
Tephritid Fruit Fly Semiochemicals: Current Knowledge and Future Perspectives
by
 , , , and
, , , and
Francesca Scolari
1,* ,
,
Federica Valerio
2 ,
,
Giovanni Benelli
3,
Nikos T. Papadopoulos
4 and
and
Lucie Vaníčková
5,6,* 1
Institute of Molecular Genetics IGM-CNR “Luigi Luca Cavalli-Sforza”, I-27100 Pavia, Italy
2
Department of Biology and Biotechnology, University of Pavia, I-27100 Pavia, Italy
3
Department of Agriculture, Food and Environment, University of Pisa, Via del Borghetto 80, 56124 Pisa, Italy
4
Department of Agriculture Crop Production and Rural Environment, University of Thessaly, Fytokou st., N. Ionia, 38446 Volos, Greece
5
Department of Chemistry and Biochemistry, Faculty of AgriSciences Mendel University in Brno, Zemedelska 1, CZ-613 00 Brno, Czech Republic
6
Department of Forest Botany, Dendrology and Geobiocoenology, Faculty of Forestry and Wood Technology, Mendel University in Brno, Zemedelska 1, CZ-613 00 Brno, Czech Republic
*
Authors to whom correspondence should be addressed.
Insects 2021, 12(5), 408; https://doi.org/10.3390/insects12050408
Submission received: 27 March 2021
/
Revised: 23 April 2021
/
Accepted: 27 April 2021
/
Published: 30 April 2021
(This article belongs to the Special Issue From Insect Pheromones to Mating Disruption: Theory and Practice)
Abstract
:Simple Summary
Tephritid fruit flies comprise pests of high agricultural relevance and species that have emerged as global invaders. Chemical signals play key roles in multiple steps of a fruit fly’s life. The production and detection of chemical cues are critical in many behavioural interactions of tephritids, such as finding mating partners and hosts for oviposition. The characterisation of the molecules involved in these behaviours sheds light on understanding the biology and ecology of fruit flies and in addition provides a solid base for developing novel species-specific pest control tools by exploiting and/or interfering with chemical perception. Here we provide a comprehensive overview of the extensive literature on different types of chemical cues emitted by tephritids, with a focus on the most relevant fruit fly pest species. We describe the chemical identity, production modality and behavioural relevance of volatile pheromones, host-marking pheromones and cuticular hydrocarbons, as well as the technological advances available for their characterisation. The variegate set of approaches integrating the use of the identified chemical signals for the control of wild populations of key pests is also explored. Last but not least, key challenges for future basic to applied research regarding tephritids are outlined.
Abstract
The Dipteran family Tephritidae (true fruit flies) comprises more than 5000 species classified in 500 genera distributed worldwide. Tephritidae include devastating agricultural pests and highly invasive species whose spread is currently facilitated by globalization, international trade and human mobility. The ability to identify and exploit a wide range of host plants for oviposition, as well as effective and diversified reproductive strategies, are among the key features supporting tephritid biological success. Intraspecific communication involves the exchange of a complex set of sensory cues that are species- and sex-specific. Chemical signals, which are standing out in tephritid communication, comprise long-distance pheromones emitted by one or both sexes, cuticular hydrocarbons with limited volatility deposited on the surrounding substrate or on the insect body regulating medium- to short-distance communication, and host-marking compounds deposited on the fruit after oviposition. In this review, the current knowledge on tephritid chemical communication was analysed with a special emphasis on fruit fly pest species belonging to the Anastrepha, Bactrocera, Ceratitis, Rhagoletis and Zeugodacus genera. The multidisciplinary approaches adopted for characterising tephritid semiochemicals, and the real-world applications and challenges for Integrated Pest Management (IPM) and biological control strategies are critically discussed. Future perspectives for targeted research on fruit fly chemical communication are highlighted.
1. Introduction
Insect semiochemicals are compounds belonging to different chemical classes that regulate intra- and inter-specific communication, affecting major behavioural and physiological responses [1,2,3]. Based on the identity of the emitter and the receiver, semiochemicals can be classified as pheromones (i.e., molecules mediating communication between co-specifics) or allelochemicals (i.e., compounds involved in communication between individuals of different species). Allelochemicals include kairomones (beneficial to the receiver but producing disadvantages for the emitter), synomones (molecules that benefit both the emitter and the receiver), allomones (beneficial to the producer and with neutral effects to the receiver), and apneumones (chemicals of non-biological origin beneficial to the receiver) [4,5,6]. Semiochemicals mediate a number of behavioural processes, such as the identification of food sources, the location of mates and hosts for oviposition, and the avoidance of predators [7].
To achieve these different functions, insects use volatile, semi-volatile, and non-volatile chemicals that are involved in long-, medium-, and short-distance communication, respectively. These stimuli can be detected by sensory neurons of the olfactory system on the antennae and maxillary palps [8], such as in the case of volatile molecules, or by neurons of the gustatory system mainly on proboscis, ovipositor and legs, which are able to perceive non-volatile chemicals through contact chemoreception [9].
Among semiochemicals, pheromones (from the Greek words “φερειν”-transfer, and “ορμαν”-excite) can be classified in two main categories: (i) releaser pheromones that produce an immediate response in a recipient individual (e.g., a male fly orienting toward a female guided by sex pheromone), (ii) primer pheromones that trigger the initiation of a complex physiological response not immediately observable [10]. The complex functions pheromones exert are mediated by sex, aggregation, alarm, trails and host-marking compounds [11,12].
Since 1959, when the term ‘pheromone’ was proposed [13] and the first pheromone, bombykol, was chemically identified in the silkworm moth, Bombyx mori (L.) (Lepidoptera: Bombycidae) [14], an increasing number of studies have focused on unravelling the chemistry and biological roles of these substances in numerous species. In most cases, pheromones are blends of individual chemicals that can be shared among species, but that are mixed in species-specific combinations (i.e., quantitatively and qualitatively) [15,16]. So far, volatile pheromones have been described as being composed of two or three compounds in moths [17,18], one in Drosophila melanogaster Meigen (Diptera: Drosophilidae) ((Z)-undec-4-enal [19]), or by complex blends in honey bees (Hymenoptera: Apidae) [20]. The chemical diversity of pheromone blends is very high, including acetate esters, alcohols, aldehydes, carboxylic acids, hydrocarbons, epoxides, ketones, benzenoid compounds, isoprenoids, terpenoids, and triacylglycerides [21]. In the case of Lepidoptera, as well as other insects that mainly rely on long-distance sexual signalling, volatile pheromones are the primary semiochemicals adopted [22]. Other insects, such as Drosophila species, are characterised by complex courtship rituals and use cuticular hydrocarbons (CHs) of both high and low-volatility [23]. The body surface of many insect species is indeed covered by a thin film of wax, composed mainly of hydrocarbons. Complex mixtures of esters, alcohols and free fatty acids are components of the cuticular wax in some insects [24]. Beside their cuticle waterproofing function, the long-chain hydrocarbons of insects are involved in chemical communication, serving as sex pheromones, kairomones, species- and gender-recognition cues, nestmate recognition compounds, fertility and dominance cues, chemical mimicry, and primer pheromones. Such key roles boosted research efforts in the past several decades on many dipteran species, including fruit flies, house flies and mosquitoes [25,26]. Insect CHs are usually a mixture of compounds that may include n-alkanes, alkenes, terminally branched monomethylalkanes, internally branched monomethylalkanes, dimethylalkanes, trimethylalkanes and others. They are synthesised by an elongation-decarboxylation pathway in oenocytes, which are associated with epidermal cells or fat bodies. After synthesis, CHs are transported through haemolymph by lipophorin carrier [27,28].
An additional type of semiochemicals is used by several parasitic and phytophagous insects immediately after egg-laying, the host-marking pheromone (HMP). Its function is to affect the oviposition behaviour of conspecifics in a way that subsequent eggs are not deposited in their already utilised resource, thus reducing the time spent on the already exploited resource and the competition for limited host resources, with advantages for both the marker and the seeker [29]. The HMP can be synthetized by female fruit flies in the form of a complex molecule [30,31,32] or a simple compound [33,34,35]. The receptors located in the tarsi and mouthparts of females searching for an oviposition site allow the detection of HMPs [29,36,37].
In the last decades, progress has been made to determine the identity and composition of semiochemicals in insects, as well as the chemical specificity and functional properties of molecules mediating semiochemical perception such as odour and taste receptors, odorant and gustatory binding proteins. Insects developed extremely refined abilities to produce and discriminate among different arrays of chemicals. In this framework, an increasingly deeper knowledge of the mechanisms underlying semiochemicals’ production and stimuli coding is being acquired, also due to advancements in analytical approaches, which, in turn, is providing multiple novel/improved tools for insect pest control.
In the scenario depicted above, true fruit flies (Diptera: Tephritidae) are excellent models to investigate the differentiation of semiochemicals’ production and perception. Including more than 5000 species and 500 genera, this family is one of the largest among dipteran, with a worldwide distribution. Many tephritids are important pests of agricultural commodities infesting a wide range of fruits and vegetables. The most pestiferous species belong to genera Anastrepha Schiner, Bactrocera Macquart, Ceratitis Macleay, Rhagoletis Loew, and Zeugodacus Hendel [38,39] (Figure 1).
As an example, the damage caused annually in Africa by the Oriental fruit fly, Bactrocera dorsalis (Hendel), has been estimated to USD 2 billion mainly due to export trade bans [41]. Moreover, due to a number of biological features including multivoltinism, long adult longevity, high fecundity, remarkable response to various stresses, increased capacity to overwinter [42], several species became aggressive global invaders, imposing strict quarantine regulations in several fruit-producing countries [43,44]. Because of their high importance, there is a list of ongoing programs worldwide that aim to eradicate, contain or suppress the populations of tephritid species [45].
Area-Wide Integrated Pest Management (AW-IPM) has been proven successful for the control of tephritid pest species and incorporates different components, such as thorough population monitoring employing sophisticated trapping systems, the Sterile Insect Technique (SIT), the Male Annihilation Technique (MAT) and often bait insecticidal sprayings [46]. To ensure effective application of the above components of the AW-IPM programs, an in-depth understanding of insect communication and mating strategies is required. This field has been widely investigated in tephritids and an expanding body of literature is available.
In this review, we critically discuss the current knowledge on pheromone-based communication in tephritid fruit flies, as well as its applied relevance for pest control. Starting from the role of these semiochemicals in tephritid reproductive behaviour, we analysed the most relevant, available literature focusing on (i) volatile pheromones released by males and/or females, (ii) HMPs, and (iii) CHs. For each of these three groups, the tissues involved in the production, their chemical identity, and the analytical methods applied for the identification, as well as the electrophysiological and behavioural tools employed to shed light on their ecological significance, are considered. Due to their high economic importance, insights are provided on pest species of the genera Anastrepha, Bactrocera, Ceratitis, Rhagoletis and Zeugodacus. Over the last decades, tephritid semiochemical research has not been equally dedicated to all three groups of semiochemicals. Thus, information related to volatile pheromones, mostly because of their more direct applied implications, is far more extended. This is also reflected in the present review, which additionally focuses on the role of semiochemical-based communication in species evolution. Tephritids display different levels of host specialisation, covering the whole spectrum from monophagy to polyphagy. Whether and how the adaptation to novel host plants drove the evolution of such an extreme diversity is still an open question and requires investigation of multiple factors (and their interactions), including those related to semiochemical-based communication. Finally, results focusing on applying the above-mentioned knowledge to fruit fly population monitoring and management, as well as to chemical taxonomy, are analysed, towards formulating major challenges for future research.
2. Semiochemicals and Reproductive Behaviour—An Overview
Semiochemicals are involved in the different phases of the reproductive behaviour of many tephritids, which include male lek formation and sexual signalling (sexual calling), courtship, acceptance of a mating partner and successful copulation (Figure 2).
The release and perception of semiochemicals are often accompanied by a range of intense behavioural interactions, including wing vibration and buzzing and head rocking [48,49,50,51].
There is a variety of mating systems in tephritid fruit flies, with lek-based ones being common in many species of economic importance [52]. Leks are mating arenas where males aggregate and perform sexual signalling without controlling the access to resources that may be critical for females or offering parental care, and provide only the sperm to females that freely choose their mates [53]. Males perform sexual signalling mainly on the under surface of leaves of preferred host trees that either bear fruits as in Anastrepha fraterculus (Wiedemann) and B. dorsalis, or do not bear fruits as in certain populations of the Mediterranean fruit fly (medfly) Ceratitis capitata (Wiedemann) (see [54] for a review). Instead, in Zeugodacus cucurbitae (Coquillett), lekking takes place more frequently on non-host plants [55,56]. Likewise, in Bactrocera cacuminata (Hering), the wild tobacco fruit fly, non-host plants containing the male attractant methyl eugenol (ME) serve as the main mating sites [57]. Lekking males, in groups of 2–10 individuals [54], perform sexual signalling that includes a set of visual, acoustic and olfactory signals, with volatile pheromone acting as the longer-distance cue.
On the other hand, at variance with many tephritids, most Rhagoletis spp. do not exhibit male lekking [52]. For example, males of Rhagoletis pomonella (Walsh), Rhagoletis cerasi (L.), Rhagoletis ribicola Doane and Rhagoletis mendax Curran individually search for potential mates mainly on host fruits and the top surface of leaves [58], mostly during the late afternoon and at dusk [52,59,60]. However, in Rhagoletis batava Hering individuals have been reported to form small groups in response to male-released pheromone that might be considered leks [61]. While olfactory cues seem not to play a role in lek location for Anastrepha obliqua (Macquart) and Anastrepha ludens (Loew), data are rather controversial for C. capitata, B. dorsalis, and Z. cucurbitae [54]. Whether the cues exploited by males to aggregate in leks derives from their own pheromone emissions, or from the plants where leks occurs is still under debate. Indeed, in C. capitata it has been proposed that plant volatiles, rather than male-emitted pheromones, contribute to male aggregation in leks [62,63], with α-copaene, a natural sesquiterpene widely present in plants, being regarded as the primary male cue for lek formation [64,65,66,67]. Male pheromone has been shown to attract conspecific males thus acting as aggregation pheromone in some Bactrocera species [68,69,70,71,72,73]. Nonetheless, Kobayashi and colleagues reported no male-male attraction in B. dorsalis [74].
Interestingly, sex pheromones are not uniquely produced by males in all tephritid species. In the Ceratitis, Anastrepha and Rhagoletis genera, the volatile chemicals identified in the adult headspace are of male origin. Instead, the role of females in emitting volatiles is evident in the genus Bactrocera. In all Bactrocera species studied so far, both sexes produce and release pheromone chemicals, albeit interspecific differences exist. In Bactrocera musae (Tryon), a polyphagous pest distributed in Australia and Papua New Guinea mainland [75], both sexes produce volatile pheromones, but its complexity is higher in females than males [76].
In the olive fruit fly, Bactrocera oleae (Rossi), females release the pheromone that plays a central role in the mating system of this species [77,78,79,80]. However, earlier and most recent studies suggested that male-emitted chemicals elicit female attraction [81,82,83]. The finding that both sexes emit and perceive chemicals during courtship [50,84], together with the identification of male wing vibration in this species [50], encourage further research to better understand the role of chemical communication in the mating behaviour of B. oleae. So far, males have been regarded to swarm before settling on olive canopy [42]. The modality of male aggregation to form leks in this species needs to be further clarified too, in order to understand the relevance of chemical communication in a species where female-borne pheromones have been well documented both as a short- and long-distance cue [42]. Recently it was proven that ethyl decanoate emitted by olive fruit fly females attracts other females, and hence may be involved in female-female aggressive interactions on oviposition sites [85].
Courtship implies a series of ritualized actions, and it is much more complex in the genus Ceratitis, particularly in C. capitata [86], than in other tephritids, such as many species of Anastrepha [87,88] and Bactrocera [89]. However, recent studies are revealing that, even in species where it was believed to be simplified, such as in Bactrocera tryoni (Froggatt), courtship is more complex than previously thought [49]. During courtship, volatile chemicals are perceived by either one of both sexes using the antennae, while less volatile compounds, such as CHs, have been suggested to be important in later stages, when male and female touch each other during mating attempts and mating per se [35,42,90]. Indeed, in several tephritids, fore- and hind-leg interactions, as well as male foreleg interaction with the female abdomen, ‘kissing’ (i.e., touching of the labella), and male tapping with the labellum on female thorax have been reported [42,49,91]. For example, in Ceratitis rosa Karsch and A. fraterculus, sex-specific differences in the quantitative composition of CH profile, together with reported mating incompatibilities, further suggest the role of CHs as short-range semiochemicals [90,92,93].
Trophallaxis (i.e., female provision with gifts -oral, genital or transdermal- by males [91,94,95]), a common behaviour in tephritids, is considered as a courtship signal as well (see [96] for a review). While in Anastrepha species a pre-mating transfer of male oral products to females by labrum-to-labrum contact is well known [91,97,98], this phenomenon has been described in medfly only recently [96]. In this species, the consumption of male-produced oral droplets by the female appeared to increase her receptivity to mating, suggesting that the chemical composition of these droplets may be a tool to assess male quality [35,96]. Further research in the medfly and in other major tephritid fruit fly species is thus essential to unravel how these substances are perceived and how this behaviour fits in the complex semiochemical-based communication frame. Interestingly, a recent work demonstrates that A. ludens males regurgitate more than females, and propose that regurgitation and deposition of series of droplets organized in lines or spirals by some fruit fly species may play multiple functions (e.g., collecting bacteria from the environment, or eliminating ingested toxicants), including the production of oral pheromones [99]. Moreover, signalling males in some Anastrepha spp. deposit pheromone on the leaf surface by abdominal tip dipping [100]. Some of these deposited components persist on leaves up to one hour after removal of signalling males and are able to attract females [100]. Abdominal dipping with deposition of a viscous substance from the male cercus (i.e., the external appendage close to the digestive tract) has also been observed in Rhagoletis boycei Cresson [101].
After mating, tephritid females undergo an almost immediate switch from response to male pheromone to host plant-oriented olfactory behaviour to seek for appropriate oviposition background. This phenomenon has been well documented for the medfly at the behavioural level [102] and molecular data suggest that genes related to olfaction and/or foraging are also changing in their transcriptional profiles [103]. Mating-related differential expression in genes involved in chemosensory perception has also been detected in Bactrocera [104,105], and Anastrepha spp. [106], with females of B. tryoni showing mating-induced switches in olfactory preference [107].
The oviposition behaviour of tephritids is highly heterogeneous and display species-specific differences, such as daily patterns of oviposition, clutch size, patterns of positioning the ovipositor, duration of the oviposition bout, and preferred plant site for the oviposition [108]. In the medfly, oviposition behaviour has been widely studied and described as being structured in four steps: arrival to the fruit, exploring (i.e., survey of fruit surface with head, labellum and ovipositor), ovipositor puncturing and drawing following oviposition conclusion [109,110]). In the last phase, the fruit surface is again explored by the females, with the aculeus of the ovipositor protracted, and a HMP is deposited [111].
The presence of conspecific eggs and developing larvae in breeding substrates may dramatically alter the oviposition behaviour in phytophagous insects. In tephritids, females often mark already used hosts with a pheromone to avoid overexploitation of the specific resource and hence reduce/eliminate competition [112,113]. To maximise the chances of survival and success of their progeny, phytophagous insects tend to avoid egg laying in already explored host resources [29,108]. Host marking is particularly important in endophytic species, such as tephritids, in which females oviposit inside fruits or other plant tissues, with no visible damage and no emission of specific plant volatiles in response to infestation and presence of fruit fly eggs [114]. HMPs are generally applied by female drawing the ovipositor following an egg-laying event [112].
Mechano-, hygro- and gustatory receptors are located on female ovipositor in tephritids [115]. Gustatory sensilla present on the tarsi of R. pomonella exhibit sensitivity to HMP [116]. Sensilla types on female ovipositor have been described in several Bactrocera and Zeugodacus species (i.e., Z. cucurbitae, Zeugodacus diaphorus (Hendel), B. dorsalis, Bactrocera minax (Enderlein), Zeugodacus scutellatus (Hendel) and Zeugodacus tau (Walker) [117]; B. tryoni [118]), as well as in R. pomonella [119,120,121], but their characterisation is still patchy in species of the Ceratitis and Anastrepha genera [122].
Interestingly, HMPs have been shown to either deter or enhance oviposition, depending on concentration and other factors [123]. Deterrent effects induce different behaviours in responding females including suppression/disruption of oviposition, reduction of the number of egg clutches per fruit and of egg number per clutch, and dispersion to less infested (occupied) areas [108,114,124,125,126,127,128]. Although HMPs are predominantly recognised by individuals of the same species [29,129], interspecific perception of HMPs has also been described in tephritids. Cross-recognition has been demonstrated among species of the Rhagoletis [130,131], Anastrepha [132] and Ceratitis genera [33,34]. The host-marking behaviour may display different features even among species of the same genus. For example, small-sized fruit specialists (e.g., Rhagoletis alternata (Fallen), Rhagoletis indifferens Curran, R. pomonella and R. cerasi) often deposit HMPs [133]. Conversely, members of the Rhagoletis suavis group rarely mark the host targets [134,135] and commonly tend to lay eggs on infested fruits [134,136,137]. Two hypotheses have been proposed for the sporadic host-marking behaviour of the suavis group. According to the “no HMP deposition” hypothesis, all species of this group use as host walnut species (Juglans spp.), which are not infested by other flies of the genus in North America, and hence provide a competition-free resource for larvae [133]. The second hypothesis proposes the occurrence of a “male host-marking behaviour”. Rhagoletis boycei males indeed usually touch the host fruit depositing a substance on its surface and females preferentially oviposit on the unmarked fruit [101]. According to this hypothesis, male host-mark replaces the female’s one, causing a reduction in female marking behaviour. Male host-marking behaviour has been described in two species of the suavis group, R. boycei and R. suavis (Loew) [113].
The drawing of the aculeus after oviposition without an evident release of HMP has been described in B. dorsalis [138], B. tryoni and B. jarvisi (Tryon) [139], and Z. cucurbitae [140]. Instead of using their ovipositor right after oviposition, olive fly females spread the olive juice that leaks from the oviposition wound over the fruit surface using their labella. This behaviour appears to prevent other females from ovipositing on the same fruit [141,142,143].
3. Volatile Pheromones
3.1. Tissues Involved in Volatile Pheromone Production
Male pheromones seem to be released primarily from glands positioned in the rectum in Ceratitis [144,145], Bactrocera [74,79,146,147,148], and Anastrepha spp. [149,150,151]. However, interesting variations have been reported. This is the case of the goldenrod gall tephritid fly Eurosta solidaginis (Fitch), which possess no anal glands but enlarged rectum and pleural epidermis that have been suggested to be involved in pheromone production and/or storage [152].
In general, the rectal valve of A. fraterculus and C. capitata is located at the distal portion of the colon, while the intestinal canal enlarges in a chamber comprising four rectal papillae projecting in the lumen of the rectal ampulla (i.e., the anterior rectum) [153]. In its proximal region, the ampulla is lined with epithelial cells that increase in number and are organized in folds in the distal portion. In several Anastrepha species, the structure of the female rectum is similar, with differences in the features of the epithelium [154]. In Bactrocera spp. (e.g., B. oleae, B. dorsalis and B. tryoni), the rectal glands of males and females display a remarkable sexual dimorphism [155,156,157]: muscles surrounding male glands are more abundant, suggesting a more complex contraction capacity to support the pheromone-storage and -release functions. The rectal sac (i.e., an evagination in the rectal gland) is present only in males. Because they are not surrounded by muscles and directly exposed to the haemolymph, the rectal pads, which extend into the gland as rectal papillae, have been suggested to be involved in transporting chemicals into the rectal gland [157].
During sexual calling, the anal tube is protruded to allow the epithelium of the distal rectum to be extruded. In medfly, when everted, the folded rectal epithelium is expanded in a balloon-like structure [144]. This evagination of the anal membranes at the abdominal tip, which appears as a droplet, has been described in Anastrepha and Ceratitis species, and it has been explained as a mean of expanding the evaporative surface, thus increasing pheromone emission and, consequently, attractiveness [100,150,158]. In addition, this behaviour is accompanied by protrusion of male expansion of the pleural abdominal region generating two lateral blisters [149,159].
Males of C. capitata and C. rosa display three types of sex-specific glands, (i) the anal glands, (ii) the pleural glands and (iii) the dimorphic salivary glands [160]. The pair of anal glands open onto the external cuticle close to the anal opening [160,161]. Males of Anastrepha species display only the pleural glands and dimorphic salivary glands [151,160]. In Bactrocera species (e.g., B. oleae, B. tryoni and B. dorsalis), both the anal glands and the dimorphic salivary glands are absent [146,160,162]. Currently, detailed information on Rhagoletis species are lacking, although the early work of Nation [160] found neither dimorphic salivary glands nor pleural glands in both R. pomonella and R. juglandis (Cresson).
In addition to the key function played by rectal tissues, pheromones have been later found to be also released orally. Salivary glands release pheromone components in Anastrepha spp. [149,150,151,163,164]. These glands are sexually dimorphic, with male salivary glands being ball-like structures associated with the crop [99,160,165]. Salivary glands have been suggested to be involved in the storage and, potentially, synthesis of pheromone components in medfly males [166]. This is supported by the fact that some chemicals of the pheromone blend identified in medfly male headspace were also detected in the extracts of salivary glands. Figure 3 summarizes the tissues involved in pheromone production in tephritids.
3.2. Composition of Volatile Pheromones
In tephritids, the volatile pheromone is a mixture of diverse chemical compounds with different isomers [167]. These chemicals are either newly synthesised or produced from precursors acquired from the diet. Plant-borne chemicals introduced with the diet become integrated into the pheromone mixture and contribute to male mating success. Diet-derived chemicals can unmodified be incorporated into the pheromone blend (e.g., raspberry ketone (RK) in Z. cucurbitae [168,169]) or can go through conversion in other compounds that are then used in the pheromone (e.g., ME in B. dorsalis that is converted in 2-allyl-4,5-dimethoxyphenol and trans-coniferyl alcohol [68,69,170,171]). Bactrocera dorsalis males are attracted to ME, and feeding on this compound was shown to directly benefit mating success [171,172].
The pheromone mixture comprises both major, minor and trace compounds, with the complete blend displaying stronger effects than individual compounds or a mix of a subset of compounds. Indeed, medfly females respond differentially to mixtures of major male pheromone compounds than to the complete blend [173]. In B. oleae, olean (1,7-dioxaspiro[5.5]undecane) is the major component of female sex pheromone, and it is more attractive to males than the other identified components (e.g., α-pinene, nonanal, and ethyl dodecanoate) [174]. However, the combination of all chemicals is more attractive than olean alone (see [171] for a review).
The composition of the pheromone mixture has been investigated using two main approaches: (i) sampling the headspace of calling males (or females in the case of Bactrocera species), (ii) extracting the chemicals from the rectal glands. Volatiles captured in the headspace have been so far collected and characterised in 18 tephritid species (Table 1; Table S1), while gland extracts have been obtained from 26 species (Table 2).
The two most represented chemical classes of the volatile compounds captured in the headspace in Anastrepha, Bactrocera, Ceratitis, Rhagoletis and Zeugodacus species are fatty acyls and organooxygen compounds. Prenol lipids, which represent most compounds in Ceratitis and Anastrepha species, and are abundant also in Rhagoletis, are instead poorly represented in Bactrocera and absent in the emissions of Z. cucurbitae. Lactones are particularly abundant in Anastrepha species (Table S1).
Within each genus, it is evident that only few compounds are shared among species (Figure 4). For example, in all four Ceratitis species investigated so far, namely C. capitata, Ceratitis fasciventris (Bezzi), Ceratitis anonae (Graham), and C. rosa, only three volatile chemicals, all belonging to the prenol lipid class, are emitted by males. These three compounds are linalool, (E)-β-ocimene, and (Z)-β-ocimene that also occur naturally in the host plants. In C. capitata, the perception of plant volatiles has been investigated by electroantennogram (EAG) and behavioural studies demonstrating that linalool, a compound representative of immature citrus fruit associated with high toxicity against immature stages of fruit flies and considered as an important compound conferring resistance against fruit fly larval development, has a significant deterrent effect [240]. Linalool was reported by numerous studies as an active constituent of medfly male sex pheromone that elicits a strong EAG response [218,221,241]. It also triggers antennal depolarisation in females of C. fasciventris, C. anonae and C. rosa (so-called Ceratitis FAR complex) [214].
In Anastrepha, (Z)-non-3-en-1-ol (a member of the fatty acyl compound class) is the unique compound shared among A. fraterculus, A. ludens, A. obliqua, and A. suspensa. This compound is a typical host plant component and has been shown to elicit an active behavioural response in all four species [175,188,193,242,243]. For example, A. fraterculus females are attracted to (Z)-non-3-en-1-ol [175]. These behaviours may be interpreted as a first step in the complex mating process of this species, i.e., attracting females to the mating site. Since mating is strongly associated with host plants, the use of plant typical compounds (e.g., limonene and pinene, among others) would help females to find simultaneously mating and oviposition sites [244].
N-(3-methylbutyl)acetamide (carboxylic acids and derivatives class) and the three organooxygen compounds (E,E)-8-ethyl-2-methyl-1,7-dioxaspiro[5.5]undecane, (E,E)-8-methyl-2-propyl-l,7-dioxaspiro[5.5]undecane, and (E,E)-2,8-dimethyl-l,7-dioxaspiro[5.5]undecane are shared among B. dorsalis s.s., B. oleae, B. zonata (Saunders) and B. tryoni. Interestingly, N-(3-methylbutyl)acetamide elicits female attraction in B. dorsalis and B. carambolae, but also in Z. cucurbitae [72,204,229]. The spiroacetal (E,E)-2,8-dimethyl-1,7-dioxaspiro[5.5]undecane elicits an antennal response in B. musae males, suggesting a biological role for this compound [76].
It is noteworthy that research efforts in investigating pheromone composition have been particularly intense in certain species such as the medfly, and this may reflect in the number of identified compounds. Future studies integrating multiple analytical approaches in all species will enable a more extensive description of the shared and unique chemical signatures of tephritid volatile pheromones.
The differences in the male pheromone composition that have been reported may be partially linked to technical aspects (e.g., different sampling techniques, experimental variables such as airflow, sampling duration, and column features), as well as to several factors affecting pheromone production and release, such as the time of the day [149,200], social context [149], food availability [199], and diet [220,245]. Larval diet (i.e., the different host fruits utilised) and adult age affect both the total amount of pheromone emitted by males, the relative quantity of major components, and the presence of specific minor compounds in the medfly [220]. Similarly, in A. ludens, the profile of the male pheromone blend and the relative abundance of the chemical components change in response to different host fruits used for larval development [245]. Moreover, qualitative and quantitative differences in the composition of male-emitted volatile pheromone have been detected among laboratory strains and different populations in A. fraterculus [176,177], and among laboratory strains and wild populations in C. capitata [221]. In addition, emission of male pheromone increases in Anastrepha species in response to treatments with methoprene (a synthetic analogue of the juvenile hormone), with important implications in gaining deeper insights into tephritid reproductive physiology and in enhancing the application of management methods such as the release of sterilised males [246,247].
