
Severe Traumatic Injury Induces Phenotypic and Functional Changes of Neutrophils and Monocytes

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Patient Cohort
2.3. Staining of Neutrophil and Monocyte Subsets for Flow Cytometry
2.4. Analysis of Bacterial Intake by Neutrophils by Flow Cytometry
2.5. Analysis of Reactive Oxygen Species Production in Neutrophils and Monocytes by Flow Cytometry
2.6. Fluorescence-Activated Cell Sorting of Neutrophil and Monocyte Subsets
2.7. Statistics
3. Results
3.1. Patient Cohort
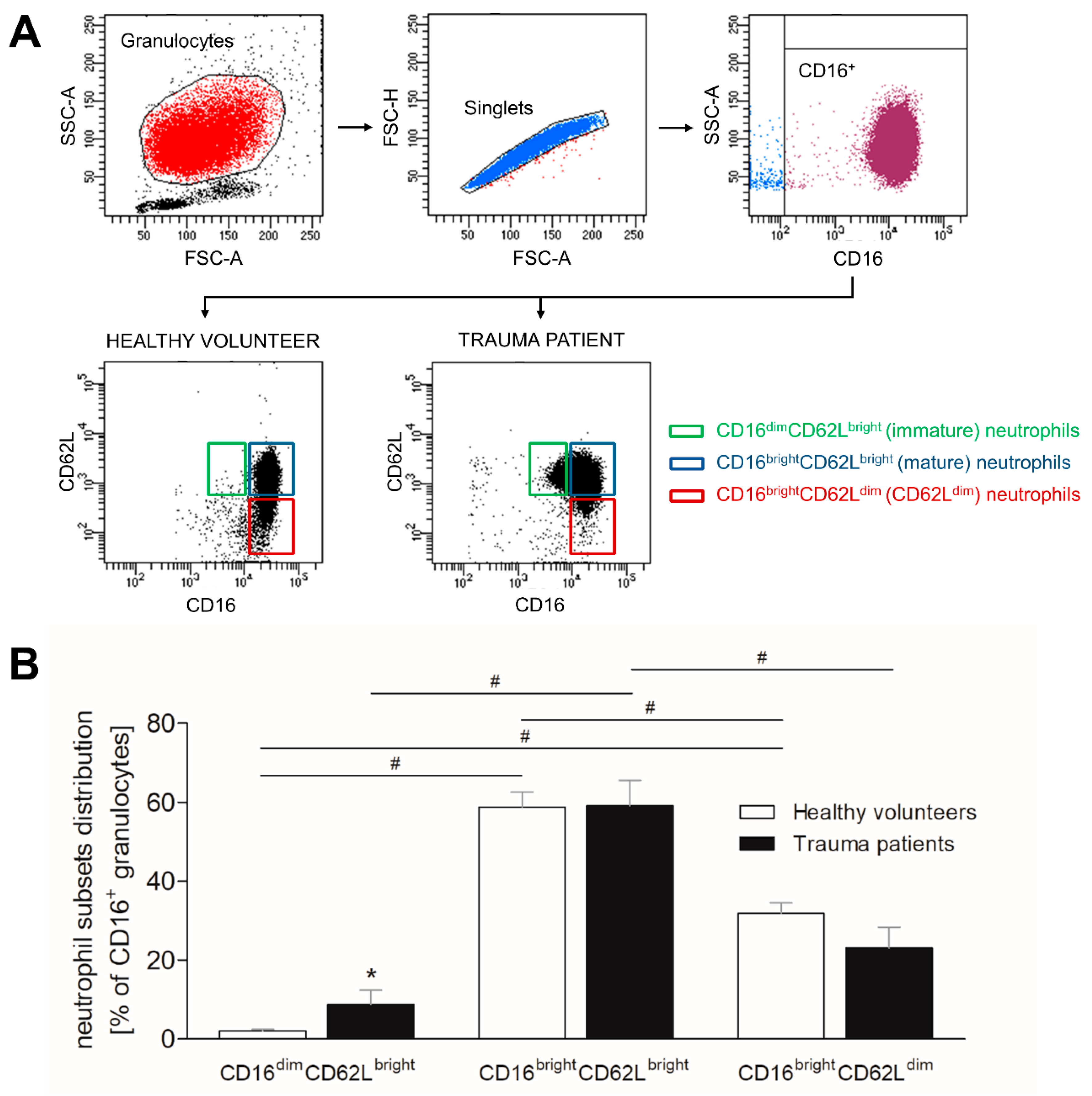
3.2. Severe Trauma Modulates the Distribution of CD16+ Neutrophil Subsets
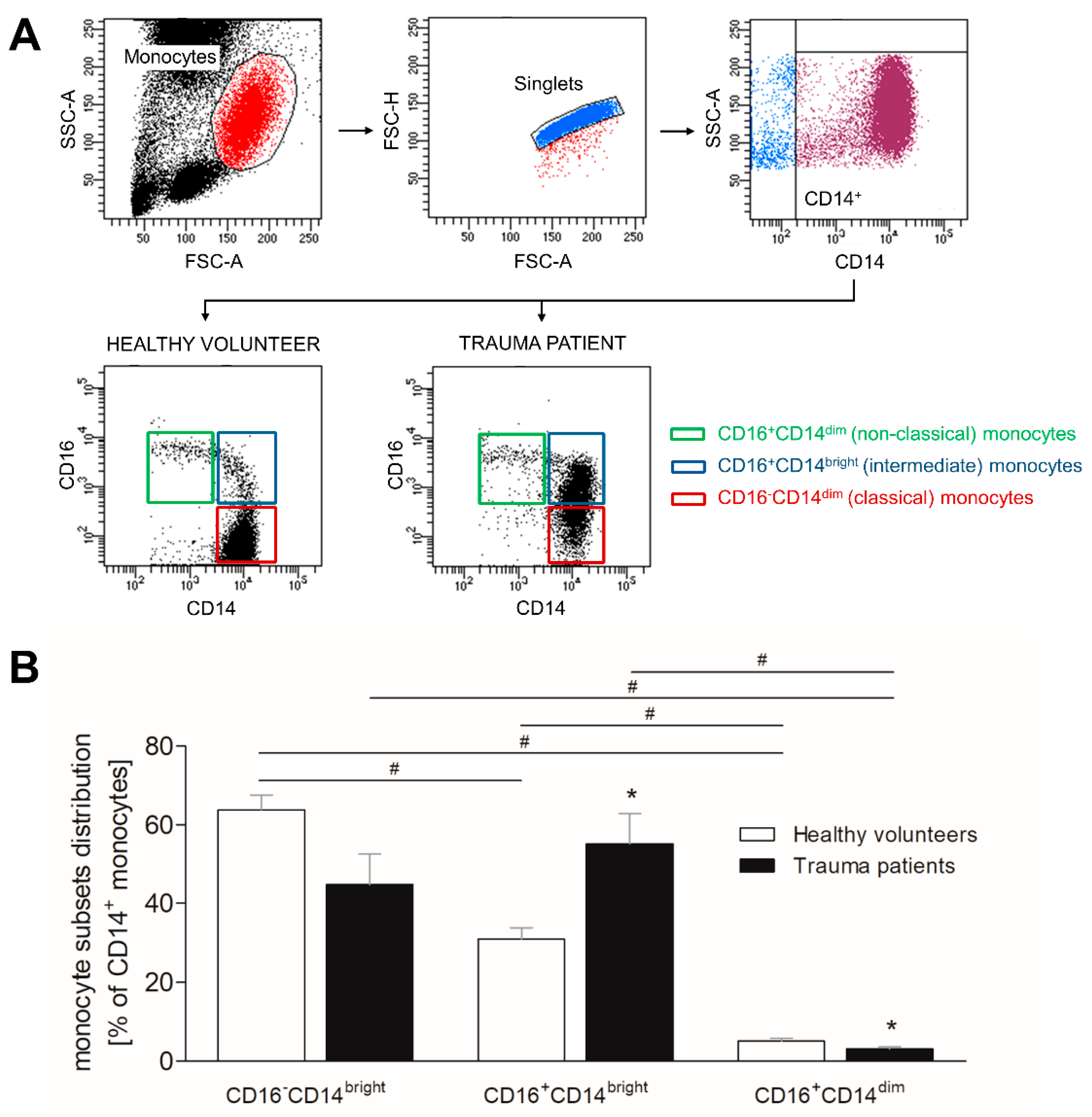
3.3. Severe Traumatic Injury Causes a Phenotypic Shift of CD14+ Monocytes
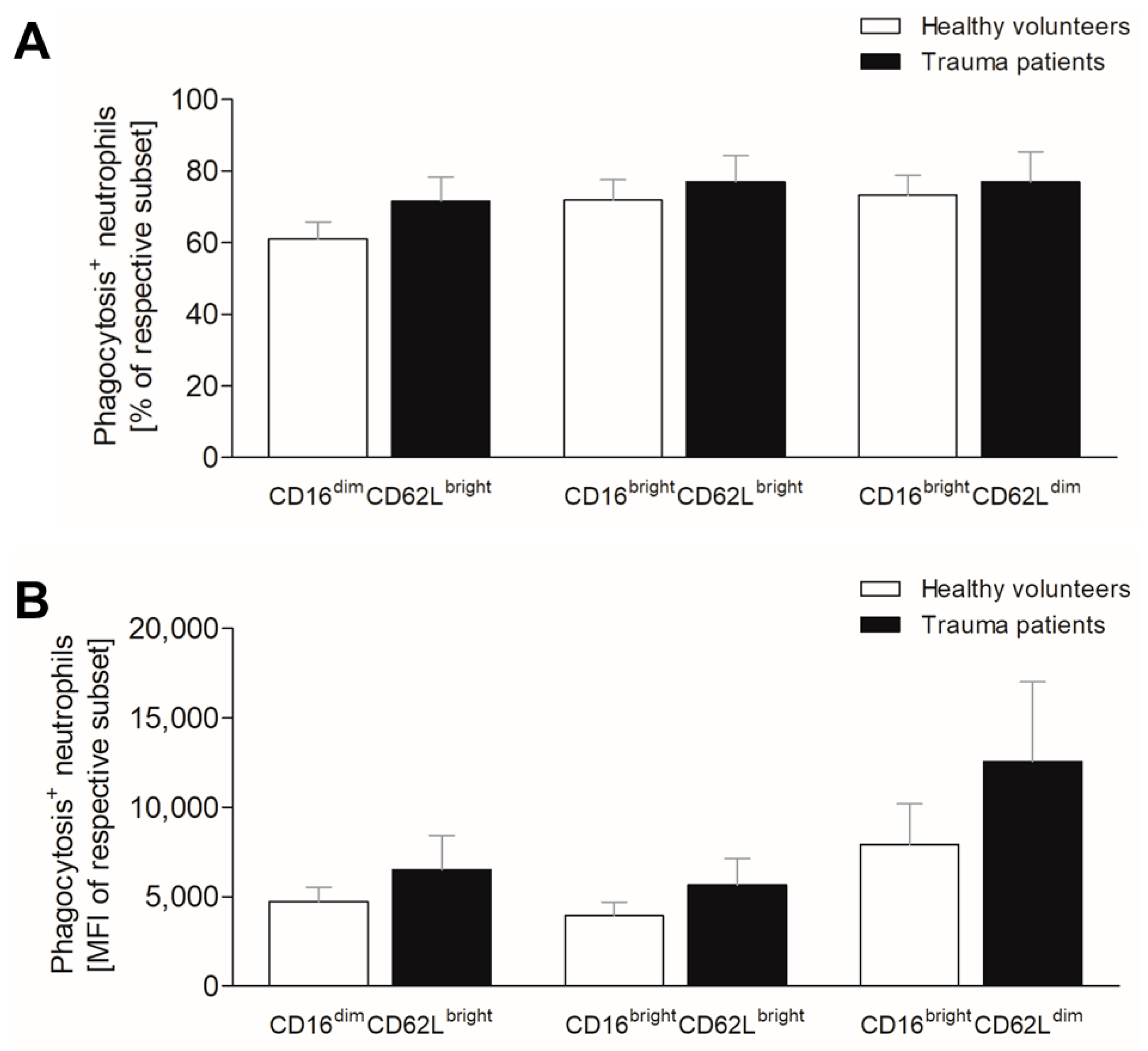
3.4. Severe Trauma Does Not Affect Phagocytic Capacity of Neutrophils at Early Time Point
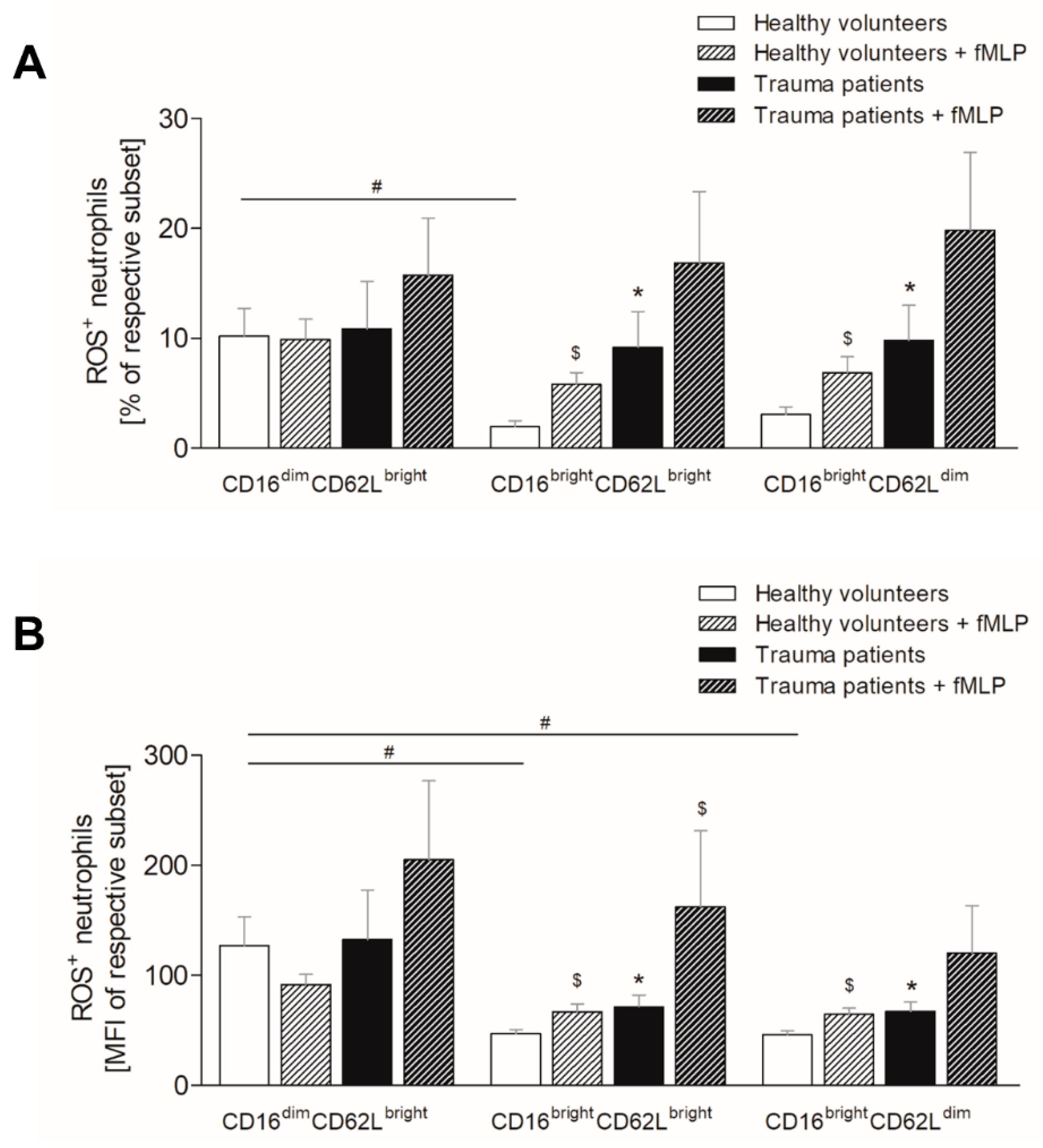
3.5. Severe Trauma Elevates the Production of Reactive Oxygen Species in Mature and CD62Ldim Neutrophils
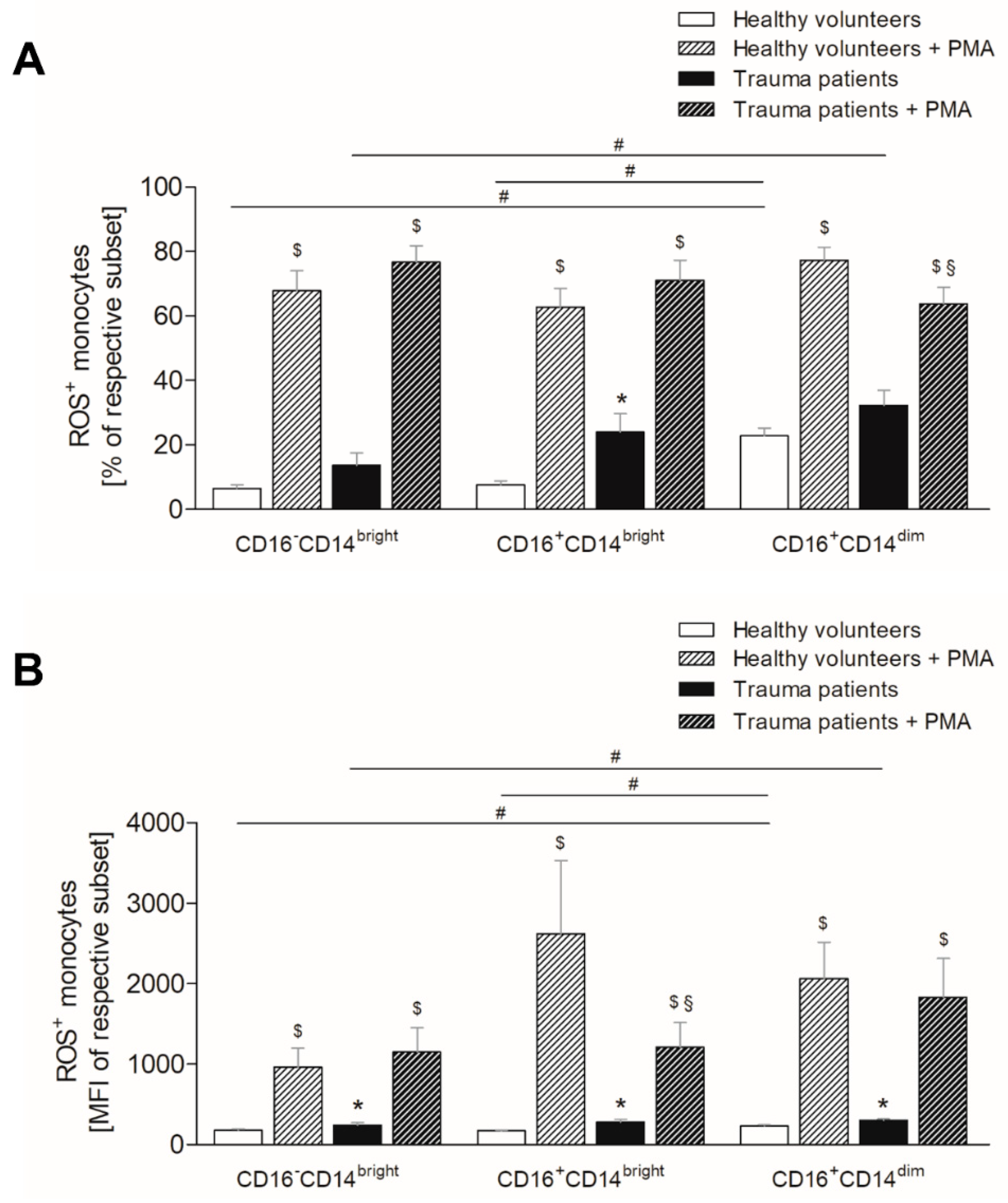
3.6. Severe Traumatic Injury Increases the Production of Reactive Oxygen Species in Monocytes
3.7. Fluorescence-Activated Cell Sorting Reveals an Exhaustion of Neutrophils and Monocytes and Aligns the Functional Differences between the Subsets
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ARDS | acute respiratory distress syndrome |
| CD | cluster of differentiation |
| FACS | fluorescence-activated cell sorting |
| fMLP | N-formyl-methionyl-leucyl-phenylalanine |
| IL | interleukin |
| ISS | injury severity score |
| MOF | multi organ failure |
| PBS | phosphate-buffered saline |
| PBMCs | peripheral blood mononuclear cells |
| PMA | phorbol 12-myristate 13-acetate |
| ROS | reactive oxygen species |
References
- Injuries and Violence: The Facts; World Health Organization: Geneve, Switzerland, 2010.
- Hellebrekers, P.; Leenen, L.P.; Hoekstra, M.; Hietbrink, F. Effect of a standardized treatment regime for infection after osteosynthesis. J. Orthop. Surg. Res. 2017, 12, 41. [Google Scholar] [CrossRef] [Green Version]
