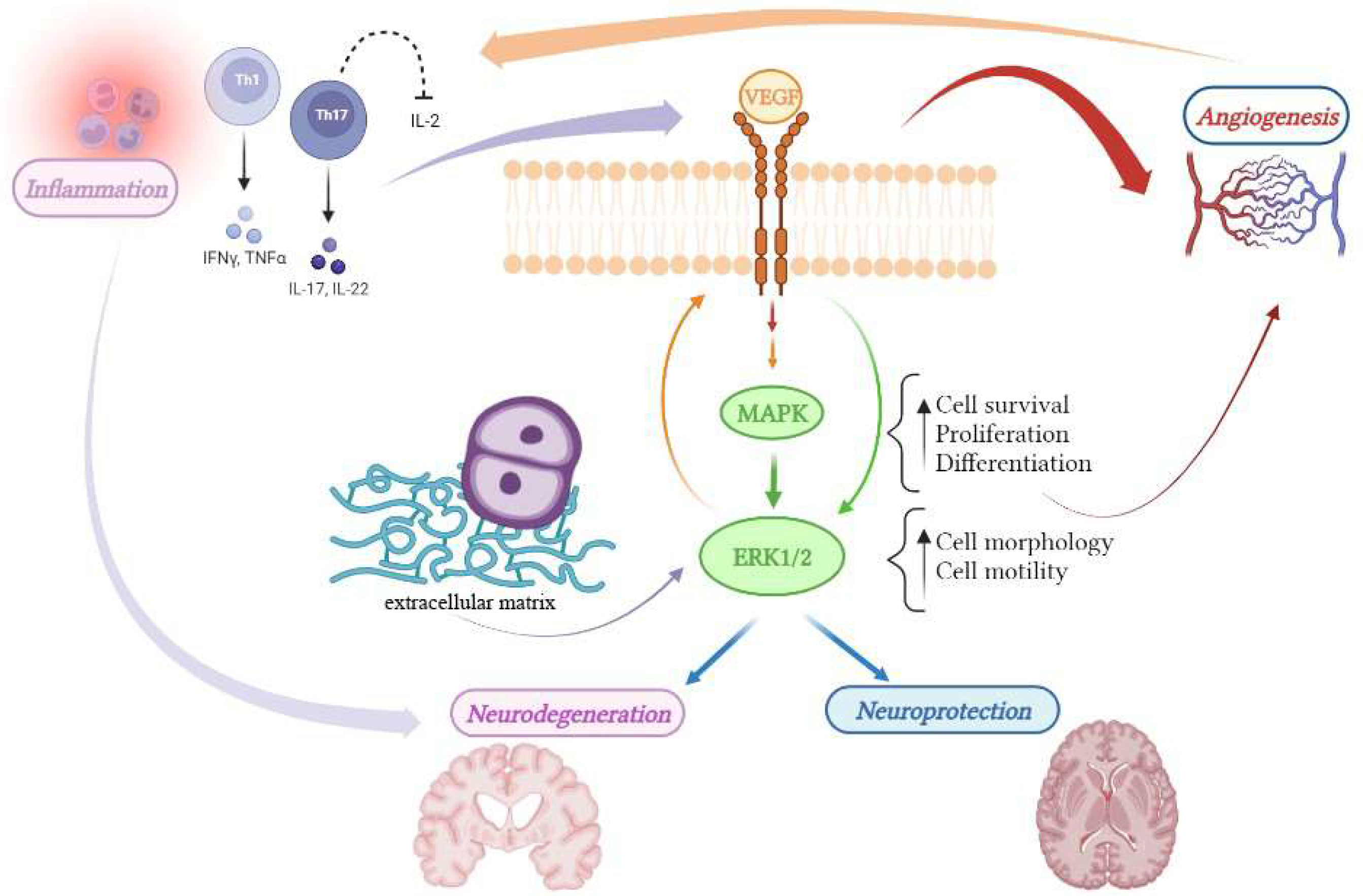{kind=link}
Abstract
:1. Introduction
2. Angiogenesis in Multiple Sclerosis
3. VEGF: A Double Edge Sword in the Treatment of MS
4. MEK/ERK1/2 Pathway: Essential to Angiogenesis Is Also a Hub Connecting Different Pathogenic Factors of MS
5. Therapy of Multiple Sclerosis
6. Natural Compound in the Therapy of Multiple Sclerosis
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lubin, F.D.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. Neurology 1996, 46, 906–911. [Google Scholar]
- Associazione Italiana Sclerosi Multipla (AISM). Available online: http://www.aism.it (accessed on 1 April 2016).
