
Phylogeny and Biogeographic History of Parnassius Butterflies (Papilionidae: Parnassiinae) Reveal Their Origin and Deep Diversification in West China

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
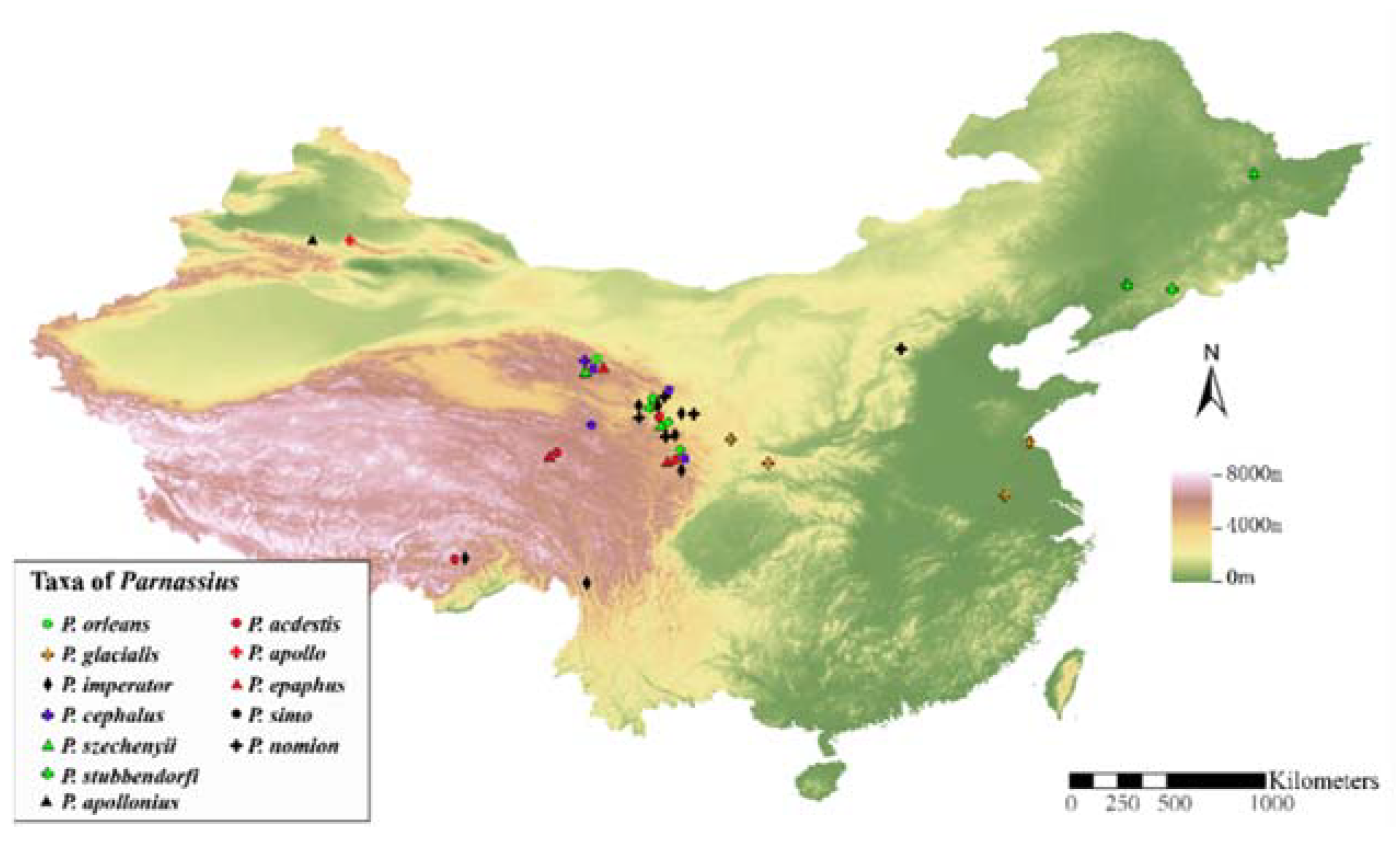
2.1. Specimen Collection
2.2. DNA Extraction, Polymerase Chain Reaction Amplification, and Sequencing
2.3. Phylogenetic Analysis
2.4. Divergence Time Estimation
2.5. Ancestral Area Reconstructions
3. Results
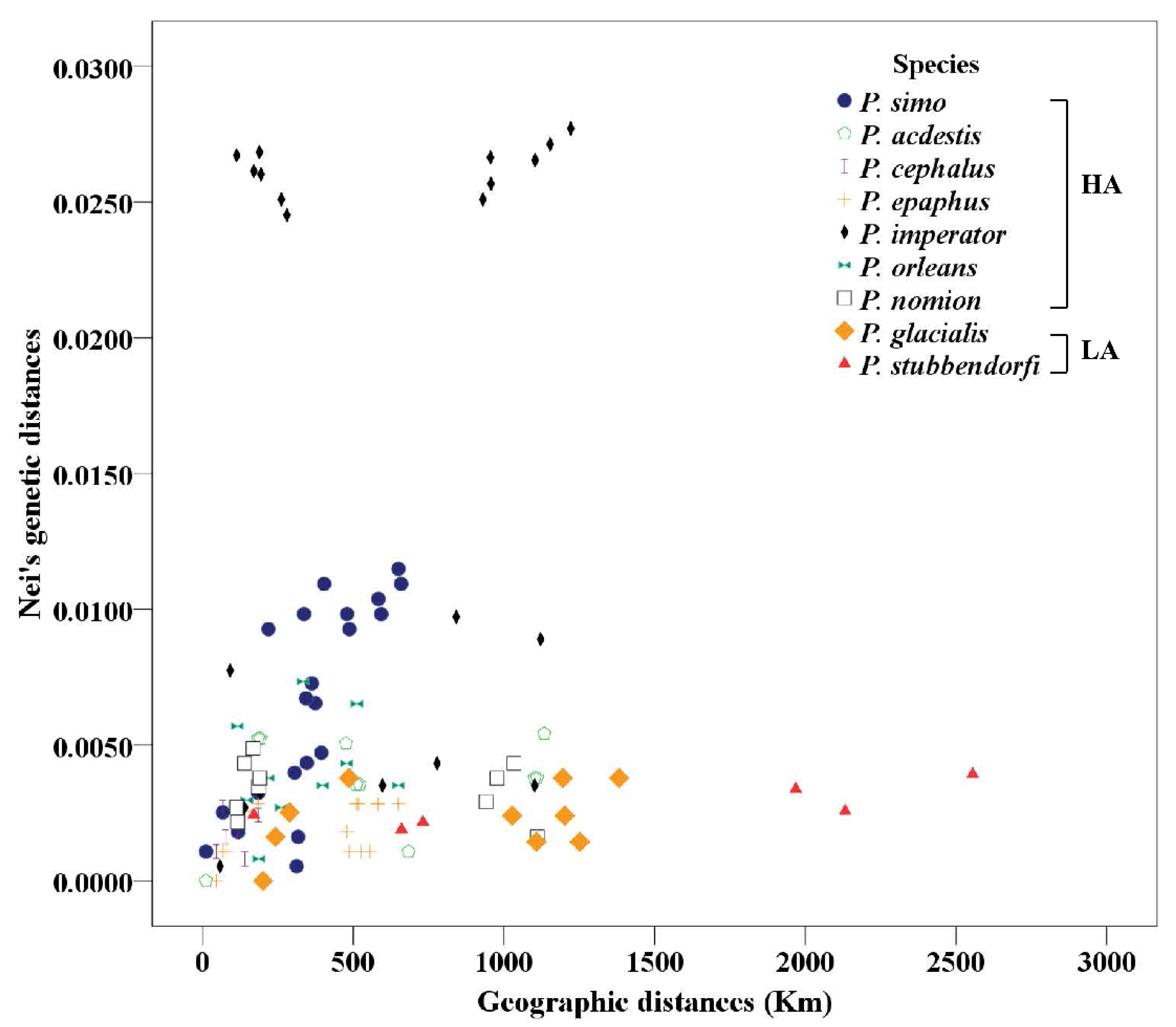
3.1. Sequence Alignment and Genetic Distances
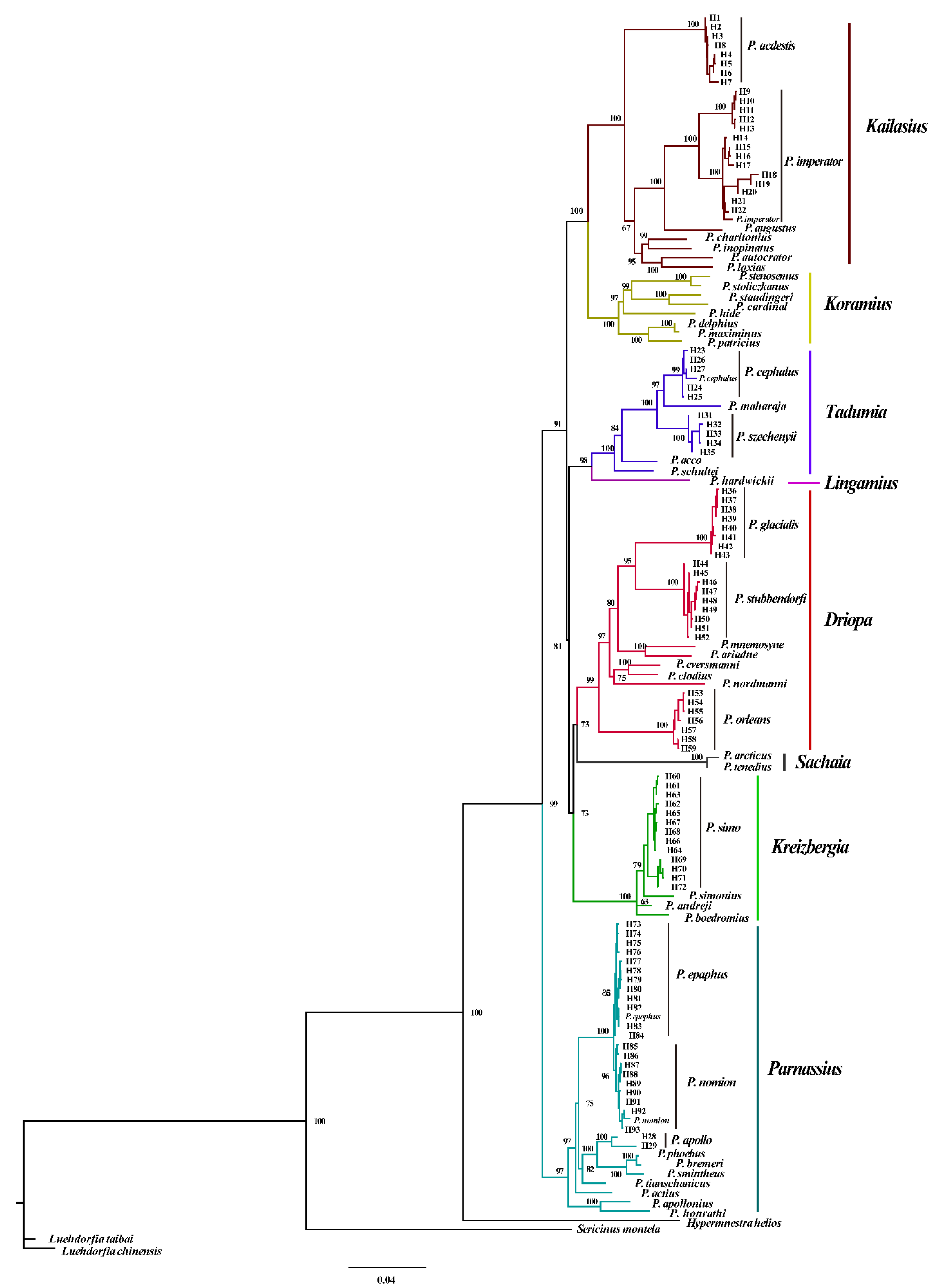
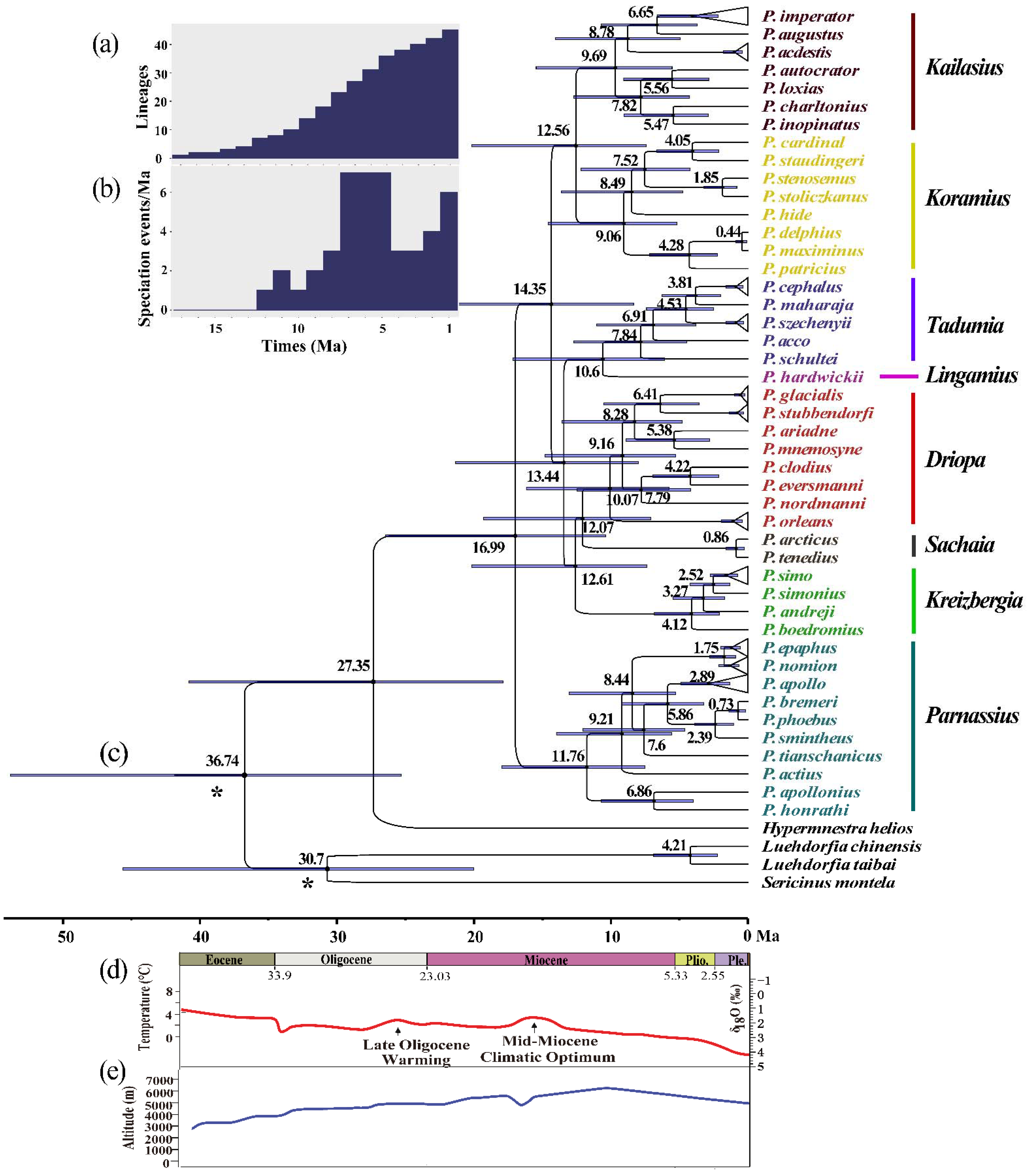
3.2. Phylogenetic Analysis and Divergence Times
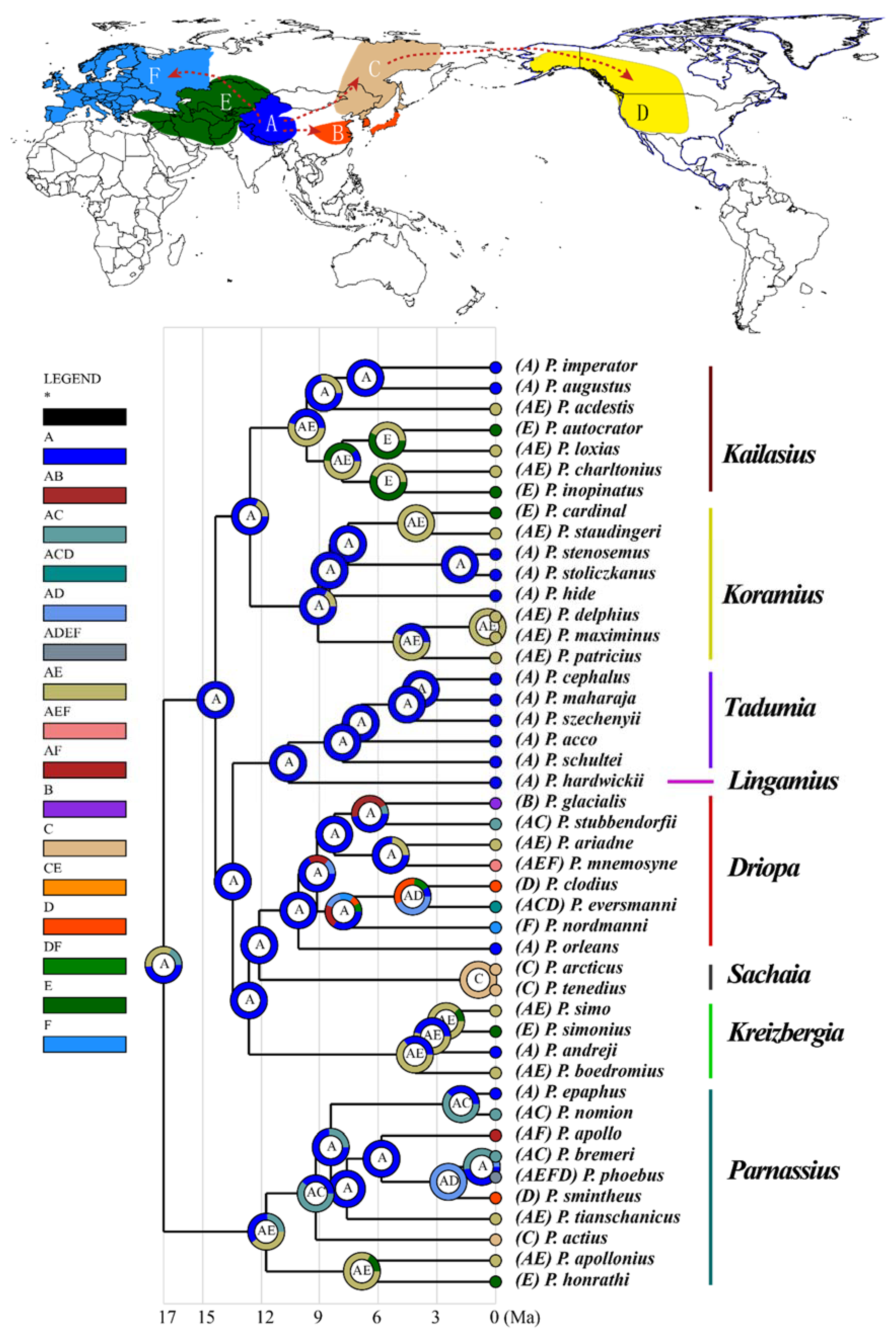
3.3. Ancestral Area Reconstruction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tapponnier, P.; Zhiqin, X.; Roger, F.; Meyer, B.; Arnaud, N.; Wittlinger, G.; Jingsui, Y. Oblique stepwise rise and growth of the tibet plateau. Science 2001, 294, 1671–1677. [Google Scholar] [CrossRef]
- Royden, L.H.; Burchfiel, B.C.; Van Der Hilst, R.D. The geological evolution of the Tibetan plateau. Science 2008, 321, 1054–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Dai, J.; Zhao, X.; Li, Y.; Graham, S.A.; He, D.; Ran, B.; Meng, J. Outward-growth of the Tibetan Plateau during the Cenozoic: A review. Tectonophysics 2014, 621, 1–43. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, H.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Herold, N.; You, Y.; Müller, R.D.; Seton, M. Climate model sensitivity to changes in Miocene paleotopography. Aust. J. Earth Sci. 2009, 56, 1049–1059. [Google Scholar] [CrossRef]