The components of male pheromone also display different volatilities [248]. Six alkanes and related compounds have been identified in the headspace of medfly males reared on a standard wheat bran-based larval diet [220]. These compounds display poor volatility and do not belong to the published CHs identified in the cuticle of adult medflies. Thus, similar to Drosophila species [249,250], medfly seems to be able to deposit on the substrate and emit in the surrounding air CH-like compounds, which may serve as short/medium distance cues for mate localisation. The ability of males to deposit pheromone on the substrate, in addition to aerial pheromone release, has also been reported in A. suspensa [63,100]. Moreover, saturated C25, C27 and C29 hydrocarbons have been detected in the volatile pheromone of male melon fly Z. cucurbitae [205].
3.3. Does Host-Preference Affect the Volatile Pheromone Bouquet?
Pheromone precursors are acquired from four main sources: (i) de novo synthesis, (ii) conversion of precursors that insects acquire from host plants or substrate, (iii) direct incorporation from the host plants, and/or (iv) from endosymbionts [21,251,252,253,254]. Thus, it is likely that different variables influence the pheromone blend.
The chemical classes of their pheromone components and their specific identity are more similar among species of Anastrepha, Ceratitis and Rhagoletis genera, and rather different from species of the genus Bactrocera. The monophagous B. oleae is the most diverse species among the genus, with the majority of its pheromone volatiles being species-specific. To elucidate whether the composition of the pheromone components is related to feeding strategies of different species, we analysed the lists of volatile compounds identified in all species of the genera Anastrepha, Ceratitis, Bactrocera and Zeugodacus. This analysis could not be extended to Rhagoletis genus, as only the volatiles emitted by R. cerasi have been identified [226] so far, and a single compound has been isolated in R. batava [61].
The species with a monophagous or oligophagous feeding strategy mostly emit chemicals that are not shared with other species with the only exception of nonanal, α-pinene and p-cymene, which are in common between B. oleae and R. cerasi. Figure 5 shows the number of chemicals identified in each species as well as the intersections of overlapping compounds, represented by connected dots (see Table S2 for the complete lists of unique and overlapping chemicals).
The polyphagous species shared more compounds, as shown by the higher number of connected dots in Figure 6 (see Table S2 for the complete lists of unique and overlapping chemicals). This finding is particularly evident between species belonging to the same genus.
Figure 6 also reports the identity of the chemicals that are shared between different genera. It generally appears that the members of the Anastrepha and Ceratitis genera share a higher number of compounds, with chemicals belonging to the prenol lipids class being most frequently in common. Conversely, Bactrocera species display quite unique pheromone blend features. The only two compounds to be shared belong to the diazine class and are in common with Ceratitis species. This suggests that at least two major evolutionary forces, i.e., genomic background and host feeding strategy, interacting in a complex manner, have shaped the pheromone blend in these species. A comprehensive molecular phylogeny of tephritid species will be of great help in disentangling the effects of genome evolution and environmental selective pressures in shaping semiochemical-based behaviour in these species. Further research is required to deeply investigate how feeding strategies affect the production of semiochemicals, also with respect to the capacity of colonizing new hosts, a feature that characterises several invasive tephritid pests. A strong effect of larval host on the pheromone bouquet produced by the polyphagous species A. ludens and A. obliqua has been recently demonstrated [245].
3.4. Analytical Approaches to Unravel Pheromone Composition
The complexity of the signals involved in tephritid chemical communication, together with their presence in small amounts in natural systems, stimulated the adoption of very sensitive high-resolution analytical methods for their detection, identification and testing. These strategies include both conventional as well as reverse chemical ecology approaches. The workflow of conventional chemical ecology (CCE) approaches generally involves the following steps: (i) preliminary observation of the existence of pheromone-mediated communication in the target species, (ii) volatile pheromone sampling, (iii) characterisation of pheromone blends through analytical techniques, (iv) chemical synthesis of the identified compounds, (v) evaluation of their electrophysiological activity, (vi) behavioural assays (bioassays) to test the role of the isolated semiochemicals in laboratory and/or field set-ups in order to confirm their pheromone identity [256]. Recently, a different approach began to be utilised for the discovery of semiochemicals, reverse chemical ecology (RCE) [257]. Similarly to what occurs for receptor-based drug discovery, RCE exploits odorant binding proteins (OBPs) as molecular targets for the screening of behaviourally-active chemicals based on their binding affinity [258]. The workflow of RCE comprises the following steps: (i) identification of OBP targets through genomics and bioinformatics approaches, (ii) characterisation of OBP expression profile, (iii) purification and 3D-structural analysis, (iv) in vitro OBP: ligand binding assays using semiochemicals collected from insects or pure compounds, (v) in silico selection of test ligands, (vi) bioassays to verify the effects of the identified semiochemicals on insects in vivo. The key aspects of these approaches are described in the following sections.
3.4.1. Conventional Chemical Ecology Approaches
The components of the complex blends of tephritid pheromones can be isolated, identified and tested using interdisciplinary approaches involving bioassays, sensory physiology, analytical and organic chemistry, and biochemistry. Recent breakthroughs in analytical techniques allow the rapid screening of semiochemicals with more sensitive bioassays and their isolation and identification from relatively smaller amounts of material. Several non-destructive and artefact-free methods are available for collecting natural odorants from living organisms. Examples include the adsorption of odours on different polymer matrices contained in cartridges or filters. Trapped odours can be desorbed thermally or eluted with organic solvents followed by analysis using gas chromatography. The direct coupling of a chemical (flame ionization detector and/or mass spectrometer) and biological detector (e.g., the use of an insect antenna) permits simultaneous isolation and identification of bioactive components from trapped odours. Furthermore, the discovery of more efficient chemical synthetic methods now allows the state-of-art synthesis of semiochemicals of high purity whose field activity may provide answers to ecological and evolutionary questions associated with the importance of the chemical in the behaviour of the target insect [259].
Volatile Collection and Analytical Techniques for Their Identification
Chemical identification of tephritid volatiles requires a chromatographic separation followed by detection using spectrometric analytical methods. Although a wide range of methods is available in principle, the number which is suitable in practice depends upon the amount of insect material that can be obtained.
Solvent extraction of tephritid rectal glands using standard solvents such as heptane, hexane, methanol, ethanol, dichloromethane and ethyl ether is commonly used [166,213,231,232]. More recently, mixtures of solvents (acetonitrile/water and methanol/acetonitrile/water) were applied for the extraction of semiochemicals from the whole body of adult medflies [260].
Solid phase microextraction (SPME) is a solvent free, pre-concentration technique developed by Arthur and Pawliszyn [261] for application in solid, liquid, or gaseous samples (reviewed in [262]). The results obtained using SPME fibres are similar to those obtained with solvent extraction [263,264,265]. SPME is a known and effective alternative to liquid-liquid extraction. It provides some advantages over liquid-liquid extraction process because of reduced solvent consumption. In tephritids, different SPME coating materials have been used for volatile collections. A polydimethylsiloxane/divinylbenzene fibre (PDMS/DVB) was applied for pre-concentration of volatile compounds emitted by male and female medflies in different mating status (virgin or mated), and age (3 or 9 days old) [222]. In total, 70 compounds of diverse chemical classes such as alcohols, acids, aldehydes, terpenes, branched hydrocarbons and esters were reported and identified by gas chromatography coupled with mass spectrometric detection (GC-MS) [222]. Similarly, PDMS fibres have been used to compare the composition of the pheromone of medfly males from a standard laboratory strain reared as larvae on laboratory media and fresh fruits [220]. Five and 30 day-old males have been used, with 36 and 27 chemicals (mostly belonging to terpenes, amides, esters and alkanes) identified to be emitted by these two age-classes, respectively [220]. In a recent work on C. capitata volatiles, divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS) combined with GC-MS and gas chromatography-flame ionization detection (GC-FID) techniques resulted in the identification of 27, 23 and 29 compounds from larvae, pupae and adults, respectively [266]. PDMS/DVB SPME fibres have been applied to collect volatiles produced by B. zonata males and females and R. batava males, respectively [61,213].
Dynamic headspace is a common method for the collection of volatiles produced by fruit flies. The volatiles emitted by virgin-calling flies are collected using a modified technique of an air-collection apparatus as described by Nation [149]. In this technique, a purified airstream is blown over living flies enclosed in a glass chamber. The volatiles are collected onto traps containing adsorbents such as SuperQ, Tenax, Activated Carbon, etc. [213,221,223]. The air flow directed through the apparatus is controlled by flowmeter. Volatile collections are usually performed for 24 h. Afterwards, the traps are washed with heptane, hexane, diethyl ether or ethanol and the obtained extract is analysed by GC-MS, electroantennography, gas chromatography coupled to electroantennographic detection (GC-EAD) methods and used for bioassays.
GC-MS is one of the most useful tools for chemical analysis of volatile semiochemicals. The gas chromatography provides high-resolution separation of components within a complex mixture, and the mass spectrometric detection supplies structural information in addition to its role as a sensitive detector (the current limit of detection is at femtomole levels). By selection of the appropriate capillary column, practically all volatile organic compounds can be separated, including carboxylic acids, ketones, aldehydes, alcohols, aromatic compounds, and hydrocarbons [180,215,226,267,268,269,270].
For identification of the absolute configuration of tephritid volatile semiochemicals, the chiral column or chiral GC-MS can be used, as recently applied for pheromone identification of R. batava [61]. Most commercial chiral GC phases currently available are composed of modified cyclodextrins, which give a wide range of enantiomeric separations, but have the disadvantage of being thermally unstable above 200 °C, and are therefore useful for relatively volatile compounds only. Like chiral nuclear magnetic resonance (NMR) studies, chiral GC requires homochiral or enantioenriched synthetic standards, but unlike the NMR technique, only nanograms of impure material are necessary [271]. Two-dimensional gas chromatography with time-of-flight mass spectrometric detection (GC × GC-TOFMS) is a recently developed analytical technique that offers a solution to the chromatographic co-elution and provides high sensitivity and selectivity. In principle, the method consists of two GC systems (GC × GC) equipped with columns of different polarity connected by an interface with an integrated cryogenic trap. The cryogenic trap repeatedly condenses compounds eluting from the primary column and releases them periodically as short pulses to the secondary column. Parameters like duration and frequency of both condensation and injection pulses are variable and allow precise tuning of the instrument according to the requirements of the analysis. Since the GC × GC produces very narrow peaks (down to 50 ms, depending on the frequency of the cryogenic modulation) a time-of-flight mass spectrometric detector (TOFMS) with a high acquisition rate (up to 500 spectra per second) is required. The pulsed nature of the TOFMS source of ionisation further enhances the system accuracy by avoiding the spectral skewing common in a continuous ionisation mode. GC × GC with TOFMS detection thus operates with a high precision independent of the concentration range [272,273]. This method has been applied for analyses of fruit fly semiochemicals produced by species of the genera Anastrepha, Bactrocera and Ceratitis [177,214,221,274].
Gas chromatography coupled with Fourier transformed infrared spectroscopy (GC-FTIR) is relatively sensitive (detection threshold 10–100 mg) and particularly useful for identifying geometrical and positional isomers or functional groups. Thus GC-FTIR is an ideal instrument for the study of volatile organic compounds. Medfly synthetic attractants composed of trans-trimedlure isomers and cis-trimedlure isomers were analysed by GC-FTIR spectroscopy [275].
Matrix-assisted laser desorption/ionization with time-of-flight mass spectrometry (MALDI-TOFMS) and desorption electrospray ionization (DESI) mass spectrometry are soft ionization techniques developed for the analysis of biomolecules (biopolymers such as proteins, peptides and sugars) and large organic molecules (such as polymers, dendrimers and other macromolecules). However, they have been applied very sporadically for studies of the fruit fly pheromone components [276].
High performance liquid chromatography (HPLC) coupled to a MS detector is at present the favoured technique for the study of polar non-volatile tephritid semiochemicals. Other types of detectors that are commonly used with HPLC separation are UV and ELSD (evaporative light scattering) detectors [277,278,279,280]. Normal phase and reversed phase HPLC separation are still popular in lipid analysis. The mobile phases usually consist of methanol, propan-2-ol, or n-hexane. Chromatographic columns with C18-phases still prevail. A few separations were reported on C8-columns. In a recent metabolomic study of B. dorsalis larvae, liquid chromatography-mass spectrometry (LC-MS) and GC-MS were applied for the characterisation of endogenous metabolite changes and biochemical effects of azadirachtin [281].
NMR spectroscopy is one of the most informative, but least sensitive, modern spectrometric methods. Fourier transform 1H and 13C NMR spectroscopy require approximately 100 μg and 10 mg pure material, respectively (50 ng – 1 μg with very expensive NMR-nanoprobe). Since many insect pheromones are only present in nanogram amounts, a very high number of individuals are required. NMR is widely used to determine the absolute configuration of optically active compounds (semiochemicals), either using chiral shift reagents, or by converting enantiomers to diastereoisomers prior to spectroscopic study, or using chiral solvent. The use of NMR for absolute identification of semiochemicals has two main disadvantages: isolation of the pure substance is required, and an enantioenriched or a homochiral synthetic standard is needed. In modern semiochemical research, NMR spectroscopy is normally used only when insufficient structural information is provided by more sensitive methods [282]. Baker and Heath [283] applied NMR spectroscopy for the identification of lactone pheromone components emitted by A. suspensa and A. ludens.
Chemical Synthesis
The chemical synthesis of tephritid pheromones is a key step towards producing pheromone compounds for use in bioassays (usually very low amounts), and for insect pest monitoring and management purposes. Moreover, their absolute configuration is an important determinant of their biological activities [284]. In 1973, Jacobson and colleagues isolated and synthetized the first medfly pheromone components, i.e., methyl (E)-non-6-enoate and (E)-non-6-en-1-ol that were attractive eliciting also sexually excitatory response to females in laboratory trials [215]. Three additional major components of the medfly pheromone, namely ethyl (E)-oct-3-enoate, geranyl acetate, and (E,E)-α-farnesene were isolated and synthetized later [225]. In 2010, Olszewski and Grison [285] reported a novel and versatile synthetic approach to the preparation of (E)-non-6-en-1-ol.
Since the first successful studies on the isolation and synthesis of the medfly pheromone components, several efforts have been made to characterise the volatiles emitted by Bactrocera and Rhagoletis species, mainly using the enantioselective approach. Moreover, in the case of chiral pheromones, which occur in tephritids, the synthesis and following bioassays using stereoisomers are essential for recognition of the stereochemistry-bioactivity relationships. This knowledge also has practical implications for pest control as in most chiral pheromones only one enantiomer is biologically active [284]. In the case of B. oleae, olean (1,7-dioxaspiro[5.5]undecane) was isolated and identified as the major component of the female-produced sex pheromone in 1980 [79]. (4R*,6R*)-4-hydroxyoleane and (3R*,6R*)-3-hydroxyoleane (the asterisks indicate the chiral centers) were then isolated as minor components [79]. Bioassays involving the synthetic enantiomers of oleane showed that (R)-oleane is active on males, while (S)-oleane on females, and GC analyses showed that females can produce (±)-oleane [207]. The synthesis of enantiomers of 4-hydroxyoleanes was conducted by Mori and co-workers [286] and a recent study identified and synthesised 11 new B. oleae female-specific components [287].
Knowledge of pheromone composition in Rhagoletis is still limited to R. cerasi and R. batava. In this last species, Buda and colleagues recently applied GC-MS analyses of fly headspace and found that males emit (-)-δ-heptalactone. The authors then synthesised the two enantiomers of (-)-δ-heptalactone using enantioselective synthesis and found that only (-)-δ-heptalactone elicited an electrophysiological response in both sexes, proposing that this chemical may act as aggregation pheromone [61].
Despite these research efforts, similar studies focusing on the optimisation of chemical synthesis of pheromone components are completely missing for the genus Anastrepha, as well as for many other species belonging to the Bactrocera, Ceratitis, Rhagoletis and Zeugodacus genera.
Identification of Electrophysiologically-Active Volatiles
GC-EAD uses the electrophysiological response from a dissected insect antenna to assign activity to a gas chromatographic peak. The GC eluent is normally split between a flame ionization detector (FID) and the insect antenna, which is connected to an amplifier and recorded by silver or platinum electrodes. The response from each detector is recorded simultaneously so that an EAD response can be correlated with a specific peak in the FID chromatogram. This technique is particularly useful for the assignment of activity to chemicals which may be present as minor components in complex mixtures, although it provides no information about the function of the active compounds. GC-EAD analyses of male pheromone or rectal glands extracts have been performed on several tephritid species, including C. fasciventris, C. anonae and C. rosa [214], C. capitata [221,241], B. oleae [288], A. serpentina [193], A. fraterculus [177,289,290,291], A. obliqua [243] and A. striata [292]. Additional studies performed GC-EAD analyses on the headspace volatiles of both sexes, in the species R. batava [61], Bactrocera frauenfeldi (Schiner) [293], B. musae [76], and C. capitata [223]. In addition to GC-EAD, gas chromatography-electropalpogram detection (GC-EPD) has begun to be used to test the responses of tephritid maxillary palps to the pheromone emissions [293].
Furthermore, electroantennography-based experiments can be performed independently from gas chromatography. In this case, pure compounds are employed to assess the response to a certain stimulus and the required dose to elicit a detectable response. In this case, chemicals are delivered to insect antennae/palps in controlled conditions by pumping the odour from a reservoir [256]. Since early works in the ’80s [244,294,295], numerous EAG-based studies have been performed in tephritids to test the electrophysiological response to volatiles of fruits and flowers [296,297,298,299], artificial attractants [300,301,302,303,304], as well as pheromone components [305,306]. The use of GC-EAD brought the EAG-based approaches to a higher level of sophistication by using the antenna/palp of the target insect as a detector for the gas chromatograph. However, while electroantennography is an excellent technique to quickly measures the change in the electrical potential between distal and proximal sections of the antenna/palp provoked by olfactory stimulation, EAG amplitude depends on the position of the electrode, the strength of the connection, and insect vitality [305,306]. Thus, EAG is considered a qualitative indicator of olfactory reception [307]. Also in tephritids, EAG signals are indeed known to vary in relation to the relative density of sensilla, which display a specific distribution on the funiculus, at the electrode location [308,309]. To increase the recording specificity, a novel method was developed integrating EAG recordings at multiple antennal positions with current source density (CSD) modelling [310,311] useful to map the functional activation of individual antennae. This method was applied to six tephritid species, i.e., C. capitata, C. catoirii (Bezzi), Neoceratitis cyanescens (Bezzi), B. zonata, Z. cucurbitae, and Dacus demmerezi (Bezzi) to measure the response to seven volatile chemicals emitted by fruits and plants at four position in the funiculus.
A more quantitative measurement of the olfactory response, although requiring significant experimental effort on multiple individuals, can be achieved using single sensillum recording (SSR). SSR allows to measure the action potentials generated by olfactory sensory neurons (OSNs) within individual sensilla on antennae or palps using an electrode in contact with the lymph of the extracellular receptor [307]. To the best of our knowledge, this approach has been used only in two tephritids so far, but involving, in both cases, fruit odours as stimuli. The sensilla located on the tarsi of C. capitata [312], and the antennae of R. pomonella [313] were investigated.
Behavioural Assays to Identify Active Compounds
Bioassays play key roles in the study of tephritid semiochemicals [271]. Bioassays allow to assess the behavioural effect(s) (e.g., lek formation, male signalling, courtship, copulation, host finding for oviposition, HMP deposition) of synthesised active compounds (pure or in mixture) with respect to host or mate interactions, differently from EAG assays that provide the electrophysiological responses of isolated organs, structures or olfactory receptors (ORs). The biological response measured by a bioassay is essential to attribute to a molecule or a mixture of chemicals the pheromone identity. Behavioural effects in tephritid species can be evaluated in laboratory, semi-field and field conditions where both the stimuli and the background in which the stimuli are presented are tightly controlled [314]. Techniques for insect bioassays have been widely reviewed (see for example [256,315,316,317]) and widely applied to tephritid species, for which most experiments have been so far performed in laboratory conditions. Initial studies were done in close-range cage bioassays estimating medfly female attraction to male pheromone components based on landings on filter paper soaked with olfactory stimuli [218]. Currently used setups adopted for the study of tephritid olfaction include (i) two-choice systems, (ii) flight tunnels, and (iii) multi-arm olfactometers. Two-choice systems, using arenas (i.e., observation chambers) [174,318,319], as well as Y-tubes/T-maze [85,231,293,320], have been used to evaluate attractive or repulsive responses following exposure to pheromone (isolated from headspace or rectal glands); in this framework, dynamic systems like the Y-tubes, allowing an air flow carrying the chemicals to be evaluated for the chemo-ecological role should be preferred, since they avoid the risk of saturating receptors of the tested flies, which is common in still-air arenas. Flight tunnels, also known as wind tunnels, are extremely useful to monitor medium-distance flight responses to mate-derived chemicals [201,270,321]. Their use, for example, allowed to assess the attraction of females to male emissions in C. capitata [270] and A. serpentina [193], or of both sexes to ME-fed B. dorsalis males [70]. Wind tunnels also permitted to prove that pheromone components can exert behavioural effects in A. ludens females [181]. These systems can integrate different components for the simultaneous identification of released volatile chemicals and the assessment of their attractiveness, as well as recording environmental parameters and fly activity [201], including the observation of flight patterns in females responding to male-derived volatiles [321]. Lastly, multi-arm olfactometers have been mainly used so far to test fruit odours and with flies released into an area composed of multiple chambers from which airflow-containing odour flows [322].
Moving laboratory results to the field for real-world applications is a timely challenge in tephritid research. Therefore, field assays to evaluate tephritid attraction should be considered after successful laboratory evaluation of a given compound. Field studies can be performed either using live insects/glands extracts as a source of pheromone or traps with different types of dispensers releasing pheromone pure chemicals or mixtures, as described especially for the medfly and the olive fruit fly. In the case of C. capitata, a synthetic blend releasing three male pheromone compounds (i.e., ethyl (E)-oct-3-enoate, geranyl acetate and (E,E)-α-farnesene), in a ratio similar to that observed in natural conditions, was effective in attracting females, as shown by trap catches [225]. Another study found that trimedlure was more effective than pheromone individual components or mixtures in trapping flies [323]. For B. oleae, field experiments aimed at determining the attractive effect of the four major pheromone components stressed the importance of finding the ideal combination between attractant formulation and trap type/colour [324]. The use of polyethylene vials as dispensers of either the complete pheromone blend, racemic mixtures of the major components, or individual synthetic chemicals resulted effective in trapping B. oleae flies [319,325]. Open field tests were performed either with wild medfly females [225] or released C. capitata and A. suspensa females [326,327,328,329]. Tests in field cages with potted trees have also been performed, providing valuable information about behavioural responses to live conspecifics or male extracts in seminatural conditions for A. obliqua, A. ludens and A. suspensa [188,244,330].
3.4.2. Reverse Chemical Ecology Approaches
In addition to the above-described techniques to identify tephritid semiochemicals, recent studies are increasingly showing that RCE has great importance in understanding the molecular basis of insect chemical perception and identifying the active volatile semiochemicals [257]. Studies began to be performed to identify genes involved in chemosensory perception in tephritids, including olfactory, ionotrophic and gustatory receptors (ORs, IRs and GRs, respectively), OBPs, odorant degrading enzymes (ODEs), and chemosensory proteins (CSPs) [331]. Insect OBPs are small soluble proteins mostly found in the chemosensillar lymph of sensory organs where they bind molecules of odorants and pheromones (see [332] for a review). Thus, OBPs are considered ideal molecular targets for binding assays to identify chemicals with a potential behaviourally-active role in tephritid biology. The expression of OBPs, followed by their purification and structural analysis, is indeed adopted to perform OBP ligand binding studies; ligands are screened from sets of volatiles emitted from host plants, pheromones, or synthetic attractants used in field applications, and once identified, behavioural responses are evaluated in vivo [256].
Identification and Functional Analysis of OBP Genes
Putative OBP genes have been initially identified through expressed sequence tag (EST) approaches in C. capitata [333], B. dorsalis [334,335], R. suavis [336], and R. pomonella [337] and their transcriptional profile started to be explored. Subsequently, with the advent of next generation sequencing, more OBPs have been discovered through mining of RNA-seq data and whole genome sequencing in several tephritid species, including B. dorsalis, B. minax [104,338,339,340,341], A. fraterculus, A. obliqua [106,342,343], and C. capitata [344]. Antennal proteomics profiling has been applied to B. dorsalis to identify differentially expressed genes, including OBP genes, in ME-responsive males [345]. Functional studies have been performed to assess their role in odour perception. These include tracing OBP genes expression profiles in different tissues, developmental stages and in response to maturation and mating, RNA interference (RNAi) combined to electrophysiology to assess the involvement of target OBPs in odour detection, followed by behavioural assays [335,341,345,346,347].
Purification and 3D-Structural Analysis of Identified/Expressed OBPs
The binding specificity of OBPs expressed in the main chemosensory organs, i.e., antennae and maxillary palps, may help in the identification of pheromone/pheromone components that are still uncharacterised. Thus, OBPs that are (i) abundant in olfactory tissues, or, ideally, specific to these tissues, and (ii) showing sequence similarities to already characterised proteins known to be involved in chemical communication in other insects can be functionally characterised by using ligand-binding assays. OBPs are expressed in bacterial or yeast systems and the recombinant proteins purified with chromatographic steps using different techniques, including anion-exchange or gel filtration chromatography, or affinity chromatography on nickel columns when histidine-tags are added to the OBP sequence (see [348] for a review). Purified proteins can then be used for ligand-binding experiments and to solve their structure through X-ray crystallography or NMR spectroscopy [348].
The first, and, so far, the only available, structural characterisation of a tephritid OBP was recently obtained in the medfly using X-ray crystallography [349]. The structure of CcapOBP22 is characterised by six α-helical elements, a typical feature of insects’ OBPs, interconnected by three disulphide bridges. Differently from other insect OBP structures, CcapOBP22 also carries a 7th α-helix at the C-terminus, which contributes to delimit the ligand-binding pocket. CcapOBP22 was co-crystallised with (E,E)-α-farnesene as ligand, further supporting the potential role of this protein in semiochemical perception in this species.
In vitro and in silico OBP: Ligand Binding Assays
Several approaches have been adopted to measure the affinity of OBPs to odorants (see [350] for a review). The most common method, which is fast and requires a limited amount of protein, is based on the use of fluorescent reporters, such as 8-anilinonaphthalene-1-sulfonic acid (ANS) and N-phenylnaphthalen-1-amine (1-NPN) in competitive binding experiments [351,352,353]. 1-NPN is a lipophilic crystalline solid that strongly binds insect OBPs [354]; when increasing amounts of a tested ligand are added to the OBP/1-NPN system, decreasing 1-NPN fluorescence emission is inferred as 1-NPN displacement since the ligand is assumed to compete for the binding pocket initially occupied by the fluorescent reporter.
This approach has been used in the medfly to evaluate the binding affinity of CcapOBP22 and CcapOBP24 to electrophysiologically-active components of the male pheromone, as well as to the two synthetic attractants trimedlure and ME [223,349]. The finding that also ME, which is a strong attractant for some Bactrocera species [170,355,356] but not for medfly, displays binding activity (although moderate) to the above medfly proteins is intriguing. Methyl eugenol is known to induce an electrophysiological response in medfly [302]. In Z. scutellatus, ME elicits significant electrophysiological responses too, but it is not behaviourally active [357]. In medfly, ME has been shown to induce poor behavioural responses in binary choice bioassays, while o-eugenol was instead strongly attractive [302]. Thus, it appears that the presence of substituents on the aromatic ring can be essential to confer attraction to chemical compounds. It will be interesting to further explore the chemistry of candidate molecules able to bind tephritid OBPs to shed new light on structures that can be exploited as novel attractants. Both OBPs showed the highest binding affinity to (E,E)-α-farnesene, which is one of the major components of medfly male pheromone bouquet, and is known to attract females [321], suggesting its role as a natural ligand for these OBPs. The verification of the behavioural responses to the presence of ligands in vivo is essential to identify volatile semiochemicals with active roles in fruit fly behaviour.
Ligand-binding assays have also been performed in B. dorsalis using 13 chemicals, including pheromone components and attractant molecules, and six proteins with high expression in the antennae (five OBPs and one CSP) [318]. Authors showed that OBPs displayed the highest affinity to the attractants, and, in the case of BdorOBP83a-2, RNAi led to a decrease in neuronal responses to tested molecules, as shown by EAG recordings and behavioural responses.
Computational reverse chemical ecology (CRCE) is another method applied to the discovery of behaviourally active chemicals [358,359]. OBP sequences can be exploited for 3D model prediction, producing 3D structure for docking studies using specific tools. Molecular docking is commonly employed in pharmaceutical research for structure-based drug design [360,361]. It implies the use of programs based on different algorithms applied to model the interaction between a small molecule and a protein at the atomic level. This allows the exploration of the behaviour of small molecules in the binding pocket of target proteins [362]. CRCE has been implemented in B. dorsalis to screen 25 chemicals for their binding potential to a general OBP (GOBP) showing that this approach may be extremely useful to quickly predict behaviourally-active semiochemicals, for example selecting chemicals belonging to specific classes [358]. The described approach is particularly beneficial especially in tephritids given the wide absence of direct crystallographic data for OBP binding modes.
Identification and Functional Analysis of OR Genes
Although OBPs are excellent study targets to either understanding the molecular and biochemical mechanisms of odour perception in insects, and to explore the development of pest control agents, they have broad binding specificity, are also distributed in non-olfactory tissues and have different functions [363]. Conversely, ORs are transmembrane proteins showing high specificity and sensitivity. Thus, genes encoding for chemosensory receptors are also becoming to be identified and characterised in tephritids, such as B. dorsalis [338,364,365,366], B. minax [341], B. oleae [367], B. latifrons and Z. cucurbitae [368], and C. capitata [344]. Olfactory receptors have been described as heteromeric ligand-gated ion channels consisting of a specific OR and the highly-conserved co-receptor Orco [369]. Olfactory receptors are transmembrane proteins for which no 3D structure is available yet, and they are a more difficult target than OBPs to be expressed and purified in heterologous systems. So far, the only three-dimensional structure currently available, obtained using a cryo-electron microscopy-based approach, is for a tetramer of Orco, described in the parasitic fig wasp Apocrypta bakeri (Joseph) [370]. Thus, only limited data on the functional activity of tephritid ORs are available. In a recent study, ten B. dorsalis ORs were co-expressed with their essential co-receptor BdorORCO in Xenopus laevis Daudin (Anura: Pipidae) oocytes. Two-electrode voltage clamp was then used to record currents from injected oocytes when ligands (i.e., 1-octen-3-ol, geranyl acetate, farnesenes, and linalyl acetate) were diluted in the assay buffer [365]. Some of the identified ORs have been shown to respond to plant volatiles [365] or ME [371]. Further research efforts oriented to clarify the structure of tephritid ORs are essential to understand the molecular recognition mechanisms they are involved in, as well as their interactions with OBPs, and thus their functional roles. Insect ORs display a different topology from those of other animal G protein-coupled receptors (GPCRs) [372], with a C-terminal faced to the extracellular section and the N-terminal to the intracellular section. This feature makes insect ORs ideal targets to be explored for the development of insect-specific pest control strategies. These may include the inhibition of either Orco or the ORx/Orco complex by antagonists able to, for example, disrupt mating behaviour through the manipulation of pheromone receptivity. Interestingly, in B. oleae, transient knockdown via RNAi gene silencing in adult individuals showed that knockdown of Orco expression reduces the mating ability in both sexes and completely inhibits oviposition [367].