- Horiguchi, H.; Loftus, T.J.; Hawkins, R.B.; Raymond, S.L.; Stortz, J.A.; Hollen, M.K.; Weiss, B.P.; Miller, E.S.; Bihorac, A.; Larson, S.D.; et al. Efron, Sepsis, and I. Critical Illness Research Center. Innate Immunity in the Persistent Inflammation, Immunosuppression, and Catabolism Syndrome and Its Implications for Therapy. Front. Immunol. 2018, 9, 595. [Google Scholar] [CrossRef]
- Mira, J.C.; Brakenridge, S.C.; Moldawer, L.L.; Moore, F.A. Persistent Inflammation, Immunosuppression and Catabolism Syndrome. Crit. Care Clin. 2017, 33, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Stormann, P.; Lustenberger, T.; Relja, B.; Marzi, I.; Wutzler, S. Role of biomarkers in acute traumatic lung injury. Injury 2017, 48, 2400–2406. [Google Scholar] [CrossRef] [PubMed]
- Villar, J.; Blanco, J.; Kacmarek, R.M. Current incidence and outcome of the acute respiratory distress syndrome. Curr. Opin. Crit. Care 2016, 22, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kellner, M.; Noonepalle, S.; Lu, Q.; Srivastava, A.; Zemskov, E.; Black, S.M. ROS Signaling in the Pathogenesis of Acute Lung Injury (ALI) and Acute Respiratory Distress Syndrome (ARDS). Adv. Exp. Med. Biol. 2017, 967, 105–137. [Google Scholar] [PubMed]
- Hesselink, L.; Spijkerman, R.; van Wessem, K.J.P.; Koenderman, L.; Leenen, L.P.H.; Huber-Lang, M.; Hietbrink, F. Neutrophil heterogeneity and its role in infectious complications after severe trauma. World J. Emerg. Surg. 2019, 14, 24. [Google Scholar] [CrossRef]
- Tak, T.; van Groenendael, R.; Pickkers, P.; Koenderman, L. Monocyte Subsets Are Differentially Lost from the Circulation during Acute Inflammation Induced by Human Experimental Endotoxemia. J. Innate. Immun. 2017, 9, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Westman, J.; Grinstein, S.; Marques, P.E. Phagocytosis of Necrotic Debris at Sites of Injury and Inflammation. Front. Immunol. 2019, 10, 3030. [Google Scholar] [CrossRef]
