
Impact of Nosema Disease and American Foulbrood on Gut Bacterial Communities of Honeybees Apis mellifera


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sterilized Honeybee Hive Diagnosis
2.3. Honeybee Preparation for P. larvae Infection
2.3.1. P. larvae Inoculum
2.3.2. Honeybees’ Food Preparation
2.3.3. P. larvae Infection in Honeybee
2.3.4. P. larvae Infection Analysis
2.4. Honeybee Preparation for Nosema infection
2.4.1. Honeybee Pathogen Preparation for N. ceranae Inoculum
2.4.2. N. ceranae Infection in Honeybee
2.5. Bacterial Community Analysis Using Next-Generation Sequencing
2.5.1. Communities Proportion Analysis
2.5.2. Alpha Diversity and Beta Diversity Analysis
2.5.3. Functional Genes Prediction
2.6. Statistical Analysis
3. Results
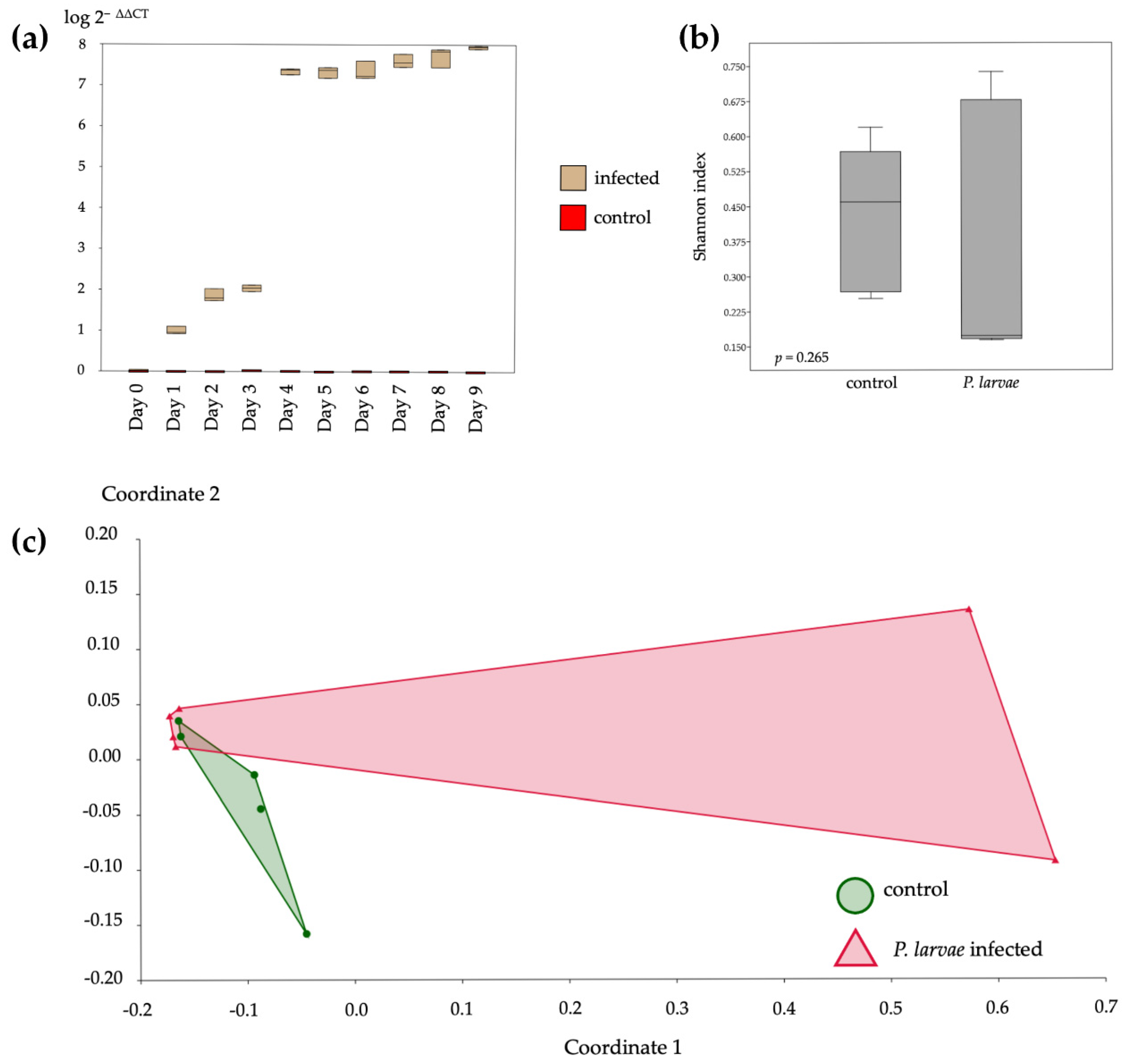
3.1. Level of P. larvae Infections and Their Bacterial Diversity Analysis
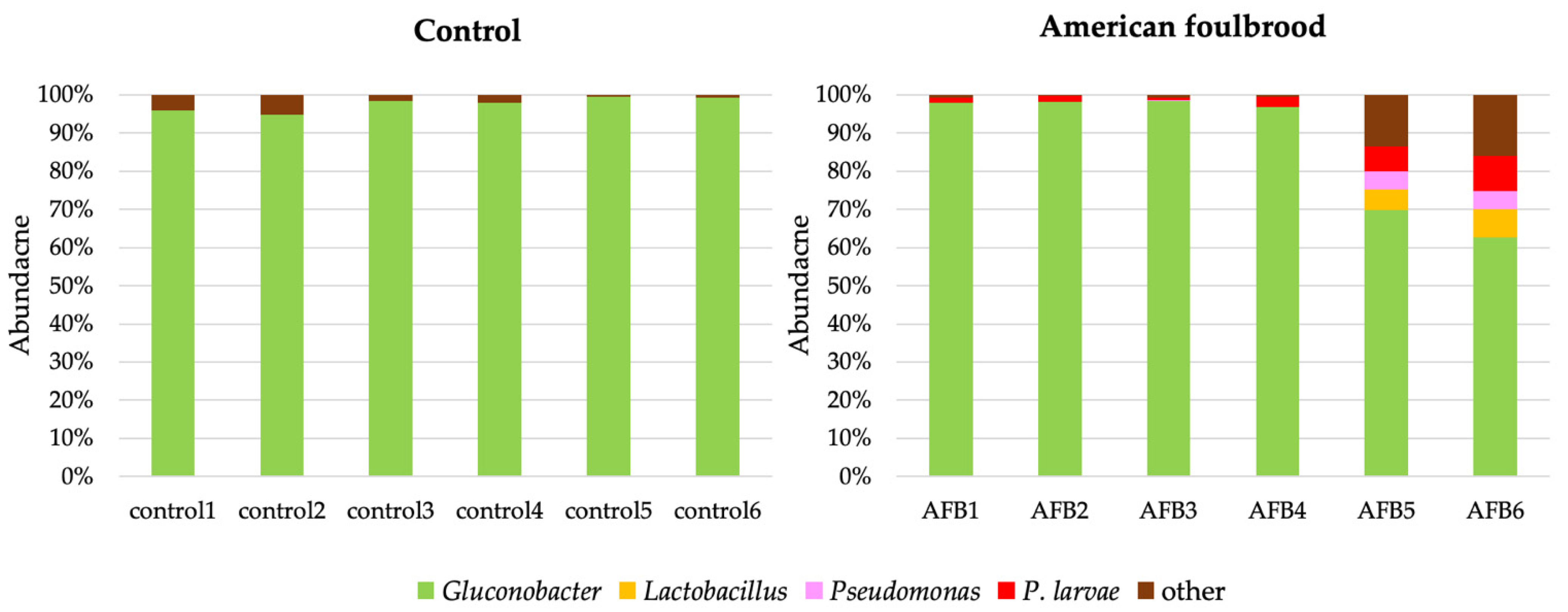
3.2. Bacterial Communities in the Gut of A. mellifera Infected by P. larvae
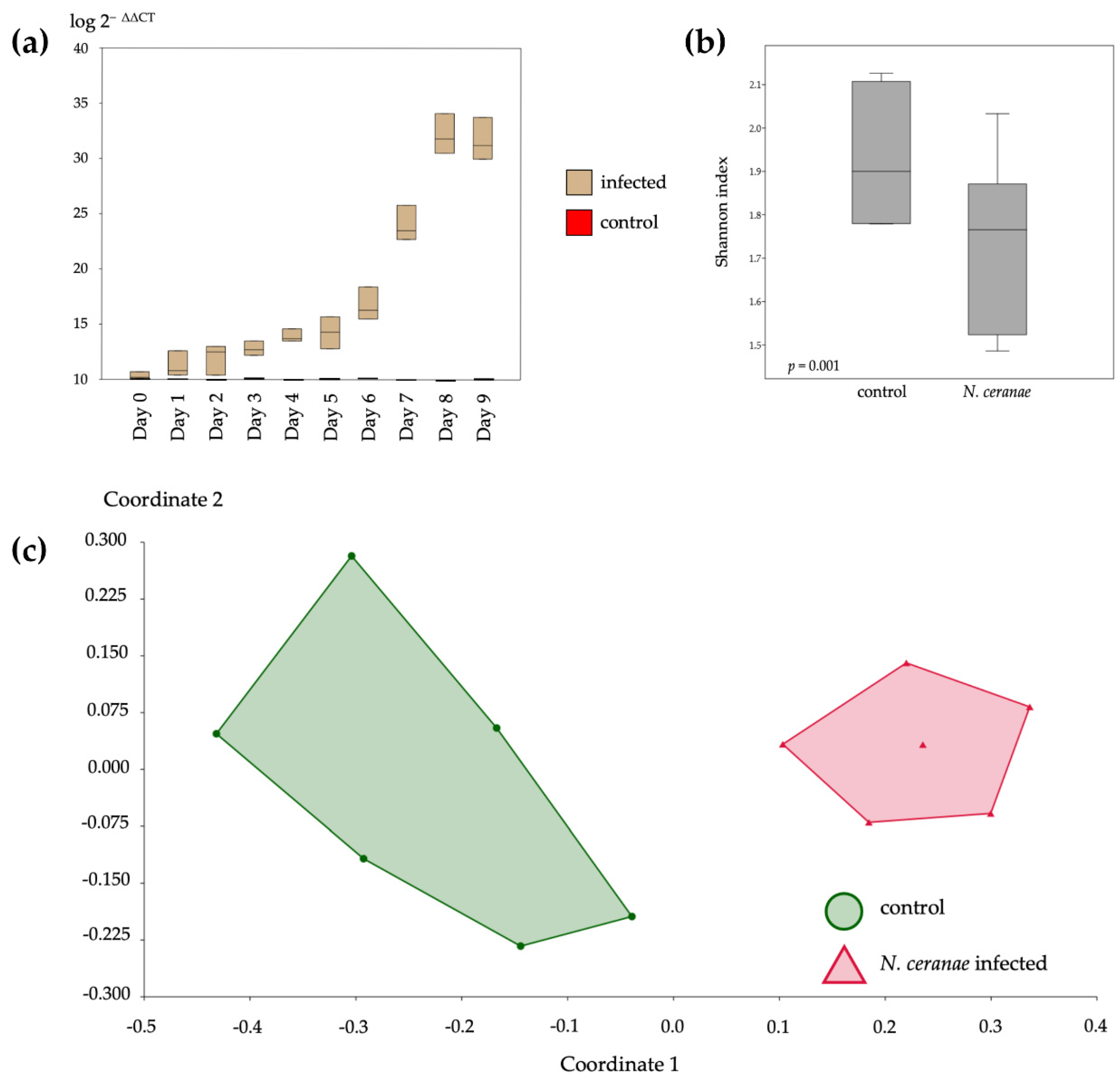
3.3. Level of Nosema Infections and Their Bacterial Diversity Analysis
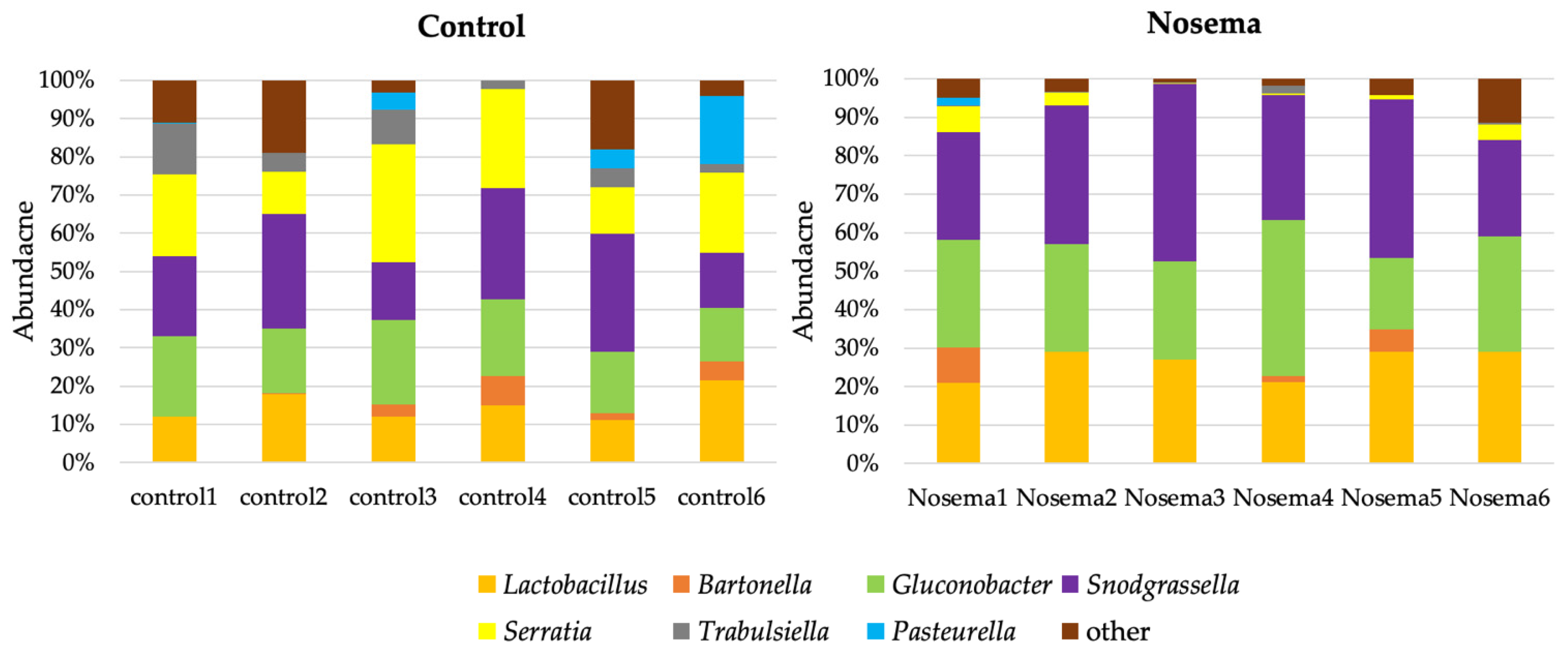
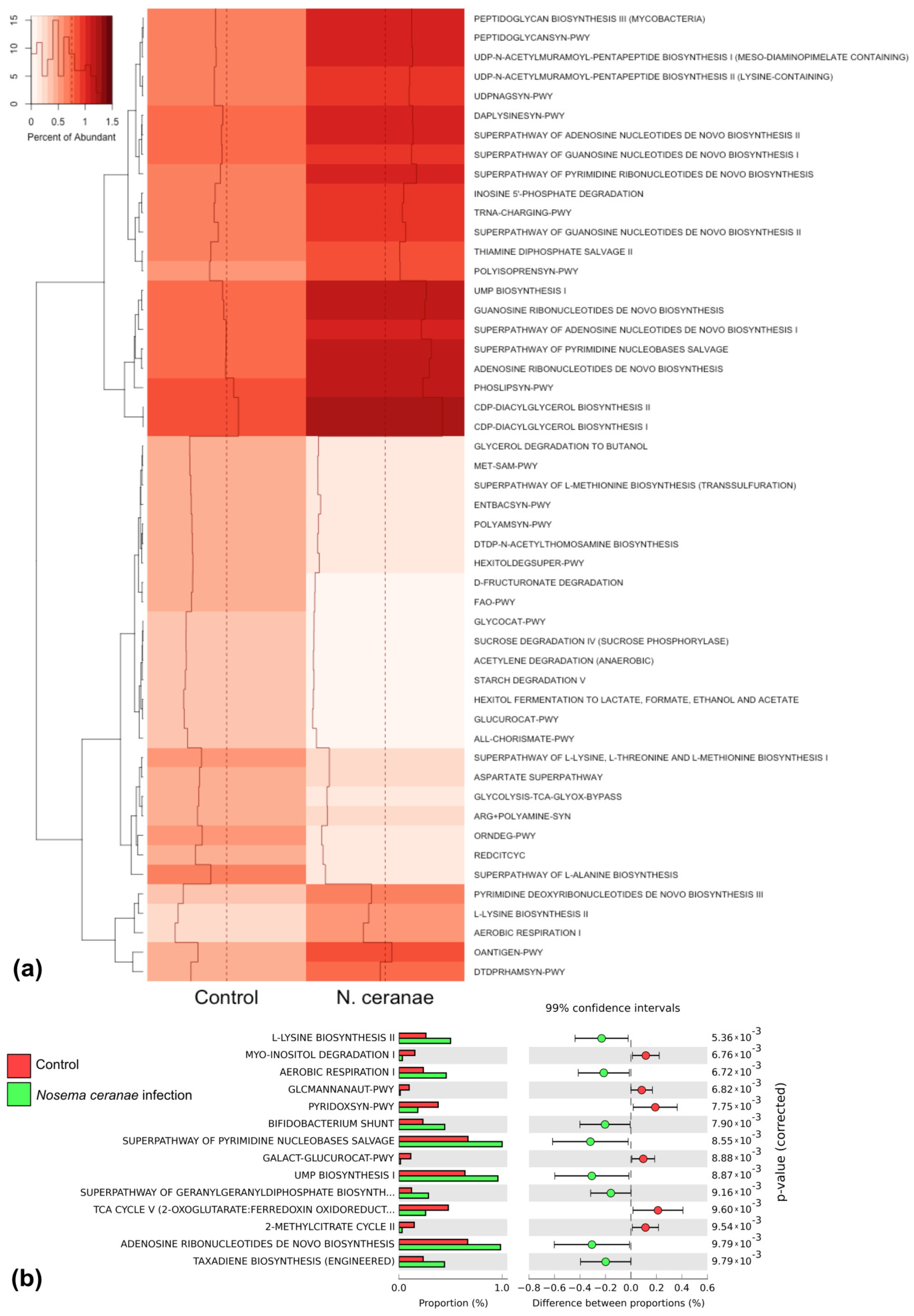
3.4. Bacterial Communities in the Gut of A. mellifera Infected by N. ceranae
3.5. Functional Gene Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hristov, P.; Shumkova, R.; Palova, N.; Neov, B. Factors Associated with Honey Bee Colony Losses: A Mini-Review. Vet. Sci. 2020, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Themudo, G.E.; Rey-Iglesia, A.; Tascón, L.R.; Jensen, A.B.; da Fonseca, R.R.; Campos, P.F. Declining genetic diversity of European honeybees along the twentieth century. Sci. Rep. 2020, 10, 10520. [Google Scholar] [CrossRef]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey bee gut microbiome is altered by in-hive pesticide exposures. Front. Microbiol. 2016, 7, 1255. [Google Scholar] [CrossRef] [PubMed]
