
Distinct Functional Cortico-Striato-Thalamo-Cerebellar Networks in Genetic Generalized and Focal Epilepsies with Generalized Tonic-Clonic Seizures

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Data Acquisition
2.3. Data Preprocessing
2.4. Parcellation Maps of Cortico-Striatum, -Thalamus and-Cerebellum Connectivity
2.5. Construction of Cortico-Striato-Thalamo-Cerebellar Networks
2.6. Statistical Analysis
2.6.1. Map Comparisons of Cortico-Striatum, Cortico-Thalamus, and Cortico-Cerebellar Connectivity
2.6.2. Comparisons of Cortico-Striato-Thalamo-Cerebellar Networks
2.6.3. Correlation with Clinical Variable of CORTICO-Striato-Thalamo-Cerebellar Networks
2.6.4. Classification of Patients
2.6.5. Reproducibility Analysis
3. Results
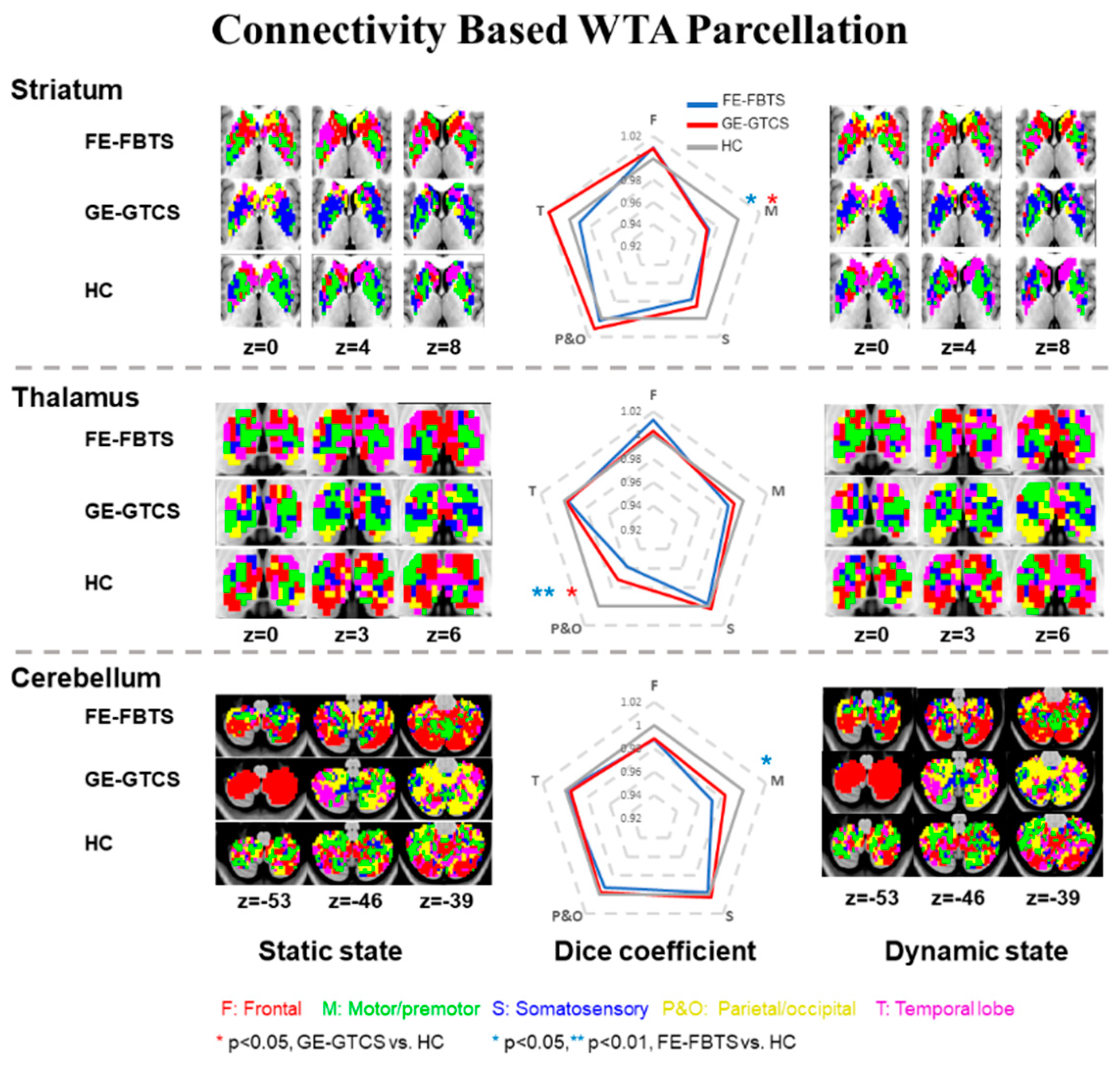
3.1. Maps of Cortico-Striatum, -Thalamus, and-Cerebellum Connectivity
3.2. Group Comparisons of Cortico-Striato-Thalamo-Cerebellar Connectivity
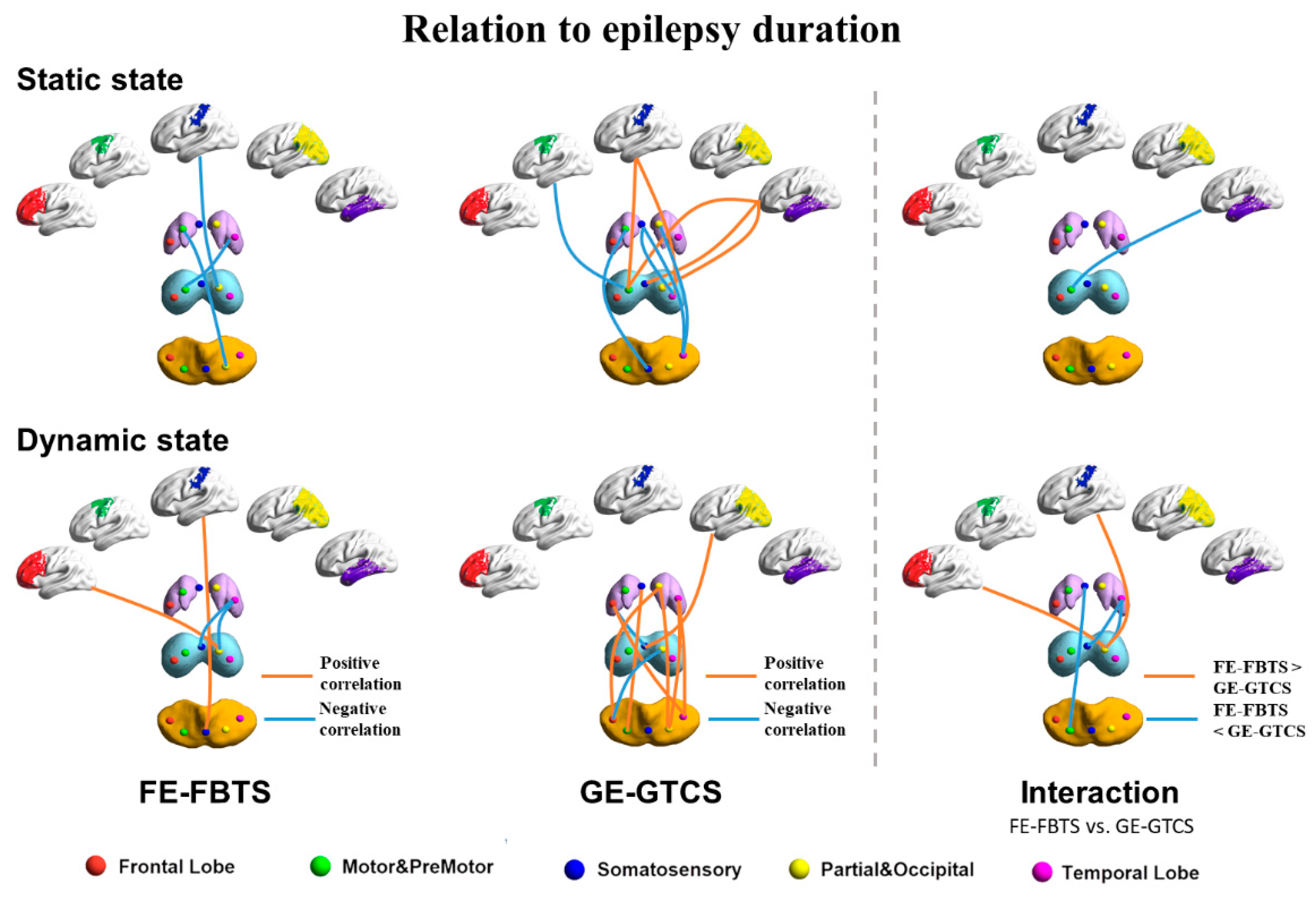
3.3. Relation to Disease Duration
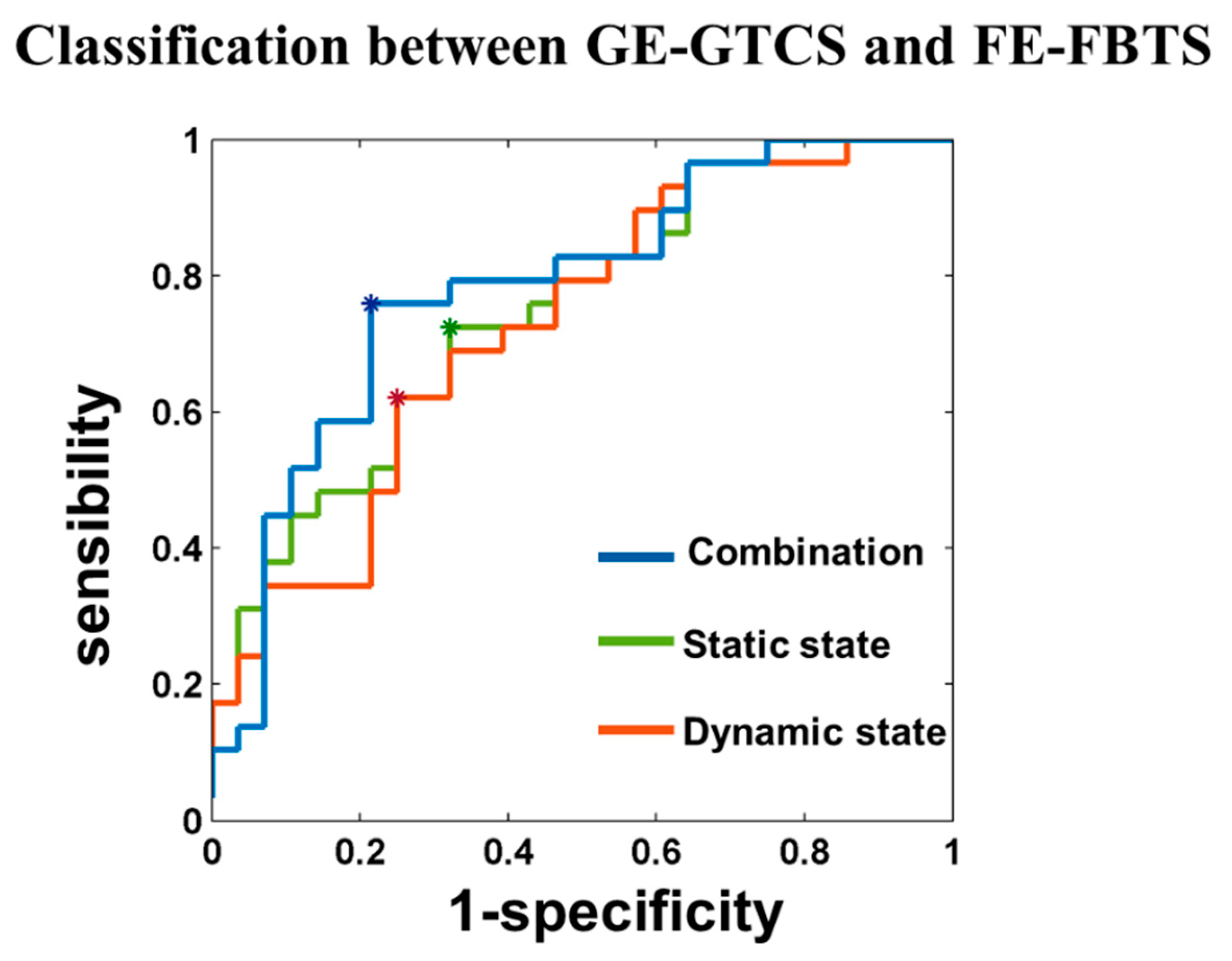
3.4. Classification
3.5. Reproducibility of Findings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blumenfeld, H.; Taylor, J. Why do seizures cause loss of consciousness? Neuroscientist 2003, 9, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.T.; Berkovic, S.F.; Brodie, M.J.; Buchhalter, J.; Cross, J.H.; van Emde Boas, W.; Engel, J.; French, J.; Glauser, T.A.; Mathern, G.W.; et al. Revised terminology and concepts for organization of seizures and epilepsies: Report of the ILAE Commission on Classification and Terminology, 2005–2009. Epilepsia 2010, 51, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Blumenfeld, H.; Varghese, G.I.; Purcaro, M.J.; Motelow, J.E.; Enev, M.; McNally, K.A.; Levin, A.R.; Hirsch, L.J.; Tikofsky, R.; Zubal, I.G.; et al. Cortical and subcortical networks in human secondarily generalized tonic-clonic seizures. Brain 2009, 132, 999–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegg, E.J.; Taylor, J.R.; Keller, S.S.; Mohanraj, R. Interictal structural and functional connectivity in idiopathic generalized epilepsy: A systematic review of graph theoretical studies. Epilepsy Behav. 