- Reich, D.S.; Lucchinetti, C.F.; Calabresi, P.A. Multiple Sclerosis. N. Engl. J. Med. 2018, 378, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Cagol, A.; Schaedelin, S.; Barakovic, M.; Benkert, P.; Todea, R.A.; Rahmanzadeh, R.; Galbusera, R.; Lu, P.J.; Weigel, M.; Melie-Garcia, L.; et al. Association of Brain Atrophy with Disease Progression Independent of Relapse Activity in Patients with Relapsing Multiple Sclerosis. JAMA Neurol. 2022, 79, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Grigoriadis, N.; van Pesch, V. A basic overview of multiple sclerosis immunopathology. Eur. J. Neurol. 2015, 22, 3–13. [Google Scholar] [CrossRef]
- Ransohoff, R.M.; Hafler, D.A.; Lucchinetti, C.F. Multiple sclerosis-a quiet revolution. Nat. Rev. Neurol. 2015, 11, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Lassmann, H. Pathogenic Mechanisms Associated with Different Clinical Courses of Multiple Sclerosis. Front. Immunol. 2019, 9, 3116. [Google Scholar] [CrossRef]
- Girolamo, F.; Coppola, C.; Ribatti, D.; Trojano, M. Angiogenesis in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol. Commun. 2014, 2, 84. [Google Scholar] [CrossRef]
- Carmeliet, P. Angiogenesis in life, disease and medicine. Nature 2005, 438, 932e6. [Google Scholar] [CrossRef]
- Rajeev, V.; Fann, D.Y.; Dinh, Q.N.; Kim, H.A.; De Silva, T.M.; Lai, M.K.P.; Chen, C.L.-H.; Drummond, G.R.; Sobey, C.G.; Arumugam, T.V. Pathophysiology of blood brain barrier dysfunction during chronic cerebral hypoperfusion in vascular cognitive impairment. Theranostics 2022, 12, 1639–1658. [Google Scholar] [CrossRef]
- Gentile, M.T.; Poulet, R.; Di Pardo, A.; Cifelli, G.; Maffei, A.; Vecchione, C.; Passarelli, F.; Landolfi, A.; Carullo, P.; Lembo, G. Beta-amyloid deposition in brain is enhanced in mouse models of arterial hypertension. Neurobiol. Aging. 2009, 30, 222–228. [Google Scholar] [CrossRef]
- Kim, D.Y.; Jeoung, D.; Ro, J.Y. Signaling pathways in the activation of mast cells cocultured with astrocytes and colocalization of both cells in experimental allergic encephalomyelitis. J. Immunol. 2010, 185, 273–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buch, S.; Subramanian, K.; Jella, P.K.; Chen, Y.; Wu, Z.; Shah, K.; Bernitsas, E.; Ge, Y.; Haacke, E.M. Revealing vascular abnormalities and measuring small vessel density in multiple sclerosis lesions using USPIO. NeuroImage Clin. 2021, 29, 102525. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Tamma, R.; Annese, T. Mast cells and angiogenesis in multiple sclerosis. Inflamm. Res. 2020, 69, 1103–1110. [Google Scholar] [CrossRef]
- Ferrara, N.; Kerbel, R.S. Angiogenesis as a therapeutic target. Nature 2005, 438, 967e74. [Google Scholar] [CrossRef] [PubMed]
- Caudio, L.; Raine, C.S.; Brosnan, C.F. Evidence of persistent bloodbrain barrier abnormalities in chronic-progressive multiple sclerosis. Acta Neuropathol. 1995, 90, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Kirk, J.; Plumb, J.; Mirakhur, M.; McQuaid, S. Tight junctional abnormality in multiple sclerosis white matter afects all calibres of vessel and is associated with blood-brain barrier leakage and active demyelination. J. Pathol. 2003, 201, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The chick embryo chorioallantoic membrane (CAM). A multifaceted experimental model. Mech. Dev. 2016, 141, 70–77. [Google Scholar] [CrossRef]