- Fries, I.; Feng, F.; Da Silva, A.; Slemeda, S.B.; Pieniazek, N.J. Nosema ceranae sp. (Microspora, Nosematitidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 1996, 32, 356–365. [Google Scholar] [CrossRef]
- Genersch, E.; Forsgren, E.; Pentika, J.; Ashiralieva, A.; Rauch, E. Reclassification of Paenibacillus larvae subsp. pulvifaciens and Paenibacillus larvae subsp. larvae as Paenibacillus larvae without subspecies differentiation. Int. J. Syst. Evol. Microbiol. 2006, 56, 501–511. [Google Scholar]
- Ritter, W. Enfermedades de las Abejas; Acribia, S.A.: Zaragoza, Spain, 2001. [Google Scholar]
- Yue, D.; Nordhoff, M.; Wieler, L.H.; Genersch, E. Fluorescence in situ hybridization (FISH) analysis of the interactions between honeybee larvae and Paenibacillus larvae, the causative agent of American foulbrood of honeybees (Apis mellifera). Environ. Microbiol. 2008, 10, 1612–1620. [Google Scholar] [CrossRef]
- Matheson, A.; Reid, M. Strategies for the prevention and control of American foulbrood. Part. I. Am. Bee J. 1992, 132, 399–402. [Google Scholar]
- TAS (Thai Agricultural Standard). The standard title has been revised from “Thai Agricultural Commodity and Food Standard (TACFS)” to in accordance with the enforcement of The Agricultural Standards Act B.E. 2551 2007. R. Gaz. 2007, 125. [Google Scholar]
- Moran, N.A. Symbiosis as an adaptive process and source of phenotypic complexity. Proc. Natl. Acad. Sci. USA 2007, 104, 8627–8633. [Google Scholar] [CrossRef]
- Martinson, V.G.; Moy, J.; Moran, N.A. Establishment of characteristic gut bacteria during development of the honeybee worker. Appl. Environ. Microbiol. 2012, 78, 2830–2840. [Google Scholar] [CrossRef]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef]
