
Pseudolycoriella hygida (Sauaia and Alves)—An Overview of a Model Organism in Genetics, with New Aspects in Morphology and Systematics

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivation and Life Cycle of Pseudolycoriella hygida
2.2. Molecular Methods and Techniques
2.2.1. Preparation of Polytene Chromosome Spreads
2.2.2. Preparation of Polytene Chromosome Maps
2.2.3. Preparation of Mitotic/Meiotic Chromosome Schemes
2.2.4. Molecular Analysis Based on COI Barcodes
2.3. Morphological Methods and Techniques
2.3.1. Slide-Mounting and Imaging of Immature Stages
2.3.2. Slide-Mounting and Imaging of Adults
2.4. Species Identification and Classification
2.5. Abbreviations
3. Results
3.1. Pseudolycoriella hygida in the Literature
3.2. Development and Behavior of Immatures and Adults
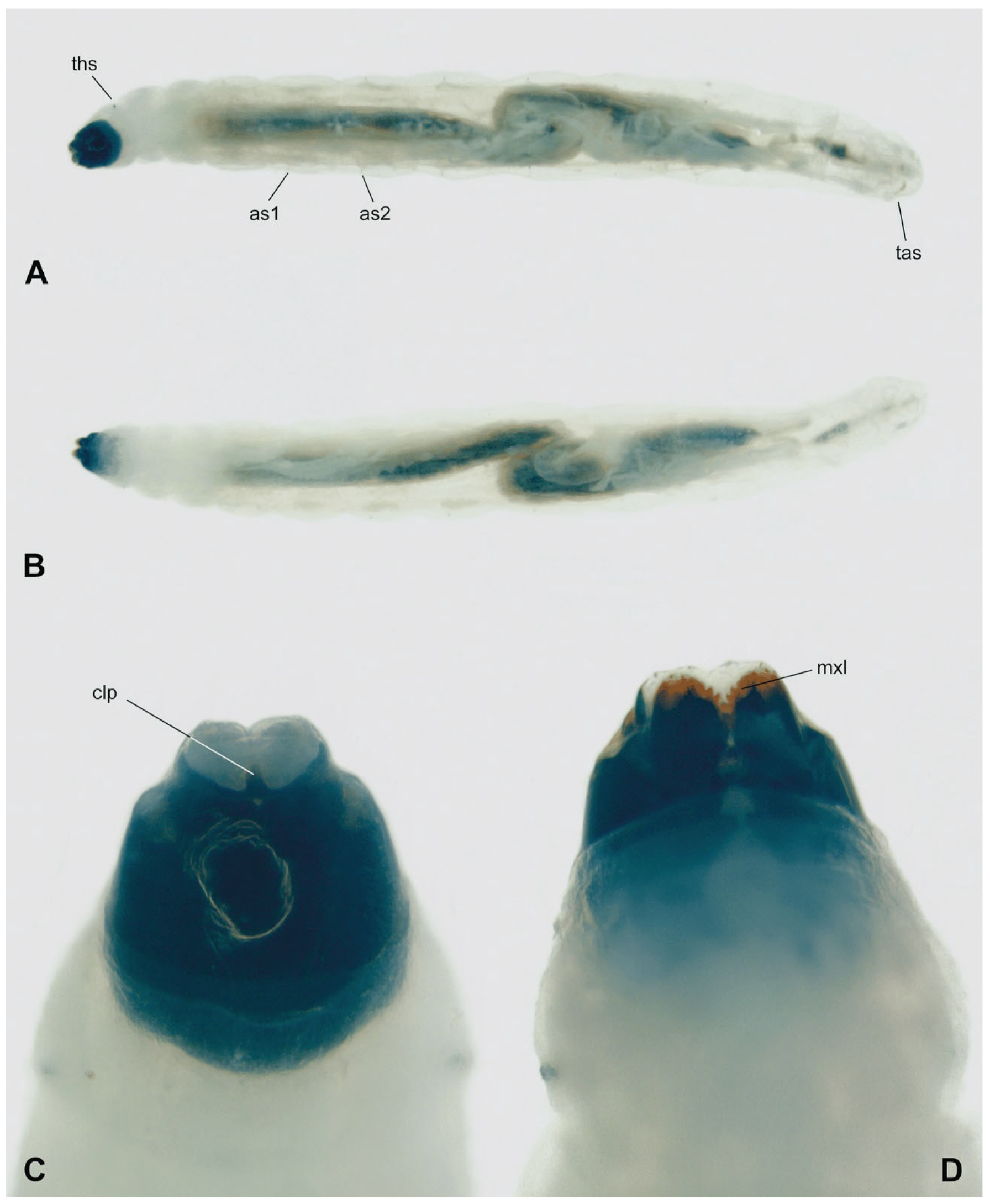
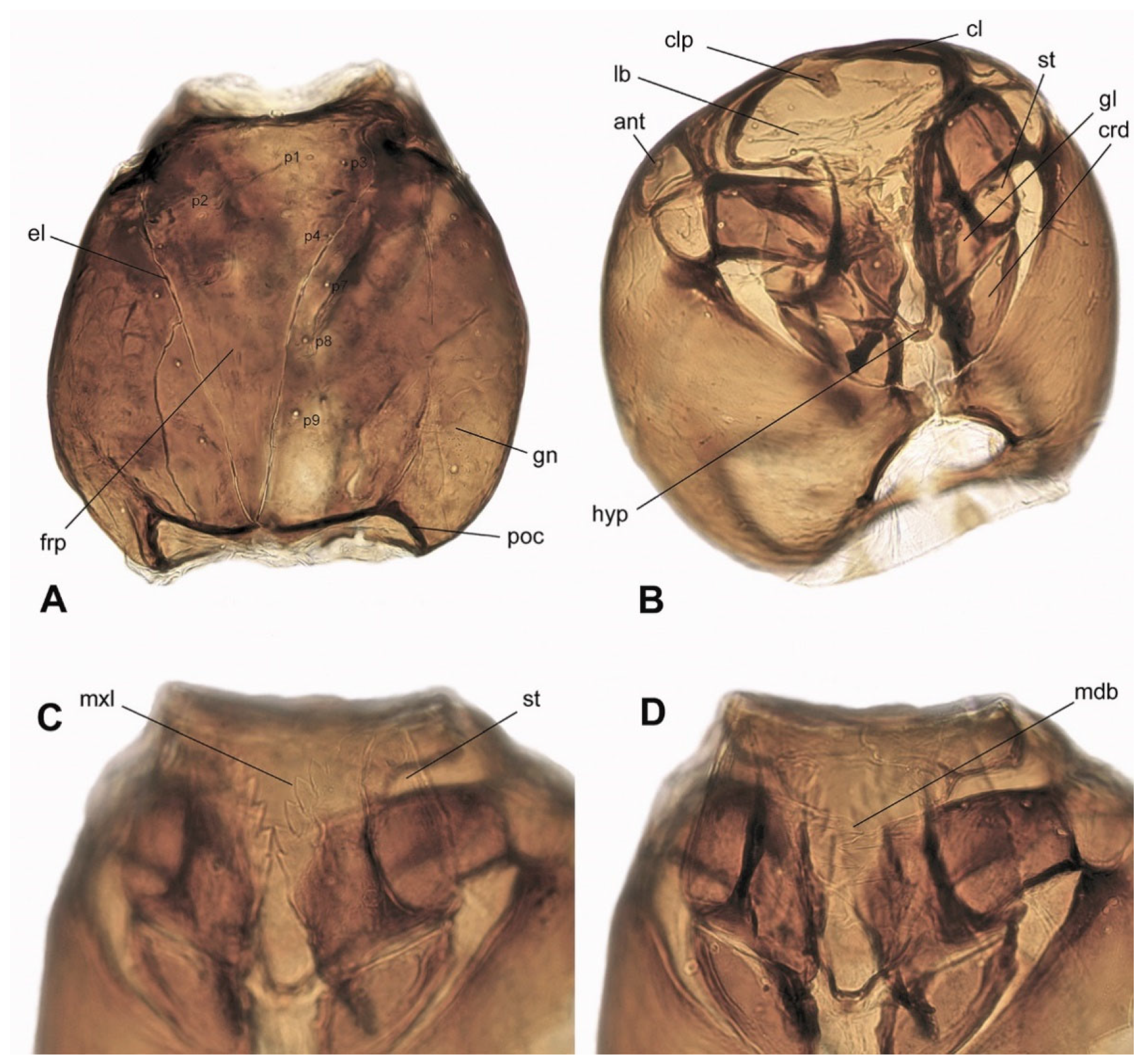
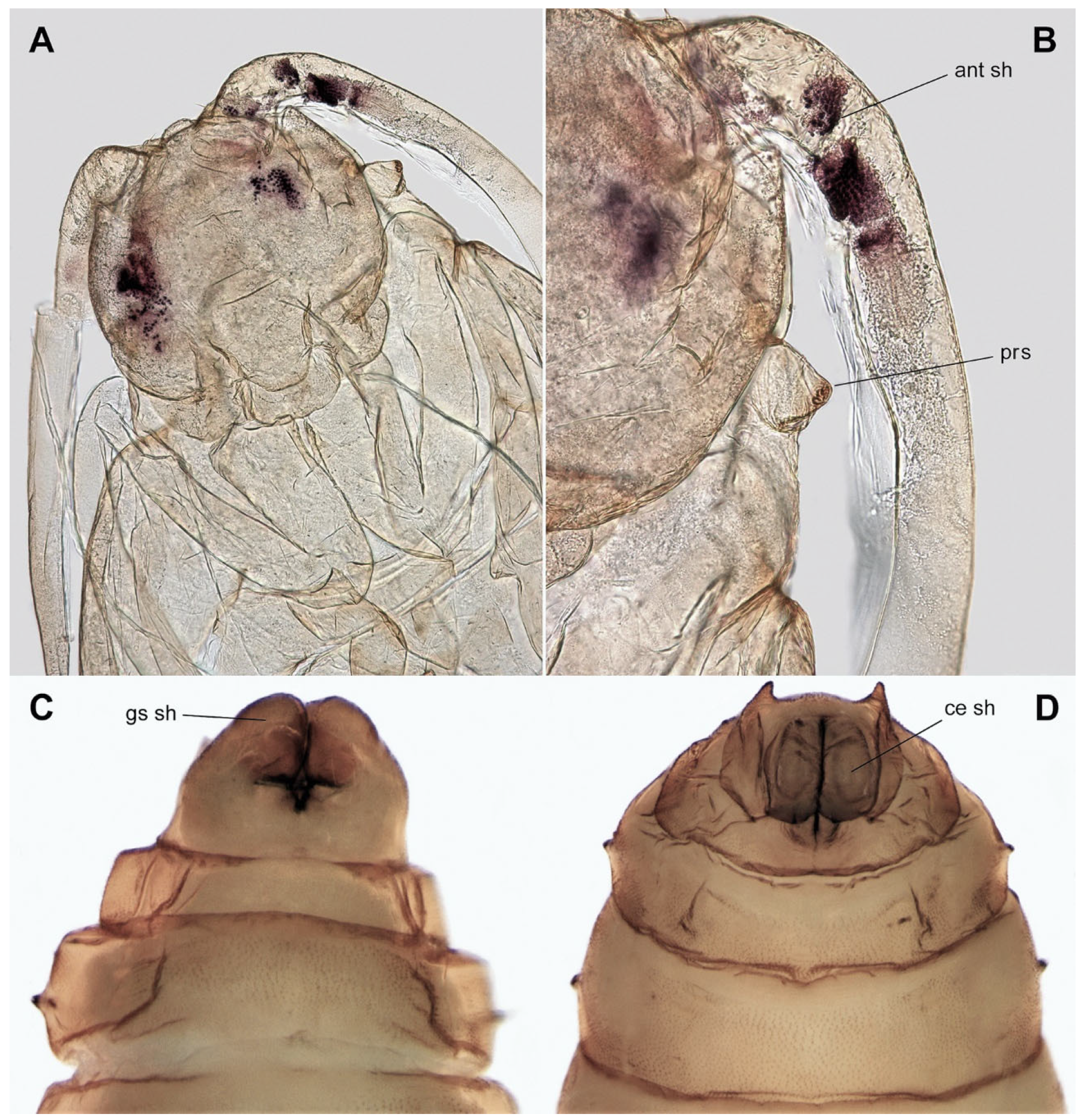
3.3. Description of Immature Stages
3.4. Redescription of Adults and Distribution of Pseudolycoriella hygida
3.5. Identification and Classification of Pseudolycoriella hygida Based on Morphological Characters of Adults
3.6. Genetic Identification and Relationship to Other COI-Analyzed Pseudolycoriella
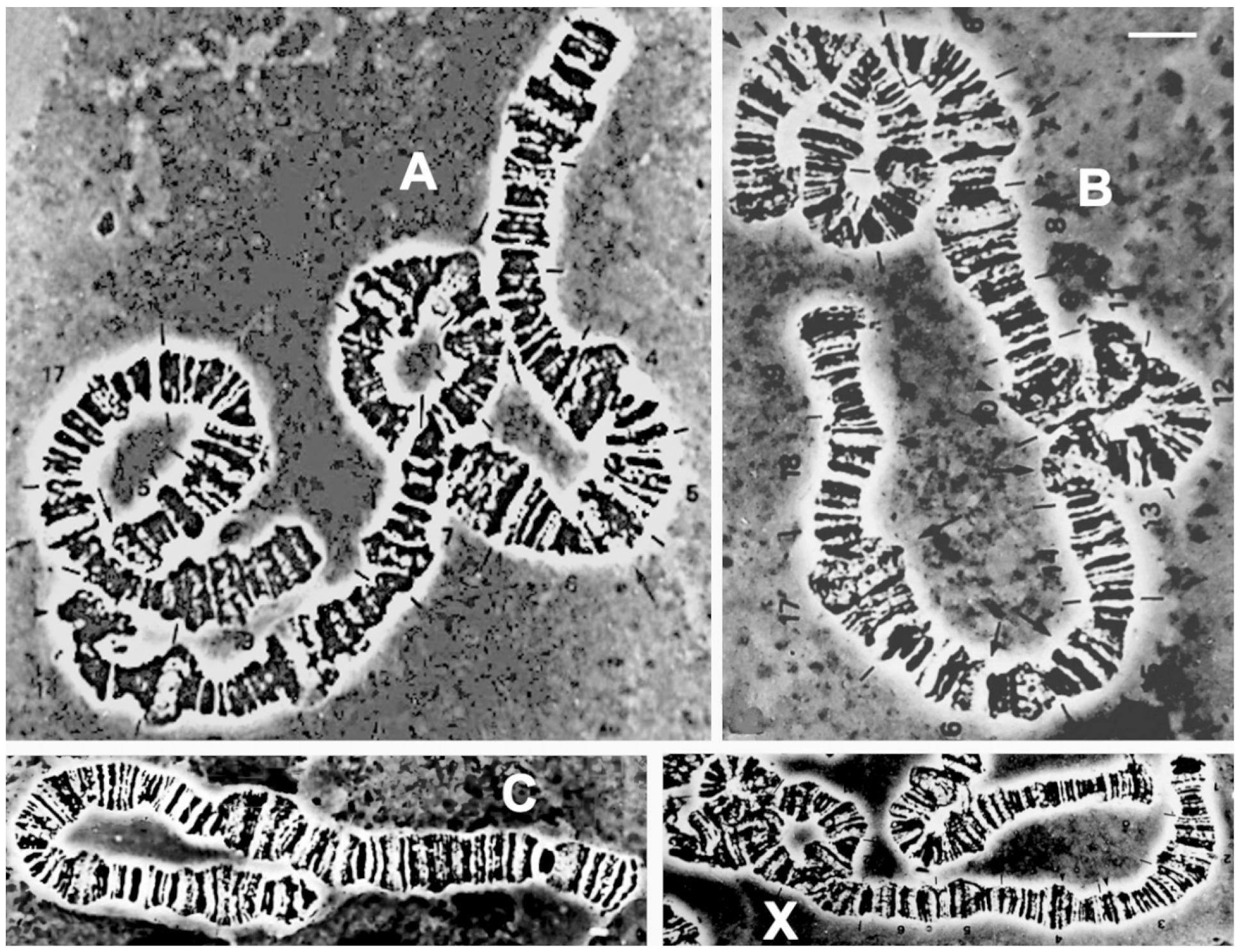
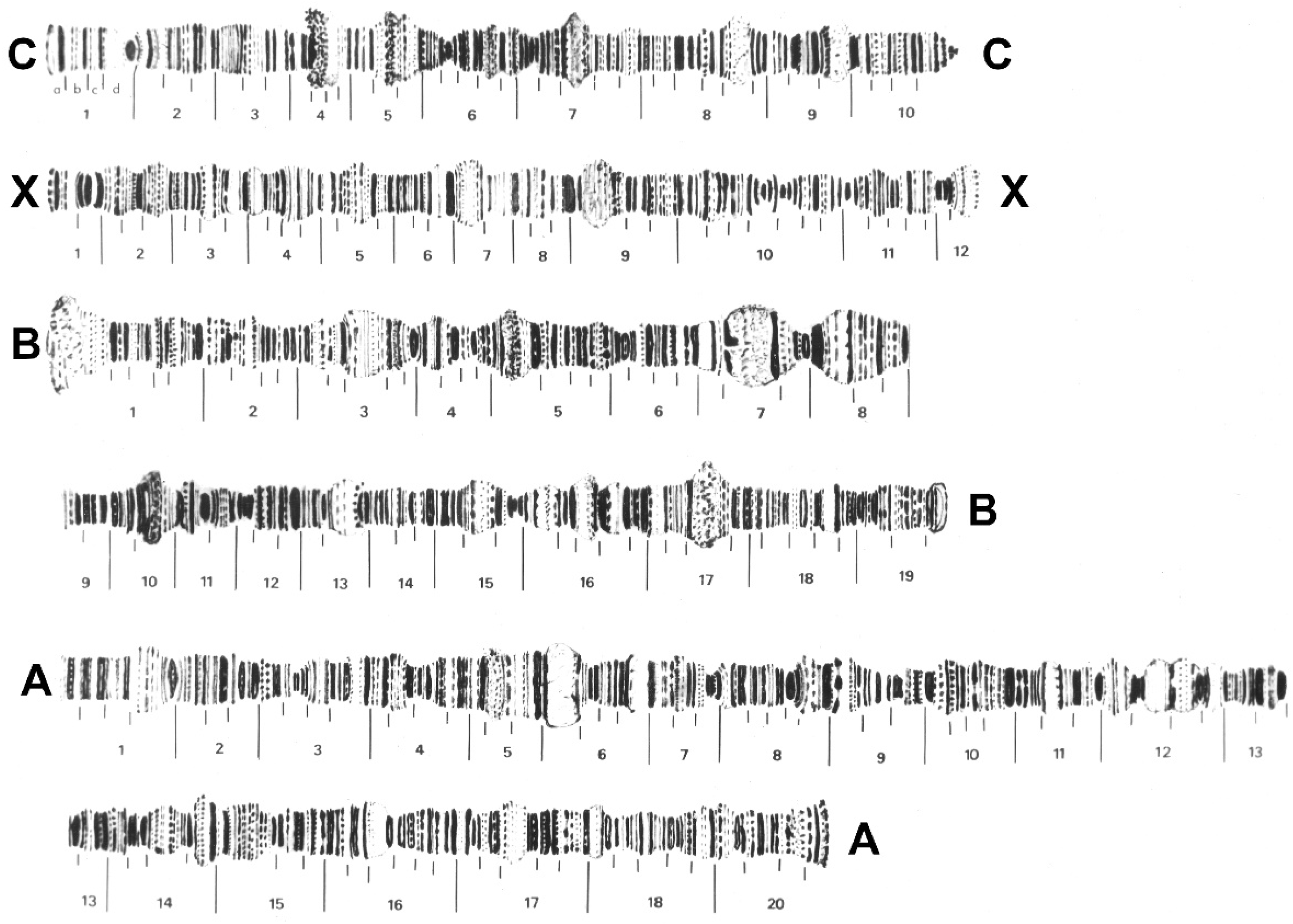
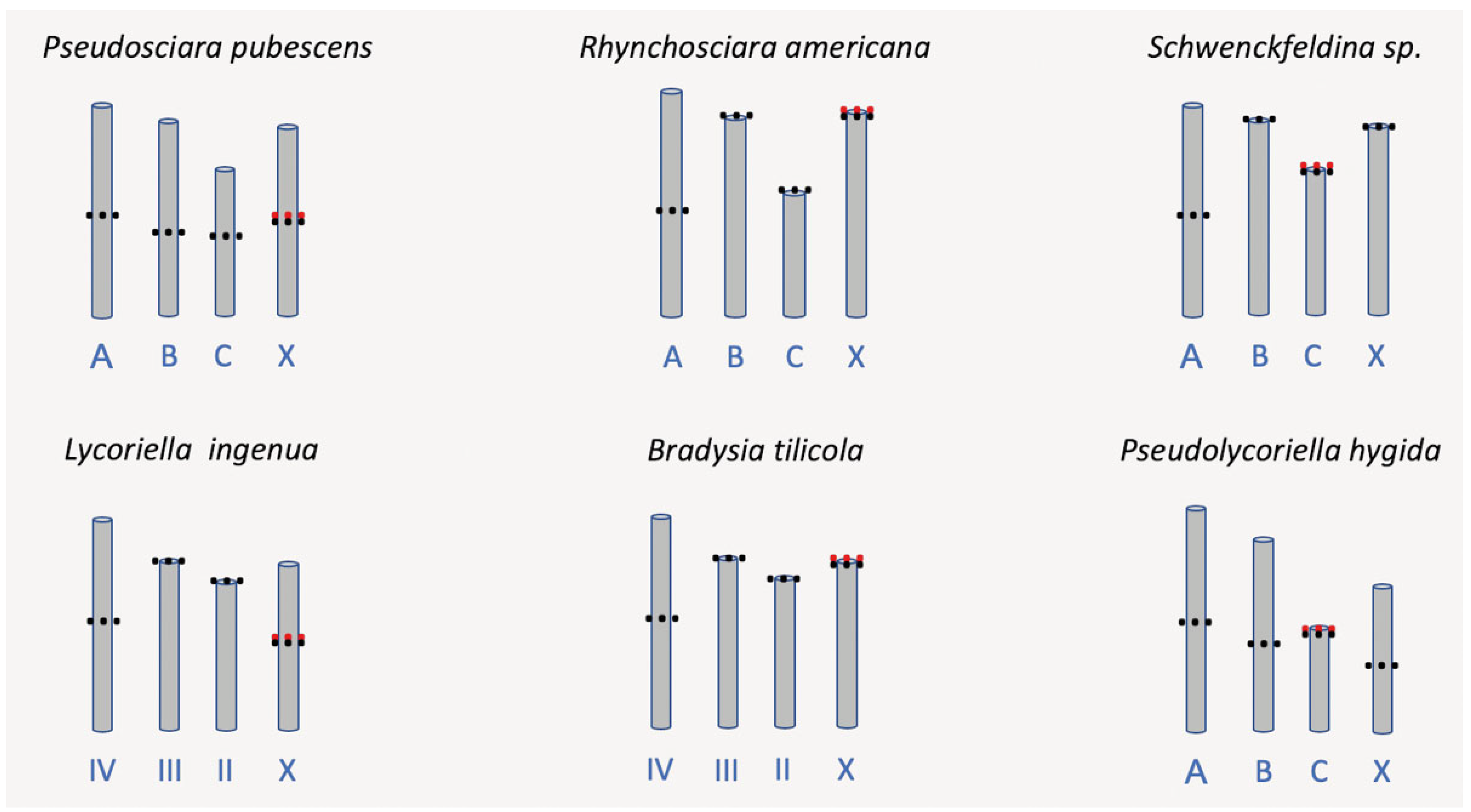
3.7. Cytogenetics
4. Discussion
4.1. Breeding of Pseudolycoriella hygida and Morphology of Immature Stages
4.2. Unknown DNA Sequences in Pseudolycoriella: A Challenge in Phylogenetic Systematics
4.3. Pseudolycoriella hygida as a Model Organism in Cytogenetics
4.4. Sciarid Names Used in Cytogenetics and Molecular Studies: Towards an Integrative Approach to Clarifying Scientific Problems
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vilkamaa, P.; Burdíková, N.; Ševčík, J. The genus Spinopygina gen. nov. (Diptera, Sciaridae) from Western North America: Preliminary molecular phylogeny and description of seven new species. Insects 2023, 14, 173. [Google Scholar] [CrossRef] [PubMed]
- Vilkamaa, P.; Menzel, F. Re-classification of Lycoriella Frey sensu lato (Diptera, Sciaridae), with description of Trichocoelina gen. n. and twenty new species. Zootaxa 2019, 4665, 1–67. [Google Scholar] [CrossRef] [PubMed]
- Köhler, A. The genus Pseudolycoriella Menzel & Mohrig, 1998 (Diptera, Sciaridae) in New Zealand. Zootaxa 2019, 4707, 1–69. [Google Scholar] [CrossRef]
- Mohrig, W.; Kauschke, E. New black fungus gnats (Diptera, Sciaridae) of North America. Part V. Genera Pseudolycoriella Menzel & Mohrig and Phytosciara Frey. Zootaxa 2019, 4543, 261–283. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Jung, S.; Menzel, F.; Heller, K.; Lee, H.; Lee, S. Molecular phylogeny of black fungus gnats (Diptera: Sciaroidea: Sciaridae) and the evolution of larval habitats. Mol. Phylogenet. Evol. 2013, 66, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Menzel, F.; Smith, J.E. 21 Sciaridae (Black Fungus Gnats). In Manual of Afrotropical Diptera. Volume 2. Nematocerous Diptera and lower Brachycera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; SANBI Graphics & Editing: Pretoria, South Africa, 2017; Volume 5, pp. 557–580. [Google Scholar]
- Mohrig, W.; Heller, K.; Hippa, H.; Vilkamaa, P.; Menzel, F. Revision of black fungus gnats (Diptera: Sciaridae) of North America. Stud. Dipterol. 2013, 19, 141–286. [Google Scholar]
- Mohrig, W.; Menzel, F. Revision der neotropischen Trauermücken—Teil I. Die Gattungen Cratyna Winnertz, Euricrium Enderlein, Metangela Rübsaamen, Pseudosciara Schiner und Sciara Meigen (Diptera: Sciaridae). Contrib. Entomol. 2014, 64, 135–190. [Google Scholar] [CrossRef]
- Babytskiy, A.I.; Moroz, M.S.; Kalashnyk, S.O.; Bezsmertna, O.O.; Dudiak, I.D.; Voitsekhivska, O.V. New findings of pest sciarid species (Diptera, Sciaridae) in Ukraine, with the first record of Bradysia difformis. Biosyst. Divers. 2019, 27, 131–141. [Google Scholar] [CrossRef]
- Broadley, A.; Kauschke, E.; Mohrig, W. Black fungus gnats (Diptera: Sciaridae) found in association with cultivated plants and mushrooms in Australia, with notes on cosmopolitan pest species and biosecurity interceptions. Zootaxa 2018, 4415, 201–242. [Google Scholar] [CrossRef]
- Zhimulev, I.F.; Belyaeva, E.S.; Semeshin, V.F.; Koryakov, D.E.; Demakov, S.A.; Demakova, O.V.; Pokholkova, G.V.; Andreyeva, E.N. Polytene chromosomes: 70 years of genetic research. Int. Rev. Cytol. 2004, 241, 203–275. [Google Scholar] [CrossRef] [PubMed]
- Balbiani, E.G. Sur la structure du noyau des cellules salivaires chez les larves de Chironomus. Zool. Anz. 1881, 4, 662–667. [Google Scholar]
- Alfert, M. Composition and structure of giant chromosomes. Int. Rev. Cytol. 1954, 3, 131–169. [Google Scholar] [CrossRef]
- Gerbi, S.A. Non-random chromosome segregation and chromosome eliminations in the fly Bradysia (Sciara). Chromosome Res. 2022, 30, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Lara, F.J.; Stocker, A.J.; Amabis, J.M. DNA sequence amplification in sciarid flies: Results and perspectives. Braz. J. Med. Biol. Res. 1991, 24, 233–248. [Google Scholar] [PubMed]