4. Host-Marking Pheromones
4.1. Chemical Identity, Production and Analytical Approaches to Their Characterisation
In tephritids, host-marking behaviour was first described in R. pomonella [374]. Later, Hafliger speculated that the biological role of this behaviour was to equally distribute the offspring in available host fruits [375]. Following these earlier observations, Prokopy and Cirio were the first that experimentally described in 1972 the HMP deposition in R. pomonella [112] and R. completa (Cresson) [135]. Since then, the host marking behaviour has been reported in 25 tephritid species, particularly frugivorous species, belonging to the Anastrepha, Ceratitis and Rhagoletis genera [114] (Table 3; Table S3).
However, it is not a general feature of the family; it seems to be common in Rhagoletis spp., sporadic in Anastrepha and Ceratitis spp. and rather absent in others (e.g., Bactrocera spp.) [29,35,129,390,391].
HMPs are low-volatility and highly polar molecules [392]. They are also soluble in water and methanol [30,131,382,393,394]. HMPs can persist on the surfaces either when they are directly deposited by the fruit flies or as extracts [108]. For instance, the HMP half-life has been estimated to 10.7 days with persisting activity for three weeks in R. pomonella [383], 9 days in R. fausta [387], 12 days in R. cerasi [386], 6 days in A. suspensa [376], 6 days in C. capitata [111] and 4 days in R. indifferens [392].
To the extent of our knowledge, HMPs are produced and stored in the posterior half of the midgut and, thus, the faecal matter contains a huge quantity of these pheromones, suggesting the existence of two main routes for HMP deposition: through ovipositor dragging after egg-lying and through defaecation [132,395,396]. In R. pomonella, HMP accumulates in the midgut, Malpighian tubules, hindgut and faeces of mature females [395].
HMPs have been isolated from faecal matter extracts using approaches based on liquid chromatography (LC) and MS, in all four species (Table 3), namely HPLC-FAB-MS (Fast Atom Bombardment Mass Spectrometry) for A. ludens [382] and R. cerasi [30], and LC-quadrupole time-of-flight-mass spectrometry (LC-QTOF-MS) in the case of Ceratitis species [390,397]. Therefore, all the HMPs that have been chemically identified were isolated from the aqueous or methanol extract of adult female faecal matter. To date, it remains to be determined whether HMPs are produced by specific glands [395].
HMP chemical identity has been so far determined in a few tephritid species (Table 3). The first chemical characterisation of an HMP was achieved in R. cerasi. The pheromone was a complex molecule, i.e., N-[15(β-glucopyranosyl)-oxy-8-hydroxypalmitoyl]-taurine, with four stereoisomers [30], showing two chiral centres at the C-8 and C-15 positions. After the synthesis of the four different stereoisomers [398], it has been demonstrated that a racemic mixture of two isomers (8R, 15S and 8S, 15R isomers) is able to deter oviposition [399].
Later, the HMP [2-(2,14-dimethylpentadecanoylamino)pentanedioic acid (or N-[2,14-dimethyl-1-oxopentadecyl)glutamic acid)] of A. ludens has been chemically characterised and synthesised [382,400]. It presented a relatively simple structure containing an isopalmitic fatty acid chain substituted by methyl at the C-2 position and coupled to glutamic acid (GA) as a single diastereomer [382,400]. The HMP of A. ludens exhibited not only intraspecific but also interspecific oviposition deterring activity to A. obliqua and A. serpentina [132]. Of note, the HMPs of R. cerasi and that of A. ludens display similarities in structure (i.e., both contain a long fatty acid residue attached to an amino acid).
Recently, the HMP of C. cosyra and C. rosa have been isolated and both chemical structures have been determined [33,34]. The C. cosyra and C. rosa HMP identified are the tripeptide glutathione (GSH) (consisting of glycine, cysteine, and GA [33]), and GA [34], respectively. Interestingly, GSH and GA levels were 5–10 and 10–20 times higher in the faecal matter than in the ovipositor or haemolymph extracts of the respective females. These results suggest that the HMPs may be transferred from the gut into the ovipositor through the haemolymph and the excess amount may be expelled with the faecal matter. GSH was shown to express pheromone and allomone action respectively, reducing the oviposition in individuals of the same species and in those of different species, such as C. rosa, C. fasciventris, C. capitata, Z. cucurbitae. Interestingly, the GSH acts as kairomone inducing arrestment behaviour in the egg parasitoid Fopius arisanus (Sonan) (Hymenoptera: Braconidae). On the other hand, GA perception resulted in oviposition reduction in C. rosa and C. fasciventris, but not in C. cosyra. It is noteworthy that the HMPs of the two Ceratitis species are highly distinct from the HMPs identified in the other fruit flies. However, they appear to be more closely related to the HMP of the Mexican fruit fly, which contains GA, than of R. cerasi that is a fatty acid glucoside. Taken together, these findings may indicate that closely related species may utilise a similar pathway for HMP synthesis.
Host marking through HMP deposition is noticeably absent in the Bactrocera genus, even if contrasting results have been reported [35,139]. To prevent other females from ovipositing on the same fruit, B. oleae females do not depose an HMP but use their labella to spread olive juice leaking from the oviposition wound, with the main compounds responsible for this repulsion being (E)-hex-2-enal and oleuropein derivatives, such as the hydroxytyrosol [143,401].
The molecular machinery underlying the perception of these substances by other females remains to be determined. However, early studies in R. pomonella suggested that D-hairs on specific segments of the ventral tarsal surface and short marginal hairs on the labellum carry the receptors for HMP detection [395].
4.2. Behavioural Assays
5. Cuticular Hydrocarbons
Cuticular hydrocarbons act as pheromones in a variety of orders, including Diptera [25,403,404,405]. Their behavioural function in flies was first described in the housefly, Musca domestica L. (Diptera: Muscidae), where (Z)-tricos-9-ene was identified as the main compound on the female cuticle acting as a sex pheromone for males [403,406,407,408]. Extensive evidence of the importance of hydrocarbons (7-monoenes) as short-range signals and contact pheromones comes from Drosophila spp. [23]. These semiochemicals are perceived by antennae and maxillary palps and/or by contact with the taste organs that are mostly located on the tarsi and proboscis [409,410,411]. Drosophila spp. show relatively stable CH profiles, although their production can vary as the flies age and even after reproductive maturation [412,413]. In tephritids, sex- and species-specificity of CHs have been described in species of the genera Anastrepha, Bactrocera, Ceratitis and Zeugodacus (Table 4; Table S4).
Characteristic CHs profiles have been successfully applied for the chemotaxonomic clarification of fruit fly species complexes of A. fraterculus, B. dorsalis and the so-called African Ceratitis FAR complex [90,274,414,416,417,418,420,421]. Nevertheless, studies focused on the elucidation of CHs behavioural function in fruit fly mating system are still missing, except for those performed on B. oleae [80,85,288].
5.1. CHs in Tephritid Species and Their Described Roles
Earlier comparative studies on adults of A. ludens, A. suspensa, C. capitata, C. rosa, Z. cucurbitae and B. dorsalis failed to identify substantial sex-specific differences in CH profiles [414,420]. It seems that the only difference detected is the much higher amount of n-alkanes in males compared to females in both B. dorsalis and Z. cucurbitae [420]. However, further research reported sex- and age-dependent differences in CH production for a laboratory population of A. fraterculus (Brazilian-1 morphotype). Sexually mature males have specific unsaturated hydrocarbons (7-monoenes) on their cuticles that lack in females [90,289]. In follow-up studies, sexual dimorphism has been evaluated in the Brazilian-1 (Argentina), Brazilian-3, Andean, Peruvian and Mexican morphotypes of the A. fraterculus species complex [92,93]. The Brazilian-1 morphotype expresses the highest sexual dimorphism (29.46%), followed by the Mexican (15.42%) and Peruvian (13.79%) ones [93]. Males and females from the five abovementioned morphotypes diverge in alkene and alkadiene content. In A. ludens, the age-dependent CHs production in males originated from a standard mass-reared colony, a genetic-sexing strain, a hybrid strain and a wild population has been recently described [415]. Wild males of A. ludens differ from the mass-reared strains in the amount of nonacosane, while genetic sexing strain expressed higher values of 2-methylhexacosane. It has been suggested that the observed differences in CHs profiles may be due to environmental pressures [415], but additional research efforts are still needed to clarify these issue.
Further research also focused on the CH composition of males and females of the African fruit fly cryptic species FAR complex, demonstrating that sex-specific differences in the CH composition do exist [214,274]. The CH sex-specificity was proved by multivariate statistical analyses of the GC×GC-MS data of 59 CHs identified in the epicuticular extracts of C. capitata, C. fasciventris, and C. rosa. In contrast, the cuticular profiles of C. anonae display no sex-specificity [274].
In species of the B. dorsalis complex, abundant complex mixtures of sex-specific oxygenated lipids (i.e., three fatty acids and 22 fatty acid esters) with so far unknown biological function were identified in epicuticle extracts from females [421]. Such sex-specificity may be driven by sexual selection if the chemical composition of the cuticle is used as a pheromone signal in mate choice. Although early studies suggested that B. dorsalis males are able to recognise females at short distance and that physical contact may play key roles in courtship and copula [424], only limited functional information are currently available. These include the data describing the strong attractiveness to males exerted by the female-specific CH 4-allyl-2,6-dimethoxyphenol (4-DMP), which has been regarded as close-range sex pheromone [425]. This compound elicits electrophysiological responses in the mid legs of B. dorsalis males [426]. Moreover, after stimulation with 4-DMP, five OBP genes are found upregulated in males, with one of them, BdorOBP2, being a promising candidate for the binding and transport of 4-DMP [426], in addition to its proposed role in the perception of ME [345]. Recently, the cuticular components of B. tryoni have been described [423]. The spiroacetals and esters were found to be female-specific, while amides were presented in both sexes. Nevertheless, the role of these and other CHs in short-range chemical communication of Bactrocera spp. and other fruit flies needs to be further elucidated through expansion of the molecular machinery underlying their production and perception and with proper behavioural assays.
5.2. Analytical Approaches to Trace CH Profiles
Preparation of samples for gas chromatographic analysis of tephritid CHs is usually made by solvent extraction of whole insect bodies with solvents such as pentane, hexane, dichlormethane, and chloroform [90,427]. The most widely used method for recovering tephritid CHs is hexane washing [274,421,428]. This process may contaminate the cuticular sample with other materials, such as those from the exocrine glands. To collect only the CH fraction of the extract, the hexane solution is placed on a short chromatographic column and the CHs eluted with a small volume of hexane. For pre-cleaning of the CHs, thin layer chromatography (TLC) has proven useful in identifying novel lipid pheromones. TLC plates consist of glass or aluminium coated with an adsorbent layer of silica gel. Using TLC, components of the chemical extract can be separated into discrete fractions according to hydrophobicity [429,430]. After the hydrocarbon fraction has been collected, the solution must be concentrated to a suitable volume before the analysis. For the determination of the final concentration of the sample, the number of insects extracted must also be known. Another reported method involves continuous extraction of the insect in a refluxing solvent [431]. This procedure also requires column chromatography and re-concentration of solutes. Solvent extraction procedure involves several steps, in which more volatile compounds may be lost during the process, and moreover the use of high-purity solvents is required. Besides, the extraction and sample preparation account for most of the analysis time [428].
The solvent-free SPME technique has been used for the analysis of cuticular components of Trupanea vicina (Wulp), an Asteraceae-feeding tephritid [432]. SPME polydimethylsiloxane coated fibre (PDMS) was used to wipe samples from various body parts of male T. vicina and the subsequent GC-MS analyses showed that 1-nonanol, the male-specific compound, was concentrated on the abdomens of males exhibiting pleural distension [432]. In A. ludens, a PDMS fibre was used to rub down the male wing CHs, which were subsequently analysed by GC-MS [415]. Recently, the direct immersion-SPME (DI-SPME) coupled with GC-MS analyses was employed for characterisation of C. capitata semiochemicals in three different mating stages. This study demonstrated that medfly compound compositions were not significantly different before and during mating. However, new chemical compounds were generated after mating, such as (Z)-tricos-9-ene and hentriacontane among ten other components [260]. Female medflies seem to discriminate mated from virgin conspecifics and express higher rates of aggressive behaviour against the later [433]. Considering the recent findings, it is plausible to argue that changes in CHs of mated females drive the differential aggressive interactions between virgin and mated female medflies.
The method of choice for the analysis of the CH profile in tephritids has been GC-MS or GC×GC-MS using two different ionization techniques: electron (EI) and chemical ionization (CI) [93,289,427,434,435]. EI-MS is the primary tool for assessing the location of methyl branch points in long-chain alkanes, but it is often difficult to identify the molecular ion. The EI mass spectra interpretation allows complete identification of a compound, but often microscale reactions or derivatizations are necessary to provide additional structural information [436]. A mass spectrum also provides a ‘finger print’ of a compound, which can be compared with the libraries and mass spectra registries such as the NIST library, the Wiley/NBS registry of mass spectral data, and the published retention indices [437,438]. Mass spectra of an CI-MS yields the (M-1)+ ion as an intense peak (sometimes as the only peak), but when the peak consists of mixture of isomeric methyl or dimethyl alkanes, placement of methyl group position becomes more difficult [435]. Another problematic aspect is the determination of the double bound position using EI-MS, because of the lack of cleavage between carbon-carbon double bonds or extensive and facile hydrogen rearrangement along the chain after molecular ion fragmentation [439]. Nevertheless, several works documented the CI-MS/MS with acetonitrile ionization gas to be a suitable method for double bound position determination [90,434,440].
There have also been studies using MALDI-TOFMS and ultraviolet laser desorption ionization orthogonal time-of-flight mass spectrometry (UV-LDI o-TOFMS) for tephritid CHs identification [90,429,441]. Cvačka and colleagues [429] applied MALDI-MS for the identification of insect CHs, using lithium 2,5-dihydroxybenzoate as a matrix [442]. This work demonstrates that MALDI-TOFMS is a convenient analytical method for the identification of high molecular weight hydrocarbons from insect cuticles, including saturated hydrocarbons and highly unsaturated and/or cyclic compounds. In A. fraterculus, application of MALDI-TOFMS method allowed for characterisation of high molecular weight saturated and unsaturated hydrocarbons, up to C37 in length [90]. Mass spectrometric imaging (MSI) of male and female A. fraterculus was performed using a previously developed protocol [443]. The preliminary MALDI-MSI experiments indicate differences in the CH distribution on the wings of males and females [289]. Nevertheless, additional detailed analyses using MALDI-MSI techniques are necessary for further conclusions concerning the CHs of A. fraterculus. The imaging data will show if some of the CHs have unique locations on A. fraterculus body surface and can also indicate if the compounds direct male/female sexual contacts.
5.3. Behavioural Assays
Behavioural tools currently used for the evaluation of CHs do not substantially differ from the set up described above for volatile pheromones. Indeed, observation chambers, as well as two- or multiple-choice systems (i.e., Y-tube/T-maze and multiple-arm olfactometers, respectively) are widely used to evaluate behavioural responses triggered by CHs with potential pheromone activity [70,75]. Further validation of the observed behavioural response can be documented in flight tunnel as well as in field and semi-field assays [171,319,444].
As a final remark, semiochemical candidates, such as potential volatile pheromones and CHs, could be evaluated for their biological functions relying on the mixed society approach [445]. Indeed, mixed societies composed of living insects and small-sized robots mimicking their conspecifics can represent a valid approach to shed light on factors guiding insect behaviour, including mating approaches. This ethorobotics-based approach has been validated on several insect species, such as cockroaches, beetles and blowflies [446,447,448]. However, no studies have been conducted relying on ethorobotics in tephritid research. In our opinion, this represents a challenge for future studies.
6. Tephritid Sexual Chemoecology: Real-World Applications and Challenges
Semiochemical-based interactions have been extensively studied in tephritid fruit flies and several aspects of the generated knowledge had already been exploited for practical purposes including (a) trapping and population monitoring, (b) direct population control approaches (lure and kill methodologies, and push and pull strategies), and (c) support and improvement of other methods, such as the SIT. More recent progress in the characterisation of (a) the olfactory molecular machinery, including OBPs and ORs, and (b) CHs may open new venues in developing inspiring approaches for artificial olfaction and hence generation of novel long- and medium-range attractants. These tools can be used to address issues regarding species complexes that may be of vast importance for regulatory and control aspects. Nonetheless, a deeper knowledge of semiochemical-based communication is essential to further understand how tephritid species adapt to complex ecosystems, also with respect to the invasive potential of several pests belonging to this family, further advancing applied research from several perspectives.
6.1. Population Monitoring and Early Detection of Tephritid Outbreaks
Given the influence semiochemicals play on insect behaviour, a better understanding of their identity, specificity and biological role(s) may benefit tephritid research by developing novel specific and environmentally-friendly attractants. Effective attractants are essential for adult trapping, which is a key tool used to (a) monitor density and seasonal patterns of established tephritid pest populations, (b) detect new infestations of exotic species, (c) delimit the detected populations, and (d) confirm the results of eradication campaigns. Early detection of small populations is particularly important to delimit the outbreak and thus implement control and eradication measures while the pest population is still present at low densities [449,450].
The list of odour attractants for tephritid fruit flies is quite long including food-based (protein-based, ammonia releasing compounds) and mating-related chemicals. The later can either be purely synthetic or derived from plants. The history of their discovery and development goes beyond the purposes of this review, and their identity, efficacy, and the related practical aspects of their use in monitoring and detection are widely reviewed in [171]. Attractants that are related to the mating behaviour or physiology of fruit flies have been more thoroughly studied in the case of Bactrocera and Ceratitis genera and they exclusively concern male lures. For example, both the plant derived α-copaene and the synthetic trimedlure and Ceralure are highly attractive to male but not female medflies [171,451,452]. Interestingly, both plant-derived and synthetic attractants are related to male reproductive success and may enhance lekking behaviour and mating competitiveness [453,454,455,456,457,458]. It should be stressed here that trimedlure is rather the most commonly used attractant for medfly regular population monitoring, control programs and in detection and eradication campaigns [459,460].
Some Bactrocera species respond to Cue Lure (CL)/RK, others to ME, the most powerful attractant so far identified, others appear to be non-responsive to both. Eugenol analogues (isoeugenol, methyl-isoeugenol and dihydroeugenol) are also proving successful in attracting Bactrocera species [461,462]. Similar to species of the genus Ceratitis, the above compounds attract only males of the Bactrocera species and again are related to their mating success [463,464,465,466,467,468]. Methyl eugenol is considered one of the most powerful attractants for male fruit flies and it is extensively used in detection, population monitoring, delimitation and eradication campaigns worldwide [170,469,470].
Conversely, no male lures are currently available for Anastrepha and Rhagoletis spp. [171]. Exploring the potential of tephritid pheromones to develop novel specific attractants is thus important, not only for Anastrepha and Rhagoletis, but also for non-responsive species belonging to the Bactrocera and Ceratitis genera. Moreover, ME has been suggested to be carcinogenic [471] and alternatives are required that also do not exert effects on non-target species. Finally, the available lures are generally effective in attracting one sex, and do not have a species-specific action.
Although food-based attractants [472] are still dominant in population monitoring of the olive fruit fly, the use of the female pheromone can provide additional information on both the population density of wild populations and the age structure of wild populations. More recently, Sarles and colleagues identified two lactones released exclusively by males of R. completa and used lactone-baited traps in walnut orchards [473]. These traps have been found particularly effective for R. completa monitoring and allowed its earlier detection in the season, supporting the idea that the analysis of pheromone components may be particularly promising for trapping tephritids in the field.
6.2. Eradication and Suppression of Fruit Fly Populations Employing Semiochemicals
Male lures such as trimedlure and especially methyl-eugenol (ME), besides being employed for population monitoring, have been used for more direct control purposes. Male annihilation technique (MAT) (i.e., elimination of males, mating and oviposition of fertile eggs, based on strong male-specific lures that are deployed in a mass trapping or lure and kill approach) is considered as a very successful option to eliminate low populations of B. dorsalis and a tool that may drive invaded population to extirpation or even eradication. For example, male annihilation against B. dorsalis has been used as a main tool in attempts to eradicate incursions or isolated established populations in California, Hawaii and Florida [474,475,476,477,478], South Africa [479,480], the Marianas Islands in Micronesia [481], the Okinawa Islands in Japan [482], and Mauritius [483]. Eradication efforts using the male attractant CL against other Bactrocera species such as B. frauenfeldi were not successful [484]. Male attraction to CL remained consistent until advanced age in B. tryoni, although it sharply declined after 12 weeks of age, with potential implications for pest management [485]. Despite its broad use as a population monitoring tool, trimedlure is not considered as an eradication tool against medfly.
Male lures and the MAT have been considered in suppression programs often in combination with other methods and preferably in the frame of an Area Wide application strategy. As it was demonstrated in Hawaii, combination of MAT with field sanitation, protein bait, sterile male releases and biological control resulted in satisfactory reduction of the fruit fly population [486]. Methyl eugenol and CL for Bactrocera species, as well as trimedlure for the medfly, have been considered. Interestingly, the use of ME for the suppression of B. dorsalis in Southern Ethiopia proved to be successful [487].
The use of the female pheromone alone or in combination with other baits has been evaluated for the control of the olive fruit fly [488,489]. These efforts include a male annihilation component but also tools against females.
Apparently, classical mating disruption approaches involving saturation of target area with species-specific pheromones are not effective against fruit flies, and the MAT is prevailing. Indeed, mating disruption approaches against B. oleae in Spain and Greece led to inconclusive results [490,491]. However, a more recent study, in which authors observed a decrease in fly catches in the presence of high pheromone (i.e., 1,7-dioxaspiro[5.5]undecane) concentrations, supports the applicability of a mating disruption approach against this species [492]. For the medfly, for both trimedlure and the iodinated trimedlure analogue Ceralure, no mating disruption effects have been so far described [492]. Such an absence of disruption effect in this species was explained by the lack of saturation in response to higher trimedlure concentrations [492].
Alternative strategies aimed at interfering with mating deserve to be explored, including the potential applications of CHs as disruptors. CHs are known to play a role in the mating behaviour of different Drosophila species [493,494,495]. In D. suzukii, alteration of C23 alkanes ratios results in disrupting mate recognition and, as a consequence, courtship and mating behaviour [496]. The identification of OBP candidates potentially able to transport the B. dorsalis female-biased CH 4-DMP [425,426] is a promising step in the clarification of the functional role of CHs in the mating behaviour of fruit flies.
Moreover, in order to improve the efficacy of any semiochemical-based approach for fruit fly eradication and suppression, additional studies should be devoted to better understand the role of abiotic factors on trapping. Indeed, although temperature, humidity, rainfall and other exogenous abiotic factors do affect the temporal and spatial activity of fruit flies, data about the role of such factors in tephritid captures are still limited and mostly available for liquid protein-baited traps (see [269] for an overview). In the case of semiochemicals, it is known that medfly attractiveness to trimedlure is related to the release rate of this compound, which is, in turn, dependent on temperature [497,498]. Recently, Cameron and colleagues examined the vapour pressures and thermodynamic properties of seven attractants (i.e., RK, CL, raspberry ketone trifluoroacetate-RKTA, ME, methyl isoeugenol, dihydroeugenol, and zingerone) currently used for trapping Bactrocera, Dacus and Zeugodacus species [499]. The authors provide valuable data regarding the volatility of these attractants. In particular, they found that (Z)-methyl isoeugenol is the most volatile of the ME-type compounds, while RKTA is the most volatile among the RK-type compounds. Interestingly though, the field life of RKTA is not long due to its susceptibility to humidity [500]. Expanding our understanding of the features of these chemicals as well as of the identity/impact of multiple abiotic factors that may affect their activity is essential to determine the design of the eradication and suppression programs, as well as the location and density of the traps to be used.
6.3. Push and Pull Approaches Based on Repellent Semiochemicals
HMPs have been regarded as attractive tools for tephritid pest control since the ‘70s. In 1976, Katsoyannos and Boller proposed to use HMPs to prevent fruit fly oviposition into the fruit. They performed the first field experiment spraying raw HMP extract obtained from the faecal matter of R. cerasi. In this way, they achieved over 90% reduction of R. cerasi infestation in cherries orchards [501,502]. Later, once the chemical structure and the synthesis of R. cerasi HMP have been obtained [29], Aluja and Boller [31] tested the synthetic R. cerasi HMP in the field and, interestingly, this was the first application of a “push-pull strategy” in fruit flies. A push-pull strategy exploits a combination of behaviour-modifying stimuli to manipulate the distribution and abundance of the insect targets. Pests are repelled from their resource (push) by using stimuli that mask the host or that acts as repellents. Simultaneously, they are driven away from the resource (pull), by using highly attractive stimuli such as traps, facilitating their elimination. A reduction of the infestation of about 90% in cherry plants was achieved by treating one half of tree canopies with a synthetic HMP. The repelled females were then trapped with visual traps placed on the other half of the canopy [31,503]. The efficacy of the synthetic HMP was further supported by another field trial in which the infestation by cherry fruit flies was eliminated [504].
Similarly, field tests were performed using raw pheromone extract from the medfly’s faecal matter achieving 84% decrease in infestation in sprayed coffee plants [390]. Exploiting the cross-recognition observed in the Anastrepha genus, Aluja and co-workers tested three potential oviposition deterrents for A. obliqua in tropical plum and mango orchards. In their experiments, they used A. ludens faecal extracts and two fully synthetic simplified analogues of the naturally occurring pheromone, namely desmethyl A. ludens HMP (DM-HMP) and anastrephamide. They obtained a significant reduction in fruit damage rates with all substances tested, and interestingly, the simplified analogues displayed comparable levels of efficacy to the natural HMP [32].
The good efficacy of the synthetic HMP analogues found in both R. cerasi and A. obliqua field tests is promising in view of their potential use as pest control strategy. However, there is evidence that, after prolonged exposure to the HMP, flies can lay eggs in the treated fruits. This behaviour could be associated with the sensorial adaptation by the insect [123,503].
Although the use of HMP in the management of fruit flies was initially suggested as a push-pull system [32,112,393,400,505], the push-pull strategy is not suitable for the species with high population growth rates [506]. A recent work used anastrephamide in combination with a protein bait to reduce grapefruit infestation by A. ludens [507]. The authors found that anastrephamide can push flies out of the treated tree, but the push-pull system requires a more effective attractant.
Lastly, an alternative use of HMP has been proposed, which implies HMP application in commercial crops in which the fruit fly populations are not resident. This option allows achieving pest suppression because of the small population and lower risk for the occurrence of adaptation [114].
6.4. Implications for Biological Control
The recent findings that fruit fly parasitoids such as F. arisanus responded to HMP of fruit fly species [114,508] may pave the way for the development of methods and approaches to enhance the biological control efforts against fruit flies. Intensifying the research towards understanding the intraspecific interactions among parasitoids and fruit fly semiochemicals, in particular, oviposition-induced volatiles, is required to further increase parasitoid ability to localise and parasitise tephritid pests in the field [509]. In particular, expanding our understanding of the identity of HMPs will facilitate approaches based on the manipulation of parasitoid behaviour to benefit fruit fly control. Indeed, the incorporation of the identified HMP chemicals in the mass rearing process of the target parasitoid has the potential to facilitate the associate learning process that allows the parasitoid to distinguish HMPs from plant-emitted volatiles [114,510]. Moreover, treatments of fields and orchards with synthetic HMPs may not only deter fruit fly oviposition, but also attract parasitoids [511,512]. Among the factors mediating semiochemicals’ production and perception by tephritid and/or parasitoids, the insect microbiota likely plays a key role. Interestingly, a recent study showed that the production of β-caryophyllene emitted by host plants and mediating oviposition avoidance in egg-infested fruit by B. dorsalis is induced by egg-surface bacteria such as Providencia sp. and Klebsiella sp. [513]. Achieving a deeper understanding of the interplay between tephritid, their microbiota and host plants will shed new light on the multifaceted field of trans-kingdom communication [514] and will provide novel targets to be exploited for pest management.
6.5. Semiochemical-Based Tools to Enhance the Sterile Insect Technique
The identification and functional characterisation of tephritid semiochemicals may be beneficial for SIT applications and can also favour the integration of SIT and MAT to maximise the efficacy of pest management campaigns.
In SIT programs, sterilised males are released in the field to mate with wild females and induce sterility and hence no viable egg production [515]. Typically, sterile males are release as immature adults, which have to survive until sexual maturation, localise females and achieve copulation for SIT programs to be successful [516]. Thus, survival, dispersal capacity and mating competitiveness, for which male signalling is essential [329,517], are key factors that have to be fulfilled. Mating-enhancing semiochemicals have been widely described in several tephritid species and include both plant-derived compounds and synthetic chemicals, such as ME, Cue-Lure, RK, ginger root oil (GRO), citrus and guava fruit volatiles, and manuka oil (see [518,519] for reviews, and [458]).
SIT campaigns are more effective when pest population density is reduced before the release of sterile males. MAT applications, as well as inundative releases of biological control agents, have the potential to reduce the density of feral population size and hence to precede the implementation of the SIT. In Bactrocera species, the pre-release exposure of males to plant-derived semiochemicals and synthetic lures has been shown to reduce their subsequent response to attractants used in MAT [35,520]. Recently, the use of a diet containing RK fed to immature sterile B. tryoni prior to release resulted in increased subsequent survival and reduced response to MAT [521]. These results are particularly promising for the implementation of SIT-MAT simultaneous used in the field, to both increase control effectiveness and reduce operational costs.
6.6. Artificial Olfaction and Pheromone-Based Nanosensors
The ability of insect to detect olfactory stimuli at low concentrations over long distances [522] stimulated researchers to exploit these phenomena to develop biosensors based on insect behaviour [523], using isolated antennae [524] to detect explosives [525,526], food toxins [527], and for disease control and diagnosis [528,529]. Biosensors require a biological and artificial component, able to make the signals readable. OBPs can be expressed and purified easily and are stable to perturbations in temperature, pH and proteases [530]. Thus, they are considered ideal candidates to be exploited in biosensors development and were used to engineer systems able to detect floral odorants, alcohols and explosives in Drosophila and Apis mellifera L. (Hymenoptera: Apidae) [531,532,533,534]. More recently, also a member of the Tephritidae family was target of this type of research: an OBP from B. dorsalis, BdorOBP2, was expressed, purified and immobilised on an interdigitated electrode and it was shown to work as an efficient biosensor for chemicals emitted by host plants (e.g., isoamyl acetate, β-ionone, benzaldehyde) [535]. As previously described, given their higher specificity and sensitivity ORs are ideal candidates to be explored for the development of biosensors. Research in the field of OR-based bioelectronic sensors is indeed recently emerging as a simpler strategy with respect to the use of the mammalian ORs to detect environmentally significant volatile organic compounds (VOCs), as shown in mosquitoes (Diptera: Culicidae) and Drosophila [536,537,538]. Although interesting, this type of application is still far from being used in the field for tephritid management, also because only a few insect ORs have been deorphanised. Conversely, the exploitation of pheromone components for the development of innovative strategies to monitor early infestations is emerging as a powerful alternative to currently adopted strategies. Indeed, by targeting the major olive fruit fly volatile pheromone component, 1,7-dioxaspiro[5.5]undecane [319], Moitra and colleagues developed a β-cyclodextrinylated nanosensor specific to the female volatile pheromone of B. oleae [539]. This device is currently being tested in open field conditions and may be important not only for the control of B. oleae but also for the development of similar sensitive microelectromechanical system (MEMS) devices for other tephritid pests.