- Zheng, H.; Nishida, A.; Kwong, W.K.; Koch, H.; Engel, P.; Steele, M.I.; Moran, N.A. Metabolism of toxic sugars by strains of the bee gut symbiont Gilliamella apicola. MBio 2016, 7, e01326-16. [Google Scholar] [CrossRef]
- Kwong, W.K.; Engel, P.; Koch, H.; Moran, N.A. Genomics and host specialization of honey bee and bumble bee gut symbionts. Proc. Natl. Acad. Sci. USA 2014, 111, 11509–11514. [Google Scholar] [CrossRef]
- Lee, F.J.; Rusch, D.B.; Stewart, F.J.; Mattila, H.R.; Newton, I.L.G. Saccharide breakdown and fermentation by the honey bee gut microbiome. Environ. Microbiol. 2015, 17, 796–815. [Google Scholar] [CrossRef]
- Ellegaard, K.M.; Tamarit, D.; Javelind, E.; Olofsson, T.C.; Andersson, S.G.; Vásquez, A. Extensive intraphylotype diversity in lactobacilli and bifidobacteria from the honeybee gut. BMC Genom. 2015, 16, 284. [Google Scholar] [CrossRef]
- Ellegaard, K.M.; Engel, P. Beyond 16S rRNA community profiling: Intra-species diversity in the gut microbiota. Front. Microbiol. 2016, 7, 1475. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Apibacter adventoris gen. nov., sp. nov., a member of the phylum Bacteroidetes isolated from honey bees. Int. J. Syst. Evol. Microbiol. 2016, 66, 1323. [Google Scholar] [CrossRef]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Disayathanoowat, T.; Young, J.P.W.; Helgason, T.; Chantawannakul, P. TRFLP analysis of bacterial communities in the midguts of Apis mellifera and Apis cerana honey bees in Thailand. FEMS Microbiol. Ecol. 2012, 79, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Cantwell, G.E. Standard methods for counting Nosema spores. Am. Bee J. 1970, 110, 222–223. [Google Scholar]