- Rani, M.; Nicholson, S.E.; Zhang, Q.; Schwacha, M.G. Damage-associated molecular patterns (DAMPs) released after burn are associated with inflammation and monocyte activation. Burns 2017, 43, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil function: From mechanisms to disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef]
- Filep, J.G.; Ariel, A. Neutrophil heterogeneity and fate in inflamed tissues: Implications for the resolution of inflammation. Am. J. Physiol. Cell Physiol. 2020, 319, C510–C532. [Google Scholar] [CrossRef]
- Andrews, T.; Sullivan, K.E. Infections in patients with inherited defects in phagocytic function. Clin. Microbiol. Rev. 2003, 16, 597–621. [Google Scholar] [CrossRef] [Green Version]
- Reine, J.; Rylance, J.; Ferreira, D.M.; Pennington, S.H.; Welters, I.D.; Parker, R.; Morton, B. The whole blood phagocytosis assay: A clinically relevant test of neutrophil function and dysfunction in community-acquired pneumonia. BMC Res. Notes 2020, 13, 203. [Google Scholar] [CrossRef] [Green Version]
- Kanyilmaz, S.; Hepguler, S.; Atamaz, F.C.; Gokmen, N.M.; Ardeniz, O.; Sin, A. Phagocytic and oxidative burst activity of neutrophils in patients with spinal cord injury. Arch. Phys. Med. Rehabil. 2013, 94, 369–374. [Google Scholar] [CrossRef]
- Liao, Y.; Liu, P.; Guo, F.; Zhang, Z.Y.; Zhang, Z. Oxidative burst of circulating neutrophils following traumatic brain injury in human. PLoS ONE 2013, 8, e68963. [Google Scholar]
- Ritzel, R.M.; Doran, S.J.; Barrett, J.P.; Henry, R.J.; Ma, E.L.; Faden, A.I.; Loane, D.J. Chronic Alterations in Systemic Immune Function after Traumatic Brain Injury. J. Neurotrauma 2018, 35, 1419–1436. [Google Scholar] [CrossRef]
- Hellebrekers, P.; Hietbrink, F.; Vrisekoop, N.; Leenen, L.P.H.; Koenderman, L. Neutrophil Functional Heterogeneity: Identification of Competitive Phagocytosis. Front. Immunol. 2017, 8, 1498. [Google Scholar] [CrossRef]
