
Competition among Flavescence Dorée Phytoplasma Strains in the Experimental Insect Vector Euscelidius variegatus
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants, Insects, and Phytoplasma Strains
2.2. Mixed Acquisition by Feeding on FD-C- and FD-D-Infected Plants
2.3. Mixed Acquisition via Abdominal Microinjection of FD-C and FD-D Suspension
2.4. Phytoplasma Transmission following “Mixed Acquisition by Feeding”
2.5. Insect Dissection
2.6. DNA Extraction, Phytoplasma Diagnosis, and Quantification of Pathogen Load
2.7. Data Analysis
3. Results
3.1. Mixed Acquisition by Feeding on FD-C- and FD-D-Infected Plants
3.1.1. Phytoplasma Loads in Infected Source Plants
3.1.2. Presence and Amount of Phytoplasma Strains in Insect Heads and Bodies
3.1.3. Insect Infection and Competition between Phytoplasma Strains
3.2. Mixed Acquisition by Abdominal Microinjection of FD-C and FD-D Suspension
3.2.1. Pathogen Load in Phytoplasma Suspension
3.2.2. Presence of Phytoplasma Strains in Insect Heads and Bodies
3.2.3. Insect Infection and Competition between Phytoplasma Strains
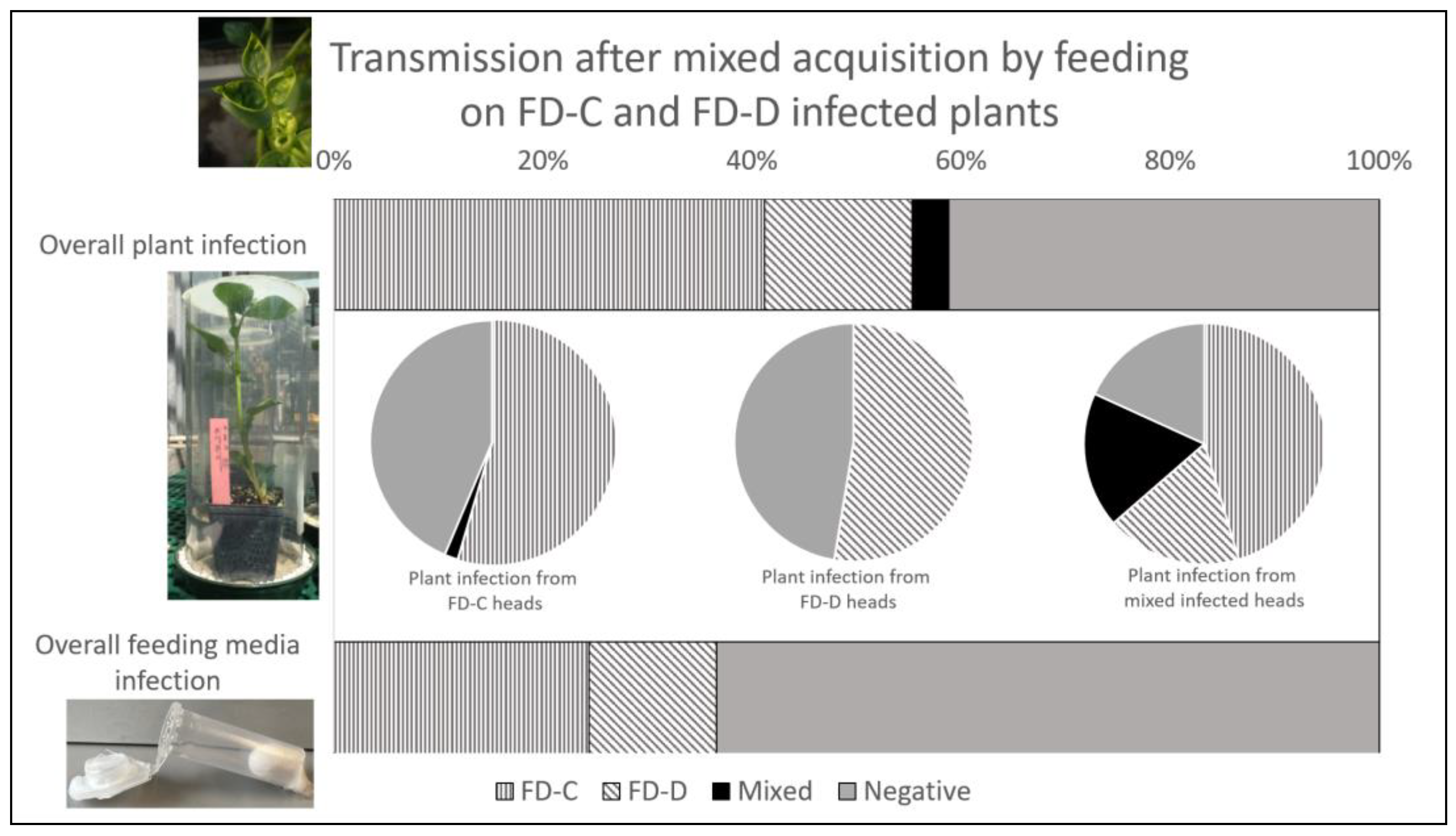
3.3. Phytoplasma Transmission following Mixed Acquisition by Feeding
3.3.1. Presence and Amount of Phytoplasma Strains in Inoculated Plants and in Feeding Media
3.3.2. Insect Infection and Competition between Phytoplasma Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Namba, S. Molecular and Biological Properties of Phytoplasmas. Proc. Jpn. Acad. Ser. B 2019, 95, 401–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomkins, M.; Kliot, A.; Marée, A.F.; Hogenhout, S.A. A Multi-Layered Mechanistic Modelling Approach to Understand How Effector Genes Extend beyond Phytoplasma to Modulate Plant Hosts, Insect Vectors and the Environment. Curr. Opin. Plant Biol. 2018, 44, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhao, Y. Phytoplasma Taxonomy: Nomenclature, Classification, and Identification. Biology 2022, 11, 1119. [Google Scholar] [CrossRef] [PubMed]
- IRPCM ‘Candidatus Phytoplasma’, a Taxon for the Wall-Less, Non-Helical Prokaryotes That Colonize Plant Phloem and Insects. Int. J. Syst. Evol. Microbiol. 2004, 54, 1243–1255. [CrossRef] [PubMed] [Green Version]
- Bertaccini, A.; Arocha-Rosete, Y.; Contaldo, N.; Duduk, B.; Fiore, N.; Montano, H.G.; Kube, M.; Kuo, C.-H.; Martini, M.; Oshima, K.; et al. Revision of the ‘Candidatus Phytoplasma’ Species Description Guidelines. Int. J. Syst. Evol. Microbiol. 2022, 72. [Google Scholar] [CrossRef]
