
Polyphenols as Food Supplement Improved Food Consumption and Longevity of Honey Bees (Apis mellifera) Intoxicated by Pesticide Thiacloprid

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bees
2.2. Chemicals
2.3. Design of the Experiment
- Treatment TL—sucrose solution with a low dosage of Thiacloprid (35 mg/L).
- Treatment FTL—sucrose solution (50% w/v) with a mixture of phenolic compounds and low dosage of Thiacloprid (35 mg/L).
- Treatment TH—sucrose solution (50% w/v) with high dosage of Thiacloprid (70 mg/L).
- Treatment FTH—sucrose solution (50% w/v) with a mixture of phenolic compounds and a high dosage of Thiacloprid (70 mg/L).
- Treatment F—sucrose solution (50% w/v) and a mixture of phenolic compounds.
- Treatment C—sucrose solution (50% w/v).
2.4. RNA Isolation and RT-qPCR
2.5. Data Analyses
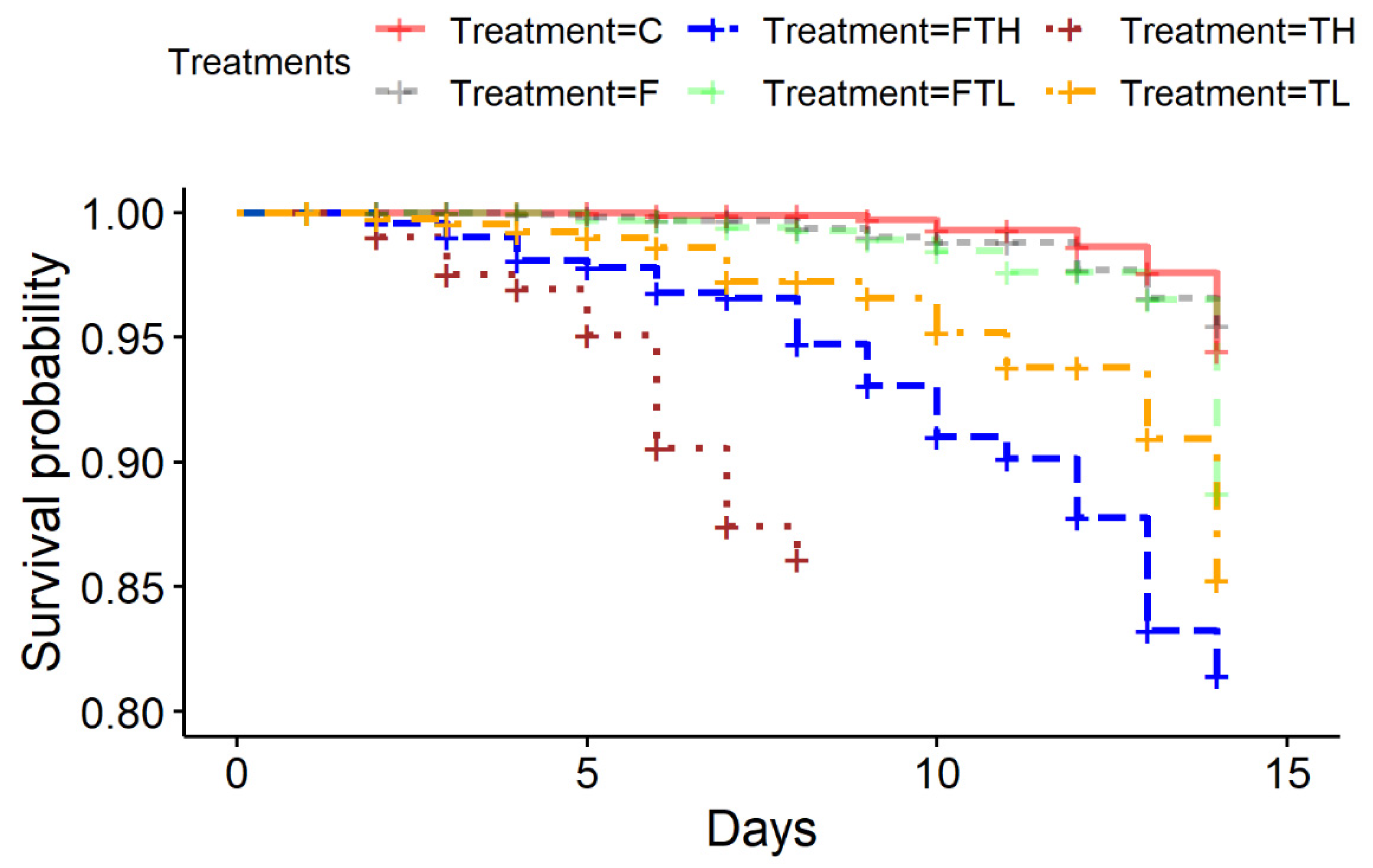
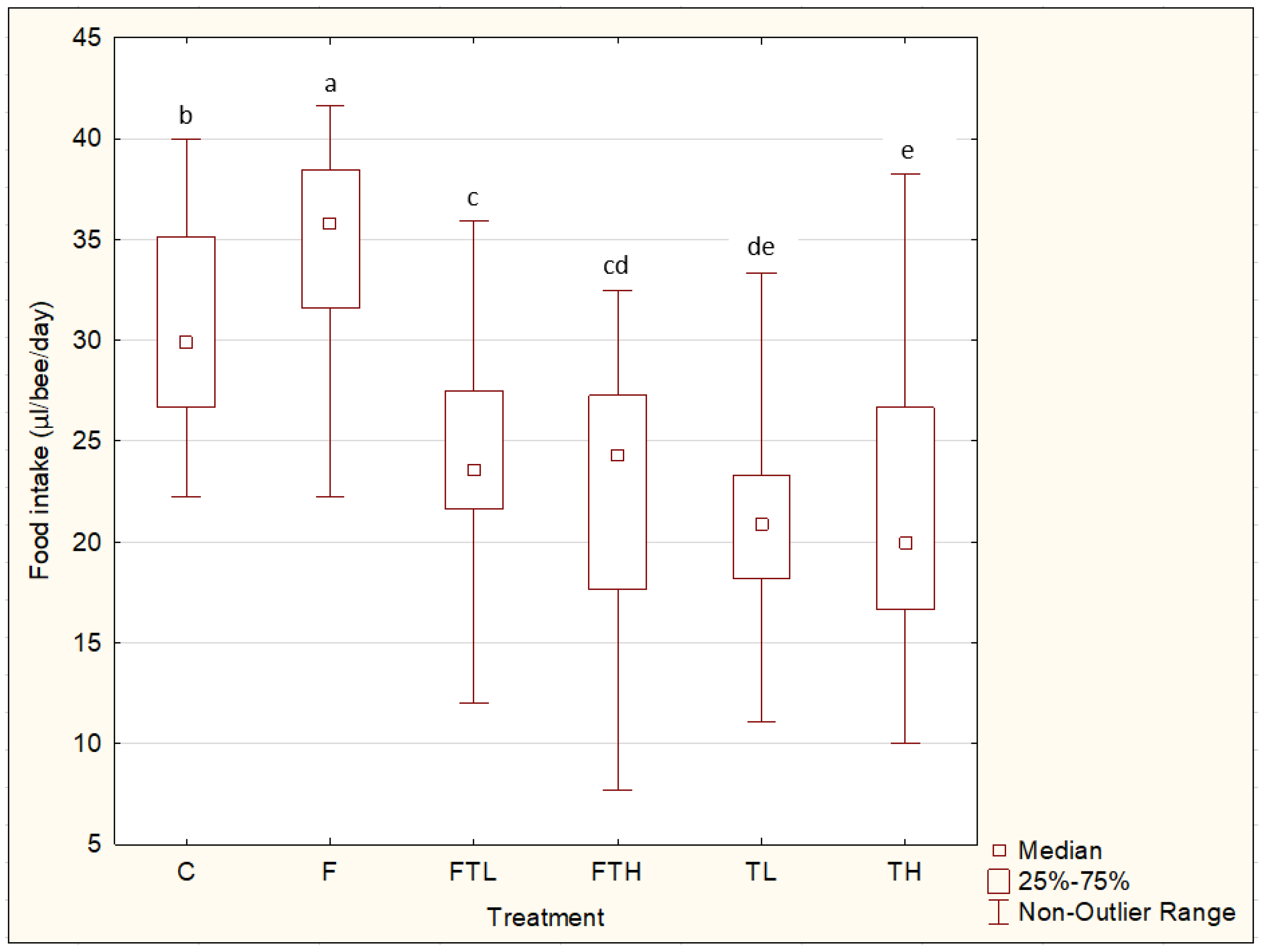
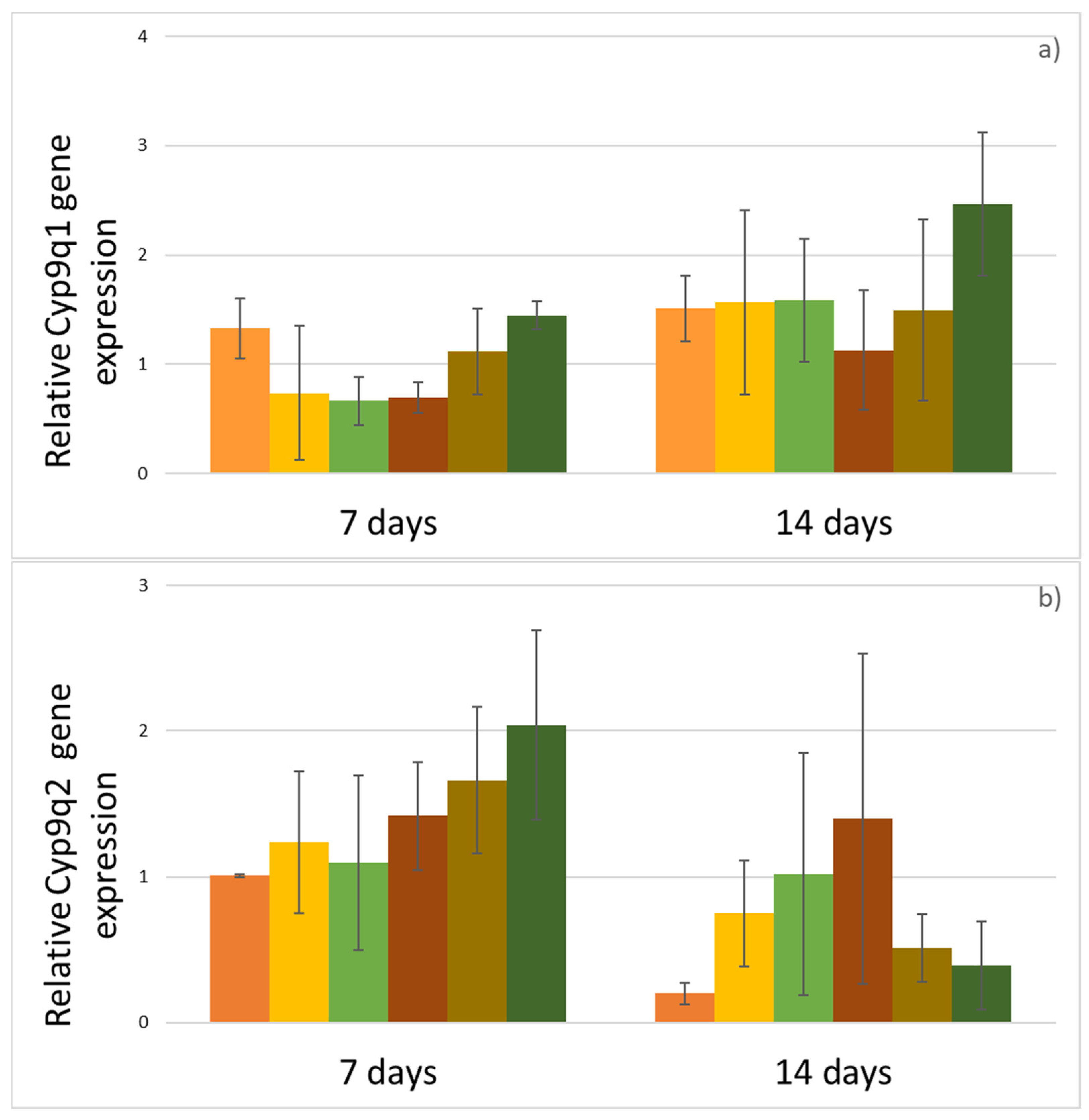
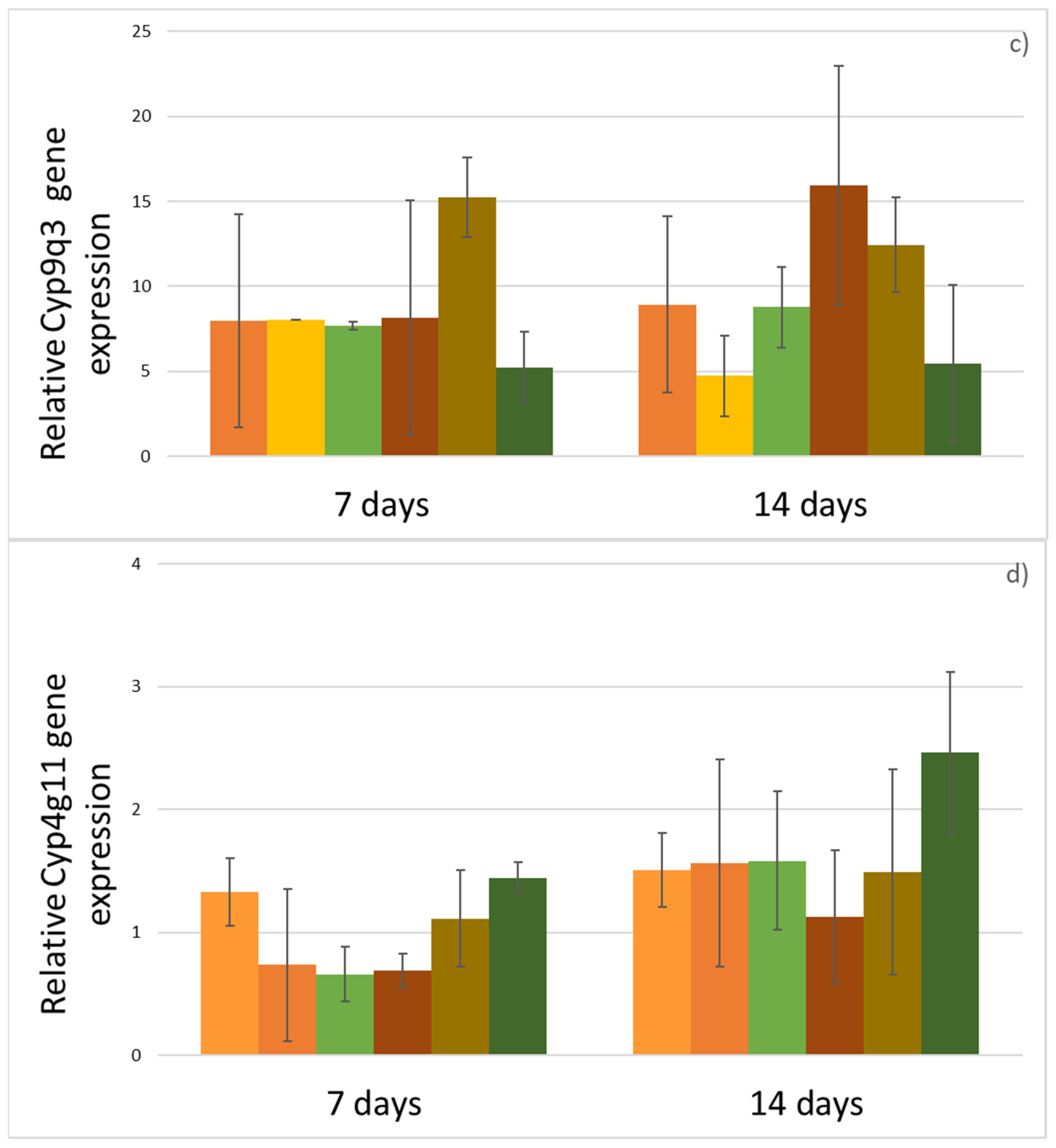
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Ann. Bot. 2009, 103, 1579–1588. [Google Scholar] [CrossRef]
- Stokstad, E. The Case of the Empty Hives. Science 2007, 316, 970–972. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and Health in Honey Bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharpe, R.J.; Heyden, L.C. Honey Bee Colony Collapse Disorder Is Possibly Caused by a Dietary Pyrethrum Deficiency. Biosci. Hypotheses 2009, 2, 439–440. [Google Scholar] [CrossRef]
- Haydak, M.H. Honey Bee Nutrition. Annu. Rev. Entomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Naug, D. Nutritional Stress Due to Habitat Loss May Explain Recent Honeybee Colony Collapses. Biol. Conserv. 2009, 142, 2369–2372. [Google Scholar] [CrossRef]