- Santos, S.S.; Brunialti, M.K.; Rigato, O.; Machado, F.R.; Silva, E.; Salomao, R. Generation of nitric oxide and reactive oxygen species by neutrophils and monocytes from septic patients and association with outcomes. Shock 2012, 38, 18–23. [Google Scholar] [CrossRef]
- Mulder, P.P.G.; Vlig, M.; Boekema, B.; Stoop, M.M.; Pijpe, A.; van Zuijlen, P.P.M.; de Jong, E.; van Cranenbroek, B.; Joosten, I.; Koenen, H. Persistent Systemic Inflammation in Patients With Severe Burn Injury Is Accompanied by Influx of Immature Neutrophils and Shifts in T Cell Subsets and Cytokine Profiles. Front. Immunol. 2020, 11, 621222. [Google Scholar] [CrossRef]
- Pillay, J.; Kamp, V.M.; van Hoffen, E.; Visser, T.; Tak, T.; Lammers, J.W.; Ulfman, L.H.; Leenen, L.P.; Pickkers, P.; Koenderman, L. A subset of neutrophils in human systemic inflammation inhibits T cell responses through Mac-1. J. Clin. Investig. 2012, 122, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Mazzei, A.; Palumbo, C.; Verri, T.; Lobreglio, G. Flow Cytometric Analysis of Monocytes Polarization and Reprogramming From Inflammatory to Immunosuppressive Phase During Sepsis. EJIFCC 2019, 30, 371–384. [Google Scholar] [PubMed]
- Kapellos, T.S.; Bonaguro, L.; Gemund, I.; Reusch, N.; Saglam, A.; Hinkley, E.R.; Schultze, J.L. Human Monocyte Subsets and Phenotypes in Major Chronic Inflammatory Diseases. Front. Immunol. 2019, 10, 2035. [Google Scholar] [CrossRef] [Green Version]
- Cros, J.; Cagnard, N.; Woollard, K.; Patey, N.; Zhang, S.Y.; Senechal, B.; Puel, A.; Biswas, S.K.; Moshous, D.; Picard, C.; et al. Human CD14dim monocytes patrol and sense nucleic acids and viruses via TLR7 and TLR8 receptors. Immunity 2010, 33, 375–386. [Google Scholar] [CrossRef] [Green Version]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gotzsche, P.C.; Vandenbroucke, J.P.; Initiative, S. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. J. Clin. Epidemiol. 2008, 61, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, S.M.; Corriden, R.; Nizet, V. The Ontogeny of a Neutrophil: Mechanisms of Granulopoiesis and Homeostasis. Microbiol. Mol. Biol. Rev. 2018, 82, e00057-17. [Google Scholar] [CrossRef] [Green Version]