6.7. Cuticular Hydrocarbons as a Tool for Chemical Taxonomy
Cuticular lipid profiles, which are species-specific both in solitary and social insects, serve as fingerprints, making it possible to discriminate species taxonomically or to recognise sibling species [443]. The first successful use of CHs for the taxonomic discrimination of tephritid fruit fly species was reported 20 years ago in five articles on the hydrocarbon profile identification of adult Malaysian B. dorsalis complex flies [420] and larvae of Anastrepha (A. acris, A. fraterculus, A. suspensa, A. ludens, A. obliqua), Ceratitis (C. capitata, C. rosa), Bactrocera (B. dorsalis), and Z. cucurbitae [414]. Recently, Vaníčková and co-workers reported CH profiling is an efficient tool for the resolution of entities in the African fruit fly cryptic species FAR complex [274,289,414,416,417,418]. In these studies, twelve potential chemotaxonomic markers were identified for the distinction of adult male and female flies of C. fasciventris, C. rosa, C. anonae and C. capitata. Some of the geographically distinct subspecies hidden in the A. fraterculus complex can also be identified using their specific CH profiles. For example, Peruvian and Brazilian-1 morphotypes have unique CH profiles, suggesting CHs could be used to distinguish between these two subspecies [93]. In the B. dorsalis complex, clear segregation of complex cuticle profiles of both B. carambolae sexes from B. dorsalis was documented, supporting both taxonomic synonymisation of B. invadens, B. papayae, and B. philippinensis with B. dorsalis, as well as the exclusion of B. carambolae from B. dorsalis [421].
7. Conclusions and Challenges
Research in the last decades amazingly expanded our knowledge in the field of tephritid fruit fly semiochemical-based communication at the genomic, molecular, physiological and behavioural level, as outlined by the huge amount of available literature. On the other side, it is clear that semiochemical-based communication has not been characterised to an equal extent in all relevant tephritid pests. Moreover, most studies have been focused on volatile pheromones, with far less information available for CHs and HMPs. Thus, a major research need is to expand our knowledge to achieve an exhaustive understanding of all the semiochemical-based communication modalities in the target species. In addition, many specific questions remain to be answered. In the case of volatile pheromones, the application of different techniques and conditions for sampling, as well as the chosen source (i.e., rectal gland content or headspace), often provided different results in the detection and quantification of volatiles, making comparisons among identified sets of chemicals challenging. This urges for the parallel adoption of more than one method to ensure a comprehensive analysis of volatile pheromones. Moreover, future research on volatile pheromones will be essential to clarify (i) the identity of the molecular/genomic machinery underlying rectal (and salivary) gland physiology leading to volatile pheromone production, (ii) the relative impact of genomic background, evolutionary history and feeding preference in shaping the volatile pheromone bouquet. With respect to HMP research, it remains to be determined: (i) whether HMP is produced by specific glands, and how it is produced, (ii) which genes and pathways are involved in its synthesis, (iii) which is the chemical composition of most HMPs; although their widespread presence in tephritids, these pheromones were so far characterised in very few species; thus, expanding their characterisation represents a major challenge; (iv) whether HMPs are really absent in the Bactrocera genus. Although CHs have been widely described and novel studies are continuously being published, we still do not know which is their exact function in true fruit fly mating. Providing an answer to these and to several other questions that may arise when diving into this multifaceted field will be essential for the implementation of novel/improved approaches to tephritid pest control. Indeed, a series of knowledge gaps do exist, which limit the toolbox in the field. First of all, strategies to improve the formulation of semiochemical-based lures currently used in field applications are needed. These include the optimisation of liquid and solid dispensers containing individual compounds or mixtures (including isomers). Moreover, no male lures are currently available for Anastrepha and Rhagoletis spp., and some Bactrocera and Ceratitis species do not respond to the existing substances. Alternative and species-specific attractants are needed and the example of the lactone-baited traps used to capture R. completa is stimulating further studies aiming at exploring volatile pheromones for trapping. In addition to the MAT-based approaches, other strategies able to interfere with mating need to be explored, including the potential applications of CHs as disruptors. Overall, expanding our understanding of the identity of HMPs will facilitate the integration of multiple approaches for fruit fly control, including biological control, with special reference to the programs relying on the use of parasitoids. Finally, novel technologies allowing the production of nanosensors able to specifically detect pheromone components may open new routes for tephritid pest control.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/insects12050408/s1, Table S1: Chemicals detected in the volatile pheromone (headspace) of 18 tephritid species of high agricultural relevance. Table S2: List of unique and overlapping chemicals characterised in the volatile pheromone of tephritid species. Table S3: Chemicals with a HMP role detected in the adult female faecal matter of four tephritid species of high agricultural relevance. Table S4: Chemicals detected in the cuticular hexane body washes of 19 tephritid species of high agricultural relevance.
Author Contributions
Conceptualization: F.S., L.V. and F.V. Literature analysis: F.S., L.V., F.V., G.B. and N.T.P. Writing of the original draft preparation: F.S., L.V. and F.V. Writing, review and editing: F.S., L.V., F.V., G.B. and N.T.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Acknowledgments
This study was performed within the Food and Agriculture Organization/International Atomic Energy Agency (FAO/IAEA) Coordinated Research Project “Assessment of Simultaneous Application of SIT and MAT to Enhance Bactrocera Fruit Fly Management” (D41027), contract no. 23126/R0 (to L.V.). The authors thank Marc De Meyer and Mark Schutze for their constructive comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations and acronyms are used in this manuscript:
| AW-IPM | Area-Wide Integrated Pest Management |
| CCE | Conventional Chemical Ecology |
| CHs | Cuticular Hydrocarbons |
| CI | Chemical Ionization |
| CL | Cue Lure |
| CRCE | Computational Reverse Chemical Ecology |
| CSD | Current Source Density |
| CSPs | Chemosensory Proteins |
| DESI | Desorption Electrospray Ionization |
| DM-HMP | Desmethyl Host-Marking Pheromone |
| DVB/CAR/PDMS | Divinylbenzene/Carboxen/Polydimethylsiloxane |
| EAG | Electroantennogram |
| EI | Electron Ionization |
| ELSD | Evaporative Light Scattering Detectors |
| FAR | Ceratitis complex including C. fasciventris, C. anonae, C. rosa, and C. quilici |
| GA | Glutamic Acid |
| GC-EAD | Gas Chromatography-Electroantennographic Detection |
| GC×GC-TOFMS | Two-dimensional Gas Chromatography with Time-Of-Flight Mass Spectrometry |
| GC-FID | Gas Chromatography-Flame Ionization Detection |
| GC-FTIR | Gas Chromatography-Fourier Transformed Infrared Spectroscopy |
| GC-MS | Gas Chromatography-Mass Spectrometry |
| GOBP | General OBP |
| GPCRs | G Protein-Coupled Receptors |
| GRO | Ginger Root Oil |
| GRs | Gustatory Receptors |
| GSH | Tripeptide Glutathione |
| HMP | Host-Marking Pheromone |
| HPLC | High Performance Liquid Chromatography |
| IPM | Integrated Pest Management |
| IRs | Ionotrophic Receptors |
| LC-MS | Liquid Chromatography-Mass Spectrometry |
| LC-QTOF-MS | LC-Quadrupole Time-Of-Flight-Mass Spectrometry |
| MALDI-TOFMS | Matrix-Assisted Laser Desorption/Ionization-Time-Of-Flight Mass Spectrometry |
| MAT | Male Annihilation Technique |
| ME | Methyl Eugenol |
| MEMS | Microelectromechanical System |
| MSI | Mass Spectrometric Imaging |
| NMR | Nuclear Magnetic Resonance |
| OBPs | Odorant Binding Proteins |
| Orco | Olfactory receptor co-receptor |
| ORs | Olfactory Receptors |
| OSNs | Olfactory Sensory Neurons |
| PDMS/DVB | Polydimethylsiloxane/Divinylbenzene |
| RCE | Reverse Chemical Ecology |
| RK | Raspberry Ketone |
| RKTA | Raspberry Ketone Trifluoroacetate |
| RNAi | RNA Interference |
| SIT | Sterile Insect Technique |
| SPME | Solid Phase Microextraction |
| SSR | Single Sensillum Recording |
| TLC | Thin Layer Chromatography |
| UV-LDI o-TOFMS | Ultraviolet Laser Desorption Ionization orthogonal Time-Of-Flight Mass Spectrometry |
| VOCs | Volatile Organic Compounds |
| 1-NPN | N-phenylnaphthalen-1-amine |
| 4-DMP | 4-allyl-2,6-dimethoxyphenol |
References
- Brezolin, A.N.; Martinazzo, J.; Muenchen, D.K.; de Cezaro, A.M.; Rigo, A.A.; Steffens, C.; Steffens, J.; Blassioli-Moraes, M.C.; Borges, M. Tools for detecting insect semiochemicals: A review. Anal. Bioanal. Chem. 2018, 410, 4091–4108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinsworth, E.F. Regulation of pheromones and other semiochemicals in the United States. In Behavior-Modifying Chemicals for Insect Management: Applications of Pheromones and Other Attractants; Ridgway, R.L., Silverstein, R.M., Inscoe, M.N., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1990; pp. 569–603. [Google Scholar]
- Rodriguez, L.C.; Niemeyer, H.M. Integrated pest management, semiochemicals and microbial pest-control agents in Latin American agriculture. Crop Prot. 2005, 24, 615–623. [Google Scholar] [CrossRef]
- McNeil, J.N.; Millar, J.G.; Chapman, R.F. Chemical communication: Pheromones and allelochemicals. In The Insects: Structure and Function; Simpson, S.J., Douglas, A.E., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 857–900. [Google Scholar]
- Abd El-Ghany, N.M. Semiochemicals for controlling insect pests. J. Plant Prot. Res. 2019, 59, 1–11. [Google Scholar] [CrossRef]
- Ruther, J.; Meiners, T.; Steidle, J.L.M. Rich in phenomena-lacking in terms. A classification of kairomones. Chemoecology 2002, 12, 161–167. [Google Scholar] [CrossRef]
- Schulz, S. The Chemistry of Pheromones and Other Semiochemicals I; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Joseph, R.M.; Carlson, J.R. Drosophila chemoreceptors: A molecular interface between the chemical world and the brain. Trends Genet. 2015, 31, 683–695. [Google Scholar] [CrossRef] [Green Version]
- Tumlinson, J.H.; Teal, P.E.A. Relationship of structure and function to biochemistry in insect pheromone systems. In Pheromone Biochemistry; Prestwich, G.D., Blomquist, G.J., Eds.; Academic Press: New York, NY, USA, 1987; pp. 3–26. [Google Scholar]
- Gut, L.J.; Stelinski, L.L.; Thomson, D.R.; Miller, J.R. Behaviour-modifying chemicals: Prospects and constraints in IPM. In Integrated Pest Management: Potential, Constraints, and Challenges; Koul, O., Dhaliwal, G.S., Cuperus, G.W., Eds.; CABI Publishing: Cambridge, MA, USA, 2004; pp. 73–120. [Google Scholar]
- Wyatt, T.D. Pheromones and Animal Behavior; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Karlson, P.; Luscher, M. “Pheromones”: A new term for a class of biologically active substances. Nature 1959, 183, 55–56. [Google Scholar] [CrossRef]
- Butenandt, V.A.; Beckmann, R.; Stamm, D.; Hecker, E. Über den sexual-lockstoff des seidenspinners Bombyx mori—Reindarstellung und konstitution. Z. Naturforsch. 1959, 14, 283–284. [Google Scholar]
- Levine, J.D.; Millar, J.G. Chemical signalling: Laser on the fly reveals a new male-specific pheromone. Curr. Biol. 2009, 19, 653–655. [Google Scholar] [CrossRef] [Green Version]
- Roelofs, W.L. Chemistry of sex attraction. Proc. Natl. Acad. Sci. USA 1995. [Google Scholar] [CrossRef] [Green Version]
- Kaissling, K.E.; Kasang, G.; Bestmann, H.J.; Stransky, W.; Vostrowsky, O. A new pheromone of the silkworm moth Bombyx mori. Sci. Nat. 1978, 65, 382–384. [Google Scholar] [CrossRef]
- Kaissling, K.-E. Pheromone reception in insects. In Neurobiology of Chemical Communication; Mucignat-Caretta, C., Ed.; CRC Press/Taylor Francis: Boca Raton, FL, USA, 2014. [Google Scholar]
- Lebreton, S.; Borrero-Echeverry, F.; Gonzalez, F.; Solum, M.; Wallin, E.A.; Hedenstrom, E.; Hansson, B.S.; Gustavsson, A.L.; Bengtsson, M.; Birgersson, G.; et al. A Drosophila female pheromone elicits species-specific long-range attraction via an olfactory channel with dual specificity for sex and food. BMC Biol. 2017, 15, 88. [Google Scholar] [CrossRef] [Green Version]
- Pankiw, T. Cued in: Honey bee pheromones as information flow and collective decision-making. Apidologie 2004, 35, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Yew, J.Y.; Chung, H. Insect pheromones: An overview of function, form, and discovery. Prog. Lipid Res. 2015, 59, 88–105. [Google Scholar] [CrossRef]
- Greenfield, M.D. Signalers and Receivers: Mechanisms and Evolution of Arthropod Communication; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Ferveur, J.F. Cuticular hydrocarbons: Their evolution and roles in Drosophila pheromonal communication. Behav. Genet. 2005, 35, 279–295. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Jackson, L.L. Chemistry and biochemistry of insect waxes. Prog. Lipid Res. 1979, 17, 319–345. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Bagnères, A.G. Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780511711909. [Google Scholar]
- Vaníčková, L.; Canale, A.; Benelli, G. Sexual chemoecology of mosquitoes (Diptera, Culicidae): Current knowledge and implications for vector control programs. Parasitol. Int. 2017, 66, 190–195. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Howard, R.W. Pheromone biosynthesis in social insects. In Insect Pheromone Biochemistry and Molecular Biology; Blomquist, G.J., Vogt, R.G., Eds.; Elsevier: New York, NY, USA, 2003; pp. 323–340. [Google Scholar]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- Nufio, C.R.; Papaj, D.R. Host marking behavior in phytophagous insects and parasitoids. Entomol. Exp. Appl. 2001, 99, 273–293. [Google Scholar] [CrossRef] [Green Version]
- Hurter, J.; Boller, E.F.; Städler, E.; Blattmann, B.; Buser, H.R.; Bosshard, N.U.; Damm, L.; Kozlowski, M.W.; Schöni, R.; Raschdorf, F.; et al. Oviposition-deterring pheromone in Rhagoletis cerasi L.: Purification and determination of the chemical constitution. Experientia 1987, 43, 157–164. [Google Scholar] [CrossRef]
- Aluja, M.; Boller, E.F. Host marking pheromone of Rhagoletis cerasi: Field deployment of synthetic pheromone as a novel cherry fruit fly management strategy. Entomol. Exp. Appl. 1992, 65, 141–147. [Google Scholar] [CrossRef]
- Aluja, M.; Díaz-Fleischer, F.; Boller, E.F.; Hurter, J.; Edmunds, A.J.F.; Hagmann, L.; Patrian, B.; Reyes, J. Application of feces extracts and synthetic analogues of the host marking pheromone of Anastrepha ludens significantly reduces fruit infestation by A. obliqua in Tropical plum and mango backyard orchards. J. Econ. Entomol. 2009, 102, 2268–2278. [Google Scholar] [CrossRef] [PubMed]
- Cheseto, X.; Kachigamba, D.L.; Ekesi, S.; Ndung’u, M.; Teal, P.E.A.; Beck, J.J.; Torto, B. Identification of the ubiquitous antioxidant tripeptide glutathione as a fruit fly semiochemical. J. Agric. Food Chem. 2017, 65, 8560–8568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheseto, X.; Kachigamba, D.L.; Benderap, M.; Ekesi, S.; Ndung’u, M.; Beck, J.J.; Torto, B. Identification of glutamic acid as a host marking pheromone of the African fruit fly species Ceratitis rosa (Diptera: Tephritidae). J. Agric. Food Chem. 2018, 66, 9933–9941. [Google Scholar] [CrossRef]
- Benelli, G.; Daane, K.M.; Canale, A.; Niu, C.Y.; Messing, R.H.; Vargas, R.I. Sexual communication and related behaviours in Tephritidae: Current knowledge and potential applications for Integrated Pest Management. J. Pest Sci. 2014, 87, 385–405. [Google Scholar] [CrossRef]
- Roitberg, B.D.; Prokopy, R.J. Insects that mark host plants. Bioscience 1987, 37, 400–406. [Google Scholar] [CrossRef]
- Hoffmeister, T.S.; Roitberg, B.D. Counterespionage in an insect herbivore-parasitoid system. Naturwissenschaften 1997, 84, 117–119. [Google Scholar] [CrossRef]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Doorenweerd, C.; Leblanc, L.; Norrbom, A.L.; Jose, M.S.; Rubinoff, D. A global checklist of the 932 fruit fly species in the tribe Dacini (Diptera, Tephritidae). Zookeys 2018, 730, 19–56. [Google Scholar] [CrossRef] [Green Version]
- Roskov, Y.; Ower, G.; Orrell, T.; Nicolson, D.; Bailly, N.; Kirk, P.M.; Bourgoin, T.; DeWalt, R.E.; Decock, W.; van Nieukerken, E.; et al. Species 2000 ITIS Catalogue of Life, 2019 Annual Checklist 2019; Naturalis: Leiden, The Netherlands, 2019. [Google Scholar]
- Malavasi, A. Introductory remarks. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T.E., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R.I., Eds.; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Clarke, A.R. Biology and Management of Bactrocera and Related Fruit Flies; CABI: Wallingford, UK, 2019. [Google Scholar]
- Nugnes, F.; Russo, E.; Viggiani, G.; Bernardo, U. First record of an invasive fruit fly belonging to Bactrocera dorsalis complex (Diptera: Tephritidae) in Europe. Insects 2018, 9, 182. [Google Scholar] [CrossRef] [Green Version]
- Malacrida, A.R.; Gomulski, L.M.; Bonizzoni, M.; Bertin, S.; Gasperi, G.; Guglielmino, C.R. Globalization and fruitfly invasion and expansion: The medfly paradigm. Genetica 2007, 131, 1–9. [Google Scholar] [CrossRef]
- Papadopoulos, N.T. Fruit Fly Invasion: Historical, Biological, Economic Aspects and Management. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Springer: Dordrecht, The Netherlands, 2014; pp. 219–252. ISBN 9789401791939. [Google Scholar]
- Vreysen, M.J.B.; Robinson, A.S.; Hendrichs, J. Area-Wide Control of Insect Pests from Research to Field Implementation; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Sivinski, J.; Aluja, M.; Dodson, G.N.; Freidberg, A.; Headrick, D.H.; Kaneshiro, K.Y.; Landolt, P.J. Topics in the evolution of fruit fly mating behavior. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 751–792. [Google Scholar]
- Feron, M. L’instinct de reproduction chez la mouche méditerranéenne des fruits Ceratitis capitata Wied. (Dipt. Trypetidae). Comportement sexuel. Comportement de ponte. Rev. Pathol. Végétale d’Entomol. Agric. Fr. 1962, 41, 1–129. [Google Scholar]
- Ekanayake, W.M.T.D.; Clarke, A.R.; Schutze, M.K. Close-distance courtship of laboratory reared Bactrocera tryoni (Diptera: Tephritidae). Austral. Entomol. 2019, 58, 578–588. [Google Scholar] [CrossRef]
- Benelli, G.; Canale, A.; Bonsignori, G.; Ragni, G.; Stefanini, C.; Raspi, A. Male wing vibration in the mating behavior of the olive fruit fly Bactrocera oleae (Rossi) (Diptera: Tephritidae). J. Insect Behav. 2012, 25, 590–603. [Google Scholar] [CrossRef]
- Poramarcom, R. Sexual communication in the Oriental fruit fly, Dacus dorsalis Hendel (Diptera: Tephritidae). Ph.D. Thesis, University of Hawaii, Honolulu, HI, USA, 1988. [Google Scholar]
- Benelli, G.; Giunti, G.; Canale, A.; Messing, R.H. Lek dynamics and cues evoking mating behavior in tephritid flies infesting soft fruits: Implications for behavior-based control tools. Appl. Entomol. Zool. 2014, 49, 363–373. [Google Scholar] [CrossRef]
- Bradbury, J.W. The evolution of leks. In Natural Selection and Social Behaviour; Alexander, R.D., Tinkle, D.W., Eds.; Chiron Press: New York, NY, USA, 1981; pp. 138–169. [Google Scholar]
- Shelly, T.E. Sexual selection on leks: A fruit fly primer. J. Insect Sci. 2018, 18. [Google Scholar] [CrossRef]
- Iwahashi, O.; Majima, T. Lek formation and male-male competition in the melon fly, Dacus cucurbitae Coquillett (Diptera, Tephritidae). Appl. Entomol. Zool. 1986, 21, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Mir, S.H.; Mir, G.M. Lekking behaviour and male-male rivalry in the melon fly Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). J. Insect Behav. 2016, 29, 379–384. [Google Scholar] [CrossRef]
- Raghu, S.; Clarke, A.R. Spatial and temporal partitioning of behaviour by adult dacines: Direct evidence for methyl eugenol as a mate rendezvous cue for Bactrocera cacuminata. Physiol. Entomol. 2003, 28, 175–184. [Google Scholar] [CrossRef]
- Messina, F.J.; Subler, J.K. Conspecific and heterospecific interactions of male Rhagoletis flies (Diptera, Tephritidae) on a shared host. J. Kansas Entomol. Soc. 1995, 68, 206–213. [Google Scholar]
- Prokopy, R.J.; Bennett, E.W.; Bush, G.L. Mating behavior in Rhagoletis pomonella (Diptera: Tephritidae): II. Temporal organization. Can. Entomol. 1972, 104, 97–104. [Google Scholar] [CrossRef]
- Smith, D.C.; Prokopy, R.J. Mating behavior of Rhagoletis mendax (Diptera: Tephritidae) flies in nature. Ann. Entomol. Soc. Am. 1982, 75, 388–392. [Google Scholar] [CrossRef]
- Buda, V.; Blazyte-Cereskiene, L.; Radziute, S.; Apsegaite, V.; Stamm, P.; Schulz, S.; Aleknavicius, D.; Mozuraitis, R. Male-produced (-)-delta-heptalactone, pheromone of fruit fly Rhagoletis batava (Diptera: Tephritidae), a sea buckthorn berries pest. Insects 2020, 11, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, R.; Shelly, T.E.; Whittier, T.S.; Kaneshiro, K.Y. Alpha-copaene, a potential rendezvous cue for the mediterranean fruit fly, Ceratitis capitata? J. Chem. Ecol. 2000, 26, 87–100. [Google Scholar] [CrossRef]
- Shelly, T.; Dang, C.; Kennelly, S. Exposure to orange (Citrus sinensis L.) trees, fruit, and oil enhances mating success of male Mediterranean fruit flies (Ceratitis capitata [Wiedemann]). J. Insect Behav. 2004, 17, 303–315. [Google Scholar] [CrossRef]
- Warthen, J.D.; McInnis, D.O. Isolation and identification of male medfly attractive components in Litchi chinensis stems and Ficus spp. stem exudates. J. Chem. Ecol. 1989, 15, 1931–1946. [Google Scholar] [CrossRef] [PubMed]
- Flath, R.A.; Cunningham, R.T.; Mon, T.R.; John, J.O. Additional male Mediterranean fruit fly (Ceratitis capitata Wied) attractants from Angelica seed oil (Angelica archangelica L). J. Chem. Ecol. 1994, 20, 1969–1984. [Google Scholar] [CrossRef]
- Flath, R.A.; Cunningham, R.T.; Mon, T.R.; John, J.O. Male lures for Mediterranean fruit fly (Ceratitis capitata Wied.): Structural analogs of α-copaene. J. Chem. Ecol. 1994, 20, 2595–2609. [Google Scholar] [CrossRef]
- Niogret, J.; Gill, M.A.; Espinoza, H.R.; Kendra, P.E. Attraction and electroantennogram responses of male Mediterranean fruit fly (Diptera: Tephritidae) to six plant essential oils. J. Entomol. Zool. Stud. 2017, 5, 958–964. [Google Scholar]
- Nishida, R.; Tan, K.H.; Serit, M.; Lajis, N.H.; Sukari, A.M.; Takahashi, S.; Fukami, H. Accumulation of phenylpropanoids in the rectal glands of males of the Oriental fruit fly, Dacus dorsalis. Experientia 1988, 44, 534–536. [Google Scholar] [CrossRef]
- Tan, K.H.; Nishida, R. Sex pheromone and mating competition after methyl eugenol consumption in Bactrocera dorsalis complex. In Fruit Fly Pests—A World Assessment of Their Biology and Management; McPheron, B.A., Steck, G.J., Eds.; St. Lucie Press: Delray Beach, FL, USA, 1996; pp. 147–153. [Google Scholar]
- Hee, A.K.W.; Tan, K.H. Attraction of female and male Bactrocera papayae to conspecific males fed with methyl eugenol and attraction of females to male sex pheromone components. J. Chem. Ecol. 1998, 24, 753–764. [Google Scholar] [CrossRef]
- Khoo, C.C.-H.; Tan, K.-H. Attraction of both sexes of melon fly, Bactrocera cucurbitae to conspecific males—A comparison after pharmacophagy of cue-lure and a new attractant—zingerone. Entomol. Exp. Appl. 2000, 97, 317–320. [Google Scholar] [CrossRef]
- Wee, S.L.; Tan, K.H. Female sexual response to male rectal volatile constituents in the fruit fly, Bactrocera carambolae (Diptera: Tephritidae). Appl. Entomol. Zool. 2005, 40, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Wee, S.L.; Tan, K.H.; Nishida, R. Pharmacophagy of methyl eugenol by males enhances sexual selection of Bactrocera carambolae (Diptera: Tephritidae). J. Chem. Ecol. 2007, 33, 1272–1282. [Google Scholar] [CrossRef]
- Kobayashi, R.M.; Ohinata, K.; Chambers, D.L.; Fujimoto, M.S. Sex pheromones of the Oriental fruit fly and the melon fly: Mating behavior, bioassay method, and attraction of females by live males and by suspected pheromone glands of males. Environ. Entomol. 1978, 7, 107–112. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Hooper, G.H.S.; Bateman, M.A. Economic fruit flies of the South Pacific Region, 2nd ed.; Queensland Department of Primary Industries: Brisbane, Australia, 1982. [Google Scholar]
- Noushini, S.; Perez, J.; Park, S.J.; Holgate, D.; Jamie, I.; Jamie, J.; Taylor, P. Rectal gland chemistry, volatile emissions, and antennal responses of male and female banana fruit fly, Bactrocera musae. Insects 2020, 11, 32. [Google Scholar] [CrossRef] [Green Version]
- Haniotakis, G.E. Sexual attraction in the olive fruit fly Dacus oleae (Gmelin). Environ. Entomol. 1974, 3, 82–86. [Google Scholar] [CrossRef]
- Haniotakis, G.E. Male olive fly attraction to virgin females in the field. Ann. Zool. Ecol. Anim. 1977, 9, 273–276. [Google Scholar]
- Baker, R.; Herbert, R.; Howse, P.E.; Jones, O.T.; Francke, W.; Reith, W. Identification and synthesis of the major sex pheromone of the olive fly (Dacus oleae). J. Chem. Soc. Chem. Commun. 1980, 1, 52. [Google Scholar] [CrossRef]
- Carpita, A.; Canale, A.; Raffaelli, A.; Saba, A.; Benelli, G.; Raspi, A. (Z)-9-tricosene identified in rectal gland extracts of Bactrocera oleae males: First evidence of a male-produced female attractant in olive fruit fly. Naturwissenschaften 2012, 99, 77–81. [Google Scholar] [CrossRef]
- De Marzo, L.; Nuzzaci, L.; Solinas, M. Studio anatomico, istologico, ultrastrutturale e fisiologico del retto ed osservazioni etologiche in relazione alla possibile produzione di feromoni sessuali nel maschio di Dacus oleae (Gmelin). Entomologica 1978, XIV, 203–266. [Google Scholar]
- Mavraganis, V.G.; Papadopoulos, N.T.; Kouloussis, N.A. Extract of olive fruit fly males (Diptera: Tephritidae) attract virgin females. Entomol. Hell. 2017, 19, 14. [Google Scholar] [CrossRef] [Green Version]
- Levi-Zada, A.; Nestel, D.; Fefer, D.; Nemni-Lavy, E.; Deloya-Kahane, I.; David, M. Analyzing diurnal and age-related pheromone emission of the olive fruit fly, Bactrocera oleae by sequential SPME-GCMS analysis. J. Chem. Ecol. 2012, 38, 1036–1041. [Google Scholar] [CrossRef]
- Benelli, G. Aggressive behavior and territoriality in the olive fruit fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae): Role of residence and time of day. J. Insect Behav. 2014, 27, 145–161. [Google Scholar] [CrossRef]
