
Could Sterile Aedes albopictus Male Releases Interfere with Aedes aegypti Population in Reunion Island?

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Preparation
2.2. Irradiation
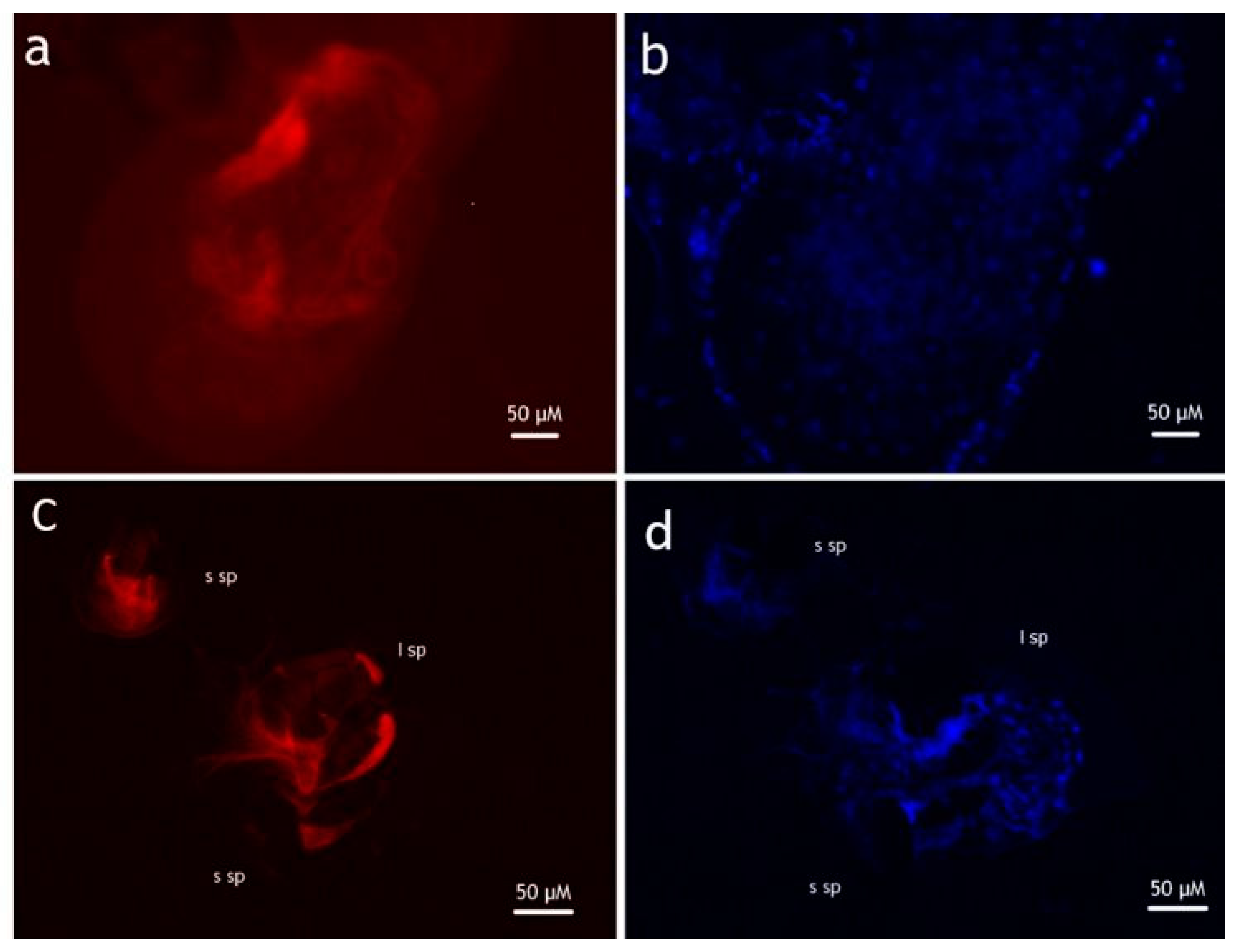
2.3. Rhodamine Marking
2.4. Mating Experiments
2.4.1. Transfer of Materials during Conspecific and Heterospecific Mating
2.4.2. Offspring Production after Conspecific and Heterospecific Mating
2.4.3. Influence of Heterospecific Mating on Subsequent Female Reproductive Behavior
2.5. Statistical Analysis
3. Results
3.1. Transfer of Materials during Conspecific and Heterospecific Mating
3.2. Offspring Production after Mating
3.3. Influence of Heterospecific Mating on the Later Female Reproductive Behavior
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salvan, M.; Mouchet, J. Aedes albopictus et Aedes aegypti a l’Ile de la Reunion. Ann. Soc. Belg. Med. Trop. 1994, 74, 323–326. [Google Scholar] [PubMed]
- Bagny, L.; Delatte, H.; Quilici, S.; Fontenille, D. Progressive decrease in Aedes aegypti distribution in Reunion Island since the 1900s. J. Med. Entomol. 2009, 46, 1541–1545. [Google Scholar] [CrossRef] [PubMed]
- Delatte, H.; Bagny, L.; Brengue, C.; Bouetard, A.; Paupy, C.; Fontenille, D. The invaders: Phylogeography of dengue and chikungunya viruses Aedes vectors, on the South West islands of the Indian Ocean. Infect. Genet. Evol. 2011, 11, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Burney, D.A.; Burney, L.P.; Godfrey, L.R.; Jungers, W.L.; Goodman, S.M.; Wright, H.T.; Timothy Jull, A.J. A chronology for late prehistoric Madagascar. J. Hum. Evol. 2004, 47, 25–63. [Google Scholar] [CrossRef] [PubMed]
- Nasci, R.S.; Hare, S.G.; Willis, F.S. Interspecific mating between Louisiana strains of Aedes albopictus and Aedes aegypti in the field and laboratory. J. Am. Mosq. Control Assoc. 1989, 5, 416–421. [Google Scholar] [PubMed]
- Lounibos, L.P. Invasion by insect vectors of human disease. Ann. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, L.; Kendell, D.; Robertson, D.; Livdahl, T.; Khatchikian, C. Aedes aegypti and Aedes albopictus in Bermuda: Extinction, invasion, invasion and extinction. Biol. Invasions 2010, 12, 3277–3288. [Google Scholar] [CrossRef]
- CIRE ARS Océan Indien. Le Point Epidémiologique N°4; CIRE ARS Océan Indien: Saint-Denis, France, 2018. [Google Scholar]
- CIRE ARS Océan Indien. Le Point Epidémiologique N°1; CIRE ARS Océan Indien: Saint-Denis, France, 2019. [Google Scholar]
- Delatte, H.; Dehecq, J.S.; Thiria, J.; Domerg, C.; Paupy, C.; Fontenille, D. Geographic distribution and developmental sites of Aedes albopictus (Diptera: Culicidae) during a Chikungunya epidemic event. Vector Borne Zoonotic Dis. 2008, 8, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Tripet, F.; Lounibos, L.P.; Robbins, D.; Moran, J.; Nishimura, N.; Blosser, E. Competitive reduction by satyrization? Evidence for interspecific mating in nature and asymmetric reproductive competition between invasive mosquito vectors. Am. J. Trop. Med. Hyg. 2011, 85, 265–270. [Google Scholar] [CrossRef]