- Johnson, R.M.; Ellis, M.D.; Mullin, C.A.; Frazier, M. Pesticides and Honey Bee Toxicity—USA. Apidologie 2010, 41, 312–331. [Google Scholar] [CrossRef] [Green Version]
- Decourtye, A.; Mader, E.; Desneux, N. Landscape Enhancement of Floral Resources for Honey Bees in Agro-Ecosystems. Apidologie 2010, 41, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Manning, R.; Rutkay, A.; Eaton, L.; Dell, B. Lipid-enhanced Pollen and Lipid-reduced Flour Diets and Their Effect on the Longevity of Honey Bees (Apis mellifera L.). Aust. J. Entomol. 2007, 46, 251–257. [Google Scholar] [CrossRef]
- Hayes, J., Jr.; Underwood, R.M.; Pettis, J. A Survey of Honey Bee Colony Losses in the US, Fall 2007 to Spring 2008. PLoS ONE 2008, 3, e4071. [Google Scholar]
- Wahl, O.; Ulm, K. Influence of Pollen Feeding and Physiological Condition on Pesticide Sensitivity of the Honey Bee Apis mellifera Carnica. Oecologia 1983, 59, 106–128. [Google Scholar] [CrossRef]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Diet Effects on Honeybee Immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, I.; Fluri, P.; Imdorf, A. Pollen Nutrition and Colony Development in Honey Bees—Part II. Bee World 2005, 86, 27–34. [Google Scholar] [CrossRef]
- Frazier, M.; Mullin, C.; Frazier, J.; Ashcraft, S. What Have Pesticides Got to Do with It? Am. Bee J. 2008, 148, 521–524. [Google Scholar]
- Medrzycki, P.; Sgolastra, F.; Bortolotti, L.; Bogo, G.; Tosi, S.; Padovani, E.; Porrini, C.; Sabatini, A.G. Influence of Brood Rearing Temperature on Honey Bee Development and Susceptibility to Poisoning by Pesticides. J. Apic. Res. 2010, 49, 52–59. [Google Scholar] [CrossRef]