- Stocker, A.J.; Yokosawa, J.; Soares, M.A.; Cadavid, E.O. DNA replication and amplification during the final cycle of politeny in sciarid gland chromosomes and their control by ecdysone. Ciência Cult. 1996, 48, 306–312. [Google Scholar]
- Gerbi, S.A.; Strezoska, Z.; Waggener, M.J. Initiation of DNA replication in multicellular eukaryotes. J. Struct. Biol. 2003, 140, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.R.; Siviero, F.; Monesi, N. Beyond DNA puffs: What can we learn from studying sciarids? Genesis 2016, 54, 361–378. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.M.; Foulk, M.S.; Casella, C.; Gerbi, S.A. The hunt for origins of DNA replication in multicellular eukaryotes. F1000Prime Rep. 2015, 7, 30. [Google Scholar] [CrossRef]
- Monesi, N.; Candido-Silva, J.A.; Paçó-Larson, M.L.; de Almeida, J.C. Chapter 6. Regulation of sciarid DNA puffs by ecdysone: Mechanisms and perspectives. In Ecdysone: Structures and Funtions; Smagghe, G., Ed.; Springer Science & Business Media B.V.: Dordrecht, The Netherlands, 2009; pp. 165–183. [Google Scholar] [CrossRef]
- Amabis, J.M.; Gorab, E. Dramatic nucleolar dispersion in the salivary gland of Schwenkfeldina sp. (Diptera: Sciaridae). Sci. Rep. 2021, 11, 8347. [Google Scholar] [CrossRef]
- de Cassia Santos Przepiura, T.; Navarro, A.M.; da Rosa Ribeiro, R.; Gomes, J.R.; Pitthan, K.V.; de Miranda Soares, M.A. Mechanisms of programmed cell death in the midgut and salivary glands from Bradysia hygida (Diptera: Sciaridae) during pupal-adult metamorphosis. Cell Biol. Int. 2020, 44, 1981–1990. [Google Scholar] [CrossRef]
- Chen, C.; Shi, X.; Desneux, N.; Han, P.; Gao, X. Detection of insecticide resistance in Bradysia odoriphaga Yang et Zhang (Diptera: Sciaridae) in China. Ecotoxicology 2017, 26, 868–875. [Google Scholar] [CrossRef]
- Hodson, C.N.; Ross, L. Evolutionary perspectives on germline-restricted chromosomes in flies (Diptera). Genome Biol. Evol. 2021, 13, evab072. [Google Scholar] [CrossRef]
- Sauaia, H.; Alves, M.A.R. A description of a new species of Bradysia (Diptera, Sciaridae). Papéis Avulsos Zool. 1968, 22, 85–88. [Google Scholar]
- Trinca, V.; Carli, S.; Cardoso Uliana, J.V.; Garbelotti, C.V.; Mendes da Silva, M.; Kunes, V.; Parras Meleiro, L.; Pereira Brancini, G.T.; Menzel, F.; Moura Andrioli, L.P.; et al. Biocatalytic potential of Pseudolycoriella CAZymes (Sciaroidea, Diptera) in degrading plant and fungal cell wall polysaccharides. iScience 2023, 26, 106449. [Google Scholar] [CrossRef] [PubMed]
- Bravo, I.S.J.; Alves, M.A.R.; Zucoloto, F.S.; Andrade, L.A.M. Aspectos alimentares e de criação de Bradysia hygida Sauaia & Alves (Diptera, Sciaridae) em laboratório. Rev. Bras. Zool. 1993, 10, 343–353. [Google Scholar] [CrossRef]
- de Lima Francisco, A.; Zucoloto, F.S. Obtenção de uma dieta artificial para Bradysia hygida (Diptera, Sciaridae). Rev. Bras. Zool. 1985, 2, 383–386. [Google Scholar] [CrossRef]
- Tramonte, V.L.C.G.; Zucoloto, F.S. Estudos de carboidratos, como alimento para Bradysia hygida (Diptera, Sciaridae). Ciência Cult. 1988, 40, 273–275. [Google Scholar]
- de Almeida, J.C.; Sauaia, H.; Viana, J.C. 5-Bromo-2̕-deoxyuridine induces visible morphological alteration in the DNA puffs of the anterior salivary gland region of Bradysia hygida (Diptera, Sciaridae). Braz. J. Med. Biol. Res. 2010, 43, 1143–1152. [Google Scholar] [CrossRef]
- Ribeiro, W.R. Efeito do 2,3 Dihidro-1H Imidazo (1,2-b) Pirazol (IMPY) Sobre o Desenvolvimento dos Pufes de DNA de Bradysia hygida. Master’s Thesis, Faculdade de Medicina de Ribeirão Prêto, Ribeirão Preto, Brazil, 1975; pp. 1–58. [Google Scholar]
- Sauaia, H.; Laicine, E.M.; Alves, M.A.R. Hydroxyurea-induced inhibition of DNA puff development in the salivary gland chromosomes of Bradysia hygida. Chromosoma 1971, 34, 128–151. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, J.C. A 28-fold increase in secretory protein synthesis is associated with DNA puff activity in the salivary gland of Bradysia hygida (Diptera, Sciaridae). Braz. J. Med. Biol. Res. 1997, 30, 605–614. [Google Scholar] [CrossRef]
- Laicine, E.M.; Alves, M.A.R.; de Almeida, J.C.; Rizzo, E.; Albernaz, W.C.; Sauaia, H. Development of DNA puffs and patterns of polypeptide synthesis in the salivary glands of Bradysia hygida. Chromosoma 1984, 89, 280–284. [Google Scholar] [CrossRef]
- Borges, A.R.; Pinatto Gaspar, V.; Fernandez, M.A. Unequal X chromosomes in Bradysia hygida (Diptera: Sciaridae) females: Karyotype assembly and morphometric analysis. Genetica 2000, 108, 101–105. [Google Scholar] [CrossRef]
