T Cell Peptides Derived from Invasive Stages of Schistosoma mansoni as Potential Schistosomiasis Vaccine

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Mice and Parasites
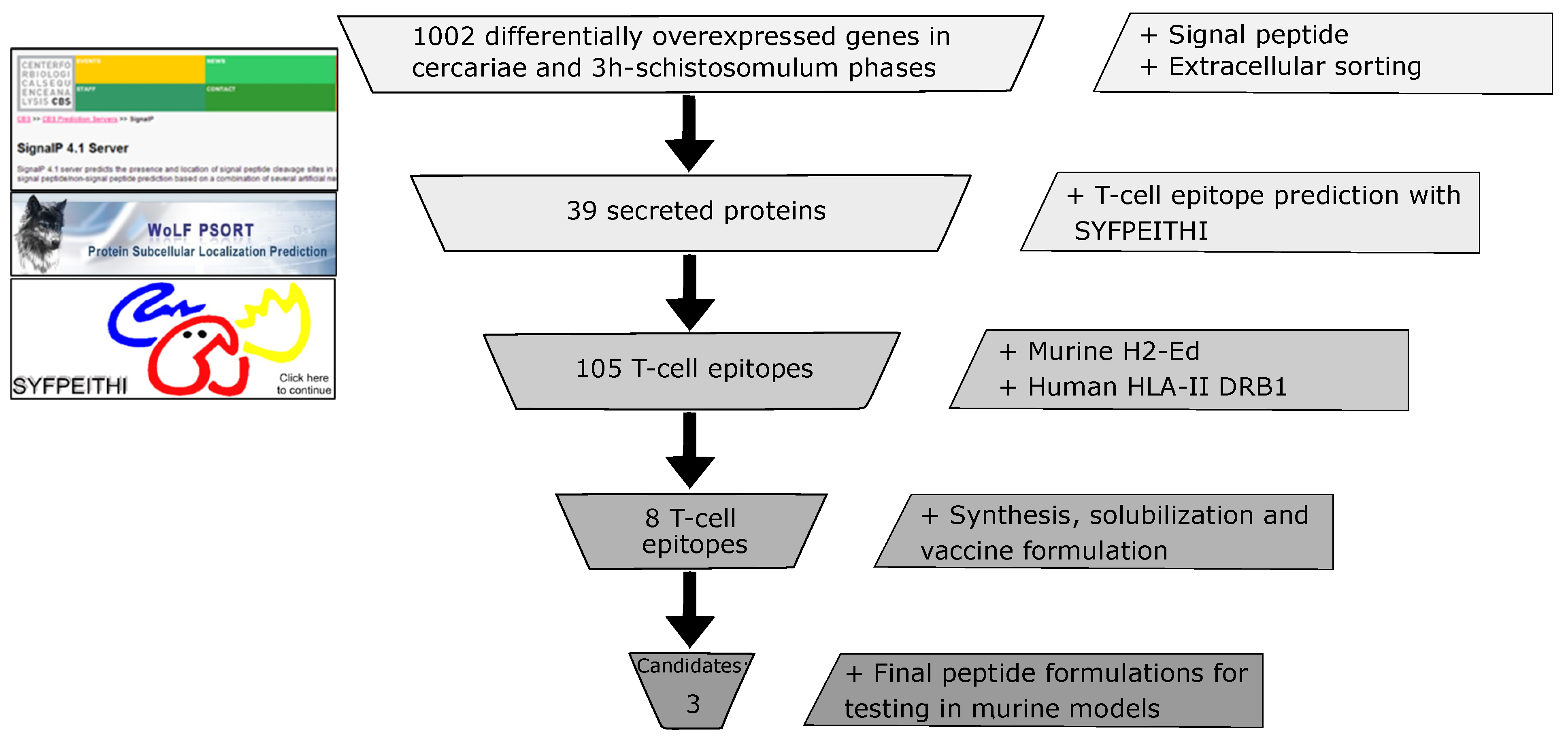
2.2. Exploring the Schistosoma mansoni Transcriptome to Identify Potential Sources of Vaccine Candidates and Design T Cell Peptide Candidates
2.3. Chemical Synthesis of T Cell Vaccine Candidates and Toxicity Study
2.4. Vaccination with the ADAD System, Challenge and Necropsy
2.5. Serum Antibodies Induced by Vaccination and Infection
2.6. Statistical Analysis
3. Results
3.1. In Silico Identification of Potential Vaccine Candidates
3.2. T Cell Epitope Characterization, Synthesis and Toxicity Assessment
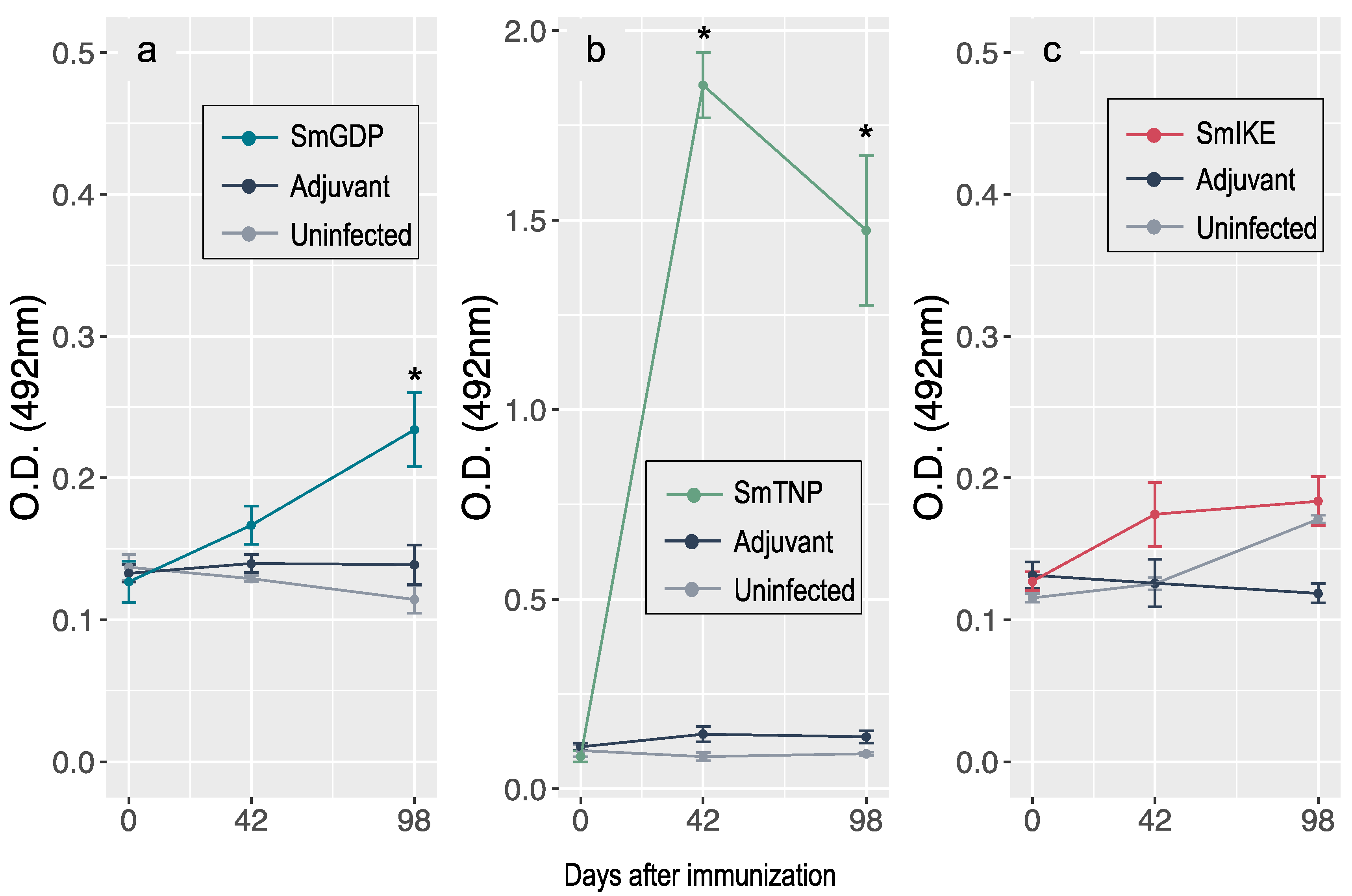
3.3. Antibody Response to Vaccination with SmGSP, SmTNP and SmIKE
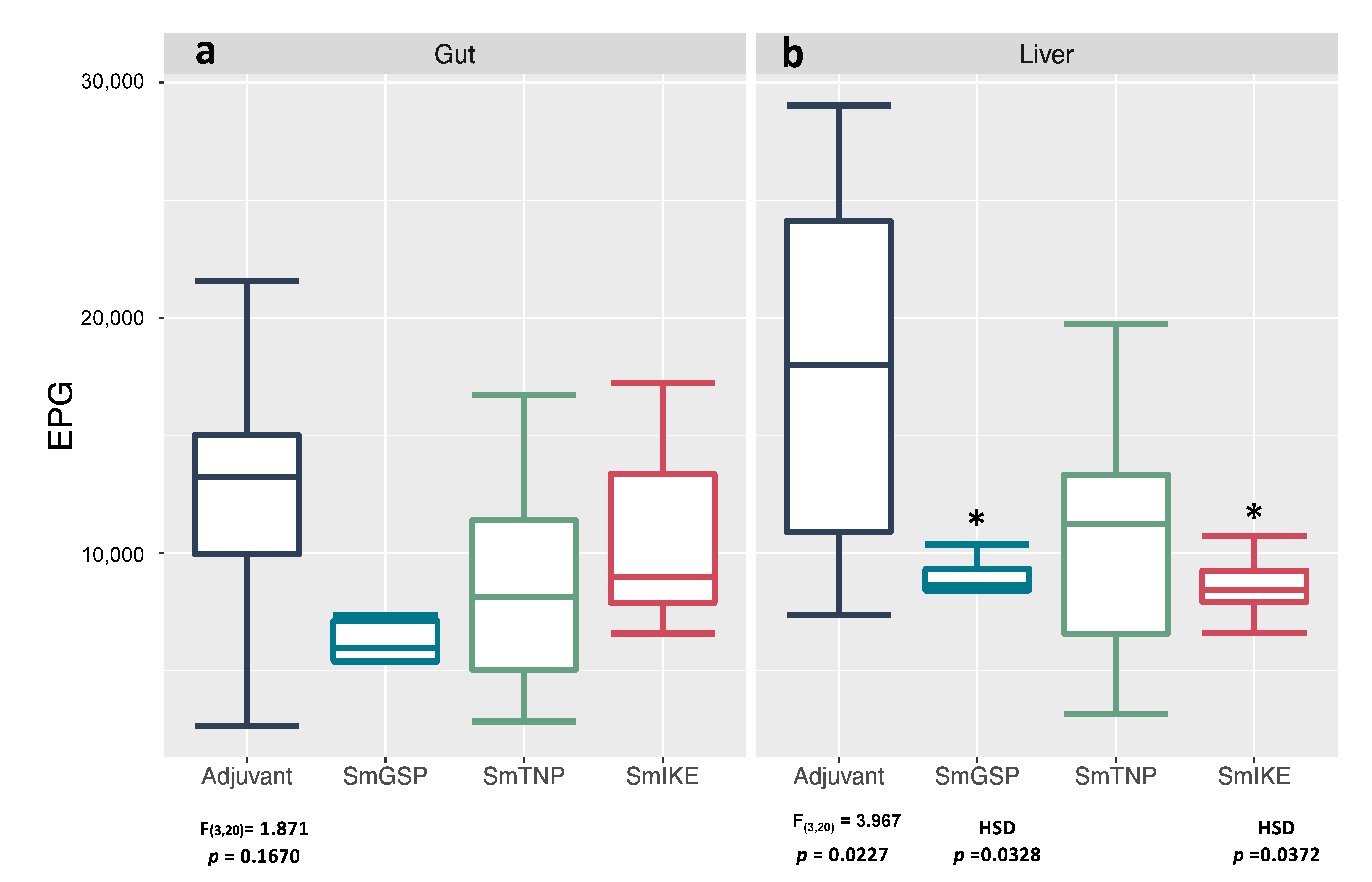
3.4. Reduction in Worm Recovery and Egg Burden
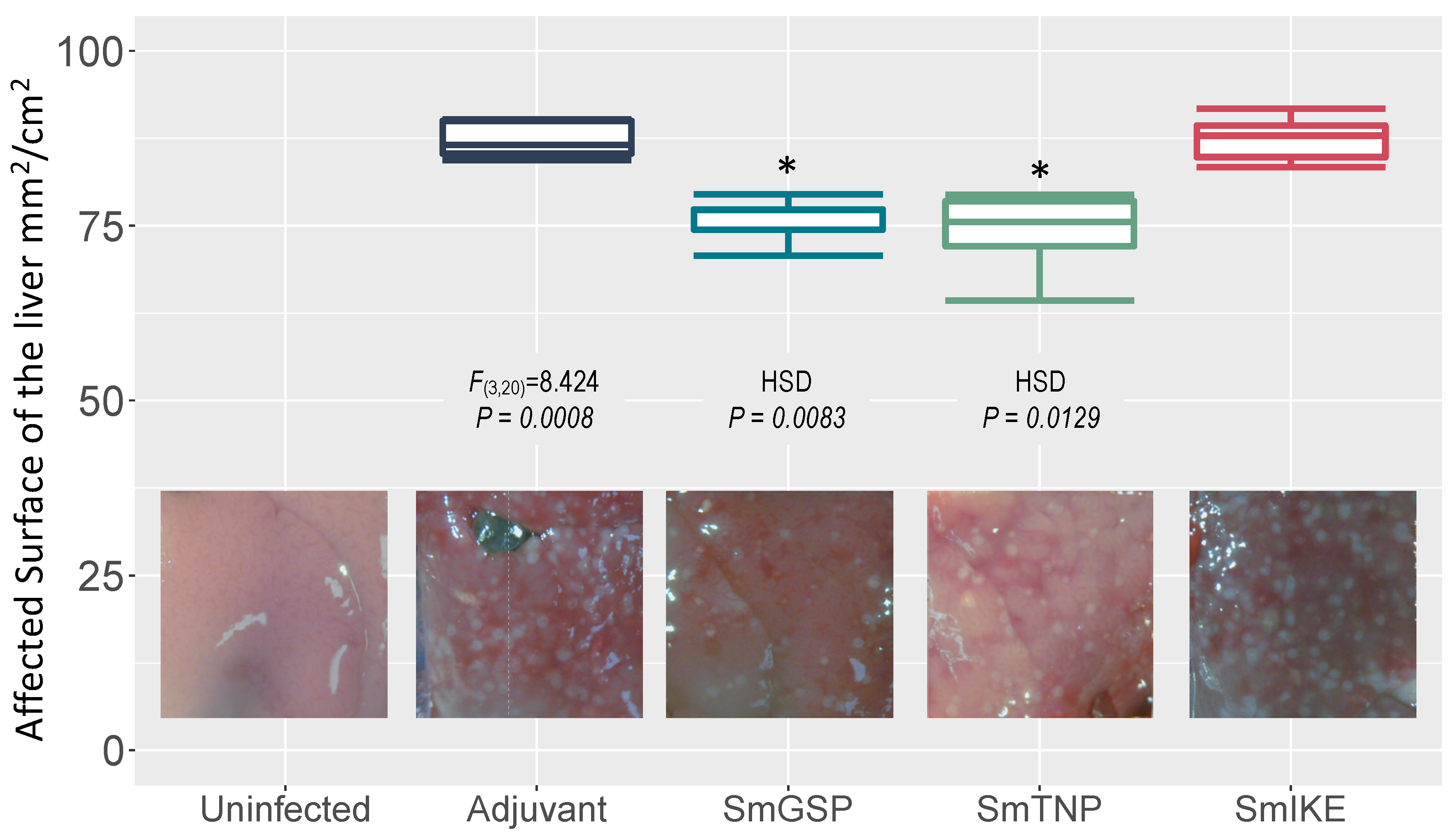
3.5. Protection against Liver Injury
3.6. Immune Response against the Infection
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 Diseases and Injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Roth, G.A.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1736–1788. [Google Scholar] [CrossRef] [Green Version]
- Kyu, H.H.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national disability-adjusted life-years (DALYs) for 359 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1859–1922. [Google Scholar] [CrossRef] [Green Version]
- Mutapi, F.; Maizels, R.; Fenwick, A.; Woolhouse, M. Human schistosomiasis in the post mass drug administration era. Lancet Infect. Dis. 2017, 17, e42–e48. [Google Scholar] [CrossRef] [Green Version]
- Riveau, G.; Schacht, A.M.; Dompnier, J.P.; Deplanque, D.; Seck, M.; Waucquier, N.; Senghor, S.; Delcroix-Genete, D.; Hermann, E.; Idris-Khodja, N.; et al. Safety and efficacy of the rSh28GST urinary schistosomiasis vaccine: A phase 3 randomized, controlled trial in Senegalese children. PLoS Negl. Trop. Dis. 2018, 12, e0006968. [Google Scholar] [CrossRef]
