
Different Performance of Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae) among Four Potato Tuber Varieties under Laboratory Condition


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Potato Tubers
2.3. Life History Characteristics of P. operculella Cultured on Four Potato Tuber Varieties
2.4. Pupal Weight of P. operculella on Four Different Potato Tuber Varieties
2.5. Oviposition Preference of P. operculella Females on Four Different Potato Tuber Varieties
2.6. Feeding Preference of P. operculella Larvae on Four Different Potato Tuber Varieties
2.7. Data Analysis
3. Results
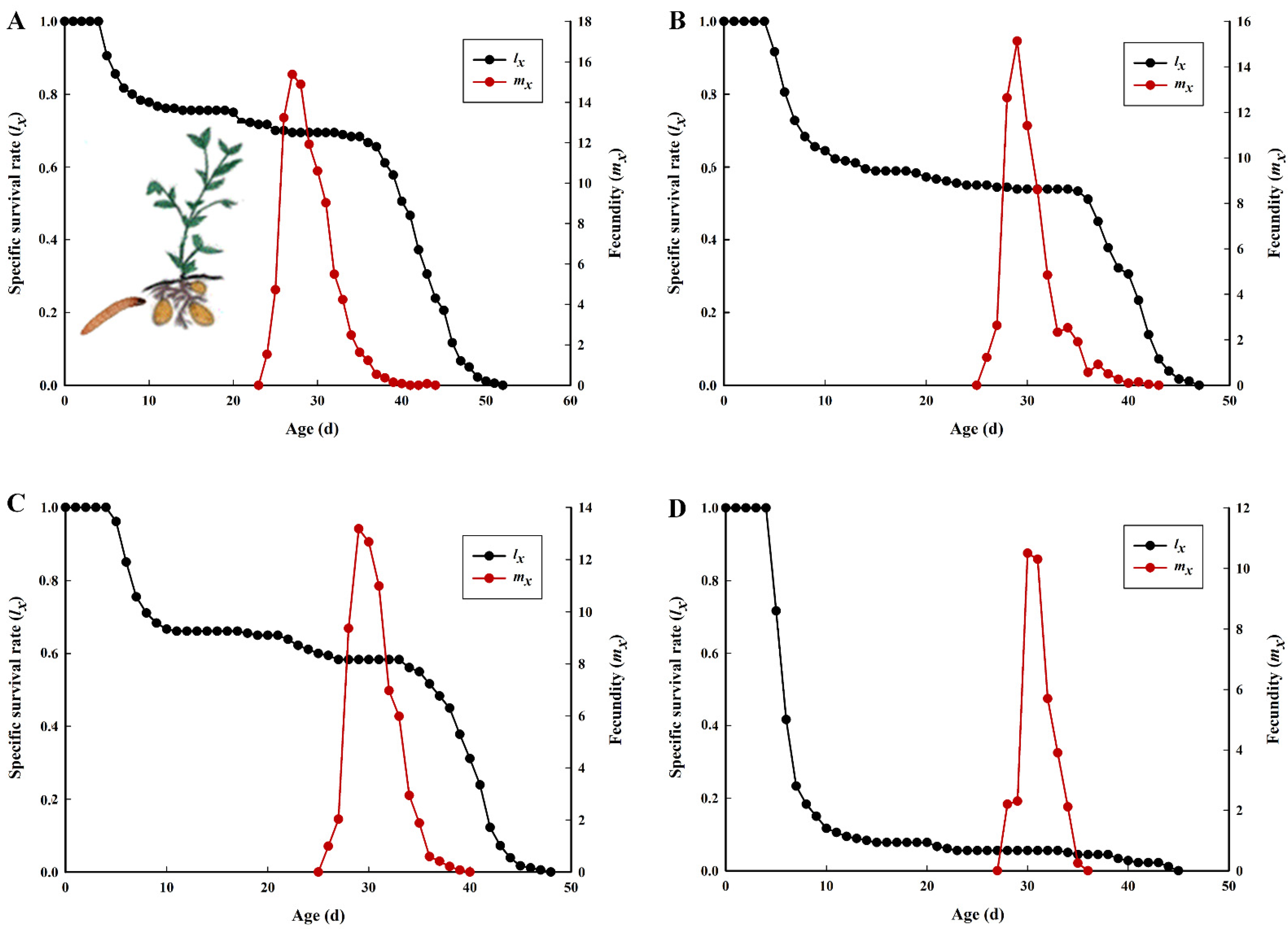
3.1. Development, Survivorship, and Reproduction of P. operculella on Four Different Potato Tuber Varieties
3.2. Population Parameters of P. operculella on Four Different Potato Tuber Varieties
3.3. Pupal Weight of P. operculella on Four Different Potato Tuber Varieties
3.4. Oviposition Preference of P. operculella on Four Different Potato Tuber Varieties
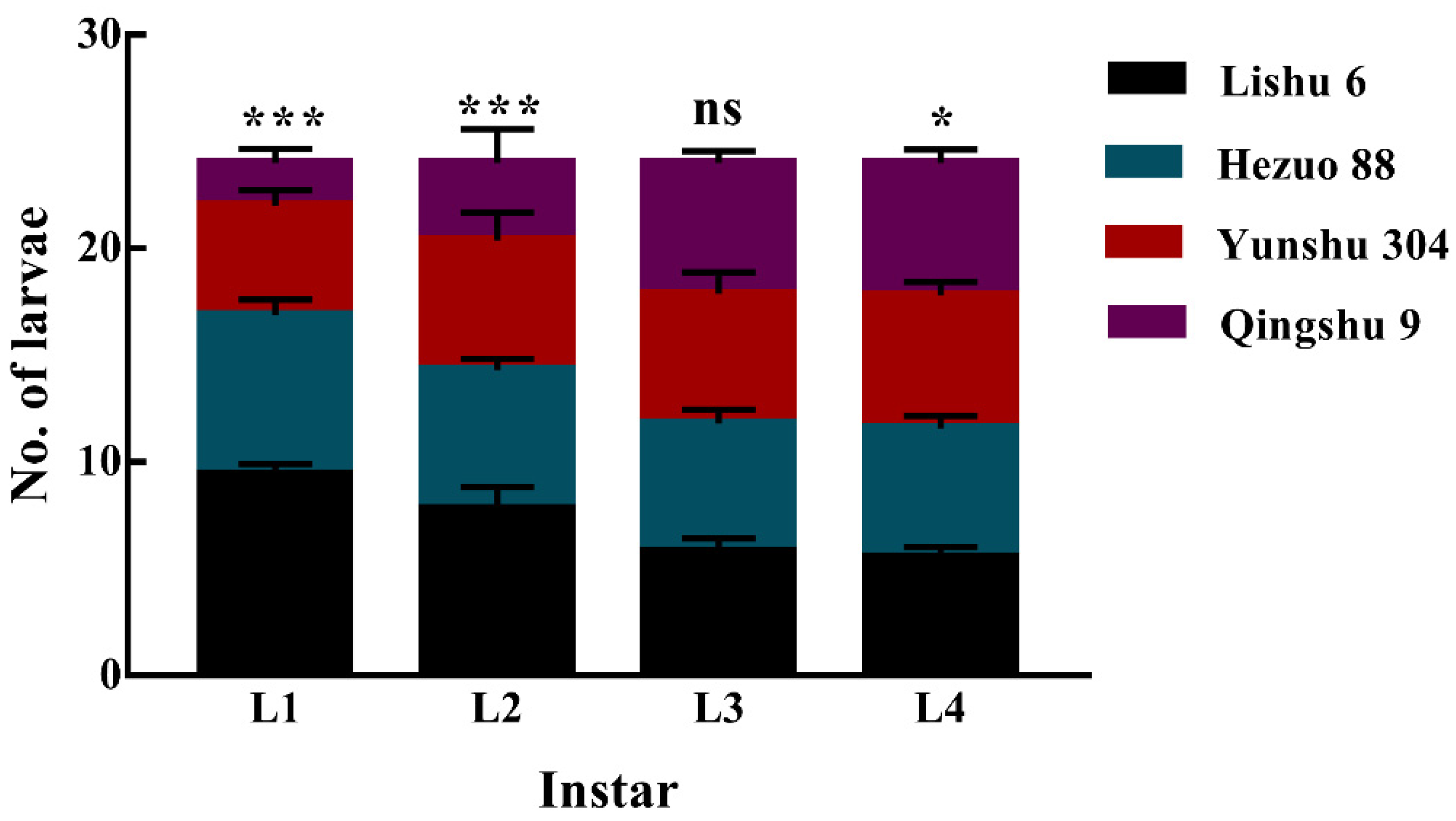
3.5. Feeding Preference of P. operculella Larvae on Four Different Potato Tuber Varieties
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gao, Y.L. Potato tuberworm: Impact and methods for control—Mini Review. CAB. Rev. 2018, 13, 1–3. [Google Scholar]
- Yan, J.J.; Zhang, M.D.; Gao, Y.L. Biology, ecology and integrated management of the potato tuber moth, Phthorimaea operculella (Lepidoptera: Gelechiidae). Acta Entomol. Sin. 2019, 62, 1469–1482. (In Chinese) [Google Scholar]
- Yuan, H.G.; Wu, S.Y.; Lei, Z.R.; Rondon, S.I.; Gao, Y.L. Sub-lethal effects of Beauveria bassiana (Balsamo) on field populations of the potato tuberworm Phthorimaea operculella Zeller in China. J. Integr. Agric. 2018, 17, 911–918. [Google Scholar] [CrossRef] [Green Version]