- Pinatto Gaspar, V.; Borges, A.R.; Fernandez, M.A. NOR sites detected by Ag-dAPI staining of an unusual autosome chromosome of Bradysia hygida (Diptera: Sciaridae) colocalize with C-banded heterochromatic region. Genetica 2002, 114, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Fontes, A.M.; Conacci, M.E.; Monesi, N.; de Almeida, J.C.; Paçó-Larson, M.L. The DNA puff BhB10-1 gene encodes a glycine-rich protein secreted by the late stage larval salivary glands of Bradysia hygida. Gene 1999, 231, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Monesi, N.; Silva, L.A.; Martins, P.C.M.; Teixeira, A.B.; Dornelas, E.C.; Moreira, J.E.; Paçó-Larson, M.L. Immunocharacterization of the DNA puff BhC4-1 protein of Bradysia hygida (Diptera: Sciaridae). Insect Biochem. Mol. Biol. 2004, 34, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Trinca, V.; Cardoso Uliana, J.V.; Sousa Ribeiro, G.K.; Teixeira Torres, T.; Monesi, N. Characterization of the mitochondrial genomes of Bradysia hygida, Phytosciara flavipes and Trichosia splendens (Diptera: Sciaridae) and novel insights on the control region of sciarid mitogenomes. Insect Mol. Biol. 2022, 31, 482–496. [Google Scholar] [CrossRef]
- Sauaia, H. Cromossomas Politȇnicos de Bradysia hygida. Inibição do Desenvolvimento dos Puffs de DNA Pela Hydroxiuréia. Ph.D. Thesis, Universidade de São Paulo, Faculdade de Medicina de Ribeirão Prêto, Departamento de Morfologia, Ribeirão Prêto, Brazil, 1971; pp. 1–53. [Google Scholar]
- Basso, L.R.; Vasconcelos, C.; Fontes, A.M.; Hartfelder, K.; Silva, J.A.; Coelho, P.S.R.; Monesi, N.; Paçó-Larson, M.L. The induction of DNA puff BhC4-1 gene is a late response to the increase in 20-hydroxyecdysone titers in last instar dipteran larvae. Mech. Dev. 2002, 110, 15–26. [Google Scholar] [CrossRef]
- Candido-Silva, J.A.; de Carvalho, D.P.; Coelho, G.R.; de Almeida, J.C. Indirect immune detection of ecdysone receptor (EcR) during the formation of DNA puffs in Bradysia hygida (Diptera, Sciaridae). Chromosome Res. 2008, 16, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Cardoso Uliana, J.V.; Pereira Brancini, G.T.; Castelli-Gair Hombría, J.; Digiampietri, L.A.; Andrioli, L.P.; Monesi, N. Characterizing the embryonic development of B. hygida (Diptera: Sciaridae) following enzymatic treatment to permeabilize the serosal cuticle. Mech. Dev. 2018, 154, 270–276. [Google Scholar] [CrossRef]
- da Conceição Silva, J.L.; Fernandez, M.A. Feulgen stain in heterogeneous egg samples from non-synchronized Bradysia hygida (Diptera: Sciaridae) laboratory culture. Cytologia 2000, 65, 167–171. [Google Scholar] [CrossRef]
- Amabis, J.M.; Reinach, F.C.; Andrews, N. Spermatogenesis in Trichosia pubescens (Diptera: Sciaridae). J. Cell Sci. 1979, 63, 199–213. [Google Scholar] [CrossRef]
- Madalena, C.R.G.; Amabis, J.M.; Stocker, A.J.; Gorab, E. The localization of ribosomal DNA in Sciaridae (Diptera: Nematocera) reassessed. Chromosome Res. 2007, 15, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Stocker, A.J.; Gorab, E. Local enrichment with homopolymeric (DA/DT) DNA in genomes of some lower dipterans and Drosophila melanogaster. Chromosoma 2003, 111, 455–460. [Google Scholar] [CrossRef]
- Gabrusewycz-Garcia, N. Studies in polytene chromosomes of sciarids. I. The salivary chromosomes of Sciara (Lycoriella) pauciseta (II), Felt. Chromosoma 1971, 33, 421–435. [Google Scholar] [CrossRef]
- Escribá, M.C.; Greciano, P.G.; Méndez-Lago, M.; de Pablos, B.; Trifonov, V.A.; Ferguson-Smith, M.A.; Goday, C.; Villasante, A. Molecular and cytological characterization of repetitive DNA sequences from the centromeric heterochromatin of Sciara coprophila. Chromosoma 2011, 120, 387–397. [Google Scholar] [CrossRef]
- Pinatto Gaspar, V.; Shimauti, E.L.T.; Fernandez, M.A. Chromosomal localization and partial sequencing of the 18S and 28S ribosomal genes from Bradysia hygida (Diptera: Sciaridae). Genet. Mol. Res. 2014, 13, 2177–2185. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270 (Suppl. S1), 96–99. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.T.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Prous, M.; Heidemaa, M.; Soon, V. Empria longicornis species group: Taxonomic revision with notes on phylogeny and ecology (Hymenoptera, Tenthredinidae). Zootaxa 2011, 2756, 1–39. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.K.; Crosby, T.K. The Preparation and Curation of Insects; DSIR Information Series, New Revised Edition; Science Information Publishing Centre: Wellington, New Zealand, 1988; Volume 163, pp. 1–91. [Google Scholar]
- Menzel, F.; Mohrig, W. Revision der paläarktischen Trauermücken (Diptera: Sciaridae). Stud. Dipterol. Suppl. 2000, 6, 1–761. [Google Scholar]
- Broadley, A.; Kauschke, E.; Mohrig, W. Revision of the black fungus gnat species (Diptera: Sciaridae) described by W. A. Steffan from Micronesia. Zootaxa 2019, 4683, 215–241. [Google Scholar] [CrossRef] [PubMed]
- Heller, K. A new species of the genus Pseudolycoriella (Diptera: Sciaridae) bred from an ornamental plant. Heteropterus Rev. Entomol. 2012, 12, 195–199. [Google Scholar]