- Spijkerman, R.; Hesselink, L.; Bongers, S.; van Wessem, K.J.P.; Vrisekoop, N.; Hietbrink, F.; Koenderman, L.; Leenen, L.P.H. Point-of-Care Analysis of Neutrophil Phenotypes: A First Step Toward Immuno-Based Precision Medicine in the Trauma ICU. Crit. Care Explor. 2020, 2, e0158. [Google Scholar] [CrossRef]
- Kovtun, A.; Messerer, D.A.C.; Scharffetter-Kochanek, K.; Huber-Lang, M.; Ignatius, A. Neutrophils in Tissue Trauma of the Skin, Bone, and Lung: Two Sides of the Same Coin. J. Immunol. Res. 2018, 2018, 8173983. [Google Scholar] [CrossRef]
- Hazeldine, J.; Naumann, D.N.; Toman, E.; Davies, D.; Bishop, J.R.B.; Su, Z.; Hampson, P.; Dinsdale, R.J.; Crombie, N.; Duggal, N.A.; et al. Prehospital immune responses and development of multiple organ dysfunction syndrome following traumatic injury: A prospective cohort study. PLoS Med. 2017, 14, e1002338. [Google Scholar] [CrossRef]
- Cappenberg, A.; Margraf, A.; Thomas, K.; Bardel, B.; McCreedy, D.A.; van Marck, V.; Mellmann, A.; Lowell, C.A.; Zarbock, A. L-selectin shedding affects bacterial clearance in the lung: A new regulatory pathway for integrin outside-in signaling. Blood 2019, 134, 1445–1457. [Google Scholar] [CrossRef]
- Lee, D.; Schultz, J.B.; Knauf, P.A.; King, R.M. Mechanical shedding of L-selectin from the neutrophil surface during rolling on sialyl Lewis x under flow. J. Biol. Chem. 2007, 282, 4812–4820. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, M.J.; Lin, K.S.; King, M.R. Fluid shear stress increases neutrophil activation via platelet-activating factor. Biophys. J. 2014, 106, 2243–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.; Chen, S.B.; Li, L.D.; Tong, C.F.; Li, N.; Lu, S.Q.; Long, M. Impact of real-time shedding on binding kinetics of membrane-remaining L-selectin to PSGL-1. Am. J. Physiol. Cell Physiol. 2019, 316, C678–C689. [Google Scholar] [CrossRef] [PubMed]