- Rondon, S.I.; Hane, D.C.; Brown, C.R.; Vales, M.I.; Dogramaci, M. Resistance of potato germplasm to the potato tuberworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2009, 102, 1649–1653. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, J.H.; Yang, Y.L.; Tang, H.; Lv, H.P.; Fan, M.T.; Shi, Y.; Dong, D.F.; Wang, G.J.; Wang, W.X.; et al. Status of major diseases and insect pests of potato and pesticide usage in China. Sci. Agric. Sin. 2019, 52, 2800–2808. (In Chinese) [Google Scholar]
- Gao, Y.L. Green control techniques for potato tuberworm (Phthorimaea operculella). Sci. Agric. Sin. 2021, 54, 533–535. (In Chinese) [Google Scholar]
- Yan, J.J.; Mei, X.D.; Feng, J.W.; Lin, Z.X.; Reitz, S.; Meng, R.X.; Gao, Y.L. Optimization of the sex pheromone-based method for trapping field populations of Phthorimaea operculella (Zeller) in South China. J. Integr. Agric. 2020, 19, 2–8. [Google Scholar]
- Graft, J.E. The potato tubermoth. Tech. Bull. USDA 1917, 427, 58. [Google Scholar]
- Arnone, S.; Musmeci, S.; Bacchetta, L.; Cordischi, N.; Pucci, E.; Cristofaro, M.; Sonnino, A. Research in Solanum spp. of sources of resistance to the potato tuber moth Phthorimaea operculella (Zeller). Potato Res. 1998, 41, 39–49. [Google Scholar] [CrossRef]
- Gao, Y.L. Potato tuberworm: A threat for China potatoes. Entomol. Ornithol. Herpetol. 2018, 7, e132. [Google Scholar] [CrossRef]
- Vaneva-Gancheva, T.; Dimitrov, Y. Chemical control of the potato tuber moth Phthorimaea operculella (Zeller) on tobacco. Bulg. J. Agric. Sci. 2013, 19, 1003–1008. [Google Scholar]
- Dekebol, A.; Aryal, S.; Jung, C. Suitability of tomato leaves for larval development of Potato tuber moth, Phthorimaea operculella (Zeller) (Lepidoptera: Gelechiidae). Entomol. Res. 2019, 49, 258–264. [Google Scholar] [CrossRef]
- Zhang, M.D.; Yan, J.J.; Gao, Y.L. The adaptive analysis of Phthorimaea operculella to different potato tuber varieties. Sci. Agric. Sin. 2021, 54, 536–546. [Google Scholar]