- Köhler, A.; Mohrig, W. Additions to the New Zealand fauna of black fungus gnats (Diptera: Sciaridae), with descriptions of six new species. N. Z. Entomol. 2016, 39, 91–109. [Google Scholar] [CrossRef]
- Menzel, F.; Mohrig, W. Beiträge zur Taxonomie und Faunistik der paläarktischen Trauermücken (Diptera, Sciaridae). Teil VI—Neue Ergebnisse aus Typenuntersuchungen und die daraus resultierenden taxonomisch-nomenklatorischen Konsequenzen. Stud. Dipterol. 1998, 5, 351–378. [Google Scholar]
- Menzel, F.; Smith, J.E. Family Sciaridae. In The Diptera of the Seychelles Islands; Pensoft Series Faunistica; Gerlach, J., Ed.; Pensoft Publisher: Moscow, Russia, 2009; Volume 85, pp. 19–45. [Google Scholar]
- Mohrig, W. Black fungus gnats of Central America. Part I. (Diptera, Sciaridae). Beitr. Entomol. 2003, 53, 1–69. [Google Scholar] [CrossRef]
- Mohrig, W. Die Trauermücken (Diptera: Sciaridae) von Papua-Neuguinea. Teil III—Gattungen Ctenosciara und Pseudolycoriella. Stud. Dipterol. 2013, 20, 123–168. [Google Scholar]
- Mohrig, W.; Jaschhof, M. Sciarid flies (Diptera, Sciaridae) of New Zealand. Stud. Dipterol. Suppl. 1999, 7, 1–101. [Google Scholar]
- Mohrig, W.; Menzel, F. 16. Sciaridae (Black Fungus Gnats). In Manual of Central American Diptera, Volume 1; Brown, B.V., Borkent, A., Cumming, J.M., Wood, D.M., Woodley, N.E., Zumbado, M.A., Eds.; NRC Research Press: Ottawa, ON, Canada, 2009; pp. 279–292. [Google Scholar]
- Mohrig, W.; Röschmann, F.; Rulik, B. The fauna of sciarid flies from the Dominican Republic (Diptera, Sciaridae). Beitr. Entomol. 2004, 54, 267–331. [Google Scholar] [CrossRef]
- Mohrig, W.; Kauschke, E.; Broadley, A. Pseudolycoriella skusei sp. nov. (Diptera: Sciaridae), a new dark-winged fungus gnat from Norfolk Island and Australia. Zootaxa 2016, 4097, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Mohrig, W.; Kauschke, E.; Broadley, A. New black fungus gnats (Diptera: Sciaridae) from Eastern Australia. Zootaxa 2018, 4450, 203–241. [Google Scholar] [CrossRef] [PubMed]
- Mohrig, W.; Kauschke, E.; Broadley, A. Revision of black fungus gnat species (Diptera, Sciaridae) described from the Hawaiian Islands by D.E. Hardy and W.A. Steffan, and a contribution to the knowledge of the sciarid fauna of the Galápagos Islands. Zootaxa 2019, 4590, 404–439. [Google Scholar] [CrossRef] [PubMed]
- Mohrig, W.; Kauschke, E.; Broadley, A. Black fungus gnats (Diptera: Sciaridae) of Queensland, Australia. Part II. Genus Pseudolycoriella Menzel & Mohrig, 1998. Zootaxa 2020, 4751, 487–506. [Google Scholar] [CrossRef]
- Rudzinski, H.-G. Neue Trauermücken aus Afrika (Diptera: Sciaridae). Entomol. Z. 1997, 107, 160–172. [Google Scholar]
- Rudzinski, H.-G. Neue Arten der Gattung Pseudolycoriella Menzel & Mohrig, 1998 aus Afrika (Diptera Nematocera: Sciaridae). Entomofauna 2003, 24, 97–119. [Google Scholar]
- Rudzinski, H.-G.; Baumjohann, K.; Wolff, M. Pseudolycoriella martita sp. nov.: The first species of the genus Pseudolycoriella Menzel & Mohrig, 1998 from Colombia. Mitteilungen Int. Entomol. Ver. E. V. 2016, 41, 5–9. [Google Scholar]
- Vilkamaa, P.; Hippa, H.; Mohrig, W. The genus Pseudolycoriella Menzel & Mohrig (Diptera, Sciaridae) in New Caledonia, with the description of thirteen new species. Zootaxa 2012, 3207, 1–21. [Google Scholar] [CrossRef]
- Winnertz, J. Beitrag zu einer Monographie der Sciarinen; W. Braumüller: Wien, Austria, 1867; pp. 1–187. [Google Scholar]
- Hippa, H.; Vilkamaa, P.; Heller, K. Review of the Holarctic Corynoptera Winnertz, 1867, s. str. (Diptera, Sciaridae). Zootaxa 2010, 2695, 1–197. [Google Scholar] [CrossRef]
- de Souza Amorim, D. A catalogue of the family Sciaridae (Diptera) of the Americas South of the United States. Rev. Bras. Entomol. 1992, 36, 55–77. [Google Scholar]
- Amabis, D.; Amabis, J.M.; Simões, L.C.G. Puffing activity in the salivary gland chromosomes of Rhynchosciara under experimental conditions. Chromosoma 1977, 62, 139–154. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, H.S.L.; Cruz, A.R. The allometric relationship between nucleus and cytoplasm in the salivary glands of Bradysia hygida. Can. J. Zool. 1983, 61, 1311–1316. [Google Scholar] [CrossRef]
- Laicine, E.M.; Fernandez, M.A.; Sauaia, H. Acid phosphatase activity in mature secretory granules of the salivary gland of Bradysia hygida. J. Morphol. 1991, 208, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Paçó-Larson, M.L.; de Almeida, J.C.; Edström, J.-E.; Sauaia, H. Cloning of a developmentally amplified gene sequence in the DNA puff C4 of Bradysia hygida (Diptera: Sciaridae) salivary glands. Insect Biochem. Mol. Biol. 1992, 22, 439–446. [Google Scholar] [CrossRef]
- Fontes, A.M.; de Almeida, J.C.; Edstrom, J.-E.; Paçó-Larson, M.L. Cloning of a B10 DNA puff sequence developmentally amplified and expressed in the salivary gland of Bradysia hygida. Braz. J. Med. Biol. Res. 1992, 25, 777–780. [Google Scholar]