- Lee, I.-M.; Martini, M.; Marcone, C.; Zhu, S.F. Classification of Phytoplasma Strains in the Elm Yellows Group (16SrV) and Proposal of ‘Candidatus Phytoplasma Ulmi’ for the Phytoplasma Associated with Elm Yellows. Int. J. Syst. Evol. Microbiol. 2004, 54, 337–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.-Y.; Sawayanagi, T.; Kakizawa, S.; Nishigawa, H.; Wei, W.; Oshima, K.; Miyata, S.; Ugaki, M.; Hibi, T.; Namba, S. ‘Candidatus Phytoplasma Ziziphi’, a Novel Phytoplasma Taxon Associated with Jujube Witches’-Broom Disease. Int. J. Syst. Evol. Microbiol. 2003, 53, 1037–1041. [Google Scholar] [CrossRef] [Green Version]
- Malembic-Maher, S.; Salar, P.; Filippin, L.; Carle, P.; Angelini, E.; Foissac, X. Genetic Diversity of European Phytoplasmas of the 16SrV Taxonomic Group and Proposal of ‘Candidatus Phytoplasma Rubi. Int. J. Syst. Evol. Microbiol. 2011, 61, 2129–2134. [Google Scholar] [CrossRef] [Green Version]
- Win, N.K.K.; Lee, S.-Y.; Bertaccini, A.; Namba, S.; Jung, H.-Y. ‘Candidatus Phytoplasma Balanitae’ Associated with Witches’ Broom Disease of Balanites Triflora. Int. J. Syst. Evol. Microbiol. 2013, 63, 636–640. [Google Scholar] [CrossRef] [Green Version]
- Debonneville, C.; Mandelli, L.; Brodard, J.; Groux, R.; Roquis, D.; Schumpp, O. The Complete Genome of the “Flavescence Dorée” Phytoplasma Reveals Characteristics of Low Genome Plasticity. Biology 2022, 11, 953. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority); Tramontini, S.; Delbianco, A.; Vos, S. Pest Survey Card on Flavescence Dorée Phytoplasma and Its Vector Scaphoideus titanus. EFSA Support. Publ. 2020, 17, EN-1909. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority); Baker, R.; Gilioli, G.; Behring, C.; Candiani, D.; Andrey, G.; Tomasz, K.; Kinkar, M.; Mosbach-Schulz, O.; Maria, N.F.; et al. Grapevine Flavescence Dorée Pest Report and Datasheet to Support Ranking of EU Candidate Priority Pests 2019; European Food Safety Authority: Parma, Italy, 2019. Available online: https://zenodo.org/record/2789595 (accessed on 19 June 2023).
- EFSA Panel on Plant Health (PLH). Scientific Opinion on Pest Categorisation of Grapevine Flavescence Dorée. EFSA J. 2014, 12, 3851. [Google Scholar] [CrossRef]
- EFSA Panel on Plant Health (PLH); Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Jaques Miret, J.A.; MacLeod, A.; et al. Risk to Plant Health of Flavescence Dorée for the EU Territory. EFSA J. 2016, 14, 4603. [Google Scholar] [CrossRef]
- Angelini, E.; Clair, D.; Borgo, M.; Bertaccini, A.; Boudon-Padieu, E. Flavescence Dorée in France and Italy—Occurrence of Closely Related Phytoplasma Isolates and Their near Relationships to Palatinate Grapevine Yellows and an Alder Yellows Phytoplasma. Vitis 2001, 40, 79–86. [Google Scholar]
- Martini, M.; Botti, S.; Marcone, C.; Marzachì, C.; Casati, P.; Bianco, P.A.; Benedetti, R.; Bertaccini, A. Genetic Variability among Flavescence Dorée Phytoplasmas from Different Origins in Italy and France. Mol. Cell. Probes 2002, 16, 197–208. [Google Scholar] [CrossRef]