- Iwasa, T.; Motoyama, N.; Ambrose, J.T.; Roe, R.M. Mechanism for the Differential Toxicity of Neonicotinoid Insecticides in the Honey Bee, Apis mellifera. Crop Prot. 2004, 23, 371–378. [Google Scholar] [CrossRef]
- Alaux, C.; Brunet, J.; Dussaubat, C.; Mondet, F.; Tchamitchan, S.; Cousin, M.; Brillard, J.; Baldy, A.; Belzunces, L.P.; Le Conte, Y. Interactions between Nosema Microspores and a Neonicotinoid Weaken Honeybees (Apis mellifera). Environ. Microbiol. 2010, 12, 774–782. [Google Scholar] [CrossRef] [Green Version]
- Retschnig, G.; Neumann, P.; Williams, G.R. Thiacloprid–Nosema Ceranae Interactions in Honey Bees: Host Survivorship but Not Parasite Reproduction Is Dependent on Pesticide Dose. J. Invertebr. Pathol. 2014, 118, 18–19. [Google Scholar] [CrossRef]
- Suchail, S.; Guez, D.; Belzunces, L.P. Characteristics of Imidacloprid Toxicity in Two Apis mellifera Subspecies. Environ. Toxicol. Chem. 2000, 19, 1901–1905. [Google Scholar] [CrossRef] [Green Version]
- Nauen, R.; Ebbinghaus-Kintscher, U.; Schmuck, R. Toxicity and Nicotinic Acetylcholine Receptor Interaction of Imidacloprid and Its Metabolites in Apis mellifera (Hymenoptera: Apidae). Pest Manag. Sci. 2001, 57, 577–586. [Google Scholar] [CrossRef]
- Wehling, M.; Ohe, W.; Brasse, D.; Forster, R. Colony Losses-Interactions of Plant Protection Products and Other Factors. Jul. Kühn Arch. 2009, 423, 153–154. [Google Scholar]