- Guo, Z.T.; Sun, B.; Zhang, Z.S.; Peng, S.Z.; Xiao, G.Q.; Ge, J.Y.; Hao, Q.Z.; Qiao, Y.S.; Liang, M.Y.; Liu, J.F.; et al. A major reorganization of Asian climate by the early Miocene. Clim. Past 2008, 4, 153–174. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Huang, J.; Ding, W. The impact of major geological events on Chinese flora. Biodivers. Sci. 2017, 25, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.M.; Mao, L.F.; Yang, T.; Ye, J.F.; Liu, B.; Li, H.L.; Sun, M.; Miller, J.T.; Mathews, S.; Hu, H.H.; et al. Evolutionary history of the angiosperm flora of China. Nature 2018, 554, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Deng, T.; Jin, Y.; Mao, L.; Zhao, D.; Ricklefs, R.E. Phylogenetic dispersion and diversity in regional assemblages of seed plants in China. Proc. Natl. Acad. Sci. USA 2019, 116, 23192–23201. [Google Scholar] [CrossRef]
- Weiss, J.C.; Rigout, J. The Parnassiinae of the World, Part 6; Goecke & Evers: Keltern, Germany, 2016; ISBN 0953224023. [Google Scholar]
- Chou, I. Monographia Rhopalocerorum Sinensium (Revised Edition); Henan Scientific and Technological Publishing House: Zhengzhou, China, 1999; ISBN 7534921937. [Google Scholar]
- Katoh, T.; Chichvarkhin, A.; Yagi, T.; Omoto, K. Phylogeny and Evolution of Butterflies of the Genus Parnassius: Inferences from Mitochondrial 16S and ND1 Sequences. Zoolog. Sci. 2005, 22, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Omoto, K.; Yonezawa, T.; Shinkawa, T. Molecular systematics and evolution of the recently discovered “Parnassian” butterfly (Parnassius davydovi Churkin, 2006) and its allied species (Lepidoptera, Papilionidae). Gene 2009, 441, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Condamine, F.L. Limited by the roof of the world: Mountain radiations of Apollo swallowtails controlled by diversity-dependence processes. Biol. Lett. 2018, 14, 20170622. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratton, P.; Konopinski, M.K.; Sbordoni, V. Pleistocene evolutionary history of the Clouded Apollo (Parnassius mnemosyne): Genetic signatures of climate cycles and a “time-dependent” mitochondrial substitution rate. Mol. Ecol. 2008, 17, 4248–4262. [Google Scholar] [CrossRef] [Green Version]
- Tao, R.; Xu, C.; Wang, Y.; Sun, X.; Li, C.; Ma, J.; Hao, J.; Yang, Q. Spatiotemporal differentiation of alpine butterfly Parnassius glacialis (Papilionidae: Parnassiinae) in China: Evidence from mitochondrial DNA and nuclear single nucleotide polymorphisms. Genes 2020, 11, 188. [Google Scholar] [CrossRef] [Green Version]
- Kebaïli, C.; Sherpa, S.; Rioux, D.; Després, L. Demographic inferences and climatic niche modelling shed light on the evolutionary history of the emblematic cold-adapted Apollo butterfly at regional scale. Mol. Ecol. 2022, 31, 448–466. [Google Scholar] [CrossRef] [PubMed]
- Bryk, F. Lepidoptera, Parnassidae. Pars II. (Subfam. Parnassiinae). Tierreich 1935, 65, 1–790. [Google Scholar]
- Ehrlich, P.R. The comparative morphology, phylogeny and higher classification of the butterflies (Lepidoptera: Papilionoidea). Kans. Univ. Bull. 1958, 39, 305–370. [Google Scholar]
- Eisner, C. Parnassiidae-Typen in der Sammlung J. C. Eisner. Zool. Verh. 1966, 81, 1–81. [Google Scholar]
- Munroe, E. The Classification of the Papilionidae (Lepidoptera). Mem. Entomol. Soc. Canada 1960, 92, 5–51. [Google Scholar] [CrossRef]