- Bester, M.; Forkert, N.D.; Stellmann, J.P.; Aly, L.; Drabik, A.; Young, K.L.; Heesen, C.; Fiehler, J.; Siemonsen, S. Increased Perfusion in Normal Appearing White Matter in High Inflammatory Multiple Sclerosis Patients. PLoS ONE 2015, 10, e0119356. [Google Scholar]
- Roscoe, W.A.; Welsh, M.E.; Carter, D.E.; Karlik, S.J. VEGF and angiogenesis in acute and chronic MOG (35–55) peptide induced EAE. J. Neuroimmunol. 2009, 209, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Pathak, A.P.; Schmainda, K.M.; Ward, B.D.; Linderman, J.R.; Rebro, K.J.; Greene, A.S. MR-derived cerebral blood volume maps: Issues regarding histological validation and assessment of tumor angiogenesis. Magn. Reson. Med. Off. J. Soc. Magn. Reson. Med. 2001, 46, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holley, J.E.; Newcombe, J.; Whatmore, J.L.; Gutowski, N.J. Increased blood vessel density and endothelial cell proliferation in multiple sclerosis cerebral white matter. Neurosci. Lett. 2010, 470, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Tumor angiogenesis and accessibility: Role of vascular endothelial growth factor. Semin. Oncol. 2002, 29, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Ludwin, S.K. The pathogenesis of multiple sclerosis: Relating human pathology to experimental studies. J. Neuropathol. Exp. Neurol. 2006, 65, 305–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polman, C.H.; O’Connor, P.W.; Havrdova, E.; Hutchinson, M.; Kappos, L.; Miller, D.H.; Phillips, J.T.; Lublin, F.D.; Giovannoni, G.; Wajgt, A.; et al. A randomized, placebo-controlled trial of natalizumab for relapsing multiple sclerosis. N. Engl. J. Med. 2006, 354, 899–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudick, R.A.; Stuart, W.H.; Calabresi, P.A.; Confavreux, C.; Galetta, S.L.; Radue, E.W.; Lublin, F.D.; Weinstock-Guttman, B.; Wynn, D.R.; Lynn, F.; et al. Natalizumab plus interferon beta-1a for relapsing multiple sclerosis. N. Engl. J. Med. 2006, 354, 911–923. [Google Scholar] [CrossRef] [Green Version]
- Khademi, M.; Bornsen, L.; Rafatnia, F.; Andersson, M.; Brundin, L.; Piehl, F.; Sellebjerg, F.; Olsson, T. The effects of natalizumab on inflammatory mediators in multiple sclerosis: Prospects for treatment-sensitive biomarkers. Eur. J. Neurol. 2009, 16, 528–536. [Google Scholar] [CrossRef]
- Iaffaldano, P.; Ruggieri, M.; Viterbo, R.G.; Mastrapasqua, M.; Trojano, M. The improvement of cognitive functions is associated with a decrease of plasma osteopontin levels in natalizumab treated relapsing multiple sclerosis. Brain Behav. Immun. 2014, 35, 176–181. [Google Scholar] [CrossRef]
- Kathy, K.; Delphine, M.; Gilles, D.; Gautier, P.; Olivier, T.; Le Mauff, B. Natalizumab in Multiple Sclerosis Treatment: From Biological Effects to Immune Monitoring. Front. Immunol. 2020, 11, 549842. [Google Scholar]
- Kacperska, M.J.; Walenczak, J.; Tomasik, B. Plasmatic microRNA as Potential Biomarkers of Multiple Sclerosis: Literature Review. Adv. Clin. Exp. Med. 2016, 25, 775–779. [Google Scholar] [CrossRef] [Green Version]