- Grillone, G.; Laurino, D.; Manino, A.; Porporato, M. Toxicity of Thiametoxam on in vitro Reared Honey Bee Brood. Apidologie 2017, 48, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Osterman, J.; Wintermantel, D.; Locke, B.; Jonsson, O.; Semberg, E.; Onorati, P.; Forsgren, E.; Rosenkranz, P.; Rahbek-Pedersen, T.; Bommarco, R. Clothianidin Seed-Treatment Has No Detectable Negative Impact on Honeybee Colonies and Their Pathogens. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rundlöf, M.; Lundin, O. Can Costs of Pesticide Exposure for Bumblebees Be Balanced by Benefits from a Mass-Flowering Crop? Environ. Sci. Technol. 2019, 53, 14144–14151. [Google Scholar] [CrossRef]
- Ghazoul, J. Buzziness as Usual? Questioning the Global Pollination Crisis. Trends Ecol. Evol. 2005, 20, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Alburaki, M.; Boutin, S.; Mercier, P.-L.; Loublier, Y.; Chagnon, M.; Derome, N. Neonicotinoid-Coated Zea Mays Seeds Indirectly Affect Honeybee Performance and Pathogen Susceptibility in Field Trials. PLoS ONE 2015, 10, e0125790. [Google Scholar] [CrossRef] [Green Version]
- Claudianos, C.; Ranson, H.; Johnson, R.; Biswas, S.; Schuler, M.; Berenbaum, M.; Feyereisen, R.; Oakeshott, J.G. A Deficit of Detoxification Enzymes: Pesticide Sensitivity and Environmental Response in the Honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.; Schuler, M.; Berenbaum, M. Task-related Differential Expression of Four Cytochrome P450 Genes in Honeybee Appendages. Insect Mol. Biol. 2015, 24, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Honey Constituents Up-Regulate Detoxification and Immunity Genes in the Western Honey Bee Apis mellifera. Proc. Natl. Acad. Sci. USA 2013, 110, 8842–8846. [Google Scholar] [CrossRef] [Green Version]