- Chen, Y.; Evans, J.D.; Smith, B.; Pettis, J. Nosema ceranae is a long present and widespread microsporidian infection of the European honey bee (Apis mellifera) in the United States. J. Invertebr. Pathol. 2008, 97, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Crailsheim, K.; Brodschneider, R.; Aupinel, P.; Behrens, D.; Genersch, E.; Vollmann, J.; Riessberger-Gallé, U. Standard methods for artificial rearing of Apis mellifera larvae. J. Apic. Res. 2013, 52, 1–16. [Google Scholar] [CrossRef]
- Evans, J.D.; Wheeler, D.E. Differential gene expression between developing queens and workers in the honey bee, Apis mellifera. Proc. Natl. Acad. Sci. USA 1999, 96, 5575–5580. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, J.D.; Shimanuki, H. Technique for rearing worker honeybees in the laboratory. J. Apic. Res. 1987, 26, 90–97. [Google Scholar] [CrossRef]
- Fourrier, J.; Deschamps, M.; Droin, L.; Alaux, C.; Fortini, D.; Beslay, D.; Le Conte, Y.; Devillers, J.; Aupinel, P.; Decourtye, A. Larval exposure to the juvenile hormone analog pyriproxyfen disrupts acceptance of and social behavior performance in adult honeybees. PLoS ONE 2015, 10, e0132985. [Google Scholar] [CrossRef] [PubMed]
- Alippi, A.M.; Aguilar, O.M. Characterization of Isolates of Paenibacillus larvae sub sp. Larvae from Diverse Geographical Origin by the Polymerase Chain Reaction and BOX Primers. J. Invertebr. Pathol. 1998, 72, 21–27. [Google Scholar] [CrossRef]
- Rembold, H.; Lackner, B.; Geistbeck, I. The chemical basis of honeybee, Apis mellifera, caste formation. Partial purification of queen bee determinator from royal jelly. J. Insect Physiol. 1974, 20, 307–314. [Google Scholar] [CrossRef]