- Emery, B. Regulation of oligodendrocyte differentiation and myelination. Science 2010, 330, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Rajewsky, K. MicroRNA control in the immune system: Basic principles. Cell 2009, 136, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemond, C.C.; Healy, B.C.; Tauhid, S.; Mazzola, M.A.; Quintana, F.J.; Gandhi, R.; Weiner, H.L.; Bakshi, R. MRI phenotypes in MS: Longitudinal changes and miRNA signatures. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, e530. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Shi, C.; Pan, H.; Meng, X.; Ji, F. MicroRNA-22 exerts its neuroprotective and angiogenic functions via regulating PI3K/Akt signaling pathway in cerebral ischemia-reperfusion rats. J. Neural. Transm. 2020, 127, 35–44. [Google Scholar] [CrossRef]
- Wang, Z.; Ge, X.; Wang, Y.; Liang, Y.; Shi, H.; Zhao, T. Mechanism of dexmedetomidine regulating osteogenesis-angiogenesis coupling through the miR-361-5p/VEGFA axis in postmenopausal osteoporosis. Life Sci. 2021, 275, 119273. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.Y.; Gui, S.Y.; Zhang, P.; Wang, M. Upregulation of miR-345-5p suppresses cell growth of lung adenocarcinoma by regulating ras homolog family member A (RhoA) and Rho/Rho associated protein kinase (Rho/ROCK) pathway. Chin. Med. J. 2021, 134, 2619–2628. [Google Scholar] [CrossRef]
- Mycko, M.P.; Baranzini, S.E. MicroRNA and exosome profiling in multiple sclerosis. Mult. Scler. 2020, 26, 599–604. [Google Scholar] [CrossRef]
- Stromness, I.M.; Goverman, J.M. Active induction of experimental allergic encephalomyelitis. Nat. Protocol. 2006, 1, 1810–1819. [Google Scholar] [CrossRef]
- Seabrook, T.J.; Littlewood-Evans, A.; BrinkMann, V.; Pollinger, B.; Schnell, C.; Hiestand, P.C. Angiogenesis is present in experimental autoimmune encephalomyelitis and pro-angiogenic factors are increased in multiple sclerosis lesions. J. Neuroinflamm. 2010, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Boroujerdi, A.; Welser-Alves, J.V.; Milner, R. Exstensive vascular remodelling on the spinal cord of pre-symptomatic experimental autoimmune encephalomyelitis mice; increased vessel expression of fibronectin and the α5β1 integrin. Exp. Neurol. 2013, 250, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brambilla, R. The contribution of astrocytes to the neuroinflammatory response in multiple sclerosis and experimental autoimmune encephalomyelitis. Acta Neuropathol. 2019, 137, 757–783. [Google Scholar] [CrossRef]
- Kunkl, M.; Amormino, C.; Tedeschi, V.; Fiorillo, M.T.; Tuosto, L. Astrocytes and Inflammatory T Helper Cells: A Dangerous Liaison in Multiple Sclerosis. Front. Immunol. 2022, 13, 824411. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Wu, P.-F.; Ma, D.-L.; Zhang, W.; Sun, M. Growth Factors and Their Roles in Multiple Sclerosis Risk. Front. Immunol. 2021, 12, 768682. [Google Scholar]
- Lin, W. Neuroprotective effects of vascular endothelial growth factor A in the experimental autoimmune encephalomyelitis model of multiple sclerosis. Neural Regen. Res. 2017, 12, 70–71. [Google Scholar] [CrossRef] [PubMed]
- MacMillan, C.J.; Furlong, S.J.; Doucette, C.D.; Chen, P.L.; Easton ASHoskin, D.W. Bevacizumab diminishes experimental autoimmune encephalomyelitis by inhibiting spinal cord angiogenesis and reducing peripheral T-cell responses. J. Neuropathol. Exp. Neurol. 2012, 71, 983–999. [Google Scholar] [CrossRef] [Green Version]