- Coelho, P.S.R.; Monesi, N.; de Almeida, J.C.; Toledo, F.; Buttin, G.; Paçó-Larson, M.L. DNA puff C4 of Bradysia hygida (Diptera: Sciaridae) contains genes unequally amplified and differentially expressed during development. Chromosome Res. 1993, 1, 121–126. [Google Scholar] [CrossRef]
- Monesi, N.; Fernandez, M.A.; Fontes, A.M.; Basso, L.R.; Nakanishi, Y.; Baron, B.; Buttin, G.; Paçó-Larson, M.L. Molecular characterization of an 18 kb segment of DNA puff C4 of Bradysia hygida (Diptera, Sciaridae). Chromosoma 1995, 103, 715–724. [Google Scholar] [CrossRef]
- Fontes, A.M. Clonagem e Caracterizacão da Estrutura e Expressão de um Gene Amplificado no Pufe de DNA B-10 de Bradysia hygida (Diptera, Scaridae). PhD Dissertation, Universidade de São Paulo, Faculdade de Medicina de Ribeirão Preto, Ribeirão Preto, Brazil, 1996; pp. 1–105. [Google Scholar]
- Penalva, L.O.F.; Yokosawa, J.; Stocker, A.J.; Soares, M.A.M.; Graessmann, M.; Orlando, T.C.; Winter, C.E.; Botella, L.M.; Graessmann, A.; Lara, F.J.S. Molecular characterization of the C-3 DNA puff gene of Rhynchosciara americana. Gene 1997, 193, 163–172. [Google Scholar] [CrossRef]
- Stocker, A.J.; Amabis, J.M.; Gorab, E.; Elke, C.; Lezzi, M. Antibodies against the D-domain of a Chironomus ecdysone receptor protein react with DNA puff sites in Trichosia pubescens. Chromosoma 1997, 106, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Monesi, N.; Jacobs-Lorena, M.; Paçó-Larson, M.L. The DNA puff gene BhC4-1 of Bradysia hygida is specifically transcribed in early prepupal salivary glands of Drosophila melanogaster. Chromosoma 1998, 107, 559–569. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, D.P.; Coelho, P.S.; de Almeida, J.C. A dual role of 20-hydroxyecdysone in the control of protein synthesis related to DNA puff activity in the anterior region of Bradysia hygida (Diptera, Sciaridae) salivary gland. Insect Biochem. Mol. Biol. 2000, 30, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Monesi, N.; de Freitas Sousa, J.; Paçó-Larson, M.L. The DNA puff BhB10-1 gene is differentially expressed in various tissues of Bradysia hygida late larvae and constitutively transcribed in transgenic Drosphila. Braz. J. Med. Biol. Res. 2001, 34, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Conacci, M.E.; Coelho, P.S.R.; Valente, V.; de Freitas Sousa, J.; Paçó-Larson, M.L.; de Almeida, J.C. The control of BhB10-1 gene expression in the salivary gland of Bradysia hygida (Diptera, Sciaridae) is disrupted in vivo by a delayed effect of cycloheximide. Insect Biochem. Mol. Biol. 2002, 32, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Shimauti, E.L.T.; Fernandez, M.A. In situ restriction endonucleases digestion by on the nucleolar organizer region (NOR) in neuroblast mitotic chromosomes of Bradysia hygida (Diptera: Sciaridae). Cytologia 2002, 67, 347–354. [Google Scholar] [CrossRef]
- Soares, M.A.M.; Monesi, N.; Basso, L.R.; Stocker, A.J.; Paçó-Larson, M.L.; Lara, F.J.S. Analysis of the amplification and transcription of the C3-22 gene of Rhynchosciara americana (Diptera: Sciaridae) in transgenic lines of Drosophila melanogaster. Chromosoma 2003, 112, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.F.C.; Fernandez, M.A. Molecular characterization of the 5S ribosomal gene of the Bradysia hygida (Diptera: Sciaridae). Genetica 2004, 122, 253–260. [Google Scholar] [CrossRef]
- Santelli, R.V.; Siviero, F.; Machado-Santelli, G.M.; Lara, F.J.S.; Stocker, A.J. Molecular characterization of the B-2 DNA puff gene of Rhynchosciara americana. Chromosoma 2004, 113, 167–176. [Google Scholar] [CrossRef]
- Simon, C.R.; de Almeida, J.C. Programmed cell death in Bradysia hygida (Diptera, Sciaridae) salivary glands presents apoptotic features. Genesis 2004, 40, 22–31. [Google Scholar] [CrossRef]
- Ferreira Saccuti, C.; de Miranda Soares, M.A.; Penteado Falco, J.R.; Fernandez, M.A. Genome size of three Brazilian flies from the Sciaridae family. Genet. Mol. Biol. 2005, 38, 743–748. [Google Scholar] [CrossRef]
- Polinarski, C.A.; da Conceição Silva, J.L.; Mikami, L.R.; Fernandez, M.A. Nuclear halo from Bradysia hygida (Diptera: Sciaridae) salivary gland polytene cells. Braz. Arch. Biol. Technol. 2005, 48, 351–357. [Google Scholar] [CrossRef]
- de Souza Gouveia, F. Análise da Estrutura e Mapeamento de Sítios de DNA Bent em Promotores de Genes de Secreção em Insetos. Master’s Thesis, Universidade Estadual de Maringá, Maringá, Brazil, 2006; pp. 1–47. [Google Scholar]
- Simon, C.R.; Pinheiro Ramos, R.G. Death by fine tunning. Braz. J. Morphol. Sci. 2006, 23, 3–13. [Google Scholar]
- Hayashi, B.; Hoff, M.M.; Fernadez, M.A.; Fiorini, A. Avaliação da curvatura intrínseca de regiões MARs, detectadas in silico, pertencentes à região amplificada do gene BHC4-1. In V Encontro Internacional de Produção Científica Cesumar, 23 a 26 de outubro de 2007; CESUMAR—Centro Universitário de Maringá: Maringá, Brazil, 2007; Available online: https://rdu.unicesumar.edu.br/handle/123456789/6492 (accessed on 5 September 2023).