- Liao, L.-H.; Wu, W.-Y.; Berenbaum, M.R. Impacts of Dietary Phytochemicals in the Presence and Absence of Pesticides on Longevity of Honey Bees (Apis mellifera). Insects 2017, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Bravo Lamas, L.; Martínez Flórez, S.; Agudo Toyos, P. Phenolic Compounds in Honey and Their Associated Health Benefits: A Review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef] [Green Version]
- Moniruzzaman, M.; Yung An, C.; Rao, P.V.; Hawlader, M.N.I.; Azlan, S.A.B.M.; Sulaiman, S.A.; Gan, S.H. Identification of Phenolic Acids and Flavonoids in Monofloral Honey from Bangladesh by High Performance Liquid Chromatography: Determination of Antioxidant Capacity. BioMed Res. Int. 2014, 2014, 737490. [Google Scholar] [CrossRef]
- Williams, G.R.; Alaux, C.; Costa, C.; Csaki, T.; Doublet, V.; Eisenhardt, D.; Brodschneider, R. Standard Methods for Maintaining Adult Apis mellifera in Cages under in Vitro Laboratory Conditions. J. Apic. Res. 2013, 52, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, A.P.; Mackert, A.; dos Santos Cristino, A.; Simões, Z.L.P. Validation of Reference Genes for Gene Expression Studies in the Honey Bee, Apis mellifera, by Quantitative Real-Time RT-PCR. Apidologie 2008, 39, 372–385. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. CYP9Q-Mediated Detoxification of Acaricides in the Honey Bee (Apis mellifera). Proc. Natl. Acad. Sci. USA 2011, 108, 12657–12662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calla, B.; MacLean, M.; Liao, L.; Dhanjal, I.; Tittiger, C.; Blomquist, G.J.; Berenbaum, M.R. Functional Characterization of CYP4G11—a Highly Conserved Enzyme in the Western Honey Bee Apis mellifera. Insect Mol. Biol. 2018, 27, 661–674. [Google Scholar] [CrossRef]