- Canale, A.; Benelli, G.; Germinara, G.S.; Fusini, G.; Romano, D.; Rapalini, F.; Desneux, N.; Rotundo, G.; Raspi, A.; Carpita, A. Behavioural and electrophysiological responses to overlooked female pheromone components in the olive fruit fly, Bactrocera oleae (Diptera: Tephritidae). Chemoecology 2015, 25, 147–157. [Google Scholar] [CrossRef]
- Eberhard, W.G. Sexual behavior and sexual selection in the Mediterranean fruit fly, Ceratitis capitata (Dacinae: Ceratitidini). In Fruit flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 459–489. [Google Scholar]
- Robacker, D.C.; Hart, W.G. Courtship and territoriality of laboratory-reared Mexican fruit-flies, Anastrepha ludens (Diptera, Tephritidae), in cages containing host and nonhost trees. Ann. Entomol. Soc. Am. 1985, 78, 488–494. [Google Scholar] [CrossRef]
- Silva, M.T.; Polloni, Y.J.; Bressan, S. Mating behavior of some fruit flies of the genus Anastrepha Schiner, 1868 (Diptera: Tephritidae) in the laboratory. Rev. Bras. Entomol. 1985, 29, 155–164. [Google Scholar]
- Tychsen, P.H. Mating behavior of the Queensland fruit fly, Dacus tryoni (Diptera: Tephritidae), in field cages. J. Aust. Entomol. Soc. 1977, 16, 459–465. [Google Scholar] [CrossRef]
- Vaníčková, L.; Svatos, A.; Kroiss, J.; Kaltenpoth, M.; Do Nascimento, R.R.; Hoskovec, M.; Břízová, R.; Kalinova, B. Cuticular hydrocarbons of the South American fruit fly Anastrepha fraterculus: Variability with sex and age. J. Chem. Ecol. 2012, 38, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Aluja, M.; Norrbom, A. Fruit flies (Tephritidae) Phylogeny and Evolution of Behavior; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Vaníčková, L.; Břízová, R.; Pompeiano, A.; Ekesi, S.; De Meyer, M. Cuticular hydrocarbons corroborate the distinction between lowland and highland Natal fruit fly (Tephritidae, Ceratitis rosa) populations. Zookeys 2015, 507–524. [Google Scholar] [CrossRef] [Green Version]
- Vaníčková, L.; Břízová, R.; Mendonca, A.L.; Pompeiano, A.; Do Nascimento, R.R. Intraspecific variation of cuticular hydrocarbon profiles in the Anastrepha fraterculus (Diptera: Tephritidae) species complex. J. Appl. Entomol. 2015, 139, 679–689. [Google Scholar] [CrossRef]
- Lewis, S.M.; Vahed, K.; Koene, J.M.; Engqvist, L.; Bussière, L.F.; Perry, J.C.; Gwynne, D.; Lehmann, G.U.C. Emerging issues in the evolution of animal nuptial gifts. Biol. Lett. 2014, 10, 20140336. [Google Scholar] [CrossRef]
- Vahed, K. The function of nuptial feeding in insects: A review of empirical studies. Biol. Rev. 2007, 73, 43–78. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D. Does indirect mating trophallaxis boost male mating success and female egg load in Mediterranean fruit flies? J. Pest Sci. 2018, 91, 181–188. [Google Scholar] [CrossRef]
- Aluja, M.; Jácome, I.; Birke, A.; Lozada, N.; Quintero, G. Basic patterns of behavior in wild Anastrepha striata (Diptera: Tephritidae) flies under field-cage conditions. Ann. Entomol. Soc. Am. 1993, 86, 776–793. [Google Scholar] [CrossRef]
- Perez-Staples, D.; Aluja, M. Anastrepha striata (Diptera: Tephritidae) females that mate with virgin males live longer. Ann. Entomol. Soc. Am. 2004, 97, 1336–1341. [Google Scholar] [CrossRef] [Green Version]
- Guillen, L.; Pascacio-Villafan, C.; Stoffolano, J.G.; Lopez-Sanchez, L.; Velazquez, O.; Rosas-Saito, G.; Altuzar-Molina, A.; Ramirez, M.; Aluja, M. Structural differences in the digestive tract between females and males could modulate regurgitation behavior in Anastrepha ludens (Diptera: Tephritidae). J. Insect Sci. 2019, 19. [Google Scholar] [CrossRef]
- Sivinski, J.M.; Epsky, N.; Heath, R.R. Pheromone deposition on leaf territories by male Caribbean fruit flies, Anastrepha suspensa (loew) (Diptera: Tephritidae). J. Insect Behav. 1994, 7, 43–51. [Google Scholar] [CrossRef]
- Papaj, D.R.; Garcia, J.M.; Alonso-Pimentel, H. Marking of host fruit by male Rhagoletis boycei Cresson flies (Diptera: Tephritidae) and its effect on egg-laying. J. Insect Behav. 1996, 9, 585–598. [Google Scholar] [CrossRef]
- Jang, E.B. Effects of mating and accessory-gland injections on olfactory-mediated behavior in the female Mediterranean fruit fly, Ceratitis capitata. J. Insect Physiol. 1995, 41, 705–710. [Google Scholar] [CrossRef]
- Gomulski, L.M.; Dimopoulos, G.; Xi, Z.Y.; Scolari, F.; Gabrieli, P.; Siciliano, P.; Clarke, A.R.; Malacrida, A.R.; Gasperi, G. Transcriptome profiling of sexual maturation and mating in the Mediterranean fruit fly, Ceratitis capitata. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.W.; Luo, D.Y.; Wu, F.Y.; Wang, J.L.; Zhang, H.Y. RNA sequencing to characterize transcriptional changes of sexual maturation and mating in the female oriental fruit fly Bactrocera dorsalis. BMC Genomics 2016, 17. [Google Scholar] [CrossRef] [Green Version]
- Kumaran, N.; van der Burg, C.A.; Qin, Y.J.; Cameron, S.L.; Clarke, A.R.; Prentis, P.J. Plant-mediated female transcriptomic changes post-mating in a tephritid fruit fly, Bactrocera tryoni. Genome Biol. Evol. 2018, 10, 94–107. [Google Scholar] [CrossRef] [Green Version]
- Campanini, E.B.; Congrains, C.; Torres, F.R.; de Brito, R.A. Odorant-binding proteins expression patterns in recently diverged species of Anastrepha fruit flies. Sci. Rep. 2017, 7, 2194. [Google Scholar] [CrossRef] [Green Version]
- Devescovi, F.; Hurtado, J.; Taylor, P.W. Mating-induced changes in responses of female Queensland fruit fly to male pheromones and fruit: A mechanism for mating-induced sexual inhibition. J. Insect Physiol. 2021, 129, 104195. [Google Scholar] [CrossRef]
- Averill, A.L.; Prokopy, R.J. Intraspecific competition in the tephritid fruit fly Rhagoletis pomonella. Ecology 1987, 68, 878–886. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Roitberg, B.D. Fruit fly foraging behavior. In Fruits Flies, Their Biology, Natural Enemies and Control; Robinson, A.S., Hooper, G., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 293–306. [Google Scholar]
- Fletcher, B.S.; Prokopy, R.J. Host location and oviposition in tephritid fruit flies. In Reproductive Behaviour of Insects: Individuals and Populations; Bailey, W.J., Ridsdill-Smith, J., Eds.; Chapman Hall: London, UK, 1991; pp. 141–171. [Google Scholar]
- Prokopy, R.J.; Ziegler, J.R.; Wong, T.T.Y. Deterrence of repeated oviposition by fruit-marking pheromone in Ceratitis capitata (Diptera: Tephritidae). J. Chem. Ecol. 1978, 4, 55–63. [Google Scholar] [CrossRef]
- Prokopy, R.J. Evidence for a marking pheromone deterring repeated oviposition in apple maggot flies. Environ. Entomol. 1972, 1, 326–332. [Google Scholar] [CrossRef]
- Díaz-Fleischer, F.; Papaj, D.R.; Prokopy, R.J.; Norrbom, A.L.; Aluja, M. Evolution of fruit fly oviposition behavior. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 811–841. [Google Scholar]
- Silva, M.A.; Bezerra-Silva, G.C.D.; Mastrangelo, T. The host marking pheromone application on the management of fruit flies—A Review. Braz. Arch. Biol. Technol. 2012, 55, 835–842. [Google Scholar] [CrossRef] [Green Version]
- Leal, T.A.B.S.; Zucoloto, F.S. Selection of artificial hosts for oviposition by wild Anastrepha obliqua (Macquart) (Diptera, Tephritidae): Influence of adult food and effect of experience. Rev. Bras. Entomol. 2008, 52, 467–471. [Google Scholar] [CrossRef]
- Crnjar, R.M.; Prokopy, R.J. Morphological and electrophysiological mapping of tarsal chemoreptors of oviposition deterring pheromone in Rhagoletis pomonella flies. J. Insect Physiol. 1982, 28, 393–400. [Google Scholar] [CrossRef]
- Zhang, G.N.; Hu, F.; Dou, W.; Wang, J.J. Morphology and distribution of sensilla on tarsi and ovipositors of six fruit flies (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2012, 105, 319–327. [Google Scholar] [CrossRef]
- Eisemann, C.H.; Rice, M.J. Behavioural evidence for hygro- and mechanoreception by ovipositor sensilla of Dacus tryoni (Diptera: Tephritidae). Physiol. Entomol. 1989, 14, 273–277. [Google Scholar] [CrossRef]
- Liscia, A.; Crnjar, R.; Angioy, A.M.; Pietra, P.; Stoffolano, J.G.J. I chemosensilli dell’ovopositore in Tabanus nigrovittatus (Macq.), Chrysops fuliginosus (Wied.), e Rhagoletis pomonella (Walsh). Boll. Soc. Ital. Biol. Sper. 1982, 58, 1325–1329. [Google Scholar]
- Girolami, V.; Crnjar, R.; Angioy, A.M.; Strapazzon, A.; Pietra, P.; Stoffolano, J.G.J.; Prokopy, R.J. Behavior and sensory physiology of Rhagoletis pomonella in relation to oviposition stimulants and deterrents in fruit. In Fruitflies of Economic Importance; Cavalloro, R., Ed.; Balkema Pub: Rotterdam, The Netherlands; Boston, MA, USA, 1986; pp. 183–190. [Google Scholar]
- Stoffolano, J.G.; Yin, L.R.S. Structure and function of the ovipositor and associated sensilla of the apple maggot, Rhagoletis pomonella (Walsh) (Diptera, Tephritidae). Int. J. Insect Morphol. Embryol. 1987, 16, 41–69. [Google Scholar] [CrossRef]
- Norrbom, A.L.; Kim, K.C. Revision of the schausi group of Anastrepha Schiner (Diptera: Tephritidae), with a discussion of the terminology of the female terminalia in the Tephritoidea. Ann. Entomol. Soc. Am. 1988, 81, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Papaj, D.R.; Aluja, M. Temporal dynamics of host-marking in the tropical tephritid fly, Anastrepha ludens. Physiol. Entomol. 1993, 18, 279–284. [Google Scholar] [CrossRef]
- Papaj, D.R.; Roitberg, B.D.; Opp, S.B.; Aluja, M.; Prokopy, R.J.; Wong, T.T.Y. Effect of marking pheromone on clutch size in the Mediterranean fruit-fly. Physiol. Entomol. 1990, 15, 463–468. [Google Scholar] [CrossRef]
- Papaj, D.R.; Roitberg, B.D.; Opp, S.B. Serial effects of host infestation on egg allocation by the Mediterranean fruit-fly—A rule of thumb and its functional-significance. J. Anim. Ecol. 1989, 58, 955–970. [Google Scholar] [CrossRef]
- Papaj, D.R.; Averill, A.L.; Prokopy, R.J.; Wong, T.T.Y. Host-marking pheromone and use of previously established oviposition sites by the Mediterranean fruit-fly (Diptera, Tephritidae). J. Insect Behav. 1992, 5, 583–598. [Google Scholar] [CrossRef]
- Roitberg, B.D.; van Lenteren, J.C.; van Alphen, J.M.M.; Galis, F.; Prokopy, R.J. Foraging of Rhagoletis pomonella, a parasite of hawthorn (Crataegus), in nature. J. Anim. Ecol. 1982, 48, 307–325. [Google Scholar] [CrossRef]
- Roitberg, B.D.; Cairl, R.S.; Prokopy, R.J. Oviposition deterring pheromone influences dispersal distance in tephritid fruit flies. Entomol. Exp. Appl. 1984, 35, 217–220. [Google Scholar] [CrossRef]
- Nufio, C.R.; Papaj, D.R. Host-marking behaviour as a quantitative signal of competition in the walnut fly Rhagoletis juglandis. Ecol. Entomol. 2004, 29, 336–344. [Google Scholar] [CrossRef]
- Averill, A.L.; Prokopy, R.J. Oviposition-deterring fruit marking pheromone in Rhagoletis basiola. Florida Entomol. 1981, 64, 221–226. [Google Scholar] [CrossRef]
- Averill, A.L.; Prokopy, R.J. Oviposition-deterring fruit marking pheromone in Rhagoletis zephyria. J. Georg. Entomol. Soc. 1982, 17, 315–319. [Google Scholar]
- Aluja, M.; Diaz-Fleischer, F. Foraging behavior of Anastrepha ludens, A. obliqua, and A. serpentina in response to feces extracts containing host marking pheromone. J. Chem. Ecol. 2006, 32, 367–389. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Papaj, D.R. Behavior of flies of the genera Rhagoletis, Zonosemata, and Carpomya. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 219–252. [Google Scholar]
- Papaj, D.R. Oviposition site guarding by male walnut flies and its possible consequences for mating success. Behav. Ecol. Sociobiol. 1994, 34, 187–195. [Google Scholar] [CrossRef]
- Cirio, U.; Italiana, S.E. Osservazioni sul comportamento di ovideposizione della Rhagoletis completa Cresson (Diptera, Trypetidae) in laboratorio. In Proceedings of the Atti del IX Congresso Nazionale Italiano di Entomologia, Tipografia Bertelly Picardi, Siena, Italy, 21–25 June 1972; Volume 1, pp. 99–117. [Google Scholar]
- Papaj, D.R. Use and avoidance of occupied hosts as a dynamic process in tephritid flies. In Insect-Plant Interactions; Bernays, E.A., Ed.; CRC Press: Boca Raton, FL, USA, 1993; Volume 5, pp. 25–46. [Google Scholar]
- Lalonde, R.G.; Mangel, M. Seasonal effects on superparasitism by Rhagoletis completa. J. Anim. Ecol. 1994, 63, 583–588. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Green, T.A.; Olson, W.A.; Vargas, R.F.; Kaneshia, D.; Wong, T.Y. Discrimination by Dacus dorsalis females (Diptera: Tephritidae) against larval-infested fruit. Florida Entomol. 1989, 72, 319–323. [Google Scholar] [CrossRef]
- Fitt, G.P. Oviposition behaviour of two tephritid fruit flies, Dacus tryoni and Dacus jarvisi, as influenced by the presence of larvae in the host fruit. Oecologia 1984, 62, 37–46. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Koyama, J. Oviposition site partitioning in Dacus cucurbitae. Entomol. Exp. Appl. 1982, 31, 428–432. [Google Scholar] [CrossRef]
- Cirio, U. Reperti sul meccanismo stimolo-risposta nell’ovideposizione del Dacus oleae (Gmelin) (Diptera: Trypetidae). Redia 1971, 52, 10. [Google Scholar]
- Girolami, V.; Vianello, A.; Strapazzon, A.; Ragazzi, E.; Veronese, G. Ovipositional deterrents in Dacus oleae. Entomol. Exp. Appl. 1981, 29, 177–188. [Google Scholar] [CrossRef]
- Lo Scalzo, R.; Scarpati, M.L.; Verzegnassi, B.; Vita, G. Olea europaea chemicals repellent to Dacus oleae females. J. Chem. Ecol. 1994, 20, 1813–1823. [Google Scholar] [CrossRef]
- Arita, L.H.; Kaneshiro, K.Y. Structure and function of the rectal epithelium and anal glands during mating behavior in the Mediterranean fruit fly. Proc. Hawaiian Entomol. Soc. 1986, 26, 27–30. [Google Scholar]
- Quilici, S.; Franck, A.; Peppuy, A.; Correia, E.D.R.; Mouniama, C.; Blard, F. Comparative studies of courtship behavior of Ceratitis spp. (Diptera: Tephritidae) in Reunion island. Florida Entomol. 2002, 85, 138–142. [Google Scholar] [CrossRef]
- Fletcher, B.S. Storage and release of a sex pheromone by the Queensland fruit fly, Dacus tryoni (Diptera: Trypetidae). Nature 1968, 219, 631–632. [Google Scholar] [CrossRef]
- Kuba, H.; Sokei, Y. The production of pheromone clouds by spraying in the melon fly, Dacus cucurbitae Coquillett (Diptera, Tephritidae). J. Ethol. 1988, 6, 105–110. [Google Scholar] [CrossRef]
- Poramarcom, R.; Baimai, V. Sexual behavior and signals used for mating of Bactrocera correcta. In Fruit Fly Pests: A World Assessment of Their Biology and Management; McPheron, B.A., Steck, G.J., Eds.; St. Lucie Press: Delray Beach, FL, USA, 1996; pp. 51–58. [Google Scholar]
- Nation, J.L. Biology of pheromone release by male Caribbean fruit-flies, Anastrepha suspensa (Diptera, Tephritidae). J. Chem. Ecol. 1990, 16, 553–572. [Google Scholar] [CrossRef]
- Nation, J.L. The role of pheromones in the mating system of Anastrepha fruit flies. In Fruit Flies: Their Biology, Natural Enemies and Control; Robinson, A.S., Hooper, G., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1989; Volume 16, pp. 189–205. [Google Scholar]
- Nation, J.L. Courtship behavior and evidence for a sex attractant in the male Caribbean fruit fly, Anastrepha suspensa. Ann. Entomol. Soc. Am. 1972, 65, 1364–1367. [Google Scholar] [CrossRef]
- Yip, E.C.; Mikó, I.; Ulmer, J.M.; Cherim, N.A.; Townley, M.A.; Poltak, S.; Helms, A.M.; De Moraes, C.M.; Mescher, M.C.; Tooker, J.F. Giant polyploid epidermal cells and male pheromone production in the tephritid fruit fly Eurosta solidaginis (Diptera: Tephritidae). J. Insect Physiol. 2021, 130, 104210. [Google Scholar] [CrossRef]
- Caetano, F.H.; Solferini, V.N.; de Britto, F.B.; Lins, D.S.; Aluani, T.; de Brito, V.G.; Zara, F.J. Ultra morphology of the digestive system of Anastrepha fraterculus and Ceratitis capitata (Diptera Tephritidae). Braz. J. Morphol. Sci. 2006, 23, 455–462. [Google Scholar]
- Barros, M.D.; Malavasi, A. Morphology of adult male rectum of seven species of Anastrepha from Brazil and mating behavior correlations. In Fruit Fly Pests—A World Assessment of Their Biology and Management; McPheron, B.A., Steck, G.J., Eds.; Taylor Francis Inc.: Abingdon, UK, 1996. [Google Scholar]
- Lee, L.W.Y.; Chang, T.H. Morphology of sex-pheromone gland in male oriental fruit fly and its suspected mechanism of pheromone release. In Proceedings of the Second International Symposium, 16–21 September; Economopoulos, A.P., Ed.; Symposium Organizing Committee: Kolymbari, Crete, Greece, 1986; pp. 16–21. [Google Scholar]
- Economopoulos, A.P.; Gianakakis, A.; Tzanakakis, M.E.; Voyatzoglou, A. Reproductive behaviour and physiology of the olive fruit fly. Anatomy of the adult rectum and odours emitted by adults. Ann. Entomol. Soc. Am. 1971, 64, 1112–1116. [Google Scholar] [CrossRef]
- Khoo, C.C.H.; Tan, K.H. Rectal gland of Bactrocera papayae: Ultrastructure, anatomy and sequestration of auto fluorescent compounds upon methyl eugenol consumption by the male fly. Microsc. Res. Tech. 2005, 67, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Arita, L.H. The Mating Behavior of the Mediterranean Fruit Fly, Ceratitis capitata (Wiedemann). Ph.D. Thesis, University of Hawaii, Honolulu, HI, USA, 1983. [Google Scholar]
- Gomez Cendra, P.; Calcagno, G.; Belluscio, L.; Vilardi, J.C. Male courtship behavior of the South American fruit fly, Anastrepha fraterculus, from an Argentinean laboratory strain. J. Insect Sci. 2011, 11, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nation, J.L. Sex-specific glands in tephritid fruit flies of the genera Anastrepha, Ceratitis, Dacus and Rhagoletis (Diptera: Tephritidae). Int. J. Insect Morphol. Embryol. 1981, 4, 27–30. [Google Scholar] [CrossRef]
- Llosie, J.; Roche, A. Organes odoriferants des males de Ceratitis capitata. Bull. Soc. Entomol. Fr. 1960, 65, 206–209. [Google Scholar]
- Fletcher, B.S. The structure and the function of the sex pheromone glands of the male Queensland fruit fly, Dacus tryoni. J. Insect Physiol. 1969, 15, 1309–1322. [Google Scholar] [CrossRef]
- Teles, M.C. Structure and development of specific sex glands in males of some Brazilian fruit flies of the genus Anastrepha Schiner, 1868 (Diptera: Tephritidae). In Proceedings of the CEC/IOBC International Symposium on Fruit Flies of Economic Importance 87, Rome, Italy, 7–10 April 1987; pp. 179–189. [Google Scholar]
- Lima, I.S.; Howse, P.E.; Stevens, D.R. Volatile components from the salivary glands of calling males of the south American fruit fly, Anastrepha fraterculus: Partial identification and behavioural activity. In Fruit Fly Pests. A World Assessment of Their Biology and Management; MacPheron, B.A., Steck, G.J., Eds.; St. Lucie Press: Delray Beach, FL, USA, 1996; pp. 107–113. [Google Scholar]
- Nation, J.L. The structure and development of two sex specific glands in male Caribbean fruit flies. Ann. Entomol. Soc. Am. 1974, 67, 731–734. [Google Scholar] [CrossRef]
- Goncalves, G.B.; Silva, C.E.; Dos Santos, J.C.G.; Dos Santos, E.S.; Do Nascimento, R.R.; Da Silva, E.L.; Mendonca, A.D.L.; De Freitas, M.D.; Sant’Ana, A.E.G. Comparison of the volatile components released by calling males of Ceratitis capitata (Diptera: Tephritidae) with those extractable from the salivary glands. Florida Entomol. 2006, 89, 375–379. [Google Scholar] [CrossRef]
- Heath, R.R.; Landolt, P.J.; Robacker, D.C.; Dueben, B.D.; Epsky, N.D. Sexual pheromones of tephritid flies: Clues to unravel phylogeny and behavior. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 793–809. [Google Scholar]
- Nishida, R.; Tan, K.H.; Takahashi, S.; Fukami, H. Volatile components of male rectal glands of the melon fly, Dacus cucurbitae Coquillett (Diptera, Tephritidae). Appl. Entomol. Zool. 1990, 25, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Nishida, R. Ecological significance of male fruit fly attractants. In Proceedings of the International Symposium Biology Control Fruit Flies, University of Ryukyus, Ginowan, Japan; Kawasaki, K., Iwahashi, O., Kaneshiro, K.Y., Eds.; 1991; pp. 246–254. [Google Scholar]
- Tan, K.H.; Nishida, R. Methyl eugenol: Its occurrence, distribution, and role in nature, especially in relation to insect behavior and pollination. J. Insect Sci. 2012, 12, 1–74. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.H.; Nishida, R.; Jang, E.B.; Shelly, T.E. Pheromones, Male Lures, and Trapping of Tephritid Fruit Flies. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T.E., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R., Eds.; Springer: New York, NY, USA, 2014; Volume 2, pp. 15–74. [Google Scholar]
- Shelly, T.E.; Dewire, A.L.M. Chemically mediated mating success in male Oriental fruit-flies (Diptera, Tephritidae). Ann. Entomol. Soc. Am. 1994, 87, 375–382. [Google Scholar] [CrossRef]
- Light, D.M.; Jang, E.B.; Binder, R.G.; Flath, R.A.; Kint, S. Minor and intermediate components enhance attraction of female Mediterranean fruit flies to natural male odor pheromone and its synthetic major components. J. Chem. Ecol. 1999, 25, 2757–2777. [Google Scholar] [CrossRef]
- Mazomenos, B.E.; Haniotakis, G.E. A multicomponent female sex pheromone of Dacus oleae Gmelin: Isolation and bioassay. J. Chem. Ecol. 1981, 7, 437–444. [Google Scholar] [CrossRef]
- Milet-Pinheiro, P.; Navarro, D.M.A.; De Aquino, N.C.; Ferreira, L.L.; Tavares, R.F.; da Silva, R.D.C.; Lima-Mendonca, A.; Vanickova, L.; Mendonca, A.L.; Do Nascimento, R.R. Identification of male-borne attractants in Anastrepha fraterculus (Diptera: Tephritidae). Chemoecology 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Caceres, C.; Segura, D.F.; Vera, M.T.; Wornoayporn, V.; Cladera, J.L.; Teal, P.; Sapountzis, P.; Bourtzis, K.; Zacharopoulou, A.; Robinson, A.S. Incipient speciation revealed in Anastrepha fraterculus (Diptera; Tephritidae) by studies on mating compatibility, sex pheromones, hybridization, and cytology. Biol. J. Linn. Soc. 2009, 97, 152–165. [Google Scholar] [CrossRef]
- Břízová, R.; Mendonca, A.L.; Vaníčková, L.; Mendonca, A.L.; Da Silva, C.E.; Tomcala, A.; Paranhos, B.A.J.; Dias, V.S.; Joachim-Bravo, I.S.; Hoskovec, M.; et al. Pheromone analyses of the Anastrepha fraterculus (Diptera: Tephritidae) cryptic species complex. Florida Entomol. 2013, 96, 1107–1115. [Google Scholar] [CrossRef] [Green Version]
- Lima, I.S.; House, P.E.; do Nascimento, R.R. Volatile substances from male Anastrepha fraterculus Wied. (Diptera: Tephritidae): Identification and behavioural activity. J. Braz. Chem. Soc. 2001, 12, 196–201. [Google Scholar] [CrossRef]
- Juárez, M.L.; Devescovi, F.; Břízová, R.; Bachmann, G.; Segura, D.F.; Kalinová, B.; Fernández, P.; Ruiz, M.J.; Yang, J.; Teal, P.E.A.; et al. Evaluating mating compatibility within fruit fly cryptic species complexes and the potential role of sex pheromones in pre-mating isolation. Zookeys 2015, 540, 125–155. [Google Scholar] [CrossRef] [Green Version]
- Rocca, J.R.; Nation, J.L.; Strekowski, L.; Battiste, M.A. Comparison of volatiles emitted by male Caribbean and Mexican fruit-flies. J. Chem. Ecol. 1992, 18, 223–244. [Google Scholar] [CrossRef]
- Robacker, D.C. Behavioral responses of female Mexican fruit flies, Anastrepha ludens, to components of male-produced sex pheromone. J. Chem. Ecol. 1988, 14, 1715–1726. [Google Scholar] [CrossRef]
- Robacker, D.C.; Hart, W.G. (Z)-3-nonenol, (Z,Z)-3,6-nonandienol and (S,S)-(-)-epianastrephin: Male produced pheromone of the Mexican fruit fly. Entomol. Exp. Appl. 1985, 39, 103–108. [Google Scholar] [CrossRef]
- Esponda-Gaxiola, R.E. Contribución al Estudio Quimico Del Atrayente Sexual de la Mosca Mexicana de la Fruta, Anastrepha ludens (Loew). Bachelor’s Thesis, Monterrey Institute of Technology and Higher Studies, Monterrey, Mexico, 1977. [Google Scholar]
- Stokes, J.B.; Uebel, E.C.; Warthen, J.D., Jr.; Jacobson, M.; Flippen-Anderson, J.L.; Gilardi, R.; Spishakoff, L.M.; Wilzer, K.R. Isolation and identification of novel lactones from male Mexican fruit flies. J. Agric. Food Chem. 1983, 31, 1162–1167. [Google Scholar] [CrossRef]
- Battiste, M.A.; Strekowski, L.; Vanderbilt, D.P.; Visnick, M.; King, R.W.; Nation, J.L. Anastrephin and epianastrephin, novel lactone components isolated from the sex pheromone blend of male Caribbean and Mexican fruit flies. Tetrahedron Lett. 1983, 24, 2611–2614. [Google Scholar] [CrossRef]
- Bosa, C.F.; Cruz-López, L.; Zepeda-Cisneros, C.S.; Valle-Mora, J.; Guillén-Navarro, K.; Liedo, P. Sexual behavior and male volatile compounds in wild and mass-reared strains of the Mexican fruit fly Anastrepha ludens (Loew) (Diptera: Tephritidae) held under different colony management regimes. Insect Sci. 2016, 23, 105–116. [Google Scholar] [CrossRef]
- Gonçalves, G.B.; Silva, C.E.; Mendonça, A.D.L.; Vaníčková, L.; Tomčala, A.; Do Nascimento, R.R. Pheromone communication in Anastrepha obliqua (Diptera: Tephritidae): A comparison of the volatiles and salivary gland extracts of two wild populations. Florida Entomol. 2013, 96, 1365–1374. [Google Scholar] [CrossRef]
- Lopez-Guillen, G.; Lopez, L.C.; Malo, E.A.; Rojas, J.C. Olfactory responses of Anastrepha obliqua (Diptera: Tephritidae) to volatiles emitted by calling males. Florida Entomol. 2011, 94, 874–881. [Google Scholar] [CrossRef]
- Dos Silva, C.S.; dos Melo, R.S.; Tavares, R.A.N.; de Aquino, N.C.; de Tavares, R.F.; Vanícková, L.; de Mendonça, A.L.; Daza, N.A.C.; Nascimento, R.R.; dos Melo, R.S.; et al. do Estudo comparativo do feromonio sexual de duas populacoes sul Americanas de Anastrepha obliqua. In A Diversidade de Debates na Pesquisa em Química; Atena Editora: Porto Alegre, Brazil, 2019. [Google Scholar]
- López-Guillén, G.; Cruz-López, L.; Malo, E.A.; González-Hernández, H.; Cazares, C.L.; López-Collado, J.; Toledo, J.; Rojas, J.C. Factors influencing the release of volatiles in Anastrepha obliqua males (Diptera: Tephritidae). Environ. Entomol. 2008, 37, 876–882. [Google Scholar] [CrossRef]
- Meza-Hernandez, J.S.; Hernandez, E.; Salvador-Figueroa, M.; Cruz-Lopez, L. Sexual compatibility, mating performance and sex pheromone release of mass-reared and wild Anastrepha obliqua (Diptera: Tephritidae) under field-cage conditions. In Proceedings of the International Fruit Fly Symposium, Stellenbosch, South Africa, 6–10 May 2002; pp. 99–104. [Google Scholar]
- Ibanez-Lopez, A.; Cruz-Lopez, L. Glandulas salivales de Anastrepha obliqua (Macquart) (Diptera: Tephritidae): Analisis quimico y morfologico, y actividad biologica de los componentes volatiles. Folia Entomol. Mex. 2001, 40, 221–231. [Google Scholar]
- Robacker, D.C.; Aluja, M.; Cosse, A.A.; Sacchetti, P. Sex pheromone investigation of Anastrepha serpentina (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2009, 102, 560–566. [Google Scholar] [CrossRef]
- Lima-Mendonça, A.; de Mendonça, A.L.; Sant’Ana, A.E.G.; Nascimento, R.R. Do Semioquímicos de moscas das frutas do gênero Anastrepha. Quim. Nova 2014, 37. [Google Scholar] [CrossRef]
- Nation, J.L. Sex pheromone components of Anastrepha suspensa and their role in mating behavior. In Proceedings of the International Symposium on the Biology and Control of Fruit Flies; Kawasaki, K., Iwahashi, O., Kaneshiro, K.Y., Eds.; University of Ryukyus: Okinawa, Ginowan, Japan, 1991; pp. 224–236. [Google Scholar]