- Arnaud, G.; Malembic-Maher, S.; Salar, P.; Bonnet, P.; Maixner, M.; Marcone, C.; Boudon-Padieu, E.; Foissac, X. Multilocus Sequence Typing Confirms the Close Genetic Interrelatedness of Three Distinct Flavescence Dorée Phytoplasma Strain Clusters and Group 16SrV Phytoplasmas Infecting Grapevine and Alder in Europe. Appl. Environ. Microbiol. 2007, 73, 4001–4010. [Google Scholar] [CrossRef] [Green Version]
- Rossi, M.; Pegoraro, M.; Ripamonti, M.; Abbà, S.; Beal, D.; Giraudo, A.; Veratti, F.; Malembic-Maher, S.; Salar, P.; Bosco, D.; et al. Genetic Diversity of Flavescence Dorée Phytoplasmas at the Vineyard Scale. Appl. Environ. Microbiol. 2019, 85, e03123-18. [Google Scholar] [CrossRef] [Green Version]
- Malembic-Maher, S.; Desqué, D.; Khalil, D.; Salar, P.; Bergey, B.; Danet, J.-L.; Duret, S.; Dubrana-Ourabah, M.-P.; Beven, L.; Ember, I.; et al. When a Palearctic Bacterium Meets a Nearctic Insect Vector: Genetic and Ecological Insights into the Emergence of the Grapevine Flavescence Dorée Epidemics in Europe. PLoS Pathog. 2020, 16, e1007967. [Google Scholar] [CrossRef] [Green Version]
- Morone, C.; Boveri, M.; Giosuè, S.; Gotta, P.; Rossi, V.; Scapin, I.; Marzachi, C. Epidemiology of Flavescence Dorée in Vineyards in Northwestern Italy. Phytopathology 2007, 97, 1422–1427. [Google Scholar] [CrossRef] [Green Version]
- Ripamonti, M.; Pegoraro, M.; Rossi, M.; Bodino, N.; Beal, D.; Panero, L.; Marzachì, C.; Bosco, D. Prevalence of Flavescence Dorée Phytoplasma-Infected Scaphoideus titanus in Different Vineyard Agroecosystems of Northwestern Italy. Insects 2020, 11, 301. [Google Scholar] [CrossRef]
- Daire, X.; Clair, D.; Reinert, W.; Boudon-Padieu, E. Detection and Differentiation of Grapevine Yellows Phytoplasmas Belonging to the Elm Yellows Group and to the Stolbur Subgroup by PCR Amplification of Non-Ribosomal DNA. Eur. J. Plant Pathol. 1997, 103, 507–514. [Google Scholar] [CrossRef]
- Quaglino, F.; Casati, P.; Bianco, P.A. Distinct RpsC Single Nucleotide Polymorphism Lineages of Flavescence Dorée Subgroup 16SrV-D Phytoplasma Co-Infect Vitis vinifera L. Folia Microbiol. 2010, 55, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Trivellone, V. An Online Global Database of Hemiptera -Phytoplasma-Plant Biological Interactions. Biodivers. Data J. 2019, 7, e32910. [Google Scholar] [CrossRef]
- Weintraub, P.G.; Beanland, L. Insect Vectors of Phytoplasmas. Annu. Rev. Entomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Trivellone, V.; Dietrich, C.H. Evolutionary Diversification in Insect Vector–Phytoplasma–Plant Associations. Ann. Entomol. Soc. Am. 2021, 114, 137–150. [Google Scholar] [CrossRef]