- Ackery, P.R. A guide to the genera and species of Parnassiinae (Lepidoptera: Papilionidae). Bull. Br. Mus. Nat. Hist. 1975, 31, 71–105. [Google Scholar] [CrossRef]
- Hancock, D. Classification of the Papilionidae: A phylogenetic approach. Smithersia 1983, 2, 1–48. [Google Scholar]
- Omoto, K.; Katoh, T.; Chichvarkhin, A.; Yagi, T. Molecular systematics and evolution of the “Apollo” butterflies of the genus Parnassius (Lepidoptera: Papilionidae) based on mitochondrial DNA sequence data. Gene 2004, 326, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Nazari, V.; Zakharov, E.V.; Sperling, F.A.H. Phylogeny, historical biogeography, and taxonomic ranking of Parnassiinae (Lepidoptera, Papilionidae) based on morphology and seven genes. Mol. Phylogenet. Evol. 2007, 42, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Michel, F.; Rebourg, C.; Cosson, E.; Descimon, H. Molecular phylogeny of Parnassiinae butterflies (Lepidoptera: Papilionidae) based on the sequences of four mitochondrial DNA segments. In Annales de la Société Entomologique de France; Taylor & Francis Group: Boca Raton, FL, USA, 2008; Volume 44, pp. 1–36. [Google Scholar]
- Condamine, F.L.; Rolland, J.; Höhna, S.; Sperling, F.A.H.; Sanmartin, I. Testing the role of the Red Queen and Court Jester as drivers of the macroevolution of Apollo butterflies. Syst. Biol. 2018, 67, 940–964. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Yagi, T.; Sasaki, G.; Takebe, H. Phylogeny of Japanese papilionid butterflies inferred from nucleotide sequences of the mitochondrial ND5 gene. J. Mol. Evol. 1999, 48, 42–48. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.H. DAMBE7: New and improved tools for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Rebourg, C.; Péténian, F.; Cosson, E.; Faure, E. Patterns of speciation and adaptive radiation in Parnassius butterflies. J. Entomol. 2006, 3, 204–215. [Google Scholar] [CrossRef]
- Su, C.; Xie, T.; Wang, Y.; Si, C.; Li, L.; Ma, J.; Li, C.; Sun, X.; Hao, J.; Yang, Q. Miocene diversification and high-altitude adaptation of Parnassius butterflies (Lepidoptera: Papilionidae) in Qinghai–Tibet plateau revealed by large-scale transcriptomic data. Insects 2020, 11, 754. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Than, C.; Ruths, D.; Nakhleh, L. PhyloNet: A software package for analyzing and reconstructing reticulate evolutionary relationships. BMC Bioinform. 2008, 9, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohn, J.C.; Labandeira, C.; Davis, D.; Mitter, C. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world. Zootaxa 2012, 3286, 1–132. [Google Scholar] [CrossRef] [Green Version]
- Magallón, S.; Gómez-Acevedo, S.; Sánchez-Reyes, L.L.; Hernández-Hernández, T. A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity. New Phytol. 2015, 207, 437–453. [Google Scholar] [CrossRef]
- Chazot, N.; Wahlberg, N.; Freitas, A.V.L.; Mitter, C.; Labandeira, C.; Sohn, J.C.; Sahoo, R.K.; Seraphim, N.; De Jong, R.; Heikkilä, M. Priors and Posteriors in Bayesian Timing of Divergence Analyses: The Age of Butterflies Revisited. Syst. Biol. 2019, 68, 797–813. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v 1.6. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 20 February 2022).