- Riveau, G.; Deplanque, D.; Remoué, F.; Schacht, A.M.; Vodougnon, H.; Capron, M.; Thiry, M.; Martial, J.; Libersa, C.; Capron, A. Safety and immunogenicity of rSh28GST antigen in humans: Phase 1 randomized clinical study of a vaccine candidate against urinary schistosomiasis. PLoS Negl. Trop. Dis. 2012, 6, e1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebeje, B.M.; Harvie, M.; You, H.; Loukas, A.; McManus, D.P. Schistosomiasis vaccines: Where do we stand? Parasites Vectors 2016, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sette, A.; Rappuoli, R. Reverse vaccinology: Developing vaccines in the era of genomics. Immunity 2010, 33, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Stern, L.; Calvo-Calle, J. HLA-DR: Molecular Insights and Vaccine Design. Curr. Pharm. Des. 2009, 15, 3249–3261. [Google Scholar] [CrossRef] [Green Version]
- Picard, M.A.L.; Boissier, J.; Roquis, D.; Grunau, C.; Allienne, J.F.; Duval, D.; Toulza, E.; Arancibia, N.; Caffrey, C.R.; Long, T.; et al. Sex-Biased Transcriptome of Schistosoma mansoni: Host-Parasite Interaction, Genetic Determinants and Epigenetic Regulators Are Associated with Sexual Differentiation. PLoS Negl. Trop. Dis. 2016, 10, e0004930. [Google Scholar] [CrossRef]
- López, C.; Yepes-Pérez, Y.; Díaz-Arévalo, D.; Patarroyo, M.E.; Patarroyo, M.A. The in vitro antigenicity of Plasmodium vivax rhoptry neck protein 2 (PvRON2) B- and T-epitopes selected by HLA-DRB1 binding profile. Front. Cell. Infect. Microbiol. 2018, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.S.; Wearsch, P.A.; Cresswell, P. Pathways of antigen processing. Annu. Rev. Immunol. 2013, 31, 443–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas-Caraballo, J.; López-Abán, J.; Pérez Del Villar, L.; Vizcaíno, C.; Vicente, B.; Fernández-Soto, P.; Del Olmo, E.; Patarroyo, M.A.; Muro, A. In vitro and in vivo studies for assessing the immune response and protection-inducing ability conferred by Fasciola hepatica-derived synthetic peptides containing B- and T-cell epitopes. PLoS ONE 2014, 9, e105323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berriman, M.; Haas, B.J.; Loverde, P.T.; Wilson, R.A.; Dillon, G.P.; Cerqueira, G.C.; Mashiyama, S.T.; Al-Lazikani, B.; Andrade, L.F.; Ashton, P.D.; et al. The genome of the blood fluke Schistosoma mansoni. Nature 2009, 460, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zheng, H.; Chen, Y.; Zhang, L.; Wang, K.; Guo, J.; Huang, Z.; Zhang, B.; Huang, W.; Jin, K.; et al. The Schistosoma japonicum genome reveals features of host-parasite interplay. Nature 2009, 460, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Young, N.D.; Jex, A.R.; Li, B.; Liu, S.; Yang, L.; Xiong, Z.; Li, Y.; Cantacessi, C.; Hall, R.S.; Xu, X.; et al. Whole-genome sequence of Schistosoma haematobium. Nat. Genet. 