- Suzuki, S.; Oshima, K.; Kakizawa, S.; Arashida, R.; Jung, H.-Y.; Yamaji, Y.; Nishigawa, H.; Ugaki, M.; Namba, S. Interaction between the Membrane Protein of a Pathogen and Insect Microfilament Complex Determines Insect-Vector Specificity. Proc. Natl. Acad. Sci. USA 2006, 103, 4252–4257. [Google Scholar] [CrossRef] [Green Version]
- Galetto, L.; Bosco, D.; Balestrini, R.; Genre, A.; Fletcher, J.; Marzachì, C. The Major Antigenic Membrane Protein of “Candidatus Phytoplasma Asteris” Selectively Interacts with ATP Synthase and Actin of Leafhopper Vectors. PLoS ONE 2011, 6, e22571. [Google Scholar] [CrossRef] [Green Version]
- Rashidi, M.; Galetto, L.; Bosco, D.; Bulgarelli, A.; Vallino, M.; Veratti, F.; Marzachì, C. Role of the Major Antigenic Membrane Protein in Phytoplasma Transmission by Two Insect Vector Species. BMC Microbiol. 2015, 15, 193. [Google Scholar] [CrossRef] [Green Version]
- Trivellone, V.; Ripamonti, M.; Angelini, E.; Filippin, L.; Rossi, M.; Marzachí, C.; Galetto, L. Evidence Suggesting Interactions between Immunodominant Membrane Protein Imp of Flavescence Dorée Phytoplasma and Protein Extracts from Distantly Related Insect Species. J. Appl. Microbiol. 2019, 127, 1801–1813. [Google Scholar] [CrossRef]
- Arricau-Bouvery, N.; Dubrana, M.-P.; Canuto, F.; Duret, S.; Brocard, L.; Claverol, S.; Malembic-Maher, S.; Foissac, X. Flavescence Dorée Phytoplasma Enters Insect Cells by a Clathrin-Mediated Endocytosis Allowing Infection of Its Insect Vector. Sci. Rep. 2023, 13, 2211. [Google Scholar] [CrossRef]
- Arricau-Bouvery, N.; Duret, S.; Dubrana, M.-P.; Batailler, B.; Desqué, D.; Béven, L.; Danet, J.-L.; Monticone, M.; Bosco, D.; Malembic-Maher, S.; et al. Variable Membrane Protein A of Flavescence Dorée Phytoplasma Binds the Midgut Perimicrovillar Membrane of Euscelidius variegatus and Promotes Adhesion to Its Epithelial Cells. Appl. Environ. Microbiol. 2018, 84, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Arricau-Bouvery, N.; Duret, S.; Dubrana, M.-P.; Desqué, D.; Eveillard, S.; Brocard, L.; Malembic-Maher, S.; Foissac, X. Interactions between the Flavescence Dorée Phytoplasma and Its Insect Vector Indicate Lectin-Type Adhesion Mediated by the Adhesin VmpA. Sci. Rep. 2021, 11, 11222. [Google Scholar] [CrossRef] [PubMed]
- Chuche, J.; Thiéry, D. Biology and Ecology of the Flavescence Dorée Vector Scaphoideus titanus: A Review. Agron. Sustain. Dev. 2014, 34, 381–403. [Google Scholar] [CrossRef] [Green Version]
- Caudwell, A.; Kuszala, C.; Larrue, J.; Bachelier, J. Transmission de La Flavescence Dorée de La Fève à La Fève Par Des Cicadelles Des Genres Euscelis et Euscelidius. Ann. Phytopathol. 1972, 1572, 181–189. [Google Scholar]