- Mansouri, S.M.; Fathi, S.A.A.; Nouri-Ganbalani, G.; Razmjou, J.; Naseri, B.; Rondon, S.I. Screening of Iranian potato germplasm for resistance to the potato tuberworm Phthorimaea operculella (Lepidoptera: Gelechiidae). Am. J. Potato Res. 2013, 90, 533–540. [Google Scholar] [CrossRef]
- Gui, F.R.; Li, Z.Y. A method for rearing the potato tuber moth Phthorimaea operculella on potato. Entomol. Knowl. 2003, 40, 187–189. [Google Scholar]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2021, 28, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.Y.; Wang, Z.Y.; He, K.L.; Cong, B. Feeding behavior of Ostrina furnacalis larvae on transgenic Bt corn expressing Cry1Ab toxin. Bull. Entomol. Res. 2005, 42, 270–274. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sex and variable development rate among individuals. Environ. Entomol. 1988, 17, 26–31. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age Stage, Two-Sex Life Table Analysis. Available online: http://140.120.197.173/Ecology/Download/Twosex-MSChart-exe-B100000.rar (accessed on 2 November 2020).
- Chi, H.; Liu, H. Two new methods for study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Johnson, R.W. An introduction to the bootstrap. Teach. Stats. 2010, 23, 49–54. [Google Scholar] [CrossRef]
- Huang, Y.B.; Chi, H. Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci. 2012, 19, 263–273. [Google Scholar] [CrossRef]
- Chi, H.; Yang, T.C. Two-sex life table and predation rate of Propylaea japonica Thunberg (Coleoptera: Coccinellidae) fed on Myzus persicae (Sulzer) (Homoptera: Aphididae). Environ. Entomol. 2003, 32, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Foot, M.A. Susceptibility of twenty potato germplasms to the potato tuber moth at Pukekohe a preliminary assessment. N. Z. J. Exp. Agric. 1976, 4, 239–242. [Google Scholar] [CrossRef]
- Raman, K.V.; Palacios, M. Screening potato for resistance to potato tuberworm. J. Econ. Entomol. 1982, 75, 47–49. [Google Scholar] [CrossRef]