2012, 44, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Crellen, T.; Allan, F.; David, S.; Durrant, C.; Huckvale, T.; Holroyd, N.; Emery, A.M.; Rollinson, D.; Aanensen, D.M.; Berriman, M.; et al. Whole genome resequencing of the human parasite Schistosoma mansoni reveals population history and effects of selection. Sci. Rep. 2016, 6, 20954. [Google Scholar] [CrossRef] [Green Version]
- Protasio, A.V.; Tsai, I.J.; Babbage, A.; Nichol, S.; Hunt, M.; Aslett, M.A.; de Silva, N.; Velarde, G.S.; Anderson, T.J.C.; Clark, R.C.; et al. A systematically improved high quality genome and transcriptome of the human blood fluke Schistosoma mansoni. PLoS Negl. Trop. Dis. 2012, 6, e1455. [Google Scholar] [CrossRef]
- Gobert, G.N.; Tran, M.H.; Moertel, L.; Mulvenna, J.; Jones, M.K.; McManus, D.P.; Loukas, A. Transcriptional changes in Schistosoma mansoni during early schistosomula development and in the presence of erythrocytes. PLoS Negl. Trop. Dis. 2010, 4, e600. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.; Amaral, M.S.; Beckedorff, F.; Silva, L.F.; Dazzani, B.; Oliveira, K.C.; Almeida, G.T.; Gomes, M.R.; Pires, D.S.; Setubal, J.C.; et al. Schistosoma mansoni Egg, Adult Male and Female Comparative Gene Expression Analysis and Identification of Novel Genes by RNA-Seq. PLoS Negl. Trop. Dis. 2015, 9, e0004334. [Google Scholar] [CrossRef]
- Nielsen, H. Predicting secretory proteins with signaIP. Methods Mol Biol. 2017, 1611, 59–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, N.; Shrestha, A.; Ruttkowski, B.; Beck, T.; Vogl, C.; Tomley, F.; Blake, D.P.; Joachim, A. The genome of the protozoan parasite Cystoisospora suis and a reverse vaccinology approach to identify vaccine candidates. Int. J. Parasitol. 2017, 47, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Raghava, G.P.S. AlgPred: Prediction of allergenic proteins and mapping of IgE epitopes. Nucleic Acids Res. 2006, 34. [Google Scholar] [CrossRef] [PubMed]