- Ponce, W.P.; Nation, J.L.; Emmel, T.C.; Smittle, B.J.; Teal, P.E.A. Quantitative analysis of pheromone production in irradiated Caribbean fruit fly males, Anastrepha suspensa (Loew). J. Chem. Ecol. 1993, 19, 3045–3056. [Google Scholar] [CrossRef]
- Chuman, T.; Sivinski, J.; Heath, R.R.; Calkins, C.O.; Tumlinson, J.H.; Battiste, M.A.; Wydra, R.L.; Strekowski, L.; Nation, J.L. Suspensolide, a new macrolide component of male Caribbean fruit fly (Anastrepha suspensa [Loew]) volatiles. Tetrahedron Lett. 1988, 29, 6561–6564. [Google Scholar] [CrossRef]
- Robacker, D.C.; Hart, W.G. Electroantennograms of male and female Caribbean fruit-flies (Diptera, Tephritidae) elicited by chemicals produced by males. Ann. Entomol. Soc. Am. 1987, 80, 508–512. [Google Scholar] [CrossRef]
- Epsky, N.D.; Heath, R.R. Food availability and pheromone production by males of Anastrepha suspensa (Diptera, Tephritidae). Environ. Entomol. 1993, 22, 942–947. [Google Scholar] [CrossRef]
- Tumlinson, J.H. Contemporary frontiers in insect semiochemical research. J. Chem. Ecol. 1988, 14, 2109–2130. [Google Scholar] [CrossRef]
- Heath, R.R.; Manukian, A.; Epsky, N.D.; Sivinski, J.; Calkins, C.O.; Landolt, P.J. A bioassay system for collecting volatiles while simultaneously attracting tephritid fruit flies. J. Chem. Ecol. 1993. [Google Scholar] [CrossRef]
- Epsky, N.D.; Heath, R.R. Pheromone production by male Anastrepha suspensa (Diptera: Tephritidae) under natural light cycles in greenhouse studies. Environ. Entomol. 1993, 22, 464–469. [Google Scholar] [CrossRef]
- Nation, J.L. Sex pheromone of the Caribbean fruit fly: Chemistry and field ecology. In Natural Products; Elsevier: Amsterdam, The Netherlands, 1983; pp. 109–110. [Google Scholar]
- Baker, R.; Bacon, A. The identification of spiroacetals in the volatile secretions of two species of fruit fly (Dacus dorsalis, Dacus cucurbitae). Cell. Mol. Life Sci. 1985, 41, 1484–1485. [Google Scholar] [CrossRef]
- Ohinata, K.; Jacobson, M.; Kobayashi, R.M.; Chambers, D.L.; Fujimoto, M.S.; Higa, H.H. Oriental fruit fly and melon fly: Biological and chemical studies of smoke produced by males. J. Environ. Sci. Health Part A 1982, 17, 197–216. [Google Scholar] [CrossRef]
- Nishida, R.; Shelly, T.E.; Kaneshiro, K.Y.; Tan, K.-H. Roles of semiochemicals in mating systems: A comparison between oriental fruit fly and medfly. In Area Wide Control Fruit Flies Other Insect Pests; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Haniotakis, G.; Francke, W.; Mori, K.; Redlich, H.; Schurig, V. Sex-specific activity of (R)-(-)- and (S)- (+)-1,7-dioxaspiro[5.5]undecane, the major pheromone of Dacus oleae. J. Chem. Ecol. 1986, 12, 1559–1568. [Google Scholar] [CrossRef]
- Gariboldi, P.; Jommi, G.; Rossi, R.; Vita, G. Studies on the chemical constitution and sex pheromone activity of volatile substances emitted by Dacus oleae. Experientia 1982, 38, 441–444. [Google Scholar] [CrossRef]
- Bellas, T.E.; Fletcher, B.S. Identification of the major components in the secretion from the rectal pheromone glands of the Queensland fruit flies Dacus tryoni and Dacus neohumeralis (Diptera:Tephritidae). J. Chem. Ecol. 1979, 5, 795–803. [Google Scholar] [CrossRef]
- Booth, Y.K.; Schwartz, B.D.; Fletcher, M.T.; Lambert, L.K.; Kitching, W.; Voss, J.J. De A diverse suite of spiroacetals, including a novel branched representative, is released by female Bactrocera tryoni (Queensland fruit fly). Chem. Commun. 2006, 3975. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Venkatesham, U.; Unelius, C.R.; Sporle, A.; Pérez, J.; Taylor, P.W.; Suckling, D.M. Chemical composition of the rectal gland and volatiles released by female Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae). Environ. Entomol. 2019, 48, 807–814. [Google Scholar] [CrossRef]
- Tan, H.K.; Nishida, R. Incorporation of raspberry ketone in the rectal glands of males of the Queensland fruit fly, Bactrocera tryoni Frogatt (Diptera: Tephritidae). Appl. Entomol. Zool. 1995, 30, 494–497. [Google Scholar] [CrossRef] [Green Version]
- Levi-Zada, A.; Levy, A.; Rempoulakis, P.; Fefer, D.; Steiner, S.; Gazit, Y.; Nestel, D.; Yuval, B.; Byers, J.A. Diel rhythm of volatile emissions of males and females of the peach fruit fly Bactrocera zonata. J. Insect Physiol. 2020, 120. [Google Scholar] [CrossRef] [PubMed]
- Břízová, R.; Vaníčková, L.; Fatarova, M.; Ekesi, S.; Hoskovec, M.; Kalinova, B. Analyses of volatiles produced by the African fruit fly species complex (Diptera, Tephritidae). Zookeys 2015, 385–404. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, M.; Ohinata, K.; Chambers, D.L.; Jones, W.A.; Fujimoto, M.S. Insect sex attractants. Isolation, identification, and synthesis of sex pheromones of male Mediterranean fruit fly. J. Med. Chem. 1973, 13, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Ohinata, K.; Nakagawa, S.; Fujimoto, M.; Higa, H.; Jacobson, M. Mediterranean fruit fly: Laboratory and field evaluations of synthetic sex pheromones. J. Environ. Sci. Health Part A 1977. [Google Scholar] [CrossRef]
- Ohinata, K.; Jacobson, M.; Nakagawa, S.; Urago, T.; Fujimoto, M.; Higa, H. Methyl (E)-6-nonenoate: A new Mediterranean fruit fly male attractant. J. Econ. Entomol. 1979, 72, 648–650. [Google Scholar] [CrossRef]
- Jang, E.B.; Light, D.M.; Dickens, J.C.; McGovern, T.P.; Nagata, J.T. Electroantennogram responses of mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae) to trimedlure and its trans isomers. J. Chem. Ecol. 1989, 15, 2219–2231. [Google Scholar] [CrossRef] [PubMed]
- Flath, R.A.; Jang, E.B.; Light, D.M.; Mon, T.R.; Carvalho, L.; Binder, R.G.; John, J.O. Volatile pheromonal emissions from the male Mediterranean fruit fly—Effects of fly age and time of day. J. Agric. Food Chem. 1993, 41, 830–837. [Google Scholar] [CrossRef]
- Merli, D.; Mannucci, B.; Bassetti, F.; Corana, F.; Falchetto, M.; Malacrida, A.R.; Gasperi, G.; Scolari, F. Larval diet affects male pheromone blend in a laboratory strain of the medfly, Ceratitis capitata (Diptera: Tephritidae). J. Chem. Ecol. 2018, 44, 339–353. [Google Scholar] [CrossRef]
- Vaníčková, L.; do Nascimento, R.R.; Hoskovec, M.; Jezkova, Z.; Břízová, R.; Tomcala, A.; Kalinova, B. Are the wild and laboratory insect populations different in semiochemical emission? The case of the medfly sex pheromone. J. Agric. Food Chem. 2012, 60, 7168–7176. [Google Scholar] [CrossRef]
- Alfaro, C.; Vacas, S.; Zarzo, M.; Navarro-Llopis, V.; Primo, J. Solid phase microextraction of volatile emissions of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae): Influence of fly sex, age, and mating status. J. Agric. Food Chem. 2011, 59, 298–306. [Google Scholar] [CrossRef]
- Siciliano, P.; He, X.L.; Woodcock, C.; Pickett, J.A.; Field, L.M.; Birkett, M.A.; Kalinova, B.; Gomulski, L.M.; Scolari, F.; Gasperi, G.; et al. Identification of pheromone components and their binding affinity to the odorant binding protein CcapOBP83a-2 of the Mediterranean fruit fly, Ceratitis capitata. Insect Biochem. Mol. Biol. 2014, 48, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.; Herbert, R.H.; Grant, G.G. Isolation and identification of the sex pheromone of the Mediterranean fruit fly, Ceratitis capitata (Wied.). J. Chem. Soc. Chem. Commun. 1985, 12, 824–825. [Google Scholar] [CrossRef]
- Heath, R.R.; Landolt, P.J.; Tumlinson, J.H.; Chambers, D.L.; Murphy, R.E.; Doolittle, R.E.; Dueben, B.D.; Sivinski, J.; Calkins, C.O. Analysis, synthesis, formulation, and field testing of three major components of male mediterranean fruit fly pheromone. J. Chem. Ecol. 1991, 17, 1925–1940. [Google Scholar] [CrossRef]
- Raptopoulos, D.; Haniotakis, G.; Koutsaftikis, A.; Kelly, D.; Mavraganis, V. Biological activity of chemicals identified from extracts and volatiles of male Rhagoletis cerasi. J. Chem. Ecol. 1995, 21, 1287–1297. [Google Scholar] [CrossRef]
- De Lima, I.S.; Howse, P.E.; Stevens, I.D.R. Volatile components from the salivary glands of calling males of the South American fruit fly, Anastrepha fraterculus: Partial identification and behavioural activity. In Fruit Fly Pests; CRC Press: Boca Raton, FL, USA, 2020; pp. 107–113. [Google Scholar] [CrossRef] [Green Version]
- Robacker, D.C.; Garcia, J.A. Responses of laboratory-strain Mexican fruit flies, Anastrepha ludens, to combinations of fermenting fruit odor and male-produced pheromone in laboratory bioassays. J. Chem. Ecol. 1990, 16, 2027–2038. [Google Scholar] [CrossRef]
- Perkins, M.; Fletcher, M.; Kitching, W.; Drew, R.; Moore, C. Chemical studies of rectal gland secretions of some species of Bactrocera dorsalis complex of fruit flies (Diptera: Tephritidae). J. Chem. Ecol. 1990, 16, 2475–2487. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Jacobs, M.F.; Kitching, W.; Krohn, S.; Drew, R.A.I.; Haniotakis, G.E.; Francke, W. Absolute stereochemistry of the 1,7-dioxaspiro[5.5]undecanols in fruit-fly species, including the olive-fly. J. Chem. Soc. Chem. Commun. 1992, 1457. [Google Scholar] [CrossRef]
- Zhang, X.G.; Wei, C.M.; Miao, J.; Zhang, X.J.; Wei, B.; Dong, W.X.; Xiao, C. Chemical compounds from female and male rectal pheromone glands of the guava fruit fly, Bactrocera correcta. Insects 2019, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Tokushima, I.; Orankanok, W.; Tan, K.H.; Ono, H.; Nishida, R. Accumulation of phenylpropanoid and sesquiterpenoid volatiles in male rectal pheromonal glands of the guava fruit fly, Bactrocera correcta. J. Chem. Ecol. 2010, 36, 1327–1334. [Google Scholar] [CrossRef]
- Fletcher, M.T.; Wells, J.A.; Jacobs, M.F.; Krohn, S.; Kitching, W.; Drew, R.A.I.; Moore, C.J.; Francke, W. Chemistry of fruit-flies. Spiroacetal-rich secretions in several Bactrocera species from the South-West Pacific region. J. Chem. Soc. Perkin Trans. 1 1992, 2827–2831. [Google Scholar] [CrossRef]
- Kitching, W.; Lewis, J.A.; Perkins, M.V.; Drew, R.; Moore, C.J.; Schurig, V.; Konig, W.A.; Francke, W. Chemistry of fruit-flies—Composition of the rectal gland secretion of (male) Dacus cucumis (cucumber fly) and Dacus halfordiae—Characterization of (Z,Z)-2,8-Dimethyl-1,7-Dioxaspiro[5.5]Undecane. J. Org. Chem. 1989, 54, 3893–3902. [Google Scholar] [CrossRef]
- Mazomenos, B.E.; Pomonis, J.G. Male olive fruit fly pheromone: Isolation, identification and lab-bioassays. In Proceedings of the CEC/IOBC International Symposium on Fruit Flies of Economic Importance, Athens, Greece, 16 November 1982; pp. 96–103. [Google Scholar]
- Noushini, S.; Park, S.J.; Jamie, I.; Jamie, J.; Taylor, P. Sampling technique biases in the analysis of fruit fly volatiles: A case study of Queensland fruit fly. Sci. Rep. 2020, 10, 19799. [Google Scholar] [CrossRef]
- Perkins, M.V.; Kitching, W.; Drew, R.A.I.; Moore, C.J.; König, W.A. Chemistry of fruit flies: Composition of the male rectal gland secretions of some species of South-East Asian Dacinae. Re-examination of Dacus cucurbitae (melon fly). J. Chem. Soc., Perkin Trans. 1 1990, 1111–1117. [Google Scholar] [CrossRef]
- Baker, R.; Herbert, R.H.; Lomer, R.A. Chemical components of the rectal gland secretions of male Dacus cucurbitae, the melon fly. Experientia 1982, 38, 232–233. [Google Scholar] [CrossRef]
- Nishida, R.; Iwahashi, O.; Tan, K.H. Accumulation of Dendrobium superbum (orchidaceae) fragrance in the rectal glands by males of the melon fly, Dacus cucurbitae. J. Chem. Ecol. 1993, 19, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, C.S.; Papadopoulos, N.T.; Kouloussis, N.A.; Tananaki, C.I.; Katsoyannos, B.I. Essential oils of citrus fruit stimulate oviposition in the Mediterranean fruit fly Ceratitis capitata (Diptera: Tephritidae). Physiol. Entomol. 2012, 37, 330–339. [Google Scholar] [CrossRef]
- Cosse, A.A.; Todd, J.L.; Millar, J.G.; Martinez, L.A.; Baker, T.C. Electroantennographic and coupled gas chromatographic-electroantennographic responses of the mediterranean fruit fly, Ceratitis capitata, to male-produced volatiles and mango odor. J. Chem. Ecol. 1995, 21, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Nation, J.L. The sex pheromone blend of Caribbean fruit fly males: Isolation biological activity, and partial chemical characterization. J. Environ. Entomol. 1975, 4, 27–30. [Google Scholar] [CrossRef]
- De Aquino, N.C.; Ferreira, L.L.; Tavares, R.; Silva, C.S.; Mendonça, A.; Joachim-Bravo, I.S.; Milet-Pinheiro, P.; Navarro, D.; De Abreu Galdino, F.C.; Do Nascimento, R.R. Bioactive male-produced volatiles from Anastrepha obliqua and their role in attraction of conspecific females. J. Chem. Ecol. 2021, 47, 167–174. [Google Scholar] [CrossRef]
- Robacker, D.C.; Hart, W.G. Behavioral-responses of male and female Mexican fruit-flies, Anastrepha ludens, to male-produced chemicals in laboratory experiments. J. Chem. Ecol. 1986, 12, 39–47. [Google Scholar] [CrossRef]
- Aluja, M.; Cabagne, G.; Altuzar-Molina, A.; Pascacio-Villafan, C.; Enciso, E.; Guillen, L. Host plant and antibiotic effects on scent bouquet composition of Anastrepha ludens and Anastrepha obliqua calling males, two polyphagous tephritid pests. Insects 2020, 11, 309. [Google Scholar] [CrossRef]
- Teal, P.E.A.; Gomez-Simuta, Y.; Proveaux, A.T. Mating experience and juvenile hormone enhance sexual signaling and mating in male Caribbean fruit flies. Proc. Natl. Acad. Sci. USA 2000, 97, 3708–3712. [Google Scholar] [CrossRef]
- Aceves-Aparicio, E.; Pérez-Staples, D.; Arredondo, J.; Corona-Morales, A.; Morales-Mávil, J.; Díaz-Fleischer, F. Combined effects of methoprene and metformin on reproduction, longevity, and stress resistance in Anastrepha ludens (Diptera: Tephritidae): Implications for the Sterile Insect Technique. J. Econ. Entomol. 2021, 114, 142–151. [Google Scholar] [CrossRef]
- Landolt, P.J.; Averill, A.L. Fruit flies. In Pheromones of Non-Lepidopteran Insects Associated with Agricultural Plants; Hardie, J., Minks, A.K., Eds.; CABI Publication: Wallingford, UK, 1999; pp. 3–25. [Google Scholar]
- Farine, J.P.; Ferveur, J.F.; Everaerts, C. Volatile Drosophila cuticular pheromones are affected by social but not sexual experience. PLoS ONE 2012, 7, e40396. [Google Scholar] [CrossRef]
- Droney, D.C.; Hock, M.B. Male sexual signals and female choice in Drosophila grimshawi (Diptera: Drosophildae). J. Insect Behav. 1998, 11, 59–71. [Google Scholar] [CrossRef]
- Muller, M.; Buchbauer, G. Essential oil components as pheromones. A review. Flavour Fragr. J. 2011, 26, 357–377. [Google Scholar] [CrossRef]
- Engl, T.; Kaltenpoth, M. Influence of microbial symbionts on insect pheromones. Nat. Prod. Rep. 2018, 35, 386–397. [Google Scholar] [CrossRef]
- Henneken, J.; Goodger, J.Q.D.; Jones, T.M.; Elgar, M.A. Diet-mediated pheromones and signature mixtures can enforce signal reliability. Front. Ecol. Evol. 2017, 4. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Ma, Y.; Xie, M.; Lu, Y.; Cheng, D. Rectal bacteria produce sex pheromones in the male oriental fruit fly. Curr. Biol. 2021, 31. [Google Scholar] [CrossRef]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R package for the visualization of intersecting sets and their properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [Green Version]
- Barbosa-Cornelio, R.; Cantor, F.; Coy-Barrera, E.; Rodriguez, D. Tools in the investigation of volatile semiochemicals on insects: From sampling to statistical analysis. Insects 2019, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Leal, W.S. Reverse chemical ecology at the service of conservation biology. Proc. Natl. Acad. Sci. USA 2017, 114, 12094–12096. [Google Scholar] [CrossRef] [Green Version]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Torto, B. Chemical signals as attractants, repellents and aggregation stimulants. In Chemical Ecology. Encyclopedia of Life Support Systems; EOLSS Publishers Co. Ltd: Oxford, UK, 2009. [Google Scholar]
- Al-Khshemawee, H.; Du, X.; Agarwal, M.; Yang, J.O.; Ren, Y.L. Application of Direct Immersion Solid-Phase Microextraction (DI-SPME) for understanding biological changes of Mediterranean Fruit Fly (Ceratitis capitata) during mating procedures. Molecules 2018, 23, 2951. [Google Scholar] [CrossRef] [Green Version]
- Arthur, C.L.; Pawliszyn, J. Solid-Phase Microextraction with thermal-desorption using fused silica optical fibers. Anal. Chem. 1990, 62, 2145–2148. [Google Scholar] [CrossRef]
- Pawliszyn, J.; Pawliszyn, B.; Pawliszyn, M. Solid Phase Microextraction (SPME). Chem. Educ. 1997, 2, 1–7. [Google Scholar] [CrossRef]
- Monnin, T.; Malosse, C.; Peeters, C. Solid-phase microextraction and cuticular hydrocarbon differences related to reproductive activity in queenless ant Dinoponera quadriceps. J. Chem. Ecol. 1998, 24, 473–490. [Google Scholar] [CrossRef]
- Turillazzi, S.; Sledge, M.F.; Moneti, G. Use of a simple method for sampling cuticular hydrocarbons from live social wasps. Ethol. Ecol. Evol. 1998, 10, 293–297. [Google Scholar] [CrossRef]
- Tentschert, J.; Bestmann, H.J.; Heinze, J. Cuticular compounds of workers and queens in two Leptothorax ant species—A comparison of results obtained by solvent extraction, solid sampling, and SPME. Chemoecology 2002, 12, 15–21. [Google Scholar] [CrossRef]
- AL-Khshema, H.; Agarwal, M.; Ren, Y.L. Optimization and validation for determination of volatile organic compounds from Mediterranean Fruit Fly (Medfly) Ceratitis capitata (Diptera: Tephritidae) by using HS-SPME-GC-FID/MS. J. Biol. Sci. 2017, 17, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Robacker, D.C.; Warfield, W.C.; Flath, R.A. A four-component attractant for the mexican fruit fly, Anastrepha ludens (Diptera: Tephritidae), from host fruit. J. Chem. Ecol. 1992, 18, 1239–1254. [Google Scholar] [CrossRef]
- Cruz-Lopez, L.; Malo, E.A.; Toledo, J.; Virgen, A.; del Mazo, A.; Rojas, J.C. A new potential attractant for Anastrepha obliqua from Spondias mombin fruits. J. Chem. Ecol. 2006, 32, 351–365. [Google Scholar] [CrossRef]
- Diaz-Fleischer, F.; C., P.J.; Shelly, T.E. Interactions between tephritid fruit fly physiological state and stimuli from baits and traps: Looking for the pied piper of Hamelin to lure pestiferous fruit flies. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T.E., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 145–172. [Google Scholar]
- Jang, E.B.; Light, D.M.; Binder, R.G.; Flath, R.A.; Carvalho, L.A. Attraction of female mediterranean fruit flies to the five major components of male-produced pheromone in a laboratory flight tunnel. J. Chem. Ecol. 1994, 20, 9–20. [Google Scholar] [CrossRef]
- Oldham, N.J. Chemical Studies on Exocrine Gland Secretion and Pheromones of Some Social Insects. Ph.D. Thesis, Keele University, Keele, UK, 1994. [Google Scholar]
- Sinha, A.E.; Fraga, C.G.; Prazen, B.J.; Synovec, R.E. Trilinear chemometric analysis of two-dimensional comprehensive gas chromatography-time-of-flight mass spectrometry data. J. Chromatogr. A 2004, 1027, 269–277. [Google Scholar] [CrossRef]
- van Deursen, M.M.; Beens, J.; Janssen, H.G.; Leclercq, P.A.; Cramers, C.A. Evaluation of time-of-flight mass spectrometric detection for fast gas chromatography. J. Chromatogr. A 2000, 878, 205–213. [Google Scholar] [CrossRef]
- Vanickova, L.; Virgilio, M.; Tomcala, A.; Brizova, R.; Ekesi, S.; Hoskovec, M.; Kalinova, B.; Do Nascimento, R.R.; De Meyer, M. Resolution of three cryptic agricultural pests (Ceratitis fasciventris, C. anonae, C. rosa, Diptera: Tephritidae) using cuticular hydrocarbon profiling. Bull. Entomol. Res. 2014, 104, 631–638. [Google Scholar] [CrossRef]
- Warthen, J.D.; Mcgovern, T.P. Gc/Ftir analyses of Trimedlure isomers and related esters. J. Chromatogr. Sci. 1986, 24, 451–457. [Google Scholar] [CrossRef]
- Karas, M.; Bachmann, D.; Bahr, U.; Hillenkamp, F. Matrix-assisted ultraviolet laser desorption of non-volatile compounds. Int. J. Mass Spectrom. Ion Process. 1987, 78, 53–68. [Google Scholar] [CrossRef]
- Patton, G.M.; Fasulo, J.M.; Robins, S.J. Analysis of lipids by high performance liquid chromatography: Part I. J. Nutr. Biochem. 1990, 1, 493–500. [Google Scholar] [CrossRef]
- McHowad, J.; Jones, J.H.; Creer, H.M. Quantification of individual phospholipid molecular species by UV absorption measurements. J. Lipid Res. 1996, 37, 2450–2460. [Google Scholar] [CrossRef]
- McHowad, J.; Jones, J.H.; Creer, H.M. Gradient elution reverse-phase chromatographic isolation of individual glycerolphospholipid molecular species. J. Chromatogr. B 1997, 702, 21–32. [Google Scholar] [CrossRef]
- Olsson, N.U.; Harding, A.J.; Harper, C.; Salem, N.J. High-performance liquid chromatography method with light-scattering detection for measurement of lipid class composition: Analysis of brain from alcoholics. J. Chromatogr. B 1996, 681, 213–218. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, Q.; Zhang, P.W.; Chen, X.T.; Liu, B.J.; Cheng, D.M.; Zhang, Z.X. Integrated LC-MS and GC-MS-based untargeted metabolomics studies of the effect of azadirachtin on Bactrocera dorsalis larvae. Sci. Rep. 2020, 10, 2306. [Google Scholar] [CrossRef] [Green Version]
- Buděšínský, M.; Pelař, J. Cyklus Organická Chemie; Institute of Organic Chemistry and Biochemistry: Praha, Czech Republic, 2000. [Google Scholar]
- Baker, J.D.; Heath, R.R. NMR spectral assignment of lactone pheromone components emitted by Caribbean and Mexican fruit flies. J. Chem. Ecol. 1993, 19, 1511–1519. [Google Scholar] [CrossRef]
- Mori, K. Significance of chirality in pheromone science. Bioorg. Med. Chem. 2007, 15, 7505–7523. [Google Scholar] [CrossRef]
- Olszewski, T.K.; Grison, C. A concise synthesis of sex pheromone of Mediterranean fruit fly, Ceratitis capitata via lithiated carbanion derived from enephosphoramide. Heteroat. Chem. 2010, 21, 139–147. [Google Scholar] [CrossRef]
- Mori, K.; Uematsu, T.; Yanagi, K.; Minobe, M. Synthesis of the optically active forms of 4,10-dihydroxy-1,7-dioxaspiro[5.5]undecane and their conversion to the enantiomers of 1,7-dioxaspiro[5.5]undecane, the olive fly pheromone. Tetrahedron 1985, 41, 2751–2758. [Google Scholar] [CrossRef]
- Fusini, G.; Barsanti, D.; Angelici, G.; Casotti, G.; Canale, A.; Benelli, G.; Lucchi, A.; Carpita, A. Identification and synthesis of new sex-specific components of olive fruit fly (Bactrocera oleae) female rectal gland, through original Negishi reactions on supported catalysts. Tetrahedron 2018, 74, 4381–4389. [Google Scholar] [CrossRef]
- Canale, A.; Germinara, S.G.; Carpita, A.; Benelli, G.; Bonsignori, G.; Stefanini, C.; Raspi, A.; Rotundo, G. Behavioural and electrophysiological responses of the olive fruit fly, Bactrocera oleae (Rossi) (Diptera: Tephritidae), to male-and female-borne sex attractants. Chemoecology 2013. [Google Scholar] [CrossRef]
- Vaníčková, L. Chemical Ecology of Fruit Flies: Genera Ceratitis and Anastrepha. Ph.D. Thesis, Institute of Chemical Technology, Prague, Czech Republic, 2012. [Google Scholar]
- Břízová, R. Analyses of Male Sex Pheromone of Anastrepha fraterculus (Diptera: Tephritidae). Master’s Thesis, Institute of Chemical Technology, Prague, Czech Republic, 2011. [Google Scholar]
- Zykova, K. Quantitative Composition Changes of Sex Pheromone in Anastrepha fraterculus Depending on Age. Master’s Thesis, Institute of Chemical Technology, Prague, Czech Republic, 2013. [Google Scholar]
- Cruz-Lopez, L.; Malo, E.A.; Rojas, J.C. Sex Pheromone of Anastrepha striata. J. Chem. Ecol. 2015, 41, 458–464. [Google Scholar] [CrossRef]
- Noushini, S.; Perez, J.; Park, S.J.; Holgate, D.; Alvarez, V.M.; Jamie, I.; Jamie, J.; Taylor, P. Attraction and electrophysiological response to identified rectal gland volatiles in Bactrocera frauenfeldi (Schiner). Molecules 2020, 25, 1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robacker, D.C.; Chapa, B.E.; Hart, W.G. Electroantennograms of Mexican fruit flies to chemicals produced by males. Entomol. Exp. Appl. 1986, 40, 123–127. [Google Scholar] [CrossRef]
- Van Der Pers, J.C.N.; Haniotakis, G.E.; King, B.M. Electroantennogram responses from olfactory receptors in Dacus oleae. Entomol. Hell. 1984, 2, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Siderhurst, M.S.; Jang, E.B. Cucumber volatile blend attractive to female melon fly, Bactrocera cucurbitae (Coquillett). J. Chem. Ecol. 2010, 36, 699–708. [Google Scholar] [CrossRef]
- Liscia, A.; Angioni, P.; Sacchetti, P.; Poddighe, S.; Granchietti, A.; Setzu, M.D.; Belcari, A. Characterization of olfactory sensilla of the olive fly: Behavioral and electrophysiological responses to volatile organic compounds from the host plant and bacterial filtrate. J. Insect Physiol. 2013, 59, 705–716. [Google Scholar] [CrossRef]
- Cunningham, J.P.; Carlsson, M.A.; Villa, T.F.; Dekker, T.; Clarke, A.R. Do fruit ripening volatiles enable resource specialism in polyphagous fruit flies? J. Chem. Ecol. 2016, 42, 931–940. [Google Scholar] [CrossRef]
- Biasazin, T.D.; Karlsson, M.F.; Hillbur, Y.; Seyoum, E.; Dekker, T. Identification of host blends that attract the African invasive fruit fly, Bactrocera invadens. J. Chem. Ecol. 2014, 40, 966–976. [Google Scholar] [CrossRef]
- Kendra, P.E.; Epsky, N.D.; Montgomery, W.S.; Heath, R.R. Response of Anastrepha suspensa (Diptera: Tephritidae) to terminal diamines in a food-based synthetic attractant. Environ. Entomol. 2008, 37, 1119–1125. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Mateo, D.M.; Puche, H.; Epsky, N.D.; Heath, R.R. Effect of age on EAG response and attraction of female Anastrepha suspensa (Diptera: Tephritidae) to ammonia and carbon dioxide. Environ. Entomol. 2005, 34, 584–590. [Google Scholar] [CrossRef]
- Tabanca, N.; Masi, M.; Epsky, N.D.; Nocera, P.; Cimmino, A.; Kendra, P.E.; Niogret, J.; Evidente, A. Laboratory evaluation of natural and synthetic aromatic compounds as potential attractants for male Mediterranean fruit fly, Ceratitis capitata. Molecules 2019, 24, 2409. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, D.A.; Kendra, P.E.; Epsky, N.D.; Montgomery, W.S.; Heath, R.R.; Jenkins, D.M.; Goenaga, R. Antennal responses of West Indian and Caribbean fruit flies (Diptera: Tephritidae) to ammonium bicarbonate and putrescine lures. Florida Entomol. 2012, 95, 28–34. [Google Scholar] [CrossRef]
- Epsky, N.; Kendra, P.E.; Heath, R.R. Response of Anastrepha suspensa to liquid protein baits and synthetic lure formulations. In Proceedings of the 7th International Symposium on Fruit Flies of Economic Importance, Salvador, Brazil, 10–15 September 2006; pp. 81–88. [Google Scholar]
- Crnjar, R.; Scalera, G.; Liscia, A.; Angioy, A.M.; Bigiani, A.; Pietra, P.; Barbarossa, I.T. Morphology and eag mapping of the antennal olfactory receptors in Dacus oleae. Entomol. Exp. Appl. 1989, 51, 77–85. [Google Scholar] [CrossRef]
- Nagai, T. On the relationship between the electroantennogram and simultaneously recorded single sensillum response of the European corn borer Ostrinia nubilalis. Arch. Insect Biochem. Physiol. 1983, 1, 85–91. [Google Scholar] [CrossRef]