- Dudvarski Stankovic, N.; Teodorczyk, M.; Ploen, R.; Zipp, F.; Schmidt, M.H. Microglia-blood vessel interactions: A double-edged sword in brain pathologies. Acta Neuropathol. 2016, 131, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Mor, F.; Quintana, F.J.; Cohen, I.R. Angiogenesis-inflammation cross-talk: Vascular endothelial growth factor is secreted by activated T cells and induces Th1 polarization. J. Immunol. 2004, 172, 4618–4623. [Google Scholar] [CrossRef] [Green Version]
- Rangachari, M.; Kuchroo, V.K. Using EAE to better understand principles of immune function and autoimmune pathology. J. Autoimmun. 2013, 45, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Totzeck, M.; Mincu, R.I.; Rassaf, T. Cardiovascular Adverse Events in Patients with Cancer Treated with Bevacizumab: A Meta-Analysis of More Than 20,000 Patients. J. Am. Heart Assoc. 2017, 6, e006278. [Google Scholar] [CrossRef] [Green Version]
- Herz, J.; Reitmeir, R.; Hagen, S.I.; Reinboth, B.S.; Guo, Z.; Zechariah, A.; El Ali, A.; Doeppner, T.R.; Bacigaluppi, M.; Pluchino, S.; et al. Intracerebroventricularly delivered VEGF promotes contralesional corticorubral plasticity after focal cerebral ischemia via mechanisms involving anti-inflammatory actions. Neurobiol. Dis. 2012, 45, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Zhang, Y.; Zhong, Y.; Ye, Y.; Hu, X.; Gu, L.; Xiong, X. Inflammation-Mediated Angiogenesis in Ischemic Stroke. Front. Cell. Neurosci. 2021, 15, 652647. [Google Scholar] [CrossRef]
- Simons, M. Angiogenesis: Where do we stand now? Circulation 2005, 111, 1556–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, R.; Duda, D.G.; Clark, J.W.; Loeffler, J.S. Lessons from phase III clinical trials on anti-VEGF therapy for cancer. Nat. Clin. Pract. Oncol. 2006, 3, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Beckner, M.E. Factors promoting tumor angiogenesis. Cancer Investig. 1999, 17, 594–623. [Google Scholar] [CrossRef] [PubMed]
- Colucci-D’Amato, L.; Perrone-Capano, C.; di Porzio, U. Chronic activation of ERK and neurodegenerative diseases. Bioessays 2003, 25, 1085–1095. [Google Scholar] [CrossRef]
- Coultas, L.; Chawengsaksophak, K.; Rossant, J. Endothelial cells and VEGF in vascular development. Nature 2005, 438, 937–945. [Google Scholar] [CrossRef]
- Abdelsaid, M.A.; El-Remessy, A.B. S-glutathionylation of LMW-PTP regulates VEGF-mediated FAK activation and endothelial cell migration. J. Cell Sci. 2012, 125, 4751–4760. [Google Scholar] [CrossRef] [Green Version]
- Kofler, N.M.; Simons, M. Angiogenesis versus arteriogenesis: Neuropilin 1 modulation of VEGF signaling. F1000prime Rep. 2015, 7, 26. [Google Scholar] [CrossRef]
- Burotto, M.; Chiou, V.L.; Lee, J.M.; Kohn, E.C. The MAPK pathway across different malignancies: A new perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, H.; Colucci-D’Amato, L.G.; Charlet-Berguerand, N.; Plouin, P.F.; Bertagna, X.; de Franciscis, V.; Thermes, C. High levels of tyrosine phosphorylated proto-ret in sporadic phenochromocytomas. Cancer Res. 2000, 60, 1365–1370. [Google Scholar]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Colucci-D’Amato, G.L.; D’Alessio, A.; Califano, D.; Cali, G.; Rizzo, C.; Nitsch, L.; Santelli, G.; de