2020, 106, 107013. [Google Scholar] [CrossRef] [PubMed]
- Shorvon, S.D.B.P.; Gibbs, A.A.; Huberfeld, G.; Kalviainen, R. Antiepileptic drug treatment of generalized tonic-clonic seizures: An evaluation of regulatory data and five criteria for drug selection. Epilepsy Behav. 2018, 82, 91–103. [Google Scholar] [CrossRef]
- Norden, A.D.; Blumenfeld, H. The role of subcortical structures in human epilepsy. Epilepsy Behav. 2002, 3, 219–231. [Google Scholar] [CrossRef]
- Luo, C.; Li, Q.; Xia, Y.; Lei, X.; Xue, K.; Yao, Z.; Lai, Y.; Martinez-Montes, E.; Liao, W.; Zhou, D.; et al. Resting state basal ganglia network in idiopathic generalized epilepsy. Hum. Brain Mapp. 2012, 33, 1279–1294. [Google Scholar] [CrossRef]
- He, X.; Chaitanya, G.; Asma, B.; Caciagli, L.; Bassett, D.S.; Tracy, J.I.; Sperling, M.R. Disrupted basal ganglia-thalamocortical loops in focal to bilateral tonic-clonic seizures. Brain 2020, 143, 175–190. [Google Scholar] [CrossRef]
- Jiang, S.; Li, X.; Li, Z.; Chang, X.; Chen, Y.; Huang, Y.; Zhang, Y.; Wang, H.; Zuo, X.; Li, X.; et al. Cerebello-cerebral connectivity in idiopathic generalized epilepsy. Eur. Radiol. 2020, 30, 3924–3933. [Google Scholar] [CrossRef]
- Moeller, F.; Siebner, H.R.; Wolff, S.; Muhle, H.; Boor, R.; Granert, O.; Jansen, O.; Stephani, U.; Siniatchkin, M. Changes in activity of striato-thalamo-cortical network precede generalized spike wave discharges. Neuroimage 2008, 39, 1839–1849. [Google Scholar] [CrossRef]