- Fries, I.; Chauzat, M.P.; Chen, Y.P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standard methods for Nosema research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Fries, I. Comparative virulence of Nosema ceranae and Nosema apis in individual European honeybees. Vet. Parasitol. 2010, 170, 212–217. [Google Scholar]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron 2001, 4/1, 1–9. [Google Scholar]
- Caspi, R.; Billington, R.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Midford, P.E.; Ong, Q.; Ong, W.K.; et al. The MetaCyc database of metabolic pathways and enzymes. Nucleic Acids Res. 2018, 46, D633–D639. [Google Scholar] [CrossRef] [PubMed]
- Krongdang, S.; Evans, J.D.; Pettis, J.S.; Chantawannakul, P. Multilocus sequence typing, biochemical and antibiotic resistance characterizations reveal diversity of North American strains of the honey bee pathogen Paenibacillus larvae. PLoS ONE 2017, 12, e0176831. [Google Scholar] [CrossRef] [PubMed]
- Poppinga, L.; Genersch, E. Molecular pathogenesis of American Foulbrood: How Paenibacillus larvae kills honey bee larvae. Curr. Opin. Insect Sci. 2015, 10, 29–36. [Google Scholar] [CrossRef]
- Chan, Q.W.; Melathopoulos, A.P.; Pernal, S.F.; Foster, L.J. The innate immune and systemic response in honey bees to a bacterial pathogen, Paenibacillus larvae. BMC Genom. 2009, 10, 387. [Google Scholar] [CrossRef]
- Schwarz, R.S.; Moran, N.A.; Evans, J.D. Early gut colonizers shape parasite susceptibility and microbiota composition in honey bee workers. Proc. Natl. Acad. Sci. USA 2016, 113, 9345–9350. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Crauser, D.; Pioz, M.; Saulnier, C.; Le Conte, Y. Parasitic and immune modulation of flight activity in honey bees tracked with optical counters. J. Exp. Biol. 2014, 217, 3416–3424. [Google Scholar] [CrossRef]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Immune system stimulation by the native gut microbiota of honey bees. R. Soc. Open Sci. 2017, 4, 170003. [Google Scholar] [CrossRef]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef]