- Rammensee, H.G.; Bachmann, J.; Emmerich, N.P.N.; Bachor, O.A.; Stevanović, S. SYFPEITHI: Database for MHC ligands and peptide motifs. Immunogenetics 1999, 50, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.; Lund, O. NN-align. An artificial neural network-based alignment algorithm for MHC class II peptide binding prediction. BMC Bioinform. 2009, 10, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrifield, R.B. Solid-Phase Peptide Synthesis. In Advances in Enzymology and Related Areas of Molecular Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1969; Volume 32, pp. 221–296. ISBN 9780470122778. [Google Scholar] [PubMed]
- Houghten, R.A. General method for the rapid solid-phase synthesis of large numbers of peptides: Specificity of antigen-antibody interaction at the level of individual amino acids. Proc. Natl. Acad. Sci. USA 1985, 82, 5131–5135. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Chen, M.; Kaushal, S.; McElroy, M.; Zhang, Y.; Ozkan, C.; Bouvet, M.; Kruse, C.; Grotjahn, D.; Ichim, T.; et al. PLGA nanoparticle-mediated delivery of tumor antigenic peptides elicits effective immune responses. Int. J. Nanomed. 2012, 7, 1475–1487. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Fernández, A.R.; Nogal-Ruiz, J.J.; López-Abán, J.; Ramajo, V.; Oleaga, A.; Manga-González, Y.; Hillyer, G.V.; Muro, A. Vaccination of mice and sheep with Fh12 FABP from Fasciola hepatica using the new adjuvant/immunomodulator system ADAD. Vet. Parasitol. 2004, 126, 287–298. [Google Scholar] [CrossRef]
- del Olmo, E.; Plaza, A.; Muro, A.; Martínez-Fernández, A.R.; Nogal-Ruiz, J.J.; López-Pérez, J.L.; San Feliciano, A. Synthesis and evaluation of some lipidic aminoalcohols and diamines as immunomodulators. Bioorganic Med. Chem. Lett. 2006, 16, 6091–6095. [Google Scholar] [CrossRef]
- Charan, J.; Kantharia, N. How to calculate sample size in animal studies? J. Pharmacol. Pharmacother. 2013, 4, 303–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festing, M.F.W.; Altman, D.G. Guidelines for the design and statistical analysis of experiments using laboratory animals. ILAR J. 2002, 43, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Vicente, B.; López-Abán, J.; Rojas-Caraballo, J.; Del Olmo, E.; Fernández-Soto, P.; Muro, A. Protection against Schistosoma mansoni infection using a Fasciola hepatica-derived fatty acid binding protein from different delivery systems. Parasites Vectors 2016, 9, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wang, L.; Liang, Y.S. Susceptibility