- Moeller, F.; LeVan, P.; Muhle, H.; Stephani, U.; Dubeau, F.; Siniatchkin, M.; Gotman, J. Absence seizures: Individual patterns revealed by EEG-fMRI. Epilepsia 2010, 51, 2000–2010. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.J.; Zhang, Z.; Xu, Q.; Wang, Z.; Wang, J.; Jiao, Q.; Yang, F.; Tan, Q.; Chen, G.; Zang, Y.F.; et al. Identifying Corticothalamic Network Epicenters in Patients with Idiopathic Generalized Epilepsy. AJNR Am. J. Neuroradiol. 2015, 36, 1494–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Lariviere, S.; Xu, Q.; Vos de Wael, R.; Hong, S.J.; Wang, Z.; Xu, Y.; Zhu, B.; Bernasconi, N.; Bernasconi, A.; et al. Community-informed connectomics of the thalamocortical system in generalized epilepsy. Neurology 2019, 93, e1112. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, B.C.; Rozen, D.A.; Worsley, K.J.; Evans, A.C.; Bernasconi, N.; Bernasconi, A. Thalamo-cortical network pathology in idiopathic generalized epilepsy: Insights from MRI-based morphometric correlation analysis. Neuroimage 2009, 46, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, I.E.; Berkovic, S.; Capovilla, G.; Connolly, M.B.; French, J.; Guilhoto, L.; Hirsch, E.; Jain, S.; Mathern, G.W.; Moshe, S.L.; et al. ILAE classification of the epilepsies: Position paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 512–521. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.G.; Zang, Y.F. DPARSF: A MATLAB Toolbox for “Pipeline” Data Analysis of Resting-State fMRI. Front. Syst. Neurosci. 2010, 4, 13. [Google Scholar] [CrossRef] [Green Version]
- Dongyang, Z.; Snyder, A.Z.; Fox, M.D.; Sansbury, M.W.; Shimony, J.S.; Raichle, M.E. Intrinsic functional relations between human cerebral cortex and thalamus. J. Neurophysiol. 2008, 100, 1740. [Google Scholar]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Liao, W.; Wu, G.R.; Xu, Q.; Ji, G.J.; Zhang, Z.; Zang, Y.F.; Lu, G. DynamicBC: A MATLAB toolbox for dynamic brain connectome analysis. Brain Connect. 2014, 4, 780–790. [Google Scholar] [CrossRef] [Green Version]