- Pap, G.; Furesz, J.; Fennt, J.; Kovacs, G.C.; Nagy, L.; Hamar, J. Self-regulation of neutrophils during phagocytosis is modified after severe tissue injury. Int. J. Mol. Med. 2006, 17, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Sturm, R.; Heftrig, D.; Mors, K.; Wagner, N.; Kontradowitz, K.; Jurida, K.; Marzi, I.; Relja, B. Phagocytizing activity of PMN from severe trauma patients in different post-traumatic phases during the 10-days post-injury course. Immunobiology 2017, 222, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Leliefeld, P.H.C.; Pillay, J.; Vrisekoop, N.; Heeres, M.; Tak, T.; Kox, M.; Rooijakkers, S.H.M.; Kuijpers, T.W.; Pickkers, P.; Leenen, L.P.H.; et al. Differential antibacterial control by neutrophil subsets. Blood Adv. 2018, 2, 1344–1355. [Google Scholar] [CrossRef] [Green Version]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front. Immunol. 2018, 9, 2171. [Google Scholar] [CrossRef] [Green Version]
- Partrick, D.A.; Moore, F.A.; Moore, E.E.; Barnett, C.C., Jr.; Silliman, C.C. Neutrophil priming and activation in the pathogenesis of postinjury multiple organ failure. New Horiz. 1996, 4, 194–210. [Google Scholar]
- Swain, S.D.; Rohn, T.T.; Quinn, M.T. Neutrophil priming in host defense: Role of oxidants as priming agents. Antioxid. Redox Signal. 2002, 4, 69–83. [Google Scholar] [CrossRef]
- Wang, L.; Ai, Z.; Khoyratty, T.; Zec, K.; Eames, H.L.; van Grinsven, E.; Hudak, A.; Morris, S.; Ahern, D.; Monaco, C.; et al. ROS-producing immature neutrophils in giant cell arteritis are linked to vascular pathologies. JCI Insight 2020, 5, e139163. [Google Scholar] [CrossRef]
- Botha, A.J.; Moore, F.A.; Moore, E.E.; Kim, F.J.; Banerjee, A.; Peterson, V.M. Postinjury neutrophil priming and activation: An early vulnerable window. Surgery 1995, 118, 358–364, discussion 364–365. [Google Scholar] [CrossRef]
- Mortaz, E.; Zadian, S.S.; Shahir, M.; Folkerts, G.; Garssen, J.; Mumby, S.; Adcock, I.M. Does Neutrophil Phenotype Predict the Survival of Trauma Patients? Front. Immunol. 2019, 10, 2122. [Google Scholar] [CrossRef]