- Malakar-Kuenen, R.; Tingey, W.M. Aspects of tuber resistance in hybrid potatoes to potato tuber worm. Entomol. Exp. Appl. 2006, 120, 131–137. [Google Scholar] [CrossRef]
- Akca, I.; Ayvaz, T.; Yazici, E.; Smith, C.L.; Chi, H. Demography and population projection of Aphis fabae (Hemiptera: Aphididae): With additional comments on life table research criteria. J. Econ. Entomol. 2015, 108, 1466–1478. [Google Scholar] [CrossRef] [PubMed]
- Tuan, S.J.; Yeh, C.C.; Atlihan, R.; Chi, H.; Tang, L.C. Demography and consumption of Spodoptera litura (Lepidoptera: Noctuidae) reared on cabbage and taro. J. Econ. Entomol. 2016, 109, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Garcia-Robledo, C.; Horvitz, C.C.; Staines, C.L. Larval morphology, development, and notes on the natural history of Cephaloleia “rolled-leaf” beetles (Coleoptera: Chrysomelidae: Cassidinae). Zootaxa 2010, 2610, 50–68. [Google Scholar] [CrossRef]
- Rojas, J.C.; Kolomiets, M.V.; Bernal, J.S.; Luis, J.F.J. Nonsensical choices? Fall armyworm moths choose seemingly best or worst hosts for their larvae, but neonate larvae make their own choices. PLoS ONE 2018, 13, e0197628. [Google Scholar] [CrossRef]
- Bernays, E.; Graham, M. On the evolution of host specificity in phytophagous arthropods. Ecology 1988, 69, 886–892. [Google Scholar] [CrossRef]
- Horgan, F.G.; Quiring, D.T.; Lagnaoui, A.; Salas, A.R.; Pelletier, Y. Periderm- and cortex-based resistance to tuber-feeding Phthorimaea operculella in two wild potato species. Entomol. Exp. Appl. 2010, 125, 249–258. [Google Scholar] [CrossRef]
- Das, G.P.; Raman, K.V. Alternate hosts of the potato tuber moth, Phthorimaea operculella (Zeller). Crop. Prot. 1994, 13, 83–86. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Stage | Lishu 6 | Hezuo 88 | Yunshu 304 | Qingshu 9 | ||||
|---|---|---|---|---|---|---|---|---|
| n | Developmental Time (d) | n | Developmental Time (d) | n | Developmental Time (d) | n | Developmental Time (d) | |