- Yao, G.; Li, J.; Wang, J.; Liu, S.; Li, X.; Cao, X.; Chen, H.; Xu, Y. Improved Resting-State Functional Dynamics in Post-stroke Depressive Patients After Shugan Jieyu Capsule Treatment. Front. Neurosci. 2020, 14, 297. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Li, J.; Duan, X.; Cui, Q.; Chen, H.; Chen, H. Static and dynamic connectomics differentiate between depressed patients with and without suicidal ideation. Hum. Brain Mapp. 2018, 39, 4105–4118. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, N.; Van De Ville, D. On spurious and real fluctuations of dynamic functional connectivity during rest. Neuroimage 2015, 104, 430–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Li, W.; Zhao, M.; Wu, J.; Wu, J.; Yang, J.; Jiao, B. Altered temporal dynamics of brain activity in patients with generalized tonic-clonic seizures. PLoS ONE 2019, 14, e0219904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Q.; Sheng, W.; Chen, Y.; Pang, Y.; Lu, F.; Tang, Q.; Han, S.; Shen, Q.; Wang, Y.; Xie, A.; et al. Dynamic changes of amplitude of low-frequency fluctuations in patients with generalized anxiety disorder. Hum. Brain Mapp. 2020, 41, 1667–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worsley, K.J.; Taylor, J.; Carbonell, F.; Chung, M.K.; Duerden, E.; Bernhardt, B.; Lyttelton, O.; Boucher, M.; Evans, A.C. SurfStat: A Matlab toolbox for the statistical analysis of univariate and multivariate surface and volumetric data using linear mixed effects models and random field theory. NeuroImage 2009, 47, S102. [Google Scholar] [CrossRef]
- Rakic, M.; Cabezas, M.; Kushibar, K.; Oliver, A.; Llado, X. Improving the detection of autism spectrum disorder by combining structural and functional MRI information. Neuroimage Clin. 2020, 25, 102181. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yuan, F.; Zhou, G.; Nie, J.; Wang, D.; Hu, P.; Ouyang, L.; Kong, L.; Liao, W. Cross-network interaction for diagnosis of major depressive disorder based on resting state functional connectivity. Brain Imaging Behav. 2021, 15, 1279–1289. [Google Scholar] [CrossRef]