- Bargielowski, I.E.; Lounibos, L.P.; Carrasquilla, M.C. Evolution of resistance to satyrization through reproductive character displacement in populations of invasive dengue vectors. Proc. Natl Acad. Sci. USA 2013, 110, 2888–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, J.M.C.; Spielman, A. The satyr effect: A model predicting parapatry and species extinction. Am. Nat. 1986, 128, 513–528. [Google Scholar] [CrossRef]
- Gröning, J.; Hochkirch, A. Reproductive interference between animal species. Q. Rev. Biol. 2008, 83, 257–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leahy, M.G.; Craig, G.B. Barriers to hybridization between Aedes aegypti and Aedes albopictus (Diptera: Culicidae). Evolution 1967, 21, 41–58. [Google Scholar] [PubMed]
- Carrasquilla, M.C.; Lounibos, L.P. Satyrization without evidence of successful insemination from interspecific mating between invasive mosquitoes. Biol. Lett. 2015, 11, 20150527. [Google Scholar] [CrossRef] [PubMed]
- Soghigian, J.; Gibbs, K.; Stanton, A.; Kaiser, R.; Livdahl, T. Sexual harassment and feeding inhibition between two invasive dengue vectors. Environ. Health Insights 2015, 8, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Bargielowski, I.; Blosser, M.; Lounibos, L.P. The effects of interspecific courtship on the mating success of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) males. Ann. Entomol. Soc. Am. 2015, 108, 513–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcela, P.; Hassan, A.A.; Hamdan, A.; Dieng, H.; Kumara, T.K. Interspecific Cross-Mating Between Aedes aegypti and Aedes albopictus Laboratory Strains: Implication of Population Density on Mating Behaviors. J. Am. Mosq. Control Assoc. 2015, 31, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.J.; Mitchell, S.N.; Paton, C.J.; Stevenson, J.; Staunton, K.M.; Snoad, N.; Beebe, N.; White, B.J.; Ritchie, S.A. Use of rhodamine B to mark the body and seminal fluid of male Aedes aegypti for mark-release-recapture experiments and estimating efficacy of sterile male releases. PLoS Negl. Trop. Dis. 2017, 11, e0005902. [Google Scholar] [CrossRef] [Green Version]
- Gouagna, L.C.; Damiens, D.; Oliva, C.; Boyer, S.; Le Goff, G.; Brengues, C.; Raude, J.; Simard, F.; Fontenille, D. Strategic Approach, Advances, and Challenges in the Development and Application of the SIT for Area-Wide Control of Aedes albopictus Mosquitoes in Reunion Island. Insects 2020, 11, 770. [Google Scholar] [CrossRef]
- Zheng, M.L.; Zhang, D.J.; Damiens, D.D.; Lees, R.S.; Gilles, J.R. Standard operating procedures for standardized mass rearing of the dengue and chikungunya vectors Aedes aegypti and Aedes albopictus (Diptera: Culicidae)-II-Egg storage and hatching. Parasites Vectors 2015, 8, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maïga, H.; Gilles, J.R.; Lees, R.S.; Yamada, H.; Bouyer, J. Demonstration of resistance to satyrization behavior in Aedes aegypti from La Réunion island. Parasite 2020, 27, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vick, K.W. Effects of interspecific matings of Trogoderma glabrum and T. inclusum on oviposition and remating. Ann. Entomol. Soc. Am. 1973, 66, 237–239. [Google Scholar] [CrossRef]