or resistance of praziquantel in human schistosomiasis: A review. Parasitol. Res. 2012, 111, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Lotfy, W.M.; Hishmat, M.G.; El Nashar, A.S.; Abu El Einin, H.M. Evaluation of a method for induction of praziquantel resistance in Schistosoma mansoni. Pharm. Biol. 2015, 53, 1214–1219. [Google Scholar] [CrossRef]
- Merrifield, M.; Hotez, P.J.; Beaumier, C.M.; Gillespie, P.; Strych, U.; Hayward, T.; Bottazzi, M.E. Advancing a vaccine to prevent human schistosomiasis. Vaccine 2016, 34, 2988–2991. [Google Scholar] [CrossRef] [Green Version]
- Nowacki, F.C.; Swain, M.T.; Klychnikov, O.I.; Niazi, U.; Ivens, A.; Quintana, J.F.; Hensbergen, P.J.; Hokke, C.H.; Buck, A.H.; Hoffmann, K.F. Protein and small non-coding RNA-enriched extracellular vesicles are released by the pathogenic blood fluke Schistosoma mansoni. J. Extracell. Vesicles 2015, 4, 28665. [Google Scholar] [CrossRef] [Green Version]
- Sotillo, J.; Pearson, M.; Potriquet, J.; Becker, L.; Pickering, D.; Mulvenna, J.; Loukas, A. Extracellular vesicles secreted by Schistosoma mansoni contain protein vaccine candidates. Int. J. Parasitol. 2016, 46, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Liu, S.; Piao, X.; Hou, N.; Gobert, G.N.; McManus, D.P.; Chen, Q. Comprehensive Transcriptome Analysis of Sex-Biased Expressed Genes Reveals Discrete Biological and Physiological Features of Male and Female Schistosoma japonicum. PLoS Negl. Trop. Dis. 2016, 10, e0004684. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Liu, S.; Piao, X.; Hou, N.; You, H.; McManus, D.P.; Chen, Q. A next-generation microarray further reveals stage-enriched gene expression pattern in the blood fluke Schistosoma japonicum. Parasites Vectors 2017, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Patarroyo, M.E.; Bermúdez, A.; Patarroyo, M.A. Structural and immunological principles leading to chemically synthesized, multiantigenic, multistage, minimal subunit-based vaccine development. Chem. Rev. 2011, 111, 3459–3507. [Google Scholar] [CrossRef] [PubMed]
- Agallou, M.; Margaroni, M.; Athanasiou, E.; Toubanaki, D.K.; Kontonikola, K.; Karidi, K.; Kammona, O.; Kiparissides, C.; Karagouni, E. Identification of BALB/c Immune Markers Correlated with a Partial Protection to Leishmania infantum after Vaccination with a Rationally Designed Multi-epitope Cysteine Protease A Peptide-Based Nanovaccine. PLoS Negl. Trop. Dis. 2017, 11, e0005311. [Google Scholar] [CrossRef] [PubMed]