- Salar, P.; Charenton, C.; Foissac, X.; Malembic-Maher, S. Multiplication Kinetics of Flavescence Dorée Phytoplasma in Broad Bean. Effect of Phytoplasma Strain and Temperature. Eur. J. Plant Pathol. 2013, 135, 371–381. [Google Scholar] [CrossRef]
- Galetto, L.; Abbà, S.; Rossi, M.; Vallino, M.; Pesando, M.; Arricau-Bouvery, N.; Dubrana, M.-P.; Chitarra, W.; Pegoraro, M.; Bosco, D.; et al. Two Phytoplasmas Elicit Different Responses in the Insect Vector Euscelidius variegatus Kirschbaum. Infect. Immun. 2018, 86, IAI.00042-18. [Google Scholar] [CrossRef] [Green Version]
- Mori, N.; Bressan, A.; Martin, M.; Guadagnini, M.; Girolami, V.; Bertaccini, A. Experimental Transmission by Scaphoideus titanus Ball of Two Flavescence Doree-Type Phytoplasmas. VITIS J. Grapevine Res. 2015, 99. [Google Scholar] [CrossRef]
- Rossi, M.; Vallino, M.; Galetto, L.; Marzachì, C. Competitive Exclusion of Flavescence Dorée Phytoplasma Strains in Catharanthus roseus Plants. Plants 2020, 9, 1594. [Google Scholar] [CrossRef]
- Galetto, L.; Miliordos, D.; Roggia, C.; Rashidi, M.; Sacco, D.; Marzachì, C.; Bosco, D. Acquisition Capability of the Grapevine Flavescence Dorée by the Leafhopper Vector Scaphoideus titanus Ball Correlates with Phytoplasma Titre in the Source Plant. J. Pest Sci. 2014, 87, 671–679. [Google Scholar] [CrossRef]
- Galetto, L.; Pegoraro, M.; Marzachì, C.; Rossi, E.; Lucchi, A.; Bosco, D. Potential Role of the Alien Planthopper Ricania speculum as Vector of Flavescence Dorée Phytoplasma. Eur. J. Plant Pathol. 2019, 154, 1103–1110. [Google Scholar] [CrossRef]
- Bressan, A.; Clair, D.; Sémétey, O.; Boudon-Padieu, E. Insect Injection and Artificial Feeding Bioassays to Test the Vector Specificity of Flavescence Dorée Phytoplasma. Phytopathology 2006, 96, 790–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Wilson, B.A.L.; Ash, G.J.; Woruba, S.B.; Fletcher, M.J.; You, M.; Yang, G.; Gurr, G.M. Determining Putative Vectors of the Bogia Coconut Syndrome Phytoplasma Using Loop-Mediated Isothermal Amplification of Single-Insect Feeding Media. Sci. Rep. 2016, 6, 35801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melamed, S.; Tanne, E.; Ben-Haim, R.; Edelbaum, O.; Yogev, D.; Sela, I. Identification and Characterization of Phytoplasmal Genes, Employing a Novel Method of Isolating Phytoplasmal Genomic DNA. J. Bacteriol. 2003, 185, 6513–6521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, C.; Salar, P.; Gillet, J.; Cloquemin, G.; Very, P.; Foissac, X.; Malembic-Maher, S. Triplex Real-Time PCR Assay for Sensitive and Simultaneous Detection of Grapevine Phytoplasmas of the 16SrV and 16SrXII-A Groups with an Endogenous Analytical Control. Vitis 2009, 48, 87–95. [Google Scholar]