- Aufauvre, J.; Misme-Aucouturier, B.; Viguès, B.; Texier, C.; Delbac, F.; Blot, N. Transcriptome analyses of the honeybee response to Nosema ceranae and insecticides. PLoS ONE 2014, 9, e91686. [Google Scholar] [CrossRef] [PubMed]
- Raymann, K.; Moran, N.A. The role of the gut microbiome in health and disease of adult honey bee workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Harris, V.C.; Maes, P.; Anderson, K.E. The bacterial communities associated with honey bee (Apis mellifera) Foragers. PLoS ONE 2014, 9, e95056. [Google Scholar]
- Moran, N.A. Genomics of honey bee microbiome. Curr. Opin. Insect Sci. 2015, 10, 22–28. [Google Scholar] [CrossRef]
- Bucher, G.E. Nonsporulating bacterial pathogens. In Insect Pathology; Steinhaus, E.A., Ed.; Academic Press Inc.: New York, NY, USA, 1963; pp. 117–147. [Google Scholar]
- Krieg, A. Diseases caused by bacteria and other prokaryotes. In Epizootiology of Insect Diseases; Fuxa, J.R., Tanada, Y., Eds.; John Wiley & Sons Inc.: New York, NY, USA, 1987; pp. 323–355. [Google Scholar]
- Broderick, N.A.; Raffa, K.F.; Goodman, R.M.; Handelsman, J. Census of the bacterial community of gypsy moth larval midgut by using culturing and culture independent methods. Appl. Environ. Microbiol. 2004, 70, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Subta, P.; Shun, I.; Disayathanoowat, T.; Manaboon Poolkeaw, M.; Williams, G.R.; Susumu, K.; Chantawannakul, P. Bacterial communities in larval diapause and pupal gut in the bamboo borer, Omphisa fuscidentalis Hampson. Chiang Mai J. Sci. 2019, 46, 456–460. [Google Scholar]
- Steinhaus, E.A. Serratia marcescens Bizio as an insect pathogen. Hilgardia 1959, 28, 351–380. [Google Scholar] [CrossRef]