- Rambaut, A. FigTree Version 1.4.3. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 20 February 2022).
- Stadler, T. Lineages-through-time plots of neutral models for speciation. Math. Biosci. 2008, 216, 163–171. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; He, X. S-DIVA (Statistical Dispersal-Vicariance Analysis): A tool for inferring biogeographic histories. Mol. Phylogenet. Evol. 2010, 56, 848–850. [Google Scholar] [CrossRef] [PubMed]
- Ree, R.H.; Smith, S.A. Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbert, T.D.; Lawrence, K.T.; Tzanova, A.; Peterson, L.C.; Caballero-Gill, R.; Kelly, C.S. Late Miocene global cooling and the rise of modern ecosystems. Nat. Geosci. 2016, 9, 843–847. [Google Scholar] [CrossRef]
- Renner, S.S. Available data point to a 4-km-high Tibetan Plateau by 40 Ma, but 100 molecular-clock papers have linked supposed recent uplift to young node ages. J. Biogeogr. 2016, 43, 1479–1487. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Meng, S.Y.; Allen, G.A.; Wen, J.; Rao, G.Y. Rapid radiation and dispersal out of the qinghai-tibetan plateau of an alpine plant lineage Rhodiola (crassulaceae). Mol. Phylogenet. Evol. 2014, 77, 147–158. [Google Scholar] [CrossRef]
- Ebersbach, J.; Muellner-Riehl, A.N.; Michalak, I.; Tkach, N.; Hoffmann, M.H.; Röser, M.; Sun, H.; Favre, A. In and out of the Qinghai-Tibet Plateau: Divergence time estimation and historical biogeography of the large arctic-alpine genus Saxifraga L. J. Biogeogr. 2017, 44, 900–910. [Google Scholar] [CrossRef]
- Favre, A.; Päckert, M.; Pauls, S.U.; Jähnig, S.C.; Uhl, D.; Michalak, I.; Muellner-Riehl, A.N. The role of the uplift of the Qinghai-Tibetan Plateau for the evolution of Tibetan biotas. Biol. Rev. Camb. Philos. Soc. 2015, 90, 236–253. [Google Scholar] [CrossRef]
- Liu, J.Q.; Wang, Y.J.; Wang, A.L.; Hideaki, O.; Abbott, R.J. Radiation and diversification within the Ligularia-Cremanthodium- Parasenecio complex (Asteraceae) triggered by uplift of the Qinghai-Tibetan Plateau. Mol. Phylogenet. Evol. 2006, 38, 31–49. [Google Scholar] [CrossRef]
- Wang, Y.J.; Liu, J.Q.; Miehe, G. Phylogenetic origins of the himalayan endemic Dolomiaea, Diplazoptilon and Xanthopappus (Asteraceae: Cardueae) based on three DNA regions. Ann. Bot. 2007, 99, 311–322. [Google Scholar] [CrossRef] [Green Version]
- Korb, S.K. An annotated checklist of the tribus parnassiini sensu korshunov of the old world (Lepidoptera, Papilionidae). Acta Biol. Sib. 2020, 6, 59–86. [Google Scholar] [CrossRef]
- Supple, M.A.; Papa, R.; Hines, H.M.; McMillan, W.O.; Counterman, B.A. Divergence with gene flow across a speciation continuum of Heliconius butterflies. BMC Evol. Biol. 2015, 15, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelman, N.B.; Frandsen, P.B.; Miyagi, M.; Clavijo, B.; Davey, J.; Dikow, R.B.; García-Accinelli, G.; Van Belleghem, S.M.; Patterson, N.; Neafsey, D.E.; et al. Genomic architecture and introgression shape a butterfly radiation. Science 2019, 366, 594–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allio, R.; Nabholz, B.; Wanke, S.; Chomicki, G.; Pérez-Escobar, O.A.; Cotton, A.M.; Clamens, A.L.; Kergoat, G.J.; Sperling, F.A.H.; Condamine, F.L. Genome-wide macroevolutionary signatures of key innovations in butterflies colonizing new host plants. Nat. Commun. 2021, 12, 354. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Drew, B.T.; Mavrodiev, E.V.; Gitzendanner, M.A.; Soltis, P.S.; Soltis, D.E. Phylogeny, divergence times, and historical biogeography of the angiosperm family Saxifragaceae. Mol. Phylogenet. Evol. 2015, 83, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Todisco, V.; Gratton, P.; Cesaroni, D.; Sbordoni, V. Phylogeography of Parnassius apollo: Hints on taxonomy and conservation of a vulnerable glacial butterfly invader. Biol. J. Linn. Soc. 2010, 101, 169–183. [Google Scholar] [CrossRef] [Green Version]