| Egg | 180 | 4.0 ± 0.0 | 180 | 4.0 ± 0.0 | 180 | 4.0 ± 0.0 | 180 | 4.0 ± 0.0 |
| larval | 180 | 14.18 ± 0.14 b | 180 | 14.30 ± 0.11 b | 180 | 14.37 ± 0.06 b | 180 | 15.50 ± 0.34 a |
| Pupa | 136 | 8.69 ± 0.09 c | 106 | 9.07 ± 0.07 b | 118 | 9.30 ± 0.09 a | 14 | 9.90 ± 0.31 a |
| Female | 58 | 14.28 ± 0.56 a | 44 | 13.33 ± 2.30 a,b | 50 | 10.74 ± 0.32 b | 5 | 9.40 ± 1.05 b |
| Male | 68 | 18.03 ± 0.27 a | 54 | 14.72 ± 0.22 b | 55 | 14.64 ± 0.24 b | 5 | 12.80 ± 1.85 b |
| Mean fecundity/egg | 58 | 213.67 ± 7.03 a | 44 | 147.40 ± 5.71 b | 50 | 142.54 ± 5.85 b | 5 | 74.00 ± 6.26 c |
| Parameter | Lishu 6 | Hezuo 88 | Yunshu 304 | Qingshu 9 |
|---|---|---|---|---|
| Intrinsic rate of increase, r (d−1) | 0.1434 ± 0.0042 a | 0.1156 ± 0.0046 b | 0.1180 ± 0.0042 b | 0.0226 ± 0.0159 c |
| Finite rate of increase, λ (d−1) | 1.1542 ± 0.0048 a | 1.1226 ± 0.0051 b | 1.1253 ± 0.0047 b | 1.0228 ± 0.0161 c |
| Net reproductive rate, R0 (offspring) | 67.6611 ± 7.7254 a | 35.2111 ± 4.8706 b | 39.5944 ± 4.9943 b | 2.0556 ± 0.9100 c |
| Mean generation time, T (d) | 29.3945 ± 0.2516 c | 30.7987 ± 0.2072 b | 31.1639 ± 0.1834 a,b | 31.9294 ± 1.1317 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Yan, J.; Ali, A.; Gao, Y. Different Performance of Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae) among Four Potato Tuber Varieties under Laboratory Condition. Insects 2021, 12, 580. https://doi.org/10.3390/insects12070580
Zhang M, Yan J, Ali A, Gao Y. Different Performance of Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae) among Four Potato Tuber Varieties under Laboratory Condition. Insects. 2021; 12(7):580. https://doi.org/10.3390/insects12070580
Chicago/Turabian StyleZhang, Mengdi, Junjie Yan, Abid Ali, and Yulin Gao. 2021. "Different Performance of Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae) among Four Potato Tuber Varieties under Laboratory Condition" Insects 12, no. 7: 580. https://doi.org/10.3390/insects12070580