- Marzachì, C.; Veratti, F.; Bosco, D. Direct PCR Detection of Phytoplasmas in Experimentally Infected Insects. Ann. Appl. Biol. 1998, 133, 45–54. [Google Scholar] [CrossRef]
- Kosmidis, I.; Kenne Pagui, E.C.; Sartori, N. Mean and Median Bias Reduction in Generalized Linear Models. Stat. Comput. 2020, 30, 43–59. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna Austria R Found. Stat. Comput. 2019, 12. Available online: https://www.R-project.org/ (accessed on 19 June 2023).
- Plavec, J.; Budinšćak, Ž.; Križanac, I.; Škorić, D.; Foissac, X.; Šeruga Musić, M. Multilocus Sequence Typing Reveals the Presence of Three Distinct Flavescence Dorée Phytoplasma Genetic Clusters in Croatian Vineyards. Plant Pathol. 2019, 68, 18–30. [Google Scholar] [CrossRef] [Green Version]
- Krstić, O.; Cvrković, T.; Marinković, S.; Jakovljević, M.; Mitrović, M.; Toševski, I.; Jović, J. Genetic Diversity of Flavescence Dorée Phytoplasmas in Vineyards of Serbia: From the Widespread Occurrence of Autochthonous Map-M51 to the Emergence of Endemic Map-FD2 (Vectotype II) and New Map-FD3 (Vectotype III) Epidemic Genotypes. Agronomy 2022, 12, 448. [Google Scholar] [CrossRef]
- Tamborindeguy, C.; Hata, F.T.; Molina, R.d.O.; Nunes, W.M.d.C. A New Perspective on the Co-Transmission of Plant Pathogens by Hemipterans. Microorganisms 2023, 11, 156. [Google Scholar] [CrossRef]
- Fu, S.; Gouin, C.; Zhou, C.; Hartung, J.S. Limited Infection by ‘Candidatus Liberibacter Asiaticus’ in ‘Valencia’ Sweet Orange Trees in the Presence of Citrus Tristeza Virus. J. Integr. Agric. 2019, 18, 2284–2293. [Google Scholar] [CrossRef]
- Wintermantel, W.M.; Cortez, A.A.; Anchieta, A.G.; Gulati-Sakhuja, A.; Hladky, L.L. Co-Infection by Two Criniviruses Alters Accumulation of Each Virus in a Host-Specific Manner and Influences Efficiency of Virus Transmission. Phytopathology 2008, 98, 1340–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautam, S.; Gadhave, K.R.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Virus-Virus Interactions in a Plant Host and in a Hemipteran Vector: Implications for Vector Fitness and Virus Epidemics. Virus Res. 2020, 286, 198069. [Google Scholar] [CrossRef]
- Dalmon, A.; Fabre, F.; Guilbaud, L.; Lecoq, H.; Jacquemond, M. Comparative Whitefly Transmission of Tomato Chlorosis Virus and Tomato Infectious Chlorosis Virus from Single or Mixed Infections. Plant Pathol. 2009, 58, 221–227. [Google Scholar] [CrossRef]
- Nolasco, G.; Fonseca, F.; Silva, G. Occurrence of Genetic Bottlenecks during Citrus Tristeza Virus Acquisition by Toxoptera citricida under Field Conditions. Arch. Virol. 2008, 153, 259–271. [Google Scholar] [CrossRef]
- Broadbent, P. Biological Characterization of Australian Isolates of Citrus Tristeza Virus and Separation of Subisolates by Single Aphid Transmissions. Plant Dis. 1996, 80, 329. [Google Scholar] [CrossRef]