- Joo, E.Y.; Tae, W.S.; Hong, S.B. Cerebral blood flow abnormality in patients with idiopathic generalized epilepsy. J. Neurol. 2008, 255, 520–525. [Google Scholar] [CrossRef]
- Vytvarova, E.; Marecek, R.; Fousek, J.; Strycek, O.; Rektor, I. Large-scale cortico-subcortical functional networks in focal epilepsies: The role of the basal ganglia. Neuroimage Clin. 2017, 14, 28–36. [Google Scholar] [CrossRef]
- Zhang, C.H.; Sha, Z.; Mundahl, J.; Liu, S.; Lu, Y.; Henry, T.R.; He, B. Thalamocortical relationship in epileptic patients with generalized spike and wave discharges—A multimodal neuroimaging study. Neuroimage Clin. 2015, 9, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Mahoney, E.C.; Zeng, A.; Yu, W.; Rowe, M.; Sahai, S.; Feustel, P.J.; Ramirez-Zamora, A.; Pilitsis, J.G.; Shin, D.S. Ventral pallidum deep brain stimulation attenuates acute partial, generalized and tonic-clonic seizures in two rat models. Epilepsy Res. 2018, 142, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Kros, L.; Eelkman Rooda, O.H.; Spanke, J.K.; Alva, P.; van Dongen, M.N.; Karapatis, A.; Tolner, E.A.; Strydis, C.; Davey, N.; Winkelman, B.H.; et al. Cerebellar output controls generalized spike-and-wave discharge occurrence. Ann. Neurol. 2015, 77, 1027–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoodley, C.J.; Valera, E.M.; Schmahmann, J.D. Functional topography of the cerebellum for motor and cognitive tasks: An fMRI study. Neuroimage 2012, 59, 1560–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krook-Magnuson, E.; Szabo, G.G.; Armstrong, C.; Oijala, M.; Soltesz, I. Cerebellar Directed Optogenetic Intervention Inhibits Spontaneous Hippocampal Seizures in a Mouse Model of Temporal Lobe Epilepsy. eNeuro 2014, 1, ENEURO.0005-14.2014. [Google Scholar] [CrossRef] [PubMed]
- Cooper, I.S. Effect of chronic stimulation of anterior cerebellum on neurological disease. Lancet 1973, 1, 206. [Google Scholar] [CrossRef]