- Anand, A.A.P.; Vennison, S.J.; Sankar, S.G.; Prabhu, D.I.G.; Vasan, P.T.; Raghuraman, T.; Geoffrey, C.J.; Vendan, S.E. Isolation and characterization of bacteria from the gut of Bombyx mori that degrade cellulose, xylan, pectin and starch and their impact on digestion. J. Insect Sci. 2010, 10, 107. [Google Scholar] [CrossRef]
- Shil, R.K.; Mojumder, S.; Sadida, F.F.; Uddin, M.; Sikdar, D. Isolation and Identification of cellulolytic bacteria from the gut of three phytophagus insect species. Braz. Arch. Biol. Technol. 2014, 57, 927–932. [Google Scholar] [CrossRef]
- Yamada, Y.; Yukphan, P. Genera and species in acetic acid bacteria. Int. J. Food Microbiol. 2008, 125, 15–24. [Google Scholar] [CrossRef]
- Crotti, E.; Rizza, A.; Chouaia, B.; Ricci, I.; Favia, G.; Alma, A.; Sacchi, L.; Bourtzis, K.; Mandrioli, M.; Cherif, A.; et al. Acetic acid bacteria, newly emerging symbionts of insects. Appl. Environ. Microbiol. 2010, 76, 6963–6970. [Google Scholar] [CrossRef] [PubMed]
- Flury, P.; Aellen, N.; Ruffner, B.; Tarr, M.P.; Fataar, S.; Metla, Z.; Ferreras, A.D.; Bloemberg, G.; Frey, J.; Goesmann, A.; et al. Insect Pathogenicity in Plant Beneficial Pseudomonads: Phylogenetic Distribution and Comparative Genomics. ISME J. 2016, 10, 2527–2542. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, A.T.; White, P.J. Species of Bacillus that make a vegetative peptidoglycan containing lysine lack diaminopimelate epimerase but have diaminopimelate dehydrogenase. Microbiology 1985, 131, 2145–2152. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Sela, D.A.; Price, N.P.; Mills, D.A. Metabolism of bifidobacteria. In Bifidobacteria: Genomics and Molecular Aspects; Caister Academic Press: Peoia, IL, USA, 2010; pp. 45–70. [Google Scholar]