- Tesovnik, T.; Cizelj, I.; Zorc, M.; Čitar, M.; Božič, J.; Glavan, G.; Narat, M. Immune Related Gene Expression in Worker Honey Bee (Apis mellifera Carnica) Pupae Exposed to Neonicotinoid Thiamethoxam and Varroa Mites (Varroa Destructor). PLoS ONE 2017, 12, e0187079. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Therneau, T.M.; Grambsch, P.M. The cox model. In Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000; pp. 39–77. [Google Scholar]
- Yu, S.J. Interactions of Allelochemicals with Detoxication Enzymes of Insecticide-Susceptible and Resistant Fall Armyworms. Pestic. Biochem. Physiol. 1984, 22, 60–68. [Google Scholar] [CrossRef]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Disruption of Quercetin Metabolism by Fungicide Affects Energy Production in Honey Bees (Apis mellifera). Proc. Natl. Acad. Sci. USA 2017, 114, 2538–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porrini, M.P.; Fernández, N.J.; Garrido, P.M.; Gende, L.B.; Medici, S.K.; Eguaras, M.J. In Vivo Evaluation of Antiparasitic Activity of Plant Extracts on Nosema Ceranae (Microsporidia). Apidologie 2011, 42, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.; Nieh, J.C.; Tosi, S. Combined Nutritional Stress and a New Systemic Pesticide (Flupyradifurone, Sivanto®) Reduce Bee Survival, Food Consumption, Flight Success, and Thermoregulation. Chemosphere 2019, 237, 124408. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, A.; Alburaki, M.; Rinderer, N.; Sampson, B.; Knight, P.R.; Karim, S.; Adamczyk, J. Effects of Coumaphos and Imidacloprid on Honey Bee (Hymenoptera: Apidae) Lifespan and Antioxidant Gene Regulations in Laboratory Experiments. Sci. Rep. 2018, 8, 15003. [Google Scholar] [CrossRef]