- Lecci, M.S.; Malta, T.M.; Flausino, V.T.; Gitaí, D.L.G.; Ruiz, J.C.; Monesi, N. Functional and bioinformatics analyses reveal conservation of cis-regulatory elements between Sciaridae and Drosophilidae. Genesis 2008, 46, 43–51. [Google Scholar] [CrossRef]
- Zanarotti, G.M.; Candido-Silva, J.A.; de Almeida, J.C. BhSGAMP-1, a gene that encodes an antimicrobial peptide, is developmentally regulated by the direct action of 20-OH ecdysone in the salivary gland of Bradysia hygida (Diptera, Sciaridae). Genesis 2009, 47, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Candido-Silva, J.A.; Monesi, N. Bradysia hygida (Diptera, Sciaridae) presents two eukaryotic Elongation Factor 1A gene homologues: Partial characterization of the eukaryotic Elongation Factor 1A-F1 gene. Braz. J. Med. Biol. Res. 2010, 43, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Francischini Rodrigues, L.; Ribeiro Passos, K.J.; Togoro, S.Y.; Fernandez, M.A. Obtenção de clones de Bradysia hygida em cromossomo artificial bacteriano (BAC). In Anais do XIX EAIC, 28 a 30 de Outubro de 2010; Universidade Estadual do Centro-Oeste, Unicentro: Guarapuava, PR, Brazil, 2010; Available online: https://anais.unicentro.br/xixeaic/pdf/1180.pdf (accessed on 5 September 2023).
- Wang, J.; Gao, Q.; Liu, Q.; Xiang, L.; Wang, Y.; Wang, D. Expression of a modified antimicrobial peptide BhSGAMP-1-S (Bradysia hygida) in Escherichia coli and characterization of its activity. Acta Microbiol. Sin. 2010, 50, 1185–1193. [Google Scholar]
- de Oliveira Gaio, A.; Rodrigues, R.C.C.; do Nascimento, C.; Secundino, N.F.C.; Lemos, F.J.A.; Pimenta, P.F.P.; Monesi, N. Use of the checkerboard DNA-DNA hybridization technique for bacteria detection in Aedes aegypti (Diptera: Culicidae) (L.). Parasites Vectors 2011, 4, 237. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.C.; Gitaí, D.L.G.; Humann, F.C.; Paçó-Larson, M.L.; Monesi, N. Functional characterization of the sciarid BhC4-1 core promoter in transgenic Drosophila. BMC Mol. Biol. 2011, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro Passos, K.J.; Togoro, S.Y.; Carignon, S.; Koundrioukoff, S.; Lachages, A.-M.; Debatisse, M.; Fernandez, M.A. Molecular combing in the analysis of developmentally regulated amplified segments of Bradysia hygida. Genet. Mol. Res. 2012, 11, 2060–2070. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.; Berry, D.L.; Kary, C.S. Insect Histology: Practical Laboratory Techniques, 1st ed.; John Wiley & Sons Ltd.: Oxford, UK, 2015; pp. 1–380. [Google Scholar] [CrossRef]
- de Moraes, A.P.R.; de Salles, C.M.C.; Bittencourt, V.R.E.P.; Bittencourt, A.J. Antimicrobial activity of Stomoxys calcitrans against Beauveria bassiana sensu lato isolates. Rev. Bras. Parasitol. Vet. 2015, 24, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Candido-Silva, J.A.; Machado, M.C.R.; Hartfelder, K.H.; de Almeida, J.C.; Paçó-Larson, M.L.; Monesi, N. Amplification and expression of a salivary gland DNA puff gene in the prothoracic gland of Bradysia hygida (Diptera: Sciaridae). J. Insect Physiol. 2015, 74, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.A.M.; Hartfelder, K.; Tesserolli de Souza, J.M.; Stocker, A.J. Developmental ecdysteroid titers and DNA puffs in larvae of two sciarid species, Rhynchosciara americana and Rhynchosciara milleri (Diptera: Sciaridae). Genetica 2015, 143, 597–612. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-H.; Zhai, Y.-F.; Cao, X.; Zhou, X.-H.; Wang, Z.-Y.; Wang, F.; Chen, H.; Yu, Y. Feeding and related morphology of salivary glands of Bradysia odoriphaga (Diptera: Sciaridae). J. Asia-Pac. Entomol. 2016, 19, 319–325. [Google Scholar] [CrossRef]
- Frank, H.O.; Garcia Sanchez, D.; de Freitas Oliveira, L.; Kobarg, J.; Monesi, N. The Drosophila melanogaster Eip74EF-PA transcription factor directly binds the sciarid BhC4-1 promoter. Genesis 2017, 55, e23075. [Google Scholar] [CrossRef]
- Cachay Wester, J.V.W.C.; Couto Lima, C.A.; Machado, M.C.R.; Zampar, P.V.; Sakagute Tavares, S.; Monesi, N. Characterization of a novel Drosophila melanogaster cis-regulatory module that drives gene expression to the larval tracheal system and adult thoracic musculature. Genesis 2018, 56, e23222. [Google Scholar] [CrossRef]
- Frank, H.O. Genes Responsivos ao Hormônio Ecdisona em Bradysia hygida (Diptera, Sciaridae). Ph.D. Thesis, Universidade de São Paulo, Faculdade de Medicina de Ribeirão Preto, Ribeirão Preto, Brazil, 2019; pp. 1–53. [Google Scholar]
- Banho, N.N.; Pattero, J.R.; Fernandez, M.A. Análise quantitativa da expressão do gene KIN em glândulas salivares de Bradysia hygida no 4º instar larval. In Proceedings of the 30º Encontro Anual de Iniciação Científica (EAIC) & 10º Encontro Anual de Iniciação Científica Júnior (EAIC-Jr), Universidade Estadual de Maringá, Campus Universitário, Maringá, Brazil, 11–12 November 2021; Volume 5. Available online: https://www.eaic.uem.br/eaic2021/anais/artigos/5223.pdf (accessed on 5 September 2023).
- Noeske, V. Single Genes Initiate Novel Epithelial Structures in Early Fly Development. Ph.D. Thesis, Combined Faculties for the Natural Sciences and Mathematics, Ruperto-Carola University of Heidelberg, Heidelberg, Germany, 2021; pp. 1–139. [Google Scholar]
- BOLD Systems. Barcode of Life Data System, Version 4. 2023. Available online: https://v4.boldsystems.org (accessed on 19 April 2023).