Franciscis, V. Abrogation of nerve growth factor-induced terminal differentiation by ret oncogene involves perturbation of nuclear translocation of ERK. J. Biol. Chem. 2000, 275, 19306–19314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Califano, D.; Rizzo, C.; D’Alessio, A.; Colucci-D’Amato, G.L.; Cali, G.; Bartoli, P.C.; Santelli, G.; Vecchio, G.; de Franciscis, V. Signaling through Ras is essential for ret oncogene-induced cell differentiation in PC12 cells. J. Biol. Chem. 2000, 275, 19297–19305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degirmenci, U.; Wang, M.; Hu, J. Targeting Aberrant RAS/RAF/MEK/ERK Signaling for Cancer Therapy. Cells 2020, 9, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logue, J.S.; Morrison, D.K. Complexity in the signaling network: Insights from the use of targeted inhibitors in cancer therapy. Genes Dev. 2012, 26, 641–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maik-Rachline, G.; Seger, R. The ERK cascade inhibitors: Towards overcoming resistance. Drug Resist. 2016, 25, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brereton, C.F.; Sutton, C.E.; Lalor, S.J.; Lavelle, E.C.; Mills, K.H.G. Inhibition of ERK MAPK suppresses IL-23 and IL-1-driven IL-17 production and attenuates autoimmune disease. J. Immunol. 2009, 183, 1715–1723. [Google Scholar] [CrossRef]
- Park, W.H. MAPK inhibitors, particularly the JNK inhibitor, increase cell death effects in H2O2-treated lung cancer cells via increased superoxide anion and glutathione depletion. Oncol. Rep. 2018, 39, 860–870. [Google Scholar] [CrossRef]
- Heumann, R.; Goemans, C.; Bartsch, D.; Lingenhöhl, K.; Waldmeier, P.C.; Hengerer, B.; Allegrini, P.R.; Schellander, K.; Wagner, E.F.; Arendt, T.; et al. Transgenic activation of Ras in neurons promotes hypertrophy and protects from lesion-induced degeneration. J. Cell Biol. 2000, 151, 1537–1548. [Google Scholar] [CrossRef]
- Birkner, K.; Wasser, B.; Loos, J.; Plotnikov, A.; Seger, R.; Zipp, F.; Witsch, E.; Bittner, S. The Role of ERK Signaling in Experimental Autoimmune Encephalomyelitis. Int. J. Mol. Sci. 2017, 18, 1990. [Google Scholar] [CrossRef] [Green Version]
- Bridel, C.; Lalive, P.H. Update on multiple sclerosis treatments. Swiss Med. Wkly. 2014, 144, w14012. [Google Scholar] [CrossRef] [PubMed]
- English, C.; Aloi, J.J. New FDA-Approved Disease-Modifying Therapies for Multiple Sclerosis. Clin. Ther. 2015, 37, 691–715. [Google Scholar] [CrossRef]
- Ten Bosch, G.J.A.; Bolk, J.; ‘t Hart, B.A.; Laman, J.D. Multiple sclerosis is linked to MAPKERKoveractivity in microglia. J. Mol. Med. 2021, 99, 1033–1042. [Google Scholar] [CrossRef]
- Kieseier, B.C.; Wiendl, H.; Hartung, H.P.; Stüve, O. The future of multiple sclerosis therapy. Pharmacol. Res. 2009, 60, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Gay, M.C.; Vrignaud, P.; Garitte, C.; Meunier, C. Predictors of depression in multiple sclerosis patients. Acta Neurol. Scand. 2010, 121, 161–170. [Google Scholar] [CrossRef] [PubMed]
- McGinley, M.P.; Goldschmidt, C.H.; Rae-Grant, A.D. Diagnosis and Treatment of Multiple Sclerosis: A Review. JAMA 2021, 325, 765–779. [Google Scholar] [CrossRef] [PubMed]