- Todisco, V.; Gratton, P.; Zakharov, E.V.; Wheat, C.W.; Sbordoni, V.; Sperling, F.A.H. Mitochondrial phylogeography of the Holarctic Parnassius phoebus complex supports a recent refugial model for alpine butterflies. J. Biogeogr. 2012, 39, 1058–1072. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ND1 (Locality) | COI (Locality) | ND5 (Locality) |
|---|---|---|---|
| Hypermnestra helios Nickerl, 1846 | AJ972131 (Uzbekistan) | AM231506 (Uzbekistan) | AB095659 (Uzbekistan) |
| Sericinus montela Gray, 1853 | AJ972136 (Fuchu, Japan) | AF170868 (Fuchu, Japan) | AB095665 (Kyoto, Japan) |
| Luehdorfia chinensis Leech, 1893 | EU622524 (NA) | EU622524 (NA) | EU622524 (NA) |
| Luehdorfia taibai Chou, 1994 | KC952673 (NA) | KC952673 (NA) | KC952673 (NA) |
| P. tianschanicus Oberthür, 1879 | DQ407806 (Dolon Pass, Kyrgyzstan) | DQ407767 (Dolon Pass, Kyrgyzstan) | AB095648 (Alai, Kyrgyzstan) |
| P. phoebus Fabricius, 1793 | AJ972122 (Tomtor, Yakutia, Russia) | AM231499 (Tomtor, Yakutia, Russia) | AB095654 (Magadan, Russia) |
| P. honrathi Staudinger, 1882 | AJ972129 (Ghissarski Mts, Uzbekistan) | DQ407772 (Ghissarski Mts, Uzbekistan) | AB096091 (Uzbekistan) |
| P. schultei Weiss & Michel, 1989 | AJ972073 (Tibet, China) | AM231445 (Tibet, China) | AB095619 (Tibet, China) |
| P. smintheus Doubleday, 1847 | AJ972125 (Wyoming, USA) | AM231495 (Wyoming, USA) | AB095653 (Colorado, USA) |
| P. maharaja Avinoff, 1916 | AJ972076 (Ladakh, India) | AM231448 (Ladakh, India) | AB095615 (Ladakh, India) |
| P. acco Gray, 1852 | AJ972070 (Ladakh, India) | AM231442 (Ladakh, India) | AB095652 (Tibet, China) |
| P. mnemosyne Linnaeus, 1758 | AM283061 (Kyrgyzstan) | AM231422 (Kyrgyzstan) | AB095626 (Kyrgyzstan) |
| P. eversmanni Ménétriés, 1850 | AJ972056 (Amur, Russia) | AM231430 (Amur, Russia) | AB095608 (Amur, Russia) |
| P. andreji Eisner, 1930 | AJ972068 (Sichuan, China) | AM231440 (Sichuan, China) | AB095643 (Sichuan, China) |
| P. stenosemus Honrath, 1890 | AJ972089 (Zanskar, India) | AM231461 (Zanskar, India) | AB095656 (Ladakh, India) |
| P. simonius Staudinger, 1889 | DQ407809 (Kyrgyzstan) | DQ407758 (Kyrgyzstan) | AB095649 (Kyrgyzstan) |
| P. charltonius Gray, 1852 | AJ972079 (Kyrgyzstan) | AM231451 (Kyrgyzstan) | AB095630 (Kyrgyzstan) |
| P. hardwickii Gray, 1831 | AJ972069 (E. Nepal) | DQ407770 (E. Nepal) | AB094969 (E. Nepal) |
| P. clodius Ménétriés, 1855 | AJ972058 (California, USA) | AF170871 (California, USA) | AB095624 (Montana, USA) |
| P. staudingeri Bang-Haas, 1882 | AJ972103 (Kaltakol, W. Gissar, Uzbekistan) | AM231477 (Kaltakol, W. Gissar, Uzbekistan) | AM283087 (Kaltakol, W. Gissar, Uzbekistan) |
| P. nordmanni Ménétriés, 1850 | AJ972059 (Caucasus, Russia) | AM231432 (Caucasus, Russia) | AB094968 (Caucasus, Russia) |
| P. autocrator Avinoff, 1913 | AJ972082 (Tajikistan) | AM231454 (Tajikistan) | AB095634 (Tajikistan) |
| P. loxias Püngeler, 1901 | AJ972080 (Kyrgyzstan) | AM231452 (Kyrgyzstan) | AB096090 (Kyrgyzstan) |
| P. delphius Eversmann, 1843 | AJ972092 (Kyrgyzstan) | DQ407762 (Kyrgyzstan) | AB095632 (Kyrgyzstan) |
| P. inopinatus Kotzsch, 1940 | AJ972081 (Afghanistan) | AM231453 (Afghanistan) | AB095641 (Afghanistan) |
| P. patricius Niepelt, 1911 | AJ972091 (Kyrgyzstan) | AM231463 (Kyrgyzstan) | AB095620 (Tianshan, Xinjiang, China) |
| P. boedromius Püngeler, 1901 | AJ972067 (Kyrgyzstan) | AM231439 (Kyrgyzstan) | AB095629 (Tianshan, Xinjiang, China) |
| P. hide Koiwaya, 1987 | AJ972090 (Tibet, China) | AM231462 (Tibet, China) | AB095613 (Qinghai, China) |
| P. ariadne Lederer, 1853 | AJ972055 (Altai, Russia) | AM231429 (Altai, Russia) | AB094970 (Altai, Russia) |
| P. stoliczkanus Felder & Felder, 1864 | AJ972087 (Ladakh, India) | AM231459 (Ladakh, India) | AB095650 (Ladakh, India) |
| P. arcticus Eisner, 1968 | AJ972062 (Yakutia, Russia) | AM231434 (Yakutia, Russia) | AB095639 (Yakutia, Russia) |
| P. maximinus Staudinger, 1891 | AJ972094 (Tianshan, Xinjiang, China) | AM231466 (Tianshan, Xinjiang, China) | AB095651 (Tianshan, Xinjiang, China) |