- Natalini, P.; Ruggieri, S.; Santarelli, I.; Vita, A.; Magni, G. Baker’s yeast UMP: Pyrophosphate phosphoribosyltransferase. Purification, enzymatic and kinetic properties. J. Biol. Chem. 1979, 254, 1558–1563. [Google Scholar] [CrossRef]
- Debast, S.B.; Buauer, M.P.; Kuijiper, E.J. Update the treatment guidance document for Clostridium difficile infection. Clin. Microbiol. Infect. 2014, 20, 1–26. [Google Scholar] [CrossRef]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision microbiome reconstitution restore bile acid mediate resistance to Clostridium difficile. Nature 2015, 517, 205–208. [Google Scholar] [CrossRef]
- Yongsawas, R.; Chaimanee, V.; Pettis, J.S.; Boncristiani, H.F., Jr.; Lopez, D.; In-On, A.; Chantawannakul, P.; Disayathanoowat, T. Impact of Sacbrood Virus on Larval Microbiome of Apis mellifera and Apis cerana. Insects 2020, 11, 439. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panjad, P.; Yongsawas, R.; Sinpoo, C.; Pakwan, C.; Subta, P.; Krongdang, S.; In-on, A.; Chomdej, S.; Chantawannakul, P.; Disayathanoowat, T. Impact of Nosema Disease and American Foulbrood on Gut Bacterial Communities of Honeybees Apis mellifera. Insects 2021, 12, 525. https://doi.org/10.3390/insects12060525
Panjad P, Yongsawas R, Sinpoo C, Pakwan C, Subta P, Krongdang S, In-on A, Chomdej S, Chantawannakul P, Disayathanoowat T. Impact of Nosema Disease and American Foulbrood on Gut Bacterial Communities of Honeybees Apis mellifera. Insects. 2021; 12(6):525. https://doi.org/10.3390/insects12060525
Chicago/Turabian StylePanjad, Poonnawat, Rujipas Yongsawas, Chainarong Sinpoo, Chonthicha Pakwan, Phakamas Subta, Sasiprapa Krongdang, Ammarin In-on, Siriwadee Chomdej, Panuwan Chantawannakul, and Terd Disayathanoowat. 2021. "Impact of Nosema Disease and American Foulbrood on Gut Bacterial Communities of Honeybees Apis mellifera" Insects 12, no. 6: 525. https://doi.org/10.3390/insects12060525