- Tallima, H.; Hanna, V.S.; El Ridi, R. Arachidonic Acid Is a Safe and Efficacious Schistosomicide, and an Endoschistosomicide in Natural and Experimental Infections, and Cysteine Peptidase Vaccinated Hosts. Front. Immunol. 2020, 11, 609994. [Google Scholar] [CrossRef] [PubMed]
- McManus, D.P. Recent progress in the development of liver fluke and blood fluke vaccines. Vaccines 2020, 8, 553. [Google Scholar] [CrossRef] [PubMed]
- Eyayu, T.; Zeleke, A.J.; Worku, L. Current status and future prospects of protein vaccine candidates against Schistosoma mansoni infection. Parasite Epidemiol. Control 2020, 11, e00176. [Google Scholar] [CrossRef] [PubMed]
- Davids, B.J.; Yoshino, T.P. Schistosoma mansoni: Excretory-secretory polypeptides exhibit selective binding to plasma components of the snail Biomphalaria glabrata. Exp. Parasitol. 1995, 81, 292–301. [Google Scholar] [CrossRef]
- Qiu, C.; Hong, Y.; Cao, Y.; Wang, F.; Fu, Z.; Shi, Y.; Wei, M.; Liu, S.; Lin, J. Molecular cloning and characterization of glutamine synthetase, a tegumental protein from Schistosoma japonicum. Parasitol. Res. 2012, 111, 2367–2376. [Google Scholar] [CrossRef]
- Bischofsberger, M.; Winkelmann, F.; Rabes, A.; Reisinger, E.C.; Sombetzki, M. Pathogen-host interaction mediated by vesicle-based secretion in schistosomes. Protoplasma 2020, 257, 1277–1287. [Google Scholar] [CrossRef]
- Ranasinghe, S.L.; Duke, M.; Harvie, M.; McManus, D.P. Kunitz-type protease inhibitor as a vaccine candidate against schistosomiasis mansoni. Int. J. Infect. Dis. 2018, 66, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Morais, S.B.; Figueiredo, B.C.; Assis, N.R.G.; Homan, J.; Mambelli, F.S.; Bicalho, R.M.; Souza, C.; Martins, V.P.; Pinheiro, C.S.; Oliveira, S.C. Schistosoma mansoni SmKI-1 or Its C-Terminal Fragment Induces Partial Protection Against S. mansoni Infection in Mice. Front. Immunol. 2018, 9, 1762. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Goenaga, J.; López-Abán, J.; Protasio, A.V.; Vicente Santiago, B.; del Olmo, E.; Vanegas, M.; Fernández-Soto, P.; Patarroyo, M.A.; Muro, A. Peptides Derived of Kunitz-Type Serine Protease Inhibitor as Potential Vaccine Against Experimental Schistosomiasis. Front. Immunol. 