- Ferilli, M.A.N.; Brunetti, V.; Costantini, E.M.; Della Marca, G. Left hemispheric status epilepticus with crossed cerebellar diaschisis. J. Neurol. Neurosurg. Psychiatry 2018, 89, 311–312. [Google Scholar] [CrossRef] [PubMed]
- Danielson, N.B.; Guo, J.N.; Blumenfeld, H. The default mode network and altered consciousness in epilepsy. Behav. Neurol. 2011, 24, 55–65. [Google Scholar] [CrossRef]
- Szabo, C.A.; Lancaster, J.L.; Lee, S.; Xiong, J.H.; Cook, C.; Mayes, B.N.; Fox, P.T. MR imaging volumetry of subcortical structures and cerebellar hemispheres in temporal lobe epilepsy. AJNR Am. J. Neuroradiol. 2006, 27, 2155–2160. [Google Scholar]
- Zhu, L.; Li, Y.; Wang, Y.; Li, R.; Zhang, Z.; Lu, G.; Chen, H. Aberrant long-range functional connectivity density in generalized tonic-clonic seizures. Medicine 2016, 95, e3893. [Google Scholar] [CrossRef]
- Pawley, A.D.; Chowdhury, F.A.; Tangwiriyasakul, C.; Ceronie, B.; Elwes, R.D.; Nashef, L.; Richardson, M.P. Cortical excitability correlates with seizure control and epilepsy duration in chronic epilepsy. Ann. Clin. Transl. Neurol. 2017, 4, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.R. The behavioral and cognitive relevance of time-varying, dynamic changes in functional connectivity. Neuroimage 2018, 180, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Liegeois, R.; Li, J.; Kong, R.; Orban, C.; Van De Ville, D.; Ge, T.; Sabuncu, M.R.; Yeo, B.T.T. Resting brain dynamics at different timescales capture distinct aspects of human behavior. Nat. Commun. 2019, 10, 2317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calhoun, V.D.; Miller, R.; Pearlson, G.; Adali, T. The chronnectome: Time-varying connectivity networks as the next frontier in fMRI data discovery. Neuron 2014, 84, 262–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, B.; Arbabshirani, M.R.; Damaraju, E.; Cetin, M.S.; Miller, R.; Pearlson, G.D.; Calhoun, V.D. Classification of schizophrenia and