- Mori, A.; Romero-Severson, J.; Black IV, W.C.; Severson, D.W. Quantitative trait loci determining autogeny and body size in the Asian tiger mosquito (Aedes albopictus). Heredity 2008, 101, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazni, W.A.; Lee, H.L.; Dayang, H.A.B.; Azahari, A.H. Cross-mating between Malaysian strains of Aedes aegypti and Aedes albopictus in the laboratory. Southeast Asian J. Trop. Med. Public Health 2009, 40, 1. [Google Scholar]
- Ariani, C.V.; Smith, S.C.; Osei-Poku, J.; Short, K.; Juneja, P.; Jiggins, F.M. Environmental and genetic factors determine whether the mosquito Aedes aegypti lays eggs without a blood meal. Am. J. Trop. Med. Hyg. 2015, 92, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.P.; Paulson, S.L. Reproductive isolation between Florida strains of Aedes aegypti and Aedes albopictus. J. Am. Mosq. Control Assoc. 1994, 10, 88–92. [Google Scholar]
- Lee, H.L.; Aramu, M.; Nazni, W.A.; Selvi, S.; Vasan, S. No evidence for successful interspecific cross-mating of transgenic Aedes aegypti (L.) and wild type Aedes albopictus Skuse. Trop. Biomed. 2009, 26, 312–319. [Google Scholar]
- Klowden, M.J. Mating and nutritional state affect the reproduction of Aedes albopictus mosquitoes. J. Am. Mosq. Control Assoc. 1993, 9, 169–173. [Google Scholar]
- Helinski, M.E.H.; Deewatthanawong, P.; Sirot, L.K.; Wolfner, M.F.; Harrington, L.C. Duration and dose-dependency of female sexual receptivity responses to seminal fluid proteins in Aedes albopictus and Ae. aegypti mosquitoes. J. Insect Physiol. 2012, 58, 1307–1313. [Google Scholar] [CrossRef] [Green Version]
- Lima-Camara, T.N.; Lima, J.B.P.; Bruno, R.V.; Peixoto, A.A. Effects of insemination and blood-feeding on locomotor activity of Aedes albopictus and Aedes aegypti (Diptera: Culicidae) females under laboratory conditions. Parasites Vectors 2014, 7, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spielman, A. The mechanics of copulation in Aedes aegypti. Biol. Bull. 1964, 127, 324–344. [Google Scholar] [CrossRef]
- Hiss, E.A.; Fuchs, M.S. The effect of matrone on oviposition in the mosquito Aedes aegypti. J. Insect Physiol. 1972, 18, 2217–2227. [Google Scholar] [CrossRef]
- Lima-Camara, T.N.; Codeço, C.T.; Honório, N.A.; Bruno, R.V.; Peixoto, A.A.; Lounibos, L.P. Male accessory gland substances from Aedes albopictus affect the locomotor activity of Aedes aegypti females. Mem. Inst. Oswaldo Cruz. 2013, 108, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Duvall, L.B.; Basrur, N.S.; Molina, H.; McMeniman, C.J.; Vosshall, L.B. A neuropeptide signaling system that rapidly enforces paternity in the Aedes aegypti mosquito. Curr. Biol. 2017, 4, 3734–3742. [Google Scholar] [CrossRef] [Green Version]