- Tosi, S.; Nieh, J.C.; Sgolastra, F.; Cabbri, R.; Medrzycki, P. Neonicotinoid Pesticides and Nutritional Stress Synergistically Reduce Survival in Honey Bees. Proc. R. Soc. B 2017, 284, 20171711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjon, C.; Troczka, B.J.; Zaworra, M.; Beadle, K.; Randall, E.; Hertlein, G.; Kumar, S.S.; Nauen, R. Unravelling the Molecular Determinants of Bee Sensitivity to Neonicotinoid Insecticides. Curr. Biol. 2018, 28, 1137–1143. [Google Scholar] [CrossRef] [Green Version]
- Alptekin, S.; Bass, C.; Nicholls, C.; Paine, M.J.; Clark, S.J.; Field, L.; Moores, G.D. Induced Thiacloprid Insensitivity in Honeybees (Apis mellifera L.) Is Assoc. with Up-regulation of Detoxification Genes. Insect Mol. Biol. 2016, 25, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, M.M.; Robinson, G.E. Diet-Dependent Gene Expression in Honey Bees: Honey vs. Sucrose or High Fructose Corn Syrup. Sci. Rep. 2014, 4, 5726. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Substance Classification | Phenolic Substance Name | Amount (%) | Amount (mg/kg) |
|---|---|---|---|
| Phenolic acids | caffeic acid | 10 | 20 |
| benzoic acid | 20 | 40 | |
| gallic acid | 7.5 | 15 | |
| ferulic acid | 20 | 40 | |
| p-Coumaric acid | 35 | 70 | |
| vanillic acid | 7.5 | 15 | |
| Flavonoids | rutin | 25 | 2.5 |
| quercetin | 25 | 2.5 | |
| naringin | 25 | 2.5 | |
| hesperidin | 25 | 2.5 |
| Gene | Sequences 5′-3′ | Reference |