bipolar patients using static and dynamic resting-state fMRI brain connectivity. Neuroimage 2016, 134, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liao, W.; Xu, Q.; Wei, W.; Zhou, H.J.; Sun, K.; Yang, F.; Mantini, D.; Ji, X.; Lu, G. Hippocampus-associated causal network of structural covariance measuring structural damage progression in temporal lobe epilepsy. Hum. Brain Mapp. 2017, 38, 753–766. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Mean ± SD) Years | Gender Male/Female | Durations (Mean ± SD) Years | Seizure Frequency (per Month) | |
|---|---|---|---|---|
| GE-GTCS | 26.44 ± 8.74 | 75/39 | 7.98 ± 8.32 | 2.212 ± 10.76 |
| FE-FBTS | 26.27 ± 7.78 | 75/39 | 9.42 ± 6.56 | 2.34 ± 6.21 |
| HC | 25.07 ± 5.99 | 75/39 | - | - |
| Statistical Value | F = 0.957 | Chi-square < 0.001 | t = 0.187 | t = 0.105 |
| p Value | 0.385 a | 1 b | 0.574 c | 0.916 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, H.; Xu, Q.; Yang, F.; Zhang, Q.; Hao, J.; Liu, G.; Liu, R.; Yu, Q.; Zhang, Z.; Xing, W.; et al. Distinct Functional Cortico-Striato-Thalamo-Cerebellar Networks in Genetic Generalized and Focal Epilepsies with Generalized Tonic-Clonic Seizures. J. Clin. Med. 2022, 11, 1612. https://doi.org/10.3390/jcm11061612
Hsieh H, Xu Q, Yang F, Zhang Q, Hao J, Liu G, Liu R, Yu Q, Zhang Z, Xing W, et al. Distinct Functional Cortico-Striato-Thalamo-Cerebellar Networks in Genetic Generalized and Focal Epilepsies with Generalized Tonic-Clonic Seizures. Journal of Clinical Medicine. 2022; 11(6):1612. https://doi.org/10.3390/jcm11061612
Chicago/Turabian StyleHsieh, Hsinyu, Qiang Xu, Fang Yang, Qirui Zhang, Jingru Hao, Gaoping Liu, Ruoting Liu, Qianqian Yu, Zixuan Zhang, Wei Xing, and et al. 2022. "Distinct Functional Cortico-Striato-Thalamo-Cerebellar Networks in Genetic Generalized and Focal Epilepsies with Generalized Tonic-Clonic Seizures" Journal of Clinical Medicine 11, no. 6: 1612. https://doi.org/10.3390/jcm11061612