- Hartberg, W.K. Observations on the mating behaviour of Aedes aegypti in nature. Bull. World Health Organ. 1971, 45, 847–850. [Google Scholar] [PubMed]
- Paton, R.S.; Bonsall, M.B. The ecological and epidemiological consequences of reproductive interference between the vectors Aedes aegypti and Aedes albopictus. J. R. Soc. Interface 2019, 16, 20190270. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Rhodamine Marked Males | Female | Mean Percent of Marked Females | Post Hoc Test Tukey for Marked Females | Mean Percent of Females with Sperm |
|---|---|---|---|---|
| Ae. albopictus | Ae. albopictus | 100 ± 0.0 | a | 100 ± 0.0 |
| Ae. albopictus sterile | Ae. albopictus | 91.1 ± 3.1 | b | 100 ± 0.0 |
| Ae. aegypti | Ae. aegypti | 100 ± 0.0 | a | 100 ± 0.0 |
| Ae. albopictus | Ae. aegypti | 14.9 ± 6.5 | c | 0 ± 0.0 |
| Ae. albopictus sterile | Ae. aegypti | 4.2 ± 4.2 | c | 0 ± 0.0 |
| Ae. aegypti | Ae. albopictus | 17.9 ± 10.4 | c | 0 ± 0.0 |
| Rhodamine Marked Males | Female | Mean Number of Eggs | Post Hoc Test Tukey for Number of Eggs | Mean Hatch Rate | Post Hoc Test Tukey for Hatch Rate |
|---|---|---|---|---|---|
| - | Ae. albopictus | 0.0 ± 0.0 | b | 0.0 ± 0.0 | c |
| - | Ae. aegypti | 20.3 ± 35.2 | b | 0.0 ± 0.0 | c |
| Ae. albopictus | Ae. albopictus | 302.0 ± 164.0 | a | 84.7 ± 1.9 | a |
| Ae. albopictus sterile | Ae. albopictus | 254.7 ± 48.2 | a | 0.1 ± 0.2 | c |
| Ae. aegypti | Ae. aegypti | 183.0 ± 83.5 | a | 70.0 ± 3.9 | b |
| Ae. albopictus | Ae. aegypti | 5.33 ± 5.5 | b | 0.0 ± 0.0 | c |
| Ae. albopictus sterile | Ae. aegypti | 0.0 ± 0.0 | b | 0.0 ± 0.0 | c |
| Ae. aegypti | Ae. albopictus | 11.0 ± 11.0 | b | 0.0 ± 0.0 | c |
| Rhodamine Marked Males | Females | Males | Mean Number of Eggs | Hatch Rate |
|---|---|---|---|---|
| First mating | Second mating | |||
| Ae. albopictus | Ae. albopictus | 302.0 ± 164.0 | 84.7 ± 2.0 | |
| Ae. aegypti | Ae. albopictus | Ae. albopictus | 303.3 ± 60.8 | 86.1 ± 8.2 |
| Ae. aegypti | Ae. aegypti | 183.0 ± 83.5 | 70.3 ± 3.9 | |
| Ae. albopictus | Ae. aegypti | Ae. aegypti | 130.3 ± 34.0 | 76.5 ± 3.3 |
| sterile Ae. albopictus | Ae. aegypti | Ae. aegypti | 130.0 ± 29.9 | 70.3 ± 9.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrianjakarivony, H.F.; Damiens, D.; Marquereau, L.; Gaudillat, B.; Habchi-Hanriot, N.; Gouagna, L.-C. Could Sterile Aedes albopictus Male Releases Interfere with Aedes aegypti Population in Reunion Island? Insects 2022, 13, 146. https://doi.org/10.3390/insects13020146
Andrianjakarivony HF, Damiens D, Marquereau L, Gaudillat B, Habchi-Hanriot N, Gouagna L-C. Could Sterile Aedes albopictus Male Releases Interfere with Aedes aegypti Population in Reunion Island? Insects. 2022; 13(2):146. https://doi.org/10.3390/insects13020146
Chicago/Turabian StyleAndrianjakarivony, Harilanto Felana, David Damiens, Lucie Marquereau, Benjamin Gaudillat, Nausicaa Habchi-Hanriot, and Louis-Clément Gouagna. 2022. "Could Sterile Aedes albopictus Male Releases Interfere with Aedes aegypti Population in Reunion Island?" Insects 13, no. 2: 146. https://doi.org/10.3390/insects13020146