- Brlansky, R.H.; Damsteegt, V.D.; Howd, D.S.; Roy, A. Molecular Analyses of Citrus Tristeza Virus Subisolates Separated by Aphid Transmission. Plant Dis. 2003, 87, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashidi, M.; D’Amelio, R.; Galetto, L.; Marzachì, C.; Bosco, D. Interactive Transmission of Two Phytoplasmas by the Vector Insect. Ann. Appl. Biol. 2014, 165, 404–413. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial Production of Bacteriocins: Latest Research Development and Applications. Biotechnol. Adv. 2018, 36, 2187–2200. [Google Scholar] [CrossRef] [PubMed]
- Hood, R.D.; Singh, P.; Hsu, F.; Güvener, T.; Carl, M.A.; Trinidad, R.R.S.; Silverman, J.M.; Ohlson, B.B.; Hicks, K.G.; Plemel, R.L.; et al. A Type VI Secretion System of Pseudomonas aeruginosa Targets a Toxin to Bacteria. Cell Host Microbe 2010, 7, 25–37. [Google Scholar] [CrossRef] [Green Version]
- MacIntyre, D.L.; Miyata, S.T.; Kitaoka, M.; Pukatzki, S. The Vibrio cholerae Type VI Secretion System Displays Antimicrobial Properties. Proc. Natl. Acad. Sci. USA 2010, 107, 19520–19524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, B.; Sule, S.; Jelkmann, W.; Seemüller, E. Suppression of Aggressive Strains of “Candidatus Phytoplasma Mali” by Mild Strains in Catharanthus roseus and Nicotiana occidentalis and Indication of Similar Action in Apple Trees. Phytopathology 2014, 104, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Abbà, S.; Galetto, L.; Carle, P.; Carrère, S.; Delledonne, M.; Foissac, X.; Palmano, S.; Veratti, F.; Marzachì, C. RNA-Seq Profile of Flavescence Dorée Phytoplasma in Grapevine. BMC Genom. 2014, 15, 1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galetto, L.; Pesando, M.; Abbà, S.; Bodino, N.; Marzachì, C. Transcriptional Profile of Selected Flavescence Dorée Phytoplasma Genes during Infection of Insect Vectors and Host Plants. J. Plant Pathol. 2023, 105, 393–407. [Google Scholar] [CrossRef]
- Jollard, C.; Foissac, X.; Desqué, D.; Razan, F.; Garcion, C.; Beven, L.; Eveillard, S. Flavescence Dorée Phytoplasma Has Multiple FtsH Genes That Are Differentially Expressed in Plants and Insects. IJMS 2019, 21, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| # Experiment | FDp Strain | Mean ± SEM (N) |
|---|---|---|
| Replicate 1 | FD-C | 5.52 × 104 ± 7.78 × 103 (2) |
| FD-D | 5.04 × 104 ± 9.19 × 102 (2) | |
| Replicate 2 | FD-C | 4.40 × 105 ± 9.90 × 104 (2) |
| FD-D | 7.10 × 104 ± 1.41 × 104 (2) |
| Body Infection | ||||||
|---|---|---|---|---|---|---|
| FD-C | FD-D | Mixed FD-C/-D | Negative | Total | ||
| Head infection | FD-C | 51 | 1 | 8 | 0 | 60 |
| FD-D | 1 | 34 | 6 | 0 | 41 | |