- Metz, C.W. Structure of the salivary gland chromosomes in Sciara. J. Hered. 1935, 26, 177–188. [Google Scholar] [CrossRef]
- Crouse, H.V.; Smith-Stocking, H. New mutants in Sciara and their genetic behavior. Genetics 1938, 23, 275–282. [Google Scholar] [CrossRef]
- Martín, I.; Ruiz, M.F.; Sánchez, L. The gene transformer-2 of Sciara (Diptera, Nematocera) and its effect on Drosophila sexual development. BMC Dev. Biol. 2011, 11, 19. [Google Scholar] [CrossRef]
- Crouse, H.V. Translocations in Sciara; their bearing on chromosome behavior, and sex determination. Res. Bull. Univ. Mo. Coll. Agric. Agric. Exp. Stn. 1943, 379, 1–75. [Google Scholar]
- Harris, M.A.; Gardner, W.A.; Oetting, R.D. A review of the scientific literature on fungus gnats (Diptera: Sciaridae) in the genus Bradysia. J. Entomol. Sci. 1996, 31, 252–276. [Google Scholar] [CrossRef]
- Blaschke-Berthold, U. Larval taxonomy in Sciaridae. Verh. Naturwissenschaftlichen Ver. Hambg. Neue Folge 1988, 30, 345–351. [Google Scholar]
- Krivosheina, N.P.; Zaitzev, A.I.; Mohrig, W.K. K biologii dvukrylykh detritniz (Diptera, Sciaridae) [On the biology of two-winged detritus (Diptera, Sciaridae)]. In Ekologiya i Morfologiya Nasekomykh—Obitatelei Gribnykh Substratov [Ecology and Morphology of Insects—Inhabitants of Fungal Substrates]; Pravdin, F.N., Ed.; Nauka: Moscow, Russia, 1987; pp. 6–24. (In Russian) [Google Scholar]
- Menzel, F.; Mohrig, W. 2.6. Family Sciaridae. In Contributions to a Manual of Palaearctic Diptera (with special reference to flies of economic importance). Volume 2: Nematocera and lower Brachycera; Papp, L., Darvas, B., Eds.; Science Herald: Budapest, Hungary, 1997; pp. 51–69. [Google Scholar]
- Steffan, W.A. A generic revision of the family Sciaridae (Diptera) of America north of Mexico. Univ. Calif. Publ. Entomol. 1966, 44, 1–77. [Google Scholar]
- Steffan, W.A. 15. Sciaridae. In Manual of Nearctic Diptera, Volume 1; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Research Branch Agriculture Canada: Ottawa, ON, Canada, 1981; Volume 27, pp. 247–255. [Google Scholar]
- Sutou, M.; Ito, M.T. Occurrence of the genus Scythropochroa (Insecta: Diptera: Sciaridae) in Japan, with descriptions of the female and larva. Species Divers. 2004, 9, 375–381. [Google Scholar] [CrossRef]
- Sutou, M.; Kato, T.; Ito, M. Description of the final larval stage and the pupa of Ctenosciara japonica (Diptera: Sciaridae) and their DNA barcodes. Stud. Dipterol. 2007, 14, 17–22. [Google Scholar]
- Steffan, W.A. Laboratory studies of Hawaiian Sciaridae (Diptera). Int. Biol. Program Tech. Rep. 1973, 25, 1–17. [Google Scholar]
- Gabrusewycz-Garcia, N. Cytological and autoradiographic studies in Sciara coprophila salivary gland chromosomes. Chromosoma 1964, 15, 312–344. [Google Scholar] [CrossRef]
- Breuer, M.E. Cromossomas politênicos das glândulas salivares de “Rhynchosciara angelae” Nonato & Pavan, 1951 (Diptera, Sciaridae). Rev. Bras. Biol. 1967, 27, 105–108. [Google Scholar]
- Amabis, J.M. The polytene chromosomes of the salivary gland of Trichosia pubescens (Diptera: Sciaridae). Rev. Bras. Genet. 1983, 6, 415–424. [Google Scholar]
- da Cunha, P.R.; Granadino, B.; Perondini, A.L.P.; Sanchez, L. Dosage compensation in sciarids is achieved by hypertranscription of the single X chromosome in males. Genetics 1994, 138, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Metz, C.W. Chromosome behavior in Sciara (Diptera). In Program and Abstracts for the Twenty-Third Annual Meeting of the American Society of Zoölogists, New Haven, Connecticut, December 28–30, 1925; American Society of Zoölogists, Ed. Anat. Rec. 1925, 31, 346–347. [Google Scholar]
- Metz, C.W. Chromosomes and sex in Sciara. Science 1925, 61, 212–214. [Google Scholar] [CrossRef]
- Ye, L.; Leng, R.; Huang, J.; Qu, C.; Wu, H. Review of three black fungus gnat species (Diptera: Sciaridae) from greenhouses in China: Three greenhouse sciarids from China. J. Asia-Pac. Entomol. 2017, 20, 179–184. [Google Scholar] [CrossRef]
- Roberts, E.C.; Lavigne, R. Fungus gnat found to inhibit development of turfgrass. USGA J. Turf. Manag. 1959, 12, 25–26. [Google Scholar]
- Shaw, F.R. A review of some of the more important contributions to our knowledge of the systematic relationships of the Sciaridae (Diptera). Proc. Hawaii Entomol. Soc. 1953, 15, 25–32. [Google Scholar]
- Tuomikoski, R. Zur Kenntnis der Sciariden (Dipt.) Finnlands. Ann. Zool. Soc. Zool. Bot. Fenn. Vanamo 1960, 21, 1–164. [Google Scholar]
- Steffan, W.A. Notes on the synonymy of Bradysia tritici (Coquillett) and Lycoriella mali (Fitch) (Diptera: Sciaridae). Pac. Insects 1965, 7, 290. [Google Scholar]
- Menzel, F.; Heller, K. Bemerkungen zur Nomenklatur der Sciariden (Diptera, Bibionomorpha: Sciaridae). Stud. Dipterol. 2007, 13, 209–229. [Google Scholar]
- Stone, A.; Laffoon, J.L. Family Sciaridae (Lycoriidae). In A Catalog of the Diptera of America North of Mexico; Stone, A., Sabrosky, C.W., Wirth, W.W., Foote, R.H., Coulson, J.R., Eds.; United States Department of Agriculture: Washington, DC, USA, 1965; Volume 276, pp. 229–236. [Google Scholar]
- Breuer, M.E. Revision of the genus Rhynchosciara Rübsaamen (Diptera, Sciaridae) in the Neotropical region. Arq. Zool. 1969, 17, 167–198. [Google Scholar] [CrossRef]
- Brito da Cunha, A.; Morgante, J.S.; Pavan, C.; Garrido, M.C.; Marques, J. Studies on cytology and differentiation in Sciaridae IV. Nuclear and cytoplasmatic differentiation in salivary glands of Bradysia elegans (Diptera, Sciaridae). Caryologia 1973, 26, 83–100. [Google Scholar] [CrossRef]
- Brito da Cunha, A.; Riess, R.W.; Pavan, C.; Biesele, J.J.; Morgante, J.S. & Garrido, M.C. Studies on cytology and differentiation in Sciaridae. 5. Electron microscopic studies on the salivary glands of Bradysia elegans Morgante (Diptera, Sciaridae). Differentiation 1973, 1, 145–166. [Google Scholar] [CrossRef]
- Brito da Cunha, A.; Morgante, J.S.; Pavan, C.; Garrido, M.C.I. Studies on cytology and differentiation in Sciaridae. III. Nuclear and cytoplasmatic differentiation in the salivary glands of Bradysia sp. Stud. Genet. 1969, 6918, 1–11. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Collecting Site | Sex | GenBank Accession No. | BOLD BIN | BOLD Sample ID | BOLD Sequence ID | Sample Holder |
|---|---|---|---|---|---|---|---|
| Bradysiopsis vittata (Meigen, 1830) | GERMANY: Rhineland-Palatinate | Female | N/A | BOLD:ACC1999 | BIOUG16734-E02 | GMGME1271-14 | ZFMK |
| Camptochaeta camptochaeta (Tuomikoski, 1960) | FINLAND: North Ostrobothnia | N/A | N/A | BOLD:ACB7851 | BIOUG04085-F06 | GMFIC094-12 | CBGC |
| Claustropyga clausa (Tuomikoski, 1960) | NORWAY: Telemark | Male | N/A | BOLD:ACY8611 | bf-sci-00255b | SCINO693-15 | BFCO |
| Diadocidia ferruginosa (Meigen, 1830) | GERMANY: Bavaria | Male | N/A | BOLD:AAY7756 | BC-ZSM-DIP-24049-G09 | AMTPD1981-15 | ZSMC |
| Dichopygina nigrohalteralis (Frey, 1948) | NORWAY: Oppland | Male | KY079365 | BOLD:ACM6664 | bf-sci-00193 | SCINO192-15 | BFCO |
| Hemineurina conspicua (Winnertz, 1867) | NORWAY: Ostfold | Male | N/A | BOLD:AAP4769 | bf-sci-01103 | SCINO1356-16 | BFCO |
| Hemineurina inflata (Winnertz, 1867) | NORWAY: Buskerud | Male | N/A | BOLD:ACJ9929 | bf-sci-01309 | SCINO1340-16 | BFCO |
| Lycoriella sativae (Johannsen, 1912) | NORWAY: Trøndelag | Male | N/A | BOLD:ABA1215 | TRD-Sci038 | SCINO608-15 | NTNU |
| Protoxylosciara longiforceps (Bukowski & Lengersdorf, 1936) | NORWAY: Hordaland | Male | N/A | BOLD:ACC1367 | bf-sci-00889 | SCINO1193-16 | BFCO |
| Pseudolycoriella aotearoa Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906359 | BOLD:AED8133 | SDEI-Dipt-0000997 | GBMNB15388-20 | SDEI |
| Pseudolycoriella bispina Mohrig, 1999 | NEW ZEALAND: North Island | Male | MK906401 | BOLD:AED6042 | SDEI-Dipt-0001522 | GBMNB15430-20 | SDEI |
| Pseudolycoriella bruckii (Winnertz, 1867) | GERMANY: Bavaria | Male | N/A | BOLD:ADW9617 | BIOUG42711-H03 | GMGMP3875-18 | ZSMC |
| Pseudolycoriella brunnea (Bukowski & Lengersdorf, 1936) | GERMANY: Bavaria | Male | N/A | BOLD:ACC1590 | BIOUG55426-H05 | GMGMW520-20 | ZSMC |