- Zhu, L.; Castranova, V.; He, P. fMLP-stimulated neutrophils increase endothelial [Ca2+]i and microvessel permeability in the absence of adhesion: Role of reactive oxygen species. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H1331–H1338. [Google Scholar] [CrossRef] [Green Version]
- Olingy, C.E.; Emeterio, C.L.S.; Ogle, M.E.; Krieger, J.R.; Bruce, A.C.; Pfau, D.D.; Jordan, B.T.; Peirce, S.M.; Botchwey, E.A. Non-classical monocytes are biased progenitors of wound healing macrophages during soft tissue injury. Sci. Rep. 2017, 7, 447. [Google Scholar] [CrossRef] [Green Version]
- Flohe, S.; Flohe, S.B.; Schade, F.U.; Waydhas, C. Immune response of severely injured patients--influence of surgical intervention and therapeutic impact. Langenbecks Arch. Surg. 2007, 392, 639–648. [Google Scholar] [CrossRef]
- Kirchhoff, C.; Biberthaler, P.; Mutschler, W.E.; Faist, E.; Jochum, M.; Zedler, S. Early down-regulation of the pro-inflammatory potential of monocytes is correlated to organ dysfunction in patients after severe multiple injury: A cohort study. Crit. Care 2009, 13, R88. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.A.; Zhang, Y.; Fullerton, J.N.; Boelen, L.; Rongvaux, A.; Maini, A.A.; Bigley, V.; Flavell, R.A.; Gilroy, D.W.; Asquith, B.; et al. The fate and lifespan of human monocyte subsets in steady state and systemic inflammation. J. Exp. Med. 2017, 214, 1913–1923. [Google Scholar] [CrossRef]
- West, S.D.; Goldberg, D.; Ziegler, A.; Krencicki, M.; Clos, T.W.D.; Mold, C. Transforming growth factor-beta, macrophage colony-stimulating factor and C-reactive protein levels correlate with CD14(high)CD16+ monocyte induction and activation in trauma patients. PLoS ONE 2012, 7, e52406. [Google Scholar] [CrossRef]