2019, 10, 2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, A.S.; Zelt, N.H.; Perera, D.J.; Ndao, M.; Ward, B.J. Vaccination against the digestive enzyme Cathepsin B using a YS1646 Salmonella enterica Typhimurium vector provides almost complete protection against Schistosoma mansoni challenge in a mouse model. PLoS Negl. Trop. Dis. 2019, 13, e0007490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallima, H.; Dvořák, J.; Kareem, S.; Abou El Dahab, M.; Abdel Aziz, N.; Dalton, J.P.; El Ridi, R. Protective immune responses against Schistosoma mansoni infection by immunization with functionally active gut-derived cysteine peptidases alone and in combination with glyceraldehyde 3-phosphate dehydrogenase. PLoS Negl. Trop. Dis. 2017, 11, e0005443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad Fuaad, A.A.H.; Roubille, R.; Pearson, M.S.; Pickering, D.A.; Loukas, A.C.; Skwarczynski, M.; Toth, I. The use of a conformational cathepsin D-derived epitope for vaccine development against Schistosoma mansoni. Bioorganic Med. Chem. 2015, 23, 1307–1312. [Google Scholar] [CrossRef] [PubMed]
- Stylianou, A.; Hadjichrysanthou, C.; Truscott, J.E.; Anderson, R.M. Developing a mathematical model for the evaluation of the potential impact of a partially efficacious vaccine on the transmission dynamics of Schistosoma mansoni in human communities. Parasites Vectors 2017, 10, 294. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SmGSP | SmTNP | SmIKE | |
|---|---|---|---|
| GeneDB | Smp_136730 | Smp_136920 | Smp_021460 |
| Genebank | CCD78846 | CCD79787 | CAZ29822 |
| Protein length | 401 | 447 | 555 |
| Product | Subfamily A1A unassigned peptidase | Polypeptide having N-acetyl galactosaminyl transferase activity | Putative bacteria glutamine synthetase |
| Signal peptide position | 1–17 | 1–23 | 1–23 |
| T epitope | GSPYLWIPSKKCDPS | TNPYHLIYPQKSPAL | IKELEDKYNLRLYSA |
| Position | 97–111 | 271–285 | 84–98 |
| Fitting murine MHC (Score < 20) | H2-Ed | H2-Ed | H2-Ed |
| Fitting human MHC (Score < 20) | DRB1*0101 DRB1*0401 DRB1*0701 | DRB1*0101 DRB1*0401 DRB1*1101 | DRB1*0301 DRB1*0701 DRB1*1501 |
| J774.2 macrophage viability (%) | 96.8 | 97.9 | 96.4 |
| Group | Total Worms | Mean ± SEM | R (%) | Female Worms | Mean ± SEM | R (%) | Male Worms | Mean ± SEM | R (%) |