- Loraschi, A.; Bellantonio, P.; Bortolon, F.; Capra, R.; Cavalla, P.; Costantino, G.; Lugaresi, A.; Martinelli, V.; Marrosu, M.G.; Patti, F.; et al. Use of herbal remedies by multiple sclerosis patients: A nation-wide survey in Italy. Neurol. Sci. 2016, 37, 613–622. [Google Scholar] [CrossRef]
- Rashidi, B.; Malekzadeh, M.; Goodarzi, M.; Masoudifar, A.; Mirzaei, H. Green tea and its anti-angiogenesis effects. Biomed. Pharmacother. 2017, 89, 949–956. [Google Scholar] [CrossRef]
- Kaplan, M.; Mutlu, E.A.; Benson, M.; Fields, J.Z.; Banan, A.; Keshavarzian, A. Use of herbal preparations in the treatment of oxidant-mediated inflammatory disorders. Complement. Ther. Med. 2007, 15, 207–216. [Google Scholar] [CrossRef]
- Olsen, S.A. A review of complementary and alternative medicine (CAM) by people with multiple sclerosis. Occup. Ther. Int. 2009, 16, 57–70. [Google Scholar] [CrossRef]
- Song, L.; Zhou, Q.-H.; Wang, H.-L.; Liao, F.-J.; Hua, L.; Zhang, H.-F.; Huang, L.-B.; Lin, Y.; Zheng, G.-Q. Chinese herbal medicine adjunct therapy in patients with acute relapse of multiple sclerosis: A systematic review and meta-analysis. Complement. Ther. Med. 2017, 31, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chang, L.; Weinstock-Guttman, B.; Gandhi, S.; Jakimovski, D.; Carl, E.; Zivadinov, R.; Ramanathan, M. Complementary and alternative medicine usage by multiple sclerosis patients: Results from a prospective clinical study. J. Altern. Complement. Med. 2018, 24, 596–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayapoglu, N.; Tan, M. Use of complementary and alternative medicine among people with multiple sclerosis in eastern Turkey. Neurol. Asia 2016, 21, 63–71. [Google Scholar]
- Mojaverrostami, S.; Bojnordi, M.N.; Ghasemi-Kasman, M.; Ebrahimzadeh, M.A.; Hamidabadi, H.G. A Review of Herbal Therapy in Multiple Sclerosis. Adv. Pharm. Bull. 2018, 8, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Gentile, M.T.; Russo, R.; Pastorino, O.; Cioffi, S.; Barbieri, F.; Illingworth, E.A.; Grieco, M.; Chambery, A.; Colucci-D’Amato, L. Ruta graveolens water extract inhibits cell-cell network formation in human umbilical endothelial cells via MEK-ERK1/2 pathways. Exp. Cell Res. 2018, 364, 50–58. [Google Scholar] [CrossRef]
- Gentile, M.T.; Ciniglia, C.; Reccia, M.G.; Volpicelli, F.; Gatti, M.; Thellung, S.; Florio, T.; Melone, M.A.; Colucci-D’Amato, L. Ruta graveolens L. Induces Death of Glioblastoma Cells and Neural Progenitors, but Not of Neurons, via ERK 1/2 and AKT Activation. PLoS ONE 2015, 10, e0118864. [Google Scholar] [CrossRef] [PubMed]
- Colucci-D’Amato, L.; Cimaglia, G. Ruta graveolens as a potential source of neuroactive compounds to promote and restore neural functions. J. Tradit. Complement. Med. 2020, 10, 309–314. [Google Scholar] [CrossRef]
- Gentile, M.T.; Pastorino, O.; Bifulco, M.; Colucci-D’Amato, L. HUVEC Tube-formation Assay to Evaluate the Impact of Natural Products on Angiogenesis. J. Vis. Exp. 2019, 148, e58591. [Google Scholar] [CrossRef]
- Amado, N.G.; Predes, D.; Fonseca, B.F.; Cerqueira, D.M.; Reis, A.H.; Dudenhoeffer, A.C.; Borges, H.L.; Mendes, F.A.; Abreu, J.G. Isoquercitrin suppresses colon cancer cell growth in vitro by targeting the Wnt/β-catenin signaling pathway. J. Biol. Chem. 2014, 289, 35456–35467. [Google Scholar] [CrossRef] [Green Version]