- Stansfield, B.K.; Ingram, D.A. Clinical significance of monocyte heterogeneity. Clin. Transl. Med. 2015, 4, 5. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Immunosuppression in sepsis: A novel understanding of the disorder and a new therapeutic approach. Lancet Infect. Dis. 2013, 13, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Bao, F.; Bailey, C.S.; Gurr, K.R.; Bailey, S.I.; Rosas-Arellano, M.P.; Dekaban, G.A.; Weaver, L.C. Increased oxidative activity in human blood neutrophils and monocytes after spinal cord injury. Exp. Neurol. 2009, 215, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, A.; Brat, G.A.; Yorkgitis, B.K.; Keegan, J.; Dolan, J.; Salim, A.; Askari, R.; Lederer, J.A. Phenotyping the Immune Response to Trauma: A Multiparametric Systems Immunology Approach. Crit. Care Med. 2017, 45, 1523–1530. [Google Scholar] [CrossRef]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | SEM | |
|---|---|---|
| Age | 38.2 years old | 4.99 |
| Gender | 10 men/5 women | - |
| ISS | 27.7 | 2.27 |
| Hospital stay | 29.0 days | 7.32 |
| ICU stay | 15.5 days | 4.15 |
| Ventilation | 8.4 days | 2.97 |
| Death | 0 | - |
| Leukocytes | 8.81/nL * | 0.92 |
| Neutrophils | 67.8% of leukocytes | 3.79 |
| Monocytes | 6.8% of leukocytes | 0.98 |
| Pneumonia | 1 patient | - |
| ARDS | 0 | - |
| Sepsis | 0 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janicova, A.; Becker, N.; Xu, B.; Simic, M.; Noack, L.; Wagner, N.; Müller, A.J.; Bertrand, J.; Marzi, I.; Relja, B. Severe Traumatic Injury Induces Phenotypic and Functional Changes of Neutrophils and Monocytes. J. Clin. Med. 2021, 10, 4139. https://doi.org/10.3390/jcm10184139
Janicova A, Becker N, Xu B, Simic M, Noack L, Wagner N, Müller AJ, Bertrand J, Marzi I, Relja B. Severe Traumatic Injury Induces Phenotypic and Functional Changes of Neutrophils and Monocytes. Journal of Clinical Medicine. 2021; 10(18):4139. https://doi.org/10.3390/jcm10184139
Chicago/Turabian StyleJanicova, Andrea, Nils Becker, Baolin Xu, Marija Simic, Laurens Noack, Nils Wagner, Andreas J. Müller, Jessica Bertrand, Ingo Marzi, and Borna Relja. 2021. "Severe Traumatic Injury Induces Phenotypic and Functional Changes of Neutrophils and Monocytes" Journal of Clinical Medicine 10, no. 18: 4139. https://doi.org/10.3390/jcm10184139