- Olsson, S.B.; Hansson, B.S. Electroantennogram and single sensillum recording in insect antennae. In Methods in Molecular Biology; Touhara, K., Ed.; Springer Science and Business Media LLC: Berlin, Germany, 2013; Volume 1068. [Google Scholar]
- Bigiani, A.; Scalera, G.; Crnjar, R.; Barbarossa, I.T.; Magherini, P.C.; Pietra, P. Distribution and function of the antennal olfactory sensilla in Ceratitis capitata Wied (Diptera, Trypetidae). Boll. Di Zool. 1989, 56, 305–311. [Google Scholar] [CrossRef]
- Bisotto-de-Oliveira, R.; Redaelli, L.R.; Sant’ana, J. Morphometry and distribution of sensilla on the antennae of Anastrepha fraterculus (Wiedemann) (Diptera: Tephritidae). Neotrop. Entomol. 2011, 40, 212–216. [Google Scholar] [CrossRef]
- Jacob, V.; Scolari, F.; Delatte, H.; Gasperi, G.; Jacquin-Joly, E.; Malacrida, A.R.; Duyck, P.F. Current source density mapping of antennal sensory selectivity reveals conserved olfactory systems between tephritids and Drosophila. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Jacob, V.E.J.M. Current source density analysis of electroantennogram recordings: A tool for mapping the olfactory response in an insect antenna. Front. Cell. Neurosci. 2018, 12. [Google Scholar] [CrossRef] [Green Version]
- Loy, F.; Solari, P.; Isola, M.; Crnjar, R.; Masala, C. Morphological and electrophysiological analysis of tarsal sensilla in the medfly Ceratitis capitata (Wiedemann, 1824) (Diptera: Tephritidae). Ital. J. Zool. 2016, 83, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Tait, C.; Batra, S.; Ramaswamy, S.S.; Feder, J.L.; Olsson, S.B. Sensory specificity and speciation: A potential neuronal pathway for host fruit odour discrimination in Rhagoletis pomonella. Proc. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, J.D. Bioassays with terrestrial invertebrates. In Methods in Chemical Ecology, Bioassays Methods; Hayes, K.F., Ed.; Kluwer Academic: Norwell, MA, USA, 1998; Volume 2, pp. 212–270. [Google Scholar]
- Howse, P.E.; Stevens, I.D.R.; Jones, O.T.; Howse, P.E.; Stevens, I.D.R.; Jones, O.T. Bioassay methods. In Insect Pheromones and Their Use in Pest Management; Springer: Dordrecht, The Netherlands, 1998. [Google Scholar]
- Baker, T.C.; Cardé, R.T. Techniques for Behavioral Bioassays. In Techniques in Pheromone Research. Springer Series in Experimental Entomology; Hummel, H.E., Miller, T.A., Eds.; Springer New York: New York, NY, USA, 1984; pp. 45–73. [Google Scholar]
- Knudsen, G.K.; Tasin, M.; Aak, A.; Thöming, G. A wind tunnel for odor mediated insect behavioural assays. J. Vis. Exp. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Lin, J.; Zhang, H.; Zeng, X. BdorOBP83a-2 mediates responses of the Oriental fruit fly to semiochemicals. Front. Physiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazomenos, B.E.; Haniotakis, G.E. Male olive fruit fly attraction to synthetic sex pheromone components in laboratory and field tests. J. Chem. Ecol. 1985, 11, 397–405. [Google Scholar] [CrossRef]
- Giunti, G.; Campolo, O.; Laudani, F. Olive fruit volatiles route intraspecific interactions and chemotaxis in Bactrocera oleae (Rossi) (Diptera: Tephritidae) females. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Landolt, P.J.; Heath, R.R.; Chambers, D.L. Oriented flight responses of female Mediterranean fruit flies to calling males, odor of calling males, and a synthetic pheromone blend. Entomol. Exp. Appl. 1992, 65, 259–266. [Google Scholar] [CrossRef]
- Biasazin, T.D.; Herrera, S.L.; Kimbokota, F.; Dekker, T. Translating olfactomes into attractants: Shared volatiles provide attractive bridges for polyphagy in fruit flies. Ecol. Lett. 2019, 22, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Baker, P.; Howse, P.; Ondarza, R.N.; Reyes, J. Field trials of synthetic sex pheromone components of the male Mediterranean fruit fly (Diptera: Tephritidae) in Southern Mexico. J. Econ. Entomol. 1990, 83, 2235–2245. [Google Scholar] [CrossRef]
- Jones, O.T.; Lisk, J.C.; Longhurst, C.; Howse, P.E. Development of a monitoring trap for the olive fruit fly, Dacus oleae (Gmelin) (Diptera: Tephritidae), using a multicomponent of its sex pheromone as lure. Bull. Entomol. Res. 1983, 73, 97–106. [Google Scholar] [CrossRef]
- Mazomenos, B.E.; Haniotakis, G.E.; Ioannou, A.; Spanakis, I.; Kozirakis, A. Field evaluation of the olive fruit fly pheromone traps with various dispensers and concentrations. In Proceedings of the International Symposium of Fruit Flies of Economic Importance, Athens, Greece, 16 November 1982; pp. 506–512. [Google Scholar]
- Perdomo, A.J.; Baranowski, R.M.; Nation, J.L. Recapture of virgin female Caribbean fruit flies from traps baited with males. Florida Entomol. 1975, 58, 291–295. [Google Scholar] [CrossRef]
- Perdomo, A.J.; Nation, J.L.; Baranowski, R.M. Attraction of female and male caribbean fruit flies to food-baited and male-baited traps under field conditions. Environ. Entomol. 1976. [Google Scholar] [CrossRef]
- Nakagawa, S.; Steiner, L.F.; Farias, G.J. Response of virgin female Mediterranean fruit flies to live mature normal males, sterile males, and Trimedlure in plastic traps. J. Econ. Entomol. 1981, 74, 566–567. [Google Scholar] [CrossRef]
- Shelly, T.E. Male signalling and lek attractiveness in the Mediterranean fruit fly. Anim. Behav. 2000, 60, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.C.; Burk, T.; Sivinski, J. Attraction of female Caribbean fruit flies, Anastrepha suspensa (Diptera: Tephritidae), to the presence of males and male-produced stimuli in field cages. Ann. Entomol. Soc. Am. 1983, 76, 996–998. [Google Scholar] [CrossRef]
- Ono, H.; Hee, A.K.-W.; Jiang, H. Recent advancements in studies on chemosensory mechanisms underlying detection of semiochemicals in Dacini fruit flies of economic importance (Diptera: Tephritidae). Insects 2021, 12, 106. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. Camb. Philos. Soc. 2018, 93, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Gomulski, L.M.; Dimopoulos, G.; Xi, Z.Y.; Soares, M.B.; Bonaldo, M.F.; Malacrida, A.R.; Gasperi, G. Gene discovery in an invasive tephritid model pest species, the Mediterranean fruit fly, Ceratitis capitata. BMC Genomics 2008, 9. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.W.; Peng, T.; He, W.; Zhang, H.Y. High-throughput sequencing to reveal genes involved in reproduction and development in Bactrocera dorsalis (Diptera: Tephritidae). PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.W.; Peng, W.; Zhu, C.P.; Zhang, Q.; Saccone, G.; Zhang, H.Y. Identification and expression profile analysis of odorant binding proteins in the Oriental fruit fly Bactrocera dorsalis. Int. J. Mol. Sci. 2013, 14, 14936–14949. [Google Scholar] [CrossRef] [Green Version]
- Ramsdell, K.M.; Lyons-Sobaski, S.A.; Robertson, H.M.; Walden, K.K.; Feder, J.L.; Wanner, K.; Berlocher, S.H. Expressed sequence tags from cephalic chemosensory organs of the northern walnut husk fly, Rhagoletis suavis, including a putative canonical odorant receptor. J. Insect Sci. 2010, 10, 51. [Google Scholar] [CrossRef]
- Schwarz, D.; Robertson, H.M.; Feder, J.L.; Varala, K.; Hudson, M.E.; Ragland, G.J.; Hahn, D.A.; Berlocher, S.H. Sympatric ecological speciation meets pyrosequencing: Sampling the transcriptome of the apple maggot Rhagoletis pomonella. BMC Genomics 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.Z.; Zhang, H.; Wang, Z.B.; Bin, S.Y.; He, H.L.; Lin, J.T. Discovery of chemosensory genes in the Oriental fruit fly, Bactrocera dorsalis. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.Z.; Kang, C.; Qu, M.Q.; Chen, J.L.; Chen, M.S.; Bin, S.Y.; Lin, J.T. Candidates for chemosensory genes identified in the Chinese citrus fly, Bactrocera minax, through a transcriptomic analysis. BMC Genomics 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-F.; Xu, L.; Zhang, Y.-X.; Wei, D.; Wang, J.-J.; Jiang, H.-B. Genome-wide identification and expression profiling of odorant-binding proteins in the oriental fruit fly, Bactrocera dorsalis. Comp. Biochem. Physiol. Part D Genomics Proteomics 2019, 31, 100605. [Google Scholar] [CrossRef]
- Xu, P.H.; Wang, Y.H.; Akami, M.; Niu, C.Y. Identification of olfactory genes and functional analysis of BminCSP and BminOBP21 in Bactrocera minax. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Campanini, E.B.; de Brito, R.A. Molecular evolution of Odorant-binding proteins gene family in two closely related Anastrepha fruit flies. BMC Evol. Biol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Rezende, V.B.; Congrains, C.; Lima, A.L.; Campanini, E.B.; Nakamura, A.M.; Oliveira, J.L.; Chahad-Ehlers, S.; Junior, I.S.; Alves de Brito, R. Head transcriptomes of two closely related species of fruit flies of the Anastrepha fraterculus group reveals divergent genes in species with extensive gene flow. G3 2016, 6, 3283–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanicolaou, A.; Schetelig, M.F.; Arensburger, P.; Atkinson, P.W.; Benoit, J.B.; Bourtzis, K.; Castanera, P.; Cavanaugh, J.P.; Chao, H.; Childers, C.; et al. The whole genome sequence of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann), reveals insights into the biology and adaptive evolution of a highly invasive pest species. Genome Biol. 2016, 17, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhao, X.F.; Fu, L.; Han, Y.Y.; Chen, J.; Lu, Y.Y. BdorOBP2 plays an indispensable role in the perception of methyl eugenol by mature males of Bactrocera dorsalis (Hendel). Sci. Rep. 2017, 7, 15894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Liang, X.F.; Xu, L.; Keesey, I.W.; Lei, Z.R.; Smagghe, G.; Wang, J.J. An antennae-specific Odorant-Binding Protein is involved in Bactrocera dorsalis olfaction. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.M.; Chahad-Ehlers, S.; Lima, A.L.A.; Taniguti, C.H.; Sobrinho, I.; Torres, F.R.; de Brito, R.A. Reference genes for accessing differential expression among developmental stages and analysis of differential expression of OBP genes in Anastrepha obliqua. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhu, J.; Knoll, W. Odorant-binding proteins as sensing elements for odour monitoring. Sensors 2018, 18, 3248. [Google Scholar] [CrossRef] [Green Version]
- Falchetto, M.; Ciossani, G.; Scolari, F.; Di Cosimo, A.; Nenci, S.; Field, L.M.; Mattevi, A.; Zhou, J.J.; Gasperi, G.; Forneris, F. Structural and biochemical evaluation of Ceratitis capitata odorant-binding protein 22 affinity for odorants involved in intersex communication. Insect Mol. Biol. 2019, 28, 431–443. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhu, J.; Knoll, W. From radioactive ligands to biosensors: Binding methods with olfactory proteins. Appl. Microbiol. Biotechnol. 2018, 102, 8213–8227. [Google Scholar] [CrossRef]
- Paolini, S.; Tanfani, F.; Fini, C.; Bertoli, E.; Pelosi, P. Porcine odorant-binding protein: Structural stability and ligand affinities measured by Fourier-transform infrared spectroscopy and fluorescence spectroscopy. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1999, 1431, 179–188. [Google Scholar] [CrossRef]
- Wojtasek, H.; Leal, W.S. Conformational change in the pheromone-binding protein from Bombyx mori induced by pH and by interaction with membranes. J. Biol. Chem. 1999, 274, 30950–30956. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.L.; Tuccori, E.; He, X.L.; Gazzano, A.; Field, L.; Zhou, J.J.; Pelosi, P. Discrimination of alarm pheromone (E)-beta-farnesene by aphid odorant-binding proteins. Insect Biochem. Mol. Biol. 2009, 39, 414–419. [Google Scholar] [CrossRef]
- Ban, L.; Scaloni, A.; D’Ambrosio, C.; Zhang, L.; Yan, Y.; Pelosi, P. Biochemical characterization and bacterial expression of an odorant-binding protein from Locusta migratoria. Cell. Mol. Life Sci. 2003, 60, 390–400. [Google Scholar] [CrossRef]
- Drew, R.A.I. The responses of fruit fly species (Diptera: Tephritidae) in the South Pacific area to male attractants. Aust. J. Entomol. 1974, 13, 267–270. [Google Scholar] [CrossRef]
- Wee, S.-L.; Peek, T.; Clarke, A.R. The responsiveness of Bactrocera jarvisi (Diptera: Tephritidae) to two naturally occurring phenylbutaonids, zingerone and raspberry ketone. J. Insect Physiol. 2018, 109, 41–46. [Google Scholar] [CrossRef]
- Park, K.C.; Jeong, S.A.; Kwon, G.; Oh, H.-W. Olfactory attraction mediated by the maxillary palps in the striped fruit fly, Bactrocera scutellata: Electrophysiological and behavioral study. Arch. Insect Biochem. Physiol. 2018, 99, e21510. [Google Scholar] [CrossRef]
- Jayanthi, P.D.K.; Kempraj, V.; Aurade, R.M.; Roy, T.K.; Shivashankara, K.S.; Verghese, A. Computational reverse chemical ecology: Virtual screening and predicting behaviorally active semiochemicals for Bactrocera dorsalis. BMC Genomics 2014, 15. [Google Scholar] [CrossRef] [Green Version]
- Jayanthi, P.D.K.; Kempraj, V.; Aurade, R. Computational reverse chemical ecology: Prospecting semiochemicals for pest management using in silico approach in Plutella xylostella Linn. Pest Manag. Hortic. Ecosyst. 2016, 22, 20–27. [Google Scholar]
- Kuntz, I.D.; Blaney, J.M.; Oatley, S.J.; Langridge, R.; Ferrin, T.E. A geometric approach to macromolecule-ligand interactions. J. Mol. Biol. 1982, 161, 269–288. [Google Scholar] [CrossRef]
- Meng, X.Y.; Zhang, H.X.; Mezei, M.; Cui, M. Molecular docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- McConkey, B.J.; Sobolev, V.; Edelman, M. The performance of current methods in ligand-protein docking. Curr. Sci. 2002, 83, 845–856. [Google Scholar]
- Venthur, H.; Zhou, J.-J. Odorant receptors and odorant-binding proteins as insect pest control targets: A comparative analysis. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Liu, Z.; Smagghe, G.; Lei, Z.R.; Wang, J.J. Identification of male- and female-specific olfaction genes in antennae of the Oriental fruit fly (Bactrocera dorsalis). PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, H.; Otake, J.; Mitsuno, H.; Ozaki, K.; Kanzaki, R.; Chieng, A.C.T.; Hee, A.K.W.; Nishida, R.; Ono, H. Functional characterization of olfactory receptors in the Oriental fruit fly Bactrocera dorsalis that respond to plant volatiles. Insect Biochem. Mol. Biol. 2018, 101, 32–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Zhou, X.F.; Gu, F.; Zhong, G.H.; Yi, X. Olfactory plasticity: Variation in the expression of chemosensory receptors in Bactrocera dorsalis in different physiological states. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Tsoumani, K.T.; Belavilas-Trovas, A.; Gregoriou, M.-E.; Mathiopoulos, K.D. Anosmic flies: What Orco silencing does to olive fruit flies. BMC Genet. 2020, 21, 140. [Google Scholar] [CrossRef]
- Ono, H.; Miyazaki, H.; Mitsuno, H.; Ozaki, K.; Kanzaki, R.; Nishida, R. Functional characterization of olfactory receptors in three Dacini fruit flies (Diptera: Tephritidae) that respond to 1-nonanol analogs as components in the rectal glands. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2020, 239. [Google Scholar] [CrossRef]
- Sato, K.; Pellegrino, M.; Nakagawa, T.; Nakagawa, T.; Vosshall, L.B.; Touhara, K. Insect olfactory receptors are heteromeric ligand-gated ion channels. Nature 2008, 452, 1002–1006. [Google Scholar] [CrossRef]
- Butterwick, J.A.; del Mármol, J.; Kim, K.H.; Kahlson, M.A.; Rogow, J.A.; Walz, T.; Ruta, V. Cryo-EM structure of the insect olfactory receptor Orco. Nature 2018, 560, 447–452. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Z.-S.; Zhang, D.-J.; Lu, Y.-Y. BdorOR88a modulates the responsiveness to methyl eugenol in mature males of Bactrocera dorsalis (Hendel). Front. Physiol. 2018, 9. [Google Scholar] [CrossRef]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Structure-function of the G Protein–Coupled Receptor superfamily. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 531–556. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.-F.; Yu, T.; Chen, Z.-J.; Chen, S.; Chen, Y.-P.; Gao, L.; Zhang, W.-H.; Jiang, B.; Bai, X.; Walker, E.D.; et al. Comparative genomic and transcriptomic analyses of chemosensory genes in the citrus fruit fly Bactrocera (Tetradacus) minax. Sci. Rep. 2020, 10, 18068. [Google Scholar] [CrossRef]
- Porter, B.A. The Apple Maggot. U.S. Dep. Agric. Tech. Bull. 1928, 66, 1–48. [Google Scholar]
- Häfliger, E. Das Auswahlvermögen der Kirschenfliege bei der Eiablage. Mitt. Schweiz. Entomol. Ges. 1953, 26, 258–264. [Google Scholar]
- Prokopy, R.J.; Greany, P.; Chambers, D. Oviposition-deterring pheromone in Anastrepha suspensa. Environ. Entomol. 1977, 6, 463–465. [Google Scholar] [CrossRef]
- Simões, M.H.; Polloni, Y.J.; Pauludetti, M.A. Biología de algunas especies de Anastrepha (Diptera: Tephritidae) en laboratório. In Proceedings of the III Latin-American Entomology Congress, Ilheus, Brazil, 23 July 1978. [Google Scholar]
- Prokopy, R.J.; Malavasi, A.; Morgante, J.S. Oviposition deterring pheromone in Anastrepha fraterculus flies. J. Chem. Ecol. 1982, 8, 763–771. [Google Scholar] [CrossRef]
- Poloni, Y.J.; Silva, M.T. Considerations on the reproductive of Anastrepha pseudoparallela Loew 1873 (Diptera: Tephritidae). In Proceedings of the Fruit Flies: II International Symposium; Elsevier Science Publishers: Kolymbari, Greece, 1986. [Google Scholar]
- Selivon, D. Alguns Aspectos do Comportamento de Anastrepha striata Schiner e Anastrepha bistrigata Bezzii (Diptera: Tephritidae). Master’s Thesis, University of São Paulo, Sao Paulo, Brazil, 1991. [Google Scholar]
- Silva, J.G. Biologia e Comportamento de Anastrepha grandis (Macquart, 1846) (Diptera: T ephritidae). Master’s Thesis, University of São Paulo, Sao Paulo, Brazil, 1991. [Google Scholar]
- Aluja-Schuneman, M.R.; Díaz-Fleischer, F.; Edmunds, A.J.F.; Hagmann, L. Isolation, Structural Determination, Synthesis, Biological Activity and Application as Control Agent of the Host Marking Pheromone (and Derivatives Thereof) of the Fruit Flies of the Type Anastrepha (Diptera: Tephritidae). US6555120B1, 29 April 2003. [Google Scholar]
- Averill, A.L.; Prokopy, R.J. Residual activity of oviposition-deterring pheromone in Rhagoletis pomonella (Diptera: Tephritidae) and female response to infested fruit. J. Chem. Ecol. 1987, 13, 167–177. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Reynolds, A.H. Ovipositional enhancement through socially facilitated behavior in Rhagoletis pomonella flies. Entomol. Exp. Appl. 1998, 86, 281–286. [Google Scholar] [CrossRef]
- Wiesmann, R. Die orientierung der kirschfliege Rhagoletis cerasi L. Landwirtsch. Jahrb. Schweiz 1937, 51, 1080–1109. [Google Scholar]
- Katsoyannos, B.I. Oviposition-deterring, male-arresting, fruit-marking pheromone in Rhagoletis cerasi. Environ. Entomol. 1975, 4, 801–807. [Google Scholar] [CrossRef]
- Prokopy, R.J. Oviposition-deterring fruit marking pheromone in Rhagoletis fausta. Environ. Entomol. 1975, 4, 298–300. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Reissig, W.H.; Moericke, V. Marking pheromones deterring repeated oviposition in Rhagoletis flies. Entomol. Exp. Appl. 1976, 20, 170–178. [Google Scholar] [CrossRef]
- Bauer, G. Life-history strategy of Rhagoletis alternata (Diptera: Trypetidae), a fruit fly operating in a ‘non-interactive’ system. J. Anim. Ecol. 1986, 55, 785. [Google Scholar] [CrossRef]
- Arredondo, J.; Diaz-Fleischer, F. Oviposition deterrents for the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae) from fly faeces extracts. Bull. Entomol. Res. 2006, 96, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Roitberg, B.D.; Prokopy, R.J. Host deprivation influence on response of Rhagoletis pomonella to its oviposition deterring pheromone. Physiol. Entomol. 1983, 8, 69–72. [Google Scholar] [CrossRef]
- Mumtaz, M.; AliNiazee, M. The oviposition-deterring pheromone in the western cherry fruit fly, Rhagoletis indifferens Curran (Dipt., Tephritidae). Biological properties. Zeitschrift für Angew. Entomol. 1983, 96, 83–93. [Google Scholar] [CrossRef]
- Boller, E. Oviposition-deterring pheromone of the European cherry fruit fly: Status of research and potential applications. In Management of Insect Pests with Semiochemicals; Mitchell, E.R., Ed.; Springer: Boston, MA, USA, 1981; pp. 457–462. [Google Scholar]
- Boller, E.F.; Hurter, J. Oviposition deterring pheromone in Rhagoletis cerasi: Behavioral laboratory test to measure pheromone activity. Entomol. Exp. Appl. 1985, 39, 163–169. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Averill, A.L.; Bardinelli, C.M.; Bowdan, E.S.; Cooley, S.S.; Crnjar, R.M.; Dundulis, E.A.; Roitberg, C.A.; Spatcher, P.J.; Tumlinson, J.H.; et al. Site of production of an oviposition-deterring pheromone component in Rhagoletis pomonella flies. J. Insect Physiol. 1982, 28, 1–7. [Google Scholar] [CrossRef]
- Hurter, J.; Katsoyannos, B.; Boller, E.F.; Wirz, P. Beitrag zur Anreicherung und teilweisen Reinigung des eiablageverhindernden Pheromons der Kirschenfliege, Rhagoletis cerasi L. (Dipt., Trypetidae). Zeitschrift für Angew. Entomol. 1976, 80, 50–56. [Google Scholar] [CrossRef]
- Kachigamba, D.L. Host-marking behaviour and pheromones in major fruit fly species (Diptera: Tephritidae) infesting mango (Mangifera indica) in Kenya. Ph.D. Thesis, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya, 2012. [Google Scholar]
- Ernst, B.; Wagner, B. Synthesis of the oviposition-deterring pheromone (Odp) in Rhagoletis cerasi L. Helv. Chim. Acta 1989, 72, 165–171. [Google Scholar] [CrossRef]
- Boller, E.F.; Aluja, M. Oviposition deterring pheromone in Rhagoletis cerasi L. Biological activity of 4 synthetic isomers and HMP discrimination of two host races as measured by an improved laboratory bioassay. J. Appl. Entomol. 1992, 113, 113–119. [Google Scholar] [CrossRef]
- Edmunds, A.J.F.; Aluja, M.; Diaz-Fleischer, F.; Patrian, B.; Hagmann, L. Host marking pheromone (HMP) in the Mexican fruit fly Anastrepha ludens. Chimia 2010, 64, 37–42. [Google Scholar] [CrossRef]
- Scarpati, M.L.; Loscalzo, R.; Vita, G. Olea europaea volatiles attractive and repellent to the Olive fruit fly (Dacus oleae, Gmelin). J. Chem. Ecol. 1993, 19, 881–891. [Google Scholar] [CrossRef]
- Kachigamba, D.L.; Ekesi, S.; Ndung’u, M.W.; Gitonga, L.M.; Teal, P.E.A.; Torto, B. Evidence for potential of managing some African fruit fly species (Diptera: Tephritidae) using the mango fruit fly host-marking pheromone. J. Econ. Entomol. 2012, 105, 2068–2075. [Google Scholar] [CrossRef]
- Carlson, D.A.; Mayer, M.S.; Silhacek, D.L.; James, J.D.; Beroza, M.; Bierl, B.A. Sex attractant pheromone of the house fly: Isolation, identification and synthesis. Science 1971. [Google Scholar] [CrossRef] [Green Version]
- Antony, C.; Jallon, J.-M. The chemical basis for sex recognition in Drosophila melanogaster. J. Insect Physiol. 1982, 28, 873–880. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Dillwith, J.W.; Adams, T.S. Biosynthesis and endocrine regulation of sex pheromone production in Diptera. In Pheromone Biochemistry; Elsevier: Amsterdam, The Netherlands, 1987; pp. 217–250. [Google Scholar]
- Rogoff, W.M.; Beltz, A.D.; Johnsen, J.O.; Plapp, F.W. A sex pheromone in the housefly, Musca domestica L. J. Insect Physiol. 1964, 10, 239–246. [Google Scholar] [CrossRef]
- Mayer, M.S.; James, J.D. Response of male Musca domestica to a specific olfactory attractant and its initial chemical purification. J. Insect Physiol. 1971, 17, 833–842. [Google Scholar] [CrossRef]
- Silhacek, D.L.; Carlson, D.A.; Mayer, M.S.; James, J.D. Composition and sex attractancy of cuticular hydrocarbons from houseflies: Effects of age, sex, and mating. J. Insect Physiol. 1972. [Google Scholar] [CrossRef]
- Stocker, R.F. The organization of the chemosensory system in Drosophila melanogaster: A review. Cell Tissue Res. 1994, 275, 3–26. [Google Scholar] [CrossRef]
- Boll, W.; Noll, M. The Drosophila Pox neuro gene: Control of male courtship behavior and fertility as revealed by a complete dissection of all enhancers. Development 2002, 129, 5667–5681. [Google Scholar] [CrossRef] [Green Version]
- Bray, S.; Amrein, H. A putative Drosophila pheromone receptor expressed in male-specific taste neurons is required for efficient courtship. Neuron 2003, 39, 1019–1029. [Google Scholar] [CrossRef] [Green Version]
- Jackson, L.L.; Bartelt, R.J. Cuticular hydrocarbons of Drosophila virilis. Comparison by age and sex. Insect Biochem. 1986, 16, 433–439. [Google Scholar] [CrossRef]
- Kim, Y.-K.; Phillips, D.R.; Chao, T.; Ehrman, L. Developmental isolation and subsequent adult behavior of Drosophila paulistorum. VI. Quantitative variation in cuticular hydrocarbons. Behav. Genet. 2004, 34, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.A.; Yocom, S.R. Cuticular hydrocarbons from six species of tephritid fruit flies. Arch. Insect Biochem. Physiol. 1986, 3, 397–412. [Google Scholar] [CrossRef]
- Bosa, C.F.; Cruz-Lopez, L.; Guillen-Navarro, K.; Zepeda-Cisneros, C.S.; Liedo, P. Variation in the cuticular hydrocarbons of the Mexican fruit fly Anastrepha ludens males between strains and age classes. Arch. Insect Biochem. Physiol. 2018, 99. [Google Scholar] [CrossRef] [PubMed]
- Lavine, B.K.; Carlson, D.A.; Calkins, C.O. Classification of tephritid fruit fly larvae by gas chromatography pattern/recognition techniques. Microchem. J. 1992, 45, 50–57. [Google Scholar] [CrossRef]
- Sutton, B.D.; Carlson, D.A. Interspecific variation in tephritid fruit fly larvae surface hydrocarbons. Arch. Insect Biochem. Physiol. 1993, 23, 53–65. [Google Scholar] [CrossRef]
- Sutton, B.D.; Steck, G.J. Discrimination of Caribbean and Mediterranean fruit fly larvae (Diptera: Tephritidae) by cuticular hydrocarbon analysis. Florida Entomol. 1994, 77, 231. [Google Scholar] [CrossRef]
- Vaníčková, L.; Hernández-Ortiz, V.; Joachim Bravo, I.S.; Dias, V.; Passos Roriz, A.K.; Laumann, R.A.; de Mendonça, A.L.; Jordão Paranhos, B.A.; do Nascimento, R.R. Current knowledge of the species complex Anastrepha fraterculus (Diptera, tephritidae) in Brazil. Zookeys 2015, 540, 211–237. [Google Scholar] [CrossRef]
- Goh, S.H.; Ooi, K.E.; Chuah, C.H.; Yong, H.S.; Khoo, S.G.; Ong, S.H. Cuticular hydrocarbons from two species of Malaysian Bactrocera fruit flies. Biochem. Syst. Ecol. 1993, 21, 215–226. [Google Scholar] [CrossRef]
- Vanickova, L.; Nagy, R.; Pompeiano, A.; Kalinova, B. Epicuticular chemistry reinforces the new taxonomic classification of the Bactrocera dorsalis species complex (Diptera: Tephritidae, Dacinae). PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Galhoum, A. Taxonomic studies on two Tephritid species (Order: Diptera), Bactrocera oleae and B. zonata, using the cuticular hydrocarbons profile. Al Azhar Bull. Sci. 2017, 28, 45–54. [Google Scholar] [CrossRef]
- Park, S.J.; Pandey, G.; Castro-Vargas, C.; Oakeshott, J.G.; Taylor, P.W.; Mendez, V. Cuticular chemistry of the Queensland fruit fly Bactrocera tryoni (Froggatt). Molecules 2020, 25, 4185. [Google Scholar] [CrossRef]
- Arakaki, N.; Kuba, H.; Soemori, H. Mating behavior of the Oriental fruit fly, Dacus dorsalis Hendel (Diptera: Tephritidae). Appl. Entomol. Zool. 1984, 19, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Hu, L.; Zhou, X.; Dai, J.; Chen, B.; Li, S. Allyl-2,6-dimethoxyphenol, a female-biased compound, is robustly attractive to conspecific males of Bactrocera dorsalis at close range. Entomol. Exp. Appl. 2019, 167, 811–819. [Google Scholar] [CrossRef]