- Ratheesh, M.; Shyni, G.L.; Sindhu, G.; Helen, A. Inhibitory effect of Ruta graveolens L. on oxidative damage, inflammation and aortic pathology in hypercholesteromic rats. Exp. Toxicol. Pathol. 2011, 63, 285–289. [Google Scholar] [CrossRef]
- Ip, F.C.F.; Ng, Y.P.; Or, T.C.T.; Sun, P.; Fu, G.; Li, J.Y.H.; Ye, W.C.; Cheung, T.H.; Ip, N.Y. Anemoside A3 ameliorates experimental autoimmune encephalomyelitis by modulating T helper 17 cell response. PLoS ONE 2017, 12, e0182069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajicek, J.; Fox, P.; Sanders, H.; Wright, D.; Vickery, J.; Nunn, A.; Thompson, A. Cannabinoids for treatment of spasticity and other symptoms related to multiple sclerosis (CAMS study): Multicentre randomised placebo-controlled trial. Lancet 2003, 362, 1517–1526. [Google Scholar] [CrossRef]
- Pertwee, R.G. Cannabinoids and multiple sclerosis. Pharmacol. Ther. 2002, 95, 165–174. [Google Scholar] [CrossRef]
- Mirossay, L.; Varinská, L.; Mojžiš, J. Antiangiogenic Effect of Flavonoids and Chalcones: An Update. Int. J. Mol. Sci. 2017, 19, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spagnuolo, C.; Moccia, S.; Russo, G.L. Anti-inflammatory effects of flavonoids in neurodegenerative disorders. Eur. J. Med. Chem. 2018, 153, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Millimouno, F.M.; Dong, J.; Yang, L.; Li, J.; Li, X. Targeting apoptosis pathways in cancer and perspectives with natural compounds from mother nature. Cancer Prev. Res. 2014, 7, 1081–1107. [Google Scholar] [CrossRef] [Green Version]
- Colucci-D’Amato, L.; Farina, A.; Vissers, J.P.C.; Chambery, A. Quantitative neuroproteomics: Classical and novel tools for studying neural differentiation and function. Stem Cell Rev. Rep. 2011, 7, 77–93. [Google Scholar] [CrossRef]
- Severino, V.; Farina, A.; Colucci-D’Amato, L.; Reccia, M.G.; Volpicelli, F.; Parente, A.; Chambery, A. Secretome profiling of differentiated neural mes-c-myc A1 cell line endowed with stem cell properties. Biochim. Biophys. Acta 2013, 1834, 2385–2395. [Google Scholar] [CrossRef]
- Colucci-D’Amato, L.; Cicatiello, A.E.; Reccia, M.G.; Volpicelli, F.; Severino, V.; Russo, R.; Sandomenico, A.; Doti, N.; D’Esposito, V.; Formisano, P.; et al. A targeted secretome profiling by multiplexed immunoassay revealed that secreted chemokine ligand 2 (MCP-1/CCL2) affects neural differentiation in mesencephalic neural progenitor cells. Proteomics 2015, 15, 714–724. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gentile, M.T.; Muto, G.; Lus, G.; Lövblad, K.-O.; Svenningsen, Å.F.; Colucci-D’Amato, L. Angiogenesis and Multiple Sclerosis Pathogenesis: A Glance at New Pharmaceutical Approaches. J. Clin. Med. 2022, 11, 4643. https://doi.org/10.3390/jcm11164643
Gentile MT, Muto G, Lus G, Lövblad K-O, Svenningsen ÅF, Colucci-D’Amato L. Angiogenesis and Multiple Sclerosis Pathogenesis: A Glance at New Pharmaceutical Approaches. Journal of Clinical Medicine. 2022; 11(16):4643. https://doi.org/10.3390/jcm11164643
Chicago/Turabian StyleGentile, Maria Teresa, Gianluca Muto, Giacomo Lus, Karl-Olof Lövblad, Åsa Fex Svenningsen, and Luca Colucci-D’Amato. 2022. "Angiogenesis and Multiple Sclerosis Pathogenesis: A Glance at New Pharmaceutical Approaches" Journal of Clinical Medicine 11, no. 16: 4643. https://doi.org/10.3390/jcm11164643