| P. cardinal Grum-Grshimailo, 1887 | AJ972095 (Tajikistan) | AM231467 (Tajikistan) | AB095644 (Tajikistan) |
| P. tenedius Eversmann, 1851 | AJ972063 (Yakutia, Russia) | AM231435 (Yakutia, Russia) | AB095658 (Yakutia, Russia) |
| P. actius Eversmann, 1843 | DQ407807 (Tianshan, Xinjiang, China) | DQ407765 (Tianshan, Xinjiang, China) | AB095622 (Tianshan, Xinjiang, China) |
| P. bremeri Bremer, 1864 | AJ972126 (Korea) | AM231501 (Korea) | AB095611 (Korea) |
| P. augustus Frühstörfer, 1903 | AJ972084 (Tibet, China) | AM231456 (Tibet, China) | AB095645 (Tibet, China) |
| P. imperator Oberthür, 1883 | AJ972083 (Qilianshan, Gansu, China) | DQ407775 (Qilianshan, Gansu, China) | AB095612 (Qinghai, China) |
| P. cephalus Grum-Grshimailo, 1891 | AJ972075 (Kun Lun Shan, China) | AM231447 (Kun Lun Shan, China) | AB095616 (Qamdo, Tibet, China) |
| P. epaphus Oberthür, 1879 | AJ972104 (Hankar, Ladakh, India) | AM231478 (Hankar, Ladakh, India) | AB095610 (Qilianshan, Gansu, China) |
| P. nomion Fischer de Waldheim, 1823 | AJ972109 (Datong Shan, Qinghai, China) | AM231480 (Datong Shan, Qinghai, China) | AB095609 (Primorye, Russia) |
| P. imperator Oberthür, 1883 | This study (7 populations) | This study (7 populations) | This study (7 populations) |
| P. cephalus Grum-Grshimailo, 1891 | This study (4 populations) | This study (4 populations) | This study (4 populations) |
| P. epaphus Oberthür, 1879 | This study (6 populations) | This study (6 populations) | This study (6 populations) |
| P. nomion Fischer de Waldheim, 1823 | This study (5 populations) | This study (5 populations) | This study (5 populations) |
| P. acdestis Grum-Grshimailo, 1891 | This study (5 populations) | This study (5 populations) | This study (5 populations) |
| P. szechenyii Frivaldszky, 1886 | This study (2 populations) | This study (2 populations) | This study (2 populations) |
| P. glacialis Butler, 1866 | This study (5 populations) | This study (5 populations) | This study (5 populations) |
| P. stubbendorfii Ménétriés, 1849 | This study (4 populations) | This study (4 populations) | This study (4 populations) |
| P. orleans Oberthür, 1890 | This study (5 populations) | This study (5 populations) | This study (5 populations) |
| P. simo Gray, 1852 | This study (7 populations) | This study (7 populations) | This study (7 populations) |
| P. apollo Linnaeus, 1758 | This study (2 populations) | This study (2 populations) | This study (2 populations) |
| P. apollonius Eversmann, 1847 | This study (1 population) | This study (1 population) | This study (1 populations) |
| Species | Mean Genetic Distance | Mean Geographic Distance (km) | Main Altitude Distribution (m) |
|---|---|---|---|
| P. glacialis | 0.0023 | 839 | 200–2000 |
| P. stubbendorfii | 0.0027 | 1370 | 300–2500 |
| P. nomion | 0.0034 | 498 | 2000–3500 |
| P. imperator | 0.0171 | 593 | 2800–5100 |
| P. orleans | 0.0041 | 330 | 3000–5000 |
| P. epaphus | 0.0020 | 368 | 3800–5100 |
| P. acdestis | 0.0037 | 593 | 4000–5000 |
| P. simo | 0.0065 | 360 | 4000–5100 |
| P. cephalus | 0.0041 | 193 | 4000–5100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; He, B.; Tao, R.; Su, C.; Ma, J.; Hao, J.; Yang, Q. Phylogeny and Biogeographic History of Parnassius Butterflies (Papilionidae: Parnassiinae) Reveal Their Origin and Deep Diversification in West China. Insects 2022, 13, 406. https://doi.org/10.3390/insects13050406
Zhao Y, He B, Tao R, Su C, Ma J, Hao J, Yang Q. Phylogeny and Biogeographic History of Parnassius Butterflies (Papilionidae: Parnassiinae) Reveal Their Origin and Deep Diversification in West China. Insects. 2022; 13(5):406. https://doi.org/10.3390/insects13050406
Chicago/Turabian StyleZhao, Youjie, Bo He, Ruisong Tao, Chengyong Su, Junye Ma, Jiasheng Hao, and Qun Yang. 2022. "Phylogeny and Biogeographic History of Parnassius Butterflies (Papilionidae: Parnassiinae) Reveal Their Origin and Deep Diversification in West China" Insects 13, no. 5: 406. https://doi.org/10.3390/insects13050406