| mixed FD-C/-D | 0 | 4 | 9 | 0 | 13 | |
| negative | 3 | 3 | 2 | 6 | 14 | |
| Total | 55 | 42 | 25 | 6 | 128 | |
| Infection Status | FDp Strain | Sample Type | Mean ± SEM |
|---|---|---|---|
| Single-infected samples | FD-C | Bodies | 6.40 × 103 ± 1.61 × 103 |
| Heads | 2.43 × 104 ± 4.85 × 103 | ||
| FD-D | Bodies | 5.08 × 103 ± 1.59 × 103 | |
| Heads | 6.81 × 104 ± 1.64 × 104 | ||
| Mixed-infected samples | FD-C | Bodies | 2.14 × 103 ± 8.57 × 102 |
| Heads | 9.97 × 103 ± 3.43 × 103 | ||
| FD-D | Bodies | 3.44 × 103 ± 9.73 × 102 | |
| Heads | 7.38 × 103 ± 3.84 × 103 |
| Body Infection | ||||||
|---|---|---|---|---|---|---|
| FD-C | FD-D | Mixed FD-C/-D | Negative | Total | ||
| Head infection | FD-C | 2 | 0 | 12 | 1 | 15 |
| FD-D | 0 | 3 | 4 | 1 | 8 | |
| mixed FD-C/-D | 2 | 2 | 11 | 0 | 15 | |
| Negative | 2 | 5 | 8 | 1 | 16 | |
| Total | 6 | 10 | 35 | 3 | 54 | |
| Head Infection | |||||
|---|---|---|---|---|---|
| FD-C | FD-D | Mixed FD-C/-D | Total | ||
| Plant infection | FD-C | 30 | 0 | 5 | 35 |
| FD-D | 0 | 10 | 2 | 12 | |
| mixed FD-C/-D | 1 | 0 | 2 | 3 | |
| Negative | 24 | 9 | 2 | 35 | |
| Total | 55 | 19 | 11 | 85 | |
| Head Infection | |||||
|---|---|---|---|---|---|
| FD-C | FD-D | Mixed FD-C/-D | Total | ||
| Feeding media detection | FD-C | 9 | 0 | 1 | 10 |
| FD-D | 0 | 3 | 2 | 5 | |
| mixed FD-C/-D | 0 | 0 | 0 | 0 | |
| Negative | 17 | 7 | 2 | 26 | |
| Total | 26 | 10 | 5 | 41 | |
| Infection Status | FDp Strain | Collection Date | Mean ± SEM |
|---|---|---|---|
| Singly infected samples | FD-C | 5 wpi | 1.53 × 105 ± 2.75 × 104 |
| 7 wpi | 3.12 × 105 ± 7.00 × 104 | ||
| FD-D | 5 wpi | 2.48 × 105 ± 8.04 × 104 | |
| 7 wpi | 4.71 × 105 ± 1.34 × 105 | ||
| Mixed-infected samples | FD-C | 5 wpi | / |
| 7 wpi | 1.27 × 104 | ||
| FD-D | 5 wpi | / | |
| 7 wpi | 9.32 × 103 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, M.; Galetto, L.; Bodino, N.; Beltramo, J.; Gamalero, S.; Pegoraro, M.; Bosco, D.; Marzachì, C. Competition among Flavescence Dorée Phytoplasma Strains in the Experimental Insect Vector Euscelidius variegatus. Insects 2023, 14, 575. https://doi.org/10.3390/insects14070575
Rossi M, Galetto L, Bodino N, Beltramo J, Gamalero S, Pegoraro M, Bosco D, Marzachì C. Competition among Flavescence Dorée Phytoplasma Strains in the Experimental Insect Vector Euscelidius variegatus. Insects. 2023; 14(7):575. https://doi.org/10.3390/insects14070575
Chicago/Turabian StyleRossi, Marika, Luciana Galetto, Nicola Bodino, Jessica Beltramo, Silvia Gamalero, Mattia Pegoraro, Domenico Bosco, and Cristina Marzachì. 2023. "Competition among Flavescence Dorée Phytoplasma Strains in the Experimental Insect Vector Euscelidius variegatus" Insects 14, no. 7: 575. https://doi.org/10.3390/insects14070575