| Pseudolycoriella cavatica (Skuse, 1888) | NEW ZEALAND: North Island | Male | MK906395 | BOLD:ABW3602 | SDEI-Dipt-0001411 | GBMNB15424-20 | SDEI |
| Pseudolycoriella compacta Heller, 2000 | GERMANY: Rhineland-Palatinate | Male | N/A | BOLD:AAN6441 | BIOUG16613-B12 | GMGMD750-14 | ZFMK |
| Pseudolycoriella dagae Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906331 | BOLD:AED4998 | SDEI-Dipt-0000655 | GBMNB15360-20 | SDEI |
| Pseudolycoriella frederickedwardsi Köhler, 2016 | NEW ZEALAND: North Island | Male | MK906341 | BOLD:AED3379 | SDEI-Dipt-0000772 | GBMNB15370-20 | SDEI |
| Pseudolycoriella gonotegmenta Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906356 | BOLD:AED8311 | SDEI-Dipt-0000977 | GBMNB15385-20 | NZAC |
| Pseudolycoriella hartmanni (Menzel & Mohrig, 1991) | NORWAY: Hedmark | Male | N/A | BOLD:ACX5047 | TRD-Sci055 | SCINO625-15 | NTNU |
| Pseudolycoriella hauta Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906322 | BOLD:AED1025 | SDEI-Dipt-0000506 | GBMNB15351-20 | SDEI |
| Pseudolycoriella horribilis (Edwards, 1931) | SOUTH KOREA: Jeollanam-do | Male | JQ613788 | BOLD:ACD4315 | 74 (JQ613788) | GBMIN12850-13 | SNUC |
| Pseudolycoriella huttoni Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906327 | BOLD:AED6374 | SDEI-Dipt-0000621 | GBMNB15356-20 | NZAC |
| Pseudolycoriella hygida (Sauaia & Alves, 1968) | BRAZIL: São Paulo | Male | N/A | BOLD:ACW7953 | SDEI-Dipt-0002256 | SSEY004-23 | SDEI |
| Pseudolycoriella hygida (Sauaia & Alves, 1968) | BRAZIL: São Paulo | Female | N/A | BOLD:ACW7953 | SDEI-Dipt-0002255 | SSEY003-23 | SDEI |
| Pseudolycoriella hygida (Sauaia & Alves, 1968) | ARGENTINA: Formosa | Female | N/A | BOLD:ACW7953 | BIOUG23471-G01 | GMAFH202-15 | MACN |
| Pseudolycoriella hygida (Sauaia & Alves, 1968) | ARGENTINA: Formosa | N/A | N/A | BOLD:ACW7953 | BIOUG23277-A10 | GMAFE075-15 | MACN |
| Pseudolycoriella hygida (Sauaia & Alves, 1968) | ARGENTINA: Formosa | N/A | N/A | BOLD:ACW7953 | BIOUG23645-D09 | GMAFL125-15 | MACN |
| Pseudolycoriella japonensis (Mohrig & Menzel, 1992) | NORWAY: Møre og Romsdal | Male | N/A | BOLD:ACZ8590 | bf-sci-00938 | SCINO1241-16 | BFCO |
| Pseudolycoriella jaschhofi Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906409 | BOLD:AED7085 | SDEI-Dipt-0001511 | GBMNB15438-20 | SDEI |
| Pseudolycoriella jejuna (Edwards, 1927) | NEW ZEALAND: North Island | Male | MK906326 | BOLD:AED3604 | SDEI-Dipt-0000603 | GBMNB15355-20 | SDEI |
| Pseudolycoriella jejunella Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906351 | BOLD:AED1935 | SDEI-Dipt-0000935 | GBMNB15380-20 | SDEI |
| Pseudolycoriella kaikoura Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906321 | BOLD:AED6542 | SDEI-Dipt-0000500 | GBMNB15350-20 | NZAC |
| Pseudolycoriella macrotegmenta Mohrig, 1999 | NEW ZEALAND: North Island | Male | MK906340 | BOLD:AED4092 | SDEI-Dipt-0000757 | GBMNB15369-20 | SDEI |
| Pseudolycoriella maddisoni Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906332 | BOLD:AED4438 | SDEI-Dipt-0000664 | GBMNB15361-20 | SDEI |
| Pseudolycoriella mahanga Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906324 | BOLD:AED1463 | SDEI-Dipt-0000595 | GBMNB15353-20 | SDEI |
| Pseudolycoriella microcteniuni (Yang & Zhang, 1987) | SEYCHELLES: La Digue | Male | N/A | BOLD:AFG5550 | SDEI-Dipt-0002109 | SSEY002-23 | SDEI |
| Pseudolycoriella nodulosa (Mohrig &Krivosheina, 1985) | NORWAY: Oslo | Male | N/A | BOLD:ACY8397 | bf-sci-00732 | SCINO772-15 | BFCO |
| Pseudolycoriella orite Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906347 | BOLD:AED1715 | SDEI-Dipt-0000863 | GBMNB15376-20 | NZAC |
| Pseudolycoriella paludum (Frey, 1948) | NORWAY: Østfold | Male | N/A | BOLD:ACP4204 | bf-sci-00343 | SCINO341-15 | BFCO |
| Pseudolycoriella plicitegmenta Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906366 | BOLD:AED8311 | SDEI-Dipt-0001153 | GBMNB15395-20 | SDEI |
| Pseudolycoriella porotaka Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906375 | BOLD:AED5151 | SDEI-Dipt-0001225 | GBMNB15404-20 | NZAC |
| Pseudolycoriella puhihi Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906396 | BOLD:AED1266 | SDEI-Dipt-0001422 | GBMNB15425-20 | SDEI |
| Pseudolycoriella raki Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906345 | BOLD:AED4711 | SDEI-Dipt-0000834 | GBMNB15374-20 | NZAC |
| Pseudolycoriella robustotegmenta Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906393 | BOLD:AED8311 | SDEI-Dipt-0001371 | GBMNB15422-20 | SDEI |
| Pseudolycoriella subbruckii (Mohrig &Hövemeyer, 1992) | GERMANY: Bavaria | Male | N/A | BOLD:ACG6269 | BIOUG07408-E12 | GMGRF345-13 | ZSMC |
| Pseudolycoriella submonticula (Mohrig & Mamaev, 1990) | GERMANY: Bavaria | Female | N/A | BOLD:ACF9727 | BIOUG05458-H11 | GMGRC1885-13 | ZSMC |
| Pseudolycoriella subtilitegmenta Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906364 | BOLD:AED8311 | SDEI-Dipt-0001151 | GBMNB15393-20 | NZAC |
| Pseudolycoriella sudhausi Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906369 | BOLD:AED1862 | SDEI-Dipt-0001170 | GBMNB15398-20 | SDEI |
| Pseudolycoriella teo Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906402 | N/A | SDEI-Dipt-0001528 | GBMNB15431-20 | NZAC |
| Pseudolycoriella tewaipounamu Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906338 | BOLD:AED4891 | SDEI-Dipt-0000752 | GBMNB15367-20 | SDEI |
| Pseudolycoriella tonnoiri Köhler, 2016 | NEW ZEALAND: North Island | Male | MK906325 | BOLD:ADM7629 | SDEI-Dipt-0000597 | GBMNB15354-20 | SDEI |
| Pseudolycoriella tuakana Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906389 | BOLD:AED4777 | SDEI-Dipt-0001337 | GBMNB15418-20 | SDEI |
| Pseudolycoriella villabonensis Heller, 2012 | ARGENTINA: Misiones | N/A | OM597266 | BOLD:ACO0105 | BIOUG24905-H03 | GMAGO963-15 | MACN |
| Pseudolycoriella wernermohrigi Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906333 | BOLD:AED4093 | SDEI-Dipt-0000665 | GBMNB15362-20 | SDEI |
| Pseudolycoriella whakahara Köhler, 2019 | NEW ZEALAND: South Island | Male | MK906408 | BOLD:AED2160 | SDEI-Dipt-0001439 | GBMNB15437-20 | NZAC |
| Pseudolycoriella whena Köhler, 2019 | NEW ZEALAND: North Island | Male | MK906388 | BOLD:AED1474 | SDEI-Dipt-0001336 | GBMNB15417-20 | NZAC |
| Pseudolycoriella zealandica (Edwards, 1927) | NEW ZEALAND: North Island | Male | MK906328 | BOLD:ADN2926 | SDEI-Dipt-0000623 | GBMNB15357-20 | SDEI |
| Spinopygina acerfalx Vilkamaa, Burdíková & Ševčík, 2023 | USA: Oregon | Male | OQ024760 | N/A | JSSCI80 | N/A | UOLC |
| Stenacanthella freyi (Tuomikoski, 1960) | NORWAY: Buskerud | Male | N/A | BOLD:ACU4800 | bf-sci-00397 | SCINO395-15 | BFCO |
| Stenacanthella secundaria (Mohrig & Menzel, 1990) | CANADA: Ontario | Male | MF841640 | BOLD:AAU6614 | BIOUG20457-G07 | CNTIA197-15 | CBGC |
| Trichocoelina vitticollis (Holmgren, 1883) | NORWAY: Svalbard | Male | MN135691 | BOLD:ABA5288 | SV967 | SVDIP306-13 | NHMO |
| Xylosciara lignicola (Winnertz, 1867) | CANADA: Ontario | Female | KR432701 | BOLD:AAU6615 | BIOUG04314-A02 | CNBPD380-12 | CBGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menzel, F.; Kramp, K.; Amorim, D.d.S.; Gorab, E.; Uliana, J.V.C.; Sauaia, H.; Monesi, N. Pseudolycoriella hygida (Sauaia and Alves)—An Overview of a Model Organism in Genetics, with New Aspects in Morphology and Systematics. Insects 2024, 15, 118. https://doi.org/10.3390/insects15020118
Menzel F, Kramp K, Amorim DdS, Gorab E, Uliana JVC, Sauaia H, Monesi N. Pseudolycoriella hygida (Sauaia and Alves)—An Overview of a Model Organism in Genetics, with New Aspects in Morphology and Systematics. Insects. 2024; 15(2):118. https://doi.org/10.3390/insects15020118
Chicago/Turabian StyleMenzel, Frank, Katja Kramp, Dalton de Souza Amorim, Eduardo Gorab, João Vitor Cardoso Uliana, Heni Sauaia, and Nadia Monesi. 2024. "Pseudolycoriella hygida (Sauaia and Alves)—An Overview of a Model Organism in Genetics, with New Aspects in Morphology and Systematics" Insects 15, no. 2: 118. https://doi.org/10.3390/insects15020118