|---|---|---|---|---|---|---|---|---|---|
| Adjuvant | 26, 27, 25, 35, 18, 18 | 24.8 ± 2.6 | - | 18, 18, 16, 19, 13, 10 | 15.7 ± 1.4 | - | 8, 9, 9, 16, 5, 8 | 9.2 ± 1.5 | - |
| SmGSP | 6, 32, 16, 8, 21, 13 | 16.0 ± 3.9 | 36 | 2, 21, 9, 4, 12, 7 | 9.2 ± 2.8 | 42 | 4, 11, 7, 4, 9, 6 | 6.8 ± 1.1 | 26 |
| SmTNP | 20, 7, 21, 14, 22, 37 | 21.2 ± 4.1 | 19 | 10, 4, 11, 10, 13, 24 | 12.0 ± 2.7 | 23 | 10, 3, 10, 4, 9, 13 | 8.2 ± 1.6 | 11 |
| SmIKE | 29, 14, 23, 27, 37, 39 | 28.2 ± 3.8 | - | 17, 7, 14, 14, 27, 24 | 17.2 ± 3.0 | - | 12, 7, 9, 13, 10, 15 | 11.0 ± 1.2 | - |
| ANOVA | F(3,20) = 2.153 p = 0.125 | F(3,20) = 2.016 p = 0.144 | F(3,20) = 1.662 p = 0.207 |
| Group | IgG | IgG1 | IgG2a |
|---|---|---|---|
| Uninfected | 0.194 ± 0.002 | 0.201 ± 0.004 | 0.111 ± 0.007 |
| Adjuvant | 0.577 ± 0.101 * HSD p = 0.002 | 0.653 ± 0.010 * HSD p = 0.003 | 0.138 ± 0.016 |
| SmGSP | 0.634 ± 0.049 * HSD p < 0.001 | 0.522 ± 0.050 | 0.210 ± 0.065 |
| SmTNP | 0.599 ± 0.047 * HSD p < 0.001 | 0.719 ± 0.094 * HSD p < 0.001 | 0.126 ± 0.012 |
| SmIKE | 0.697 ± 0.061 * HSD p < 0.001 | 0.703 ± 0.104 * HSD p = 0.001 | 0.153 ± 0.022 |
| ANOVA | F(4,25) = 10.60 p < 0.001 | F(4,25) = 7.62 p < 0.001 | F(4,25) = 1.43 p = 0.2543 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Abán, J.; Vicente, B.; Kabbas-Piñango, E.; Hernández-Goenaga, J.; Sánchez-Montejo, J.; Aguiriano, M.; del Olmo, E.; Vanegas, M.; Patarroyo, M.A.; Muro, A. T Cell Peptides Derived from Invasive Stages of Schistosoma mansoni as Potential Schistosomiasis Vaccine. J. Clin. Med. 2021, 10, 445. https://doi.org/10.3390/jcm10030445
López-Abán J, Vicente B, Kabbas-Piñango E, Hernández-Goenaga J, Sánchez-Montejo J, Aguiriano M, del Olmo E, Vanegas M, Patarroyo MA, Muro A. T Cell Peptides Derived from Invasive Stages of Schistosoma mansoni as Potential Schistosomiasis Vaccine. Journal of Clinical Medicine. 2021; 10(3):445. https://doi.org/10.3390/jcm10030445
Chicago/Turabian StyleLópez-Abán, Julio, Belén Vicente, Elías Kabbas-Piñango, Juan Hernández-Goenaga, Javier Sánchez-Montejo, María Aguiriano, Esther del Olmo, Magnolia Vanegas, Manuel Alfonso Patarroyo, and Antonio Muro. 2021. "T Cell Peptides Derived from Invasive Stages of Schistosoma mansoni as Potential Schistosomiasis Vaccine" Journal of Clinical Medicine 10, no. 3: 445. https://doi.org/10.3390/jcm10030445