|---|---|---|
| Cyp9q1 | F: TCGAGAAGTTTTTCCACCG R: CTCTTTCCTCCTCGATTG | Mao et al. [37] |
| Cyp9q2 | F: GATTATCGCCTATTATTA R: GTTCTCCTTCCCTCTGAT | Mao et al. [37] |
| Cyp4g11 | F: AATGCGAGAAGTGTCGTCGA R: AGCGGTTTCCAGAAGGATGT | Calla et al. [38] |
| AmRp49 | F: CGTCATATGTTGCCAACTGGT R: TTGAGCACGTTCAACAATGG | Tesovnik et al. [39] |
| Treatment | Degrees of Freedom | Chi-Square Statistic | p-Value |
|---|---|---|---|
| TH/C | 1 | 310 | <0.001 |
| TH/F | 1 | 270 | <0.001 |
| FTH/TH | 1 | 72 | <0.001 |
| FTH/C | 1 | 62.9 | <0.001 |
| FTH/F | 1 | 51.2 | <0.001 |
| FTL/TL | 1 | 6 | 0.01 |
| TL/C | 1 | 5.7 | 0.01 |
| FTL/C | 1 | 4.6 | 0.03 |
| FTL/F | 1 | 1.8 | 0.17 |
| F/C | 1 | 0.6 | 0.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hýbl, M.; Mráz, P.; Šipoš, J.; Hoštičková, I.; Bohatá, A.; Čurn, V.; Kopec, T. Polyphenols as Food Supplement Improved Food Consumption and Longevity of Honey Bees (Apis mellifera) Intoxicated by Pesticide Thiacloprid. Insects 2021, 12, 572. https://doi.org/10.3390/insects12070572
Hýbl M, Mráz P, Šipoš J, Hoštičková I, Bohatá A, Čurn V, Kopec T. Polyphenols as Food Supplement Improved Food Consumption and Longevity of Honey Bees (Apis mellifera) Intoxicated by Pesticide Thiacloprid. Insects. 2021; 12(7):572. https://doi.org/10.3390/insects12070572
Chicago/Turabian StyleHýbl, Marian, Petr Mráz, Jan Šipoš, Irena Hoštičková, Andrea Bohatá, Vladislav Čurn, and Tomáš Kopec. 2021. "Polyphenols as Food Supplement Improved Food Consumption and Longevity of Honey Bees (Apis mellifera) Intoxicated by Pesticide Thiacloprid" Insects 12, no. 7: 572. https://doi.org/10.3390/insects12070572