- Hu, L.; Chen, B.; Liu, K.; Yu, G.; Chen, Y.; Dai, J.; Zhao, X.; Zhong, G.; Zhang, Y.; Shen, J. OBP2 in the midlegs of the male Bactrocera dorsalis is involved in the perception of the female-biased sex pheromone 4-allyl-2,6-dimethoxyphenol. J. Agric. Food Chem. 2021, 69, 126–134. [Google Scholar] [CrossRef]
- Bagneres, A.G.; Morgan, E.D. A simple method for analysis of insect cuticular hydrocarbons. J. Chem. Ecol. 1990, 16, 3263–3276. [Google Scholar] [CrossRef]
- Brill, J.H.; Bertsch, W. An investigation of sampling methods for the analysis of cuticular hydrocarbons. J. Entomol. Sci. 1985, 20, 435–443. [Google Scholar] [CrossRef]
- Cvacka, J.; Jiroš, P.; Šobotník, J.; Hanus, R.; Svatoš, A. Analysis of insect cuticular hydrocarbons using matrix-assisted laser desorption/ionization mass spectrometry. J. Chem. Ecol. 2006, 32, 409–434. [Google Scholar] [CrossRef]
- Chin, J.S.R.; Yew, J.Y. Pheromones in the Fruit Fly. Methods Mol. Biol. 2013, 1068, 15–27. [Google Scholar] [CrossRef]
- Lockey, K.H. Cuticular hydrocarbons of adult Cylindrinotus laevioctostriatus (Goeze) and Phylan gibbus (Fabricius) (Coleoptera: Tenebrionidae). Insect Biochem. 1981, 11, 549–561. [Google Scholar] [CrossRef]
- Kosi, A.Z.; Chinta, S.; Headrick, D.H.; Cokl, A.; Millar, J.G. Do chemical signals mediate reproductive behavior of Trupanea vicina, an emerging pest of ornamental marigold production in California? Entomol. Exp. Appl. 2013, 149, 44–56. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Carey, J.R.; Liedo, P.; Muller, H.-G.; Senturk, D. Virgin females compete for mates in the male lekking species Ceratitis capitata. Physiol. Entomol. 2009, 34, 238–245. [Google Scholar] [CrossRef]
- Kroiss, J.; Svatos, A.; Kaltenpoth, M. Rapid identification of insect cuticular hydrocarbons using Gas Chromatography-Ion-Trap Mass Spectrometry. J. Chem. Ecol. 2011, 37, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; McDaniel, C.A.; Nelson, D.R.; Blomquist, G.J. Chemical ionization mass spectrometry. Application to insect-derived cuticular alkanes. J. Chem. Ecol. 1980, 6, 609–623. [Google Scholar] [CrossRef]
- Moravcova, D.; Kahle, V.; Rehulkova, H.; Chmelik, J.; Rehulka, P. Short monolithic columns for purification and fractionation of peptide samples for matrix-assisted laser desorption/ionization time-of-flight/time-of-flight mass spectrometry analysis in proteomics. J. Chromatogr. A 2009, 1216, 3629–3636. [Google Scholar] [CrossRef]
- McLafferty, F.W. Registry of Mass Spectral Data, 5th ed.; Wiley: New York, NY, USA, 1989. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy; Allured: Carol Stream, IL, USA, 2007. [Google Scholar]
- Carlson, D.A.; Roan, C.S.; Yost, R.A.; Hector, J. Dimethyl disulfide derivatives of long-chain alkenes, alkadienes, and alkatrienes for gas-chromatography mass-spectrometry. Anal. Chem. 1989, 61, 1564–1571. [Google Scholar] [CrossRef]
- Oldham, N.J.; Svatos, A. Determination of the double bond position in functionalized monoenes by chemical ionization ion-trap mass spectrometry using acetonitrile as a reagent gas. Rapid Commun. Mass Spectrom. 1999, 13, 331–336. [Google Scholar] [CrossRef]
- Yew, J.Y.; Dreisewerd, K.; Luftmann, H.; Müthing, J.; Pohlentz, G.; Kravitz, E.A. A new male sex pheromone and novel cuticular cues for chemical communication in Drosophila. Curr. Biol. 2009, 19, 1245–1254. [Google Scholar] [CrossRef] [Green Version]
- Cvacka, J.; Svatos, A. Matrix-assisted laser desorption/ionization analysis of lipids and high molecular weight hydrocarbons with lithium 2,5-dihydroxybenzoate matrix. Rapid Commun. Mass Spectrom. 2003, 17, 2203–2207. [Google Scholar] [CrossRef]
- Vrkoslav, V.; Muck, A.; Cvacka, J.; Svatos, A. MALDI imaging of neutral cuticular lipids in insects and plants. J. Am. Soc. Mass Spectrom. 2010, 21, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Sivinski, J.M.; Calkins, C. Pheromones and parapheromones in the control of tephritids. Florida Entomol. 1986, 69, 157. [Google Scholar] [CrossRef]
- Romano, D.; Donati, E.; Benelli, G.; Stefanini, C. A review on animal–robot interaction: From bio-hybrid organisms to mixed societies. Biol. Cybern. 2019, 113, 201–225. [Google Scholar] [CrossRef] [PubMed]
- Halloy, J.; Sempo, G.; Caprari, G.; Rivault, C.; Asadpour, M.; Tache, F.; Said, I.; Durier, V.; Canonge, S.; Ame, J.M.; et al. Social integration of robots into groups of cockroaches to control self-organized choices. Science 2007, 318, 1155–1158. [Google Scholar] [CrossRef] [Green Version]
- Romano, D.; Benelli, G.; Kavallieratos, N.G.; Athanassiou, C.G.; Canale, A.; Stefanini, C. Beetle-robot hybrid interaction: Sex, lateralization and mating experience modulate behavioural responses to robotic cues in the larger grain borer Prostephanus truncatus (Horn). Biol. Cybern. 2020, 114, 473–483. [Google Scholar] [CrossRef]
- Romano, D.; Benelli, G.; Stefanini, C. Opposite valence social information provided by bio-robotic demonstrators shapes selection processes in the green bottle fly. J. R. Soc. Interface 2021, 18, rsif.2021.0056. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Katsoyannos, B.I.; Kouloussis, N.A.; Hendrichs, J.; Carey, J.R.; Heath, R.R. Early detection and population monitoring of Ceratitis capitata (Diptera: Tephritidae) in a mixed-fruit orchard in northern Greece. J. Econ. Entomol. 2001, 94, 971–978. [Google Scholar] [CrossRef]
- Suckling, D.M.; Kean, J.M.; Stringer, L.D.; Cáceres-Barrios, C.; Hendrichs, J.; Reyes-Flores, J.; Dominiak, B.C. Eradication of tephritid fruit fly pest populations: Outcomes and prospects. Pest Manag. Sci. 2016, 72, 456–465. [Google Scholar] [CrossRef]
- Casaña-Giner, V.; Levi, V.; Navarro-Llopis, V.; Jang, E.B. Implication of SAR of male medfly attractants in insect olfaction. SAR QSAR Environ. Res. 2002, 13, 629–640. [Google Scholar] [CrossRef]
- Jang, E.; Khrimian, A.; Holler, T. Field response of Mediterranean fruit flies to ceralure B1 relative to most active isomer and commercial formulation of trimedlure. J. Econ. Entomol. 2010. [Google Scholar] [CrossRef]
- Shelly, T.E.; Cowan, A.N.; Edu, J.; Pahio, E. Mating success of male Mediterranean fruit flies following exposure to two sources of α-copaene, manuka oil and mango. Florida Entomol. 2008, 91, 9–15. [Google Scholar] [CrossRef]
- Shelly, T.E.; Whittier, T.S.; Villalobos, E.M. Trimedlure affects mating success and mate attraction in male Mediterranean fruit flies. Entomol. Exp. Appl. 1996, 78, 181–185. [Google Scholar] [CrossRef]
- Papadopoulos, N.T.; Katsoyannos, B.I.; Kouloussis, N.A.; Hendrichs, J. Effect of orange peel substances on mating competitiveness of male Ceratitis capitata. Entomol. Exp. Appl. 2001, 99, 253–261. [Google Scholar] [CrossRef]
- Shelly, T.E.; Epsky, N.D. Exposure to tea tree oil enhances the mating success of male Mediterranean fruit flies (Diptera: Tephritidae). Florida Entomol. 2015, 98, 1127–1133. [Google Scholar] [CrossRef]
- Shelly, T.E. Exposure to α-copaene and α-copaene-containing oils enhances mating success of male Mediterranean fruit flies (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2001, 94, 497–502. [Google Scholar] [CrossRef]
- Kouloussis, N.A.; Gerofotis, C.D.; Ioannou, C.S.; Iliadis, I.V.; Papadopoulos, N.T.; Koveos, D.S. Towards improving sterile insect technique: Exposure to orange oil compounds increases sexual signalling and longevity in Ceratitis capitata males of the Vienna 8 GSS. PLoS ONE 2017, 12, e0188092. [Google Scholar] [CrossRef] [Green Version]
- Dean, D.; Pierre, H.; Mosser, L.; Kurashima, R.; Shelly, T. Field longevity and attractiveness of trimedlure plugs to male Ceratitis capitata in Florida and Hawaii. Florida Entomol. 2018, 101, 441–446. [Google Scholar] [CrossRef] [Green Version]
- (IAEA) International Atomic Energy Agency. Trapping Guidelines for Area-Wide Fruit Fly Programmes; IAEA: Vienna, Austria, 2003. [Google Scholar]
- Royer, J.E.; Agovaua, S.; Bokosou, J.; Kurika, K.; Mararuai, A.; Mayer, D.G.; Niangu, B. Responses of fruit flies (Diptera: Tephritidae) to new attractants in Papua New Guinea. Austral. Entomol. 2018, 57, 40–49. [Google Scholar] [CrossRef]
- Royer, J.E. Responses of fruit flies (Tephritidae: Dacinae) to novel male attractants in north Queensland, Australia, and improved lures for some pest species. Austral. Entomol. 2015, 54, 411–426. [Google Scholar] [CrossRef] [Green Version]
- Shelly, T.E.; Villalobos, E.M. Cue Lure and the mating behavior of male melon flies (Diptera: Tephritidae). Florida Entomol. 1995, 78, 473. [Google Scholar] [CrossRef]
- Shelly, T.E. Effects of methyl eugenol and raspberry ketone/cue lure on the sexual behavior of Bactrocera species (Diptera: Tephritidae). Appl. Entomol. Zool. 2010, 45, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.M.; Shuttleworth, L.A.; Osborne, T.; Collins, D.; Gurr, G.M.; Reynolds, O.L. Raspberry ketone accelerates sexual maturation and improves mating performance of sterile male Queensland fruit fly, Bactrocera tryoni (Froggatt). Pest Manag. Sci. 2019, 75, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Akter, H.; Mendez, V.; Morelli, R.; Pérez, J.; Taylor, P.W. Raspberry ketone supplement promotes early sexual maturation in male Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae). Pest Manag. Sci. 2017, 73, 1764–1770. [Google Scholar] [CrossRef] [PubMed]
- Wee, S.-L.; Clarke, A.R. Male-lure type, lure dosage, and fly age at feeding all influence male mating success in Jarvis’ fruit fly. Sci. Rep. 2020, 10, 15004. [Google Scholar] [CrossRef] [PubMed]
- Akter, H.; Pérez, J.; Park, S.J. Raspberry ketone supplements provided to immature male Queensland fruit fly, Bactrocera tryoni (Froggatt), increase the amount of volatiles in rectal glands. Chemoecology 2020, 31, 89–99. [Google Scholar] [CrossRef]
- Cunningham, R.T.; Suda, D.Y. Male annihilation through mass-trapping of male flies with methyleugenol to reduce infestation of Oriental fruit fly (Diptera: Tephritidae) larvae in papaya. J. Econ. Entomol. 1986, 79, 1580–1582. [Google Scholar] [CrossRef]
- Haq, I.U.; Cáceres, C.; Meza, J.S.; Hendrichs, J.; Vreysen, M.J.B. Different methods of methyl eugenol application enhance the mating success of male Oriental fruit fly (Dipera: Tephritidae). Sci. Rep. 2018, 8, 6033. [Google Scholar] [CrossRef] [PubMed]
- De Vincenzi, M.; Silano, M.; Stacchini, P.; Scazzocchio, B. Constituents of aromatic plants: I. Methyleugenol. Fitoterapia 2000, 71, 216–221. [Google Scholar] [CrossRef]
- Haniotakis, G.; Kozyrakis, M.; Fitsakis, T.; Antonidaki, A. An effective mass trapping method for the control of Dacus oleae (Diptera, tephritidae). J. Econ. Entomol. 1991, 84, 564–569. [Google Scholar] [CrossRef]
- Sarles, L.; Fassotte, B.; Boullis, A.; Lognay, G.; Verhaeghe, A.; Marko, I.; Verheggen, F.J. Improving the monitoring of the walnut husk fly (Diptera: Tephritidae) using male-produced lactones. J. Econ. Entomol. 2018, 111, 2032–2037. [Google Scholar] [CrossRef]
- Vargas, R.I.; Souder, S.K.; Hoffman, K.; Mercogliano, J.; Smith, T.R.; Hammond, J.; Davis, B.J.; Brodie, M.; Dripps, J.E. Attraction and mortality of Bactrocera dorsalis (Diptera: Tephritidae) to STATIC spinosad ME weathered under operational conditions in California and Florida: A reduced-risk male annihilation treatment. J. Econ. Entomol. 2014, 107, 1362–1369. [Google Scholar] [CrossRef] [Green Version]
- (CDFA) California Department of Food and Agriculture. Oriental Fruit Fly Fact Sheet; CDFA: Sacramento, CA, USA, 2008. [Google Scholar]
- Vargas, R.; Piñero, J.; Leblanc, L. An overview of pest species of Bactrocera fruit flies (Diptera: Tephritidae) and the integration of biopesticides with other biological approaches for their management with a focus on the Pacific region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, N.T.; Plant, R.E.; Carey, J.R. From trickle to flood: The large-scale, cryptic invasion of California by tropical fruit flies. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoukis, N.C.; Vargas, R.I.; Carvalho, L.; Fezza, T.; Wilson, S.; Collier, T.; Shelly, T.E. A field test on the effectiveness of male annihilation technique against Bactrocera dorsalis (Diptera: Tephritidae) at varying application densities. PLoS ONE 2019, 14, e0213337. [Google Scholar] [CrossRef] [Green Version]
- Manrakhan, A.; Venter, J.H.; Hattingh, V. The progressive invasion of Bactrocera dorsalis (Diptera: Tephritidae) in South Africa. Biol. Invasions 2015, 17, 2803–2809. [Google Scholar] [CrossRef]
- Manrakhan, A.; Hattingh, V.; Venter, J.-H.; Holtzhausen, M. Eradication of Bactrocera invadens (Diptera: Tephritidae) in Limpopo Province, South Africa. Afr. Entomol. 2011, 19, 650–659. [Google Scholar] [CrossRef]
- Steiner, L.F.; Hart, W.G.; Harris, E.J.; Cunningham, R.T.; Ohinata, K.; Kamakahi, D.C. Eradication of the Oriental fruit fly from the Mariana Islands by the methods of male annihilation and sterile insect release. J. Econ. Entomol. 1970, 63, 131–135. [Google Scholar] [CrossRef]
- Koyama, J.; Teruya, T.; Tanaka, K. Eradication of the Oriental fruit fly (Diptera: Tephritidae) from the Okinawa Islands by a male annihilation method. J. Econ. Entomol. 1984, 77, 468–472. [Google Scholar] [CrossRef]
- Seewooruthun, S.I.; Permalloo, S.; Gungah, B.; Soonnoo, A.R.; Alleck, M. Eradication of an exotic fruit fly from Mauritius. In Area-Wide Control of Fruit Flies and Other Insect Pests; Penerbit Universiti Sains Malaysia: Penang, Malaysia, 2000; pp. 389–394. [Google Scholar]
- Ekesi, S.; De Meyer, M.; Mohamed, S.A.; Virgilio, M.; Borgemeister, C. Taxonomy, ecology, and management of native and exotic fruit fly species in Africa. Annu. Rev. Entomol. 2016, 61, 219–238. [Google Scholar] [CrossRef]
- Tasnin, M.S.; Merkel, K.; Clarke, A.R. Effects of advanced age on olfactory response of male and female Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). J. Insect Physiol. 2020, 122, 104024. [Google Scholar] [CrossRef]
- Mau, R.F.L.; Jang, E.B.; Vargas, R. The Hawaii area-wide fruit fly management programme. In Area-Wide Control of Insect Pests from research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 671–683. [Google Scholar]
- Ballo, S.; Demissie, G.; Tefera, T.; Mohamed, S.A.; Khamis, F.M.; Niassy, S.; Ekesi, S. Use of para-pheromone methyl eugenol for suppression of the mango fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) in Southern Ethiopia. In Sustainability in Plant and Crop Protection; Springer: Cham, Switzerland, 2020; pp. 203–217. [Google Scholar]
- Speranza, S.; Bellocchi, G.; Pucci, C. IPM trials on attract-and-kill mixtures against the olive fly Bactrocera oleae (Diptera Tephritidae). Bull. Insectol. 2004, 57, 111–115. [Google Scholar]
- Broumas, T.; Haniotakis, G.; Liaropoulos, C.; Tomazou, T.; Ragoussis, N. The efficacy of an improved form of the mass-trapping method, for the control of the olive fruit fly, Bactrocera oleae (Gmelin) (Dipt., Tephritidae): Pilot-scale feasibility studies. J. Appl. Entomol. 2002, 126, 217–223. [Google Scholar] [CrossRef]
- Montiel, A.; Ramos, P.; Jones, O.T.; Lisk, J.C.; Howse, P.E.; Baker, R. Interferencias en el apareamiento de la mosca del olivo (Dacus oleae Gmel.) con el principal componente de su feromona sexual. Bol. Serv. Plagas 1982, 8, 193–200. [Google Scholar]
- Montiel, A.; Jones, O.T. Alternative methods for controlling the olive fly, Bactrocera oleae, involving semiochemicals. IOBC WPRS Bull. 2002, 25, 147–156. [Google Scholar]
- Navarro-Llopis, V.; Alfaro, C.; Primo, J.; Vacas, S. Response of two tephritid species, Bactrocera oleae and Ceratitis capitata, to different emission levels of pheromone and parapheromone. Crop Prot. 2011, 30, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Ferveur, J.F. Genetic control of pheromones in Drosophila simulans. I. Ngbo, a locus on the second chromosome. Genetics 1991, 128, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Gleason, J.M.; Jallon, J.M.; Rouault, J.D.; Ritchie, M.G. Quantitative trait loci for cuticular hydrocarbons associated with sexual isolation between Drosophila simulans and D. sechellia. Genetics 2005, 171, 1789–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liimatainen, J.O.; Jallon, J.M. Genetic analysis of cuticular hydrocarbons and their effect on courtship in Drosophila virilis and D. lummei. Behav. Genet. 2007, 37. [Google Scholar] [CrossRef]
- Snellings, Y.; Herrera, B.; Wildemann, B.; Beelen, M.; Zwarts, L.; Wenseleers, T.; Callaerts, P. The role of cuticular hydrocarbons in mate recognition in Drosophila suzukii. Sci Rep 2018, 8, 4996. [Google Scholar] [CrossRef]
- Leonhardt, B.A.; Rice, R.E.; Harte, E.M.; Cunningham, R.T. Evaluation of dispensers containing trimedlure, the attractant for the Mediterranean fruit fly (Diptera: Tephritidae). J. Econ. Entomol. 1984, 77, 744–749. [Google Scholar] [CrossRef]
- Domínguez-Ruiz, J.; Sanchis, J.; Navarro-Llopis, V.; Primo, J. A new long-life trimedlure dispenser for Mediterranean fruit fly. J. Econ. Entomol. 2008, 101, 1325–1330. [Google Scholar] [CrossRef]
- Cameron, D.N.S.; McRae, C.; Park, S.J.; Taylor, P.W.; Jamie, I.M. Vapor pressures and thermodynamic properties of phenylpropanoid and phenylbutanoid attractants of male Bactrocera, Dacus, and Zeugodacus fruit flies at ambient temperatures. J. Agric. Food Chem. 2020, 68, 9654–9663. [Google Scholar] [CrossRef] [PubMed]
- Lehman, K.A.; Barahona, D.C.; Manoukis, N.C.; Carvalho, L.A.F.N.; De Faveri, S.G.; Auth, J.E.; Siderhurst, M.S. Raspberry ketone trifluoroacetate trapping of Zeugodacus cucurbitae (Diptera: Tephritidae)in Hawaii. J. Econ. Entomol. 2019, 112, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Katsoyannos, B.; Boller, E. Second field application of oviposition-deterring pheromone of the European cherry fruit fly, Rhagoletis cerasi L. (Diptera: Tephritidae). Zeitschrift für Angew. Entomol. 1980, 89, 278–281. [Google Scholar] [CrossRef]
- Katsoyannos, B.I.; Boller, E.F. First field application of oviposition-deterring marking pheromone of European cherry fruit fly. Environ. Entomol. 1976, 5, 151–152. [Google Scholar] [CrossRef]
- Aluja, M.; Boller, E.F. Host marking pheromone of Rhagoletis cerasi: Foraging behavior in response to synthetic pheromonal isomers. J. Chem. Ecol. 1992, 18, 1299–1311. [Google Scholar] [CrossRef]
- Boller, E.; Hurter, J. The marking pheromone of the cherry fruit fly: A novel non-toxic and ecologically safe technique to protect cherries against cherry fruit fly infestation. In Proceedings of the Second International Symposium on Insect Pheromones; 1998. [Google Scholar]
- Prokopy, R.J. Epideitic pheromones that influence spacing patterns of phytophagous insects. In Semiochemicals: Their Role in Pest Control; Nordlund, D.A., Jones, R.L., Lewis, W.J., Eds.; Wiley Press: New York, NY, USA, 1981; pp. 181–213. [Google Scholar]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Birke, A.; Lopez-Ramirez, S.; Jimenez-Mendoza, R.; Acosta, E.; Ortega, R.; Edmunds, A.; Aluja, M. Host marking pheromone and GF120(TM) applied in a push-pull scheme reduce grapefruit infestation by Anastrepha ludens in field-cage studies. J. Pest Sci. 2020, 93, 507–518. [Google Scholar] [CrossRef]
- Cai, P.; Song, Y.; Huo, D.; Lin, J.; Zhang, H.; Zhang, Z.; Xiao, C.; Huang, F.; Ji, Q. Chemical cues induced from fly-oviposition mediate the host-seeking behaviour of Fopius arisanus (Hymenoptera: Braconidae), an effective egg parasitoid of Bactrocera dorsalis (Diptera: Tephritidae), within a tritrophic context. Insects 2020, 11, 231. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Revadi, S.; Carpita, A.; Giunti, G.; Raspi, A.; Anfora, G.; Canale, A. Behavioral and electrophysiological responses of the parasitic wasp Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae) to Ceratitis capitata-induced fruit volatiles. Biol. Control 2013, 64, 116–124. [Google Scholar] [CrossRef]
- Giunti, G.; Canale, A.; Messing, R.H.; Donati, E.; Stefanini, C.; Michaud, J.P.; Benelli, G. Parasitoid learning: Current knowledge and implications for biological control. Biol. Control 2015, 90, 208–219. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Rodriguez-Saona, C.; Meyer, W.L. Recognition of foreign oviposition-marking pheromone in a multi-trophic context. Naturwissenschaften 2009, 96, 585–592. [Google Scholar] [CrossRef]
- Faraone, N.; Hillier, N.K.; Cutler, G.C. Collection of host-marking pheromone from Rhagoletis mendax (Diptera: Tephritidae). Can. Entomol. 2016, 148, 552–555. [Google Scholar] [CrossRef]
- Li, H.; Ren, L.; Xie, M.; Gao, Y.; He, M.; Hassan, B.; Lu, Y.; Cheng, D. Egg-surface bacteria are indirectly associated with oviposition aversion in Bactrocera dorsalis. Curr. Biol. 2020, 30, 4432–4440. [Google Scholar] [CrossRef]
- Calcagnile, M.; Tredici, S.M.; Talà, A.; Alifano, P. Bacterial semiochemicals and transkingdom interactions with insects and plants. Insects 2019, 10, 441. [Google Scholar] [CrossRef] [Green Version]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Gurr, G.M.; Kvedaras, O.L. Synergizing biological control: Scope for sterile insect technique, induced plant defences and cultural techniques to enhance natural enemy impact. Biol. Control 2010, 52, 198–207. [Google Scholar] [CrossRef]
- Whittier, T.S.; Nam, F.Y.; Shelly, T.E.; Kaneshiro, K.Y. Male courtship success and female discrimination in the mediterranean fruit fly (Diptera: Tephritidae). J. Insect Behav. 1994, 7, 159–170. [Google Scholar] [CrossRef]
- Pereira, R.; Yuval, B.; Liedo, P.; Teal, P.E.A.; Shelly, T.E.; McInnis, D.O.; Hendrichs, J. Improving sterile male performance in support of programmes integrating the sterile insect technique against fruit flies. J. Appl. Entomol. 2013, 137, 178–190. [Google Scholar] [CrossRef]
- Segura, D.F.; Belliard, S.A.; Vera, M.T.; Bachmann, G.E.; Ruiz, M.J.; Jofre-Barud, F.; Fernández, P.C.; López, M.L.; Shelly, T.E. Plant chemicals and the sexual behavior of male tephritid fruit flies. Ann. Entomol. Soc. Am. 2018, 111, 239–264. [Google Scholar] [CrossRef]
- Vargas, R.I.; Shelly, T.E.; Leblanc, L.; Piñero, J.C. Recent advances in methyl eugenol and Cue-Lure technologies for fruit fly detection, monitoring, and control in Hawaii. In Vitamins and Hormones; Elsevier: Amsterdam, The Netherlands, 2010; pp. 575–595. [Google Scholar]
- Khan, M.A.M.; Manoukis, N.C.; Osborne, T.; Barchia, I.M.; Gurr, G.M.; Reynolds, O.L. Semiochemical mediated enhancement of males to complement sterile insect technique in management of the tephritid pest Bactrocera tryoni (Froggatt). Sci. Rep. 2017, 7, 13366. [Google Scholar] [CrossRef] [Green Version]
- Wehrenfennig, C.; Schott, M.; Gasch, T.; During, R.A.; Vilcinskas, A.; Kohl, C.D. On-site airborne pheromone sensing. Anal. Bioanal. Chem. 2013, 405, 6389–6403. [Google Scholar] [CrossRef]
- Fernandez-Grandon, G.M.; Girling, R.D.; Poppy, G.M. Utilizing insect behavior in chemical detection by a behavioral biosensor. J. Plant Interact. 2011, 6, 109–112. [Google Scholar] [CrossRef]
- Schroth, P.; Schoning, M.J.; Schutz, S.; Malkoc, U.; Steffen, A.; Marso, M.; Hummel, H.E.; Kordos, P.; Luth, H. Coupling of insect antennae to field-effect transistors for biochemical sensing. Electrochim. Acta 1999, 44, 3821–3826. [Google Scholar] [CrossRef]
- Repasky, K.S.; Shaw, J.A.; Scheppele, R.; Melton, C.; Carsten, J.L.; Spangler, L.H. Optical detection of honeybees by use of wing-beat modulation of scattered laser light for locating explosives and land mines. Appl. Opt. 2006, 45, 1839–1843. [Google Scholar] [CrossRef] [Green Version]
- Tomberlin, J.K.; Tertuliano, M.; Rains, G.; Lewis, W.J. Conditioned Microplitis croceipes Cresson (Hymenoptera: Braconidae) detect and respond to 2,4-DNT: Development of a biological sensor. J. Forensic. Sci. 2005, 50, 1187–1190. [Google Scholar] [CrossRef]
- Tertuliano, M.; Tomberlin, J.K.; Jurjevic, Z.; Wilson, D.; Rains, G.C.; Lewis, W.J. The ability of conditioned Microplitis croceipes (Hymenoptera: Braconidae) to distinguish between odors of aflatoxigenic and non-aflatoxigenic fungal strains. Chemoecology 2005, 15, 89–95. [Google Scholar] [CrossRef]
- Carey, A.F.; Carlson, J.R. Insect olfaction from model systems to disease control. Proc. Natl. Acad. Sci. USA 2011, 108, 12987–12995. [Google Scholar] [CrossRef] [Green Version]
- Strauch, M.; Lüdke, A.; Münch, D.; Laudes, T.; Galizia, C.G.; Martinelli, E.; Lavra, L.; Paolesse, R.; Ulivieri, A.; Catini, A.; et al. More than apples and oranges—Detecting cancer with a fruit fly’s antenna. Sci. Rep. 2015, 4, 3576. [Google Scholar] [CrossRef] [Green Version]
- Pelosi, P.; Mastrogiacomo, R.; Iovinella, I.; Tuccori, E.; Persaud, K.C. Structure and biotechnological applications of odorant-binding proteins. Appl. Microbiol. Biotechnol. 2014, 98, 61–70. [Google Scholar] [CrossRef]
- Lu, Y.L.; Li, H.L.; Zhuang, S.L.; Zhang, D.M.; Zhang, Q.; Zhou, J.; Dong, S.Y.; Liu, Q.J.; Wang, P. Olfactory biosensor using odorant-binding proteins from honeybee: Ligands of floral odors and pheromones detection by electrochemical impedance. Sensors Actuators B Chem. 2014, 193, 420–427. [Google Scholar] [CrossRef]
- Sankaran, S.; Panigrahi, S.; Mallik, S. Odorant binding protein based biomimetic sensors for detection of alcohols associated with Salmonella contamination in packaged beef. Biosens. Bioelectron. 2011, 26, 3103–3109. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.F.; Kim, S.N.; Crookes-Goodson, W.J.; Farmer, B.L.; Naik, R.R. Biomimetic chemosensor: Designing peptide recognition elements for surface functionalization of carbon nanotube field effect transistors. ACS Nano 2010, 4, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Kotlowski, C.; Larisika, M.; Guerin, P.M.; Kleber, C.; Krober, T.; Mastrogiacomo, R.; Nowak, C.; Pelosi, P.; Schutz, S.; Schwaighofer, A.; et al. Fine discrimination of volatile compounds by graphene-immobilized odorant-binding proteins. Sensors Actuators B Chem. 2018, 256, 564–572. [Google Scholar] [CrossRef]
- Lu, Y.L.; Yao, Y.; Li, S.; Zhang, Q.; Liu, Q.J. Olfactory biosensor based on odorant-binding proteins of Bactrocera dorsalis with electrochemical impedance sensing for pest management. Sens. Rev. 2017, 37, 396–403. [Google Scholar] [CrossRef]
- Bohbot, J.D.; Dickens, J.C. Characterization of an enantioselective odorant receptor in the yellow fever mosquito Aedes aegypti. PLoS ONE 2009, 4, e7032. [Google Scholar] [CrossRef] [Green Version]
- Stensmyr, M.C.; Dweck, H.K.M.; Farhan, A.; Ibba, I.; Strutz, A.; Mukunda, L.; Linz, J.; Grabe, V.; Steck, K.; Lavista-Llanos, S.; et al. A conserved dedicated olfactory circuit for detecting harmful microbes in Drosophila. Cell 2012, 151, 1345–1357. [Google Scholar] [CrossRef] [Green Version]
- Bohbot, J.D.; Vernick, S. The emergence of insect Odorant Receptor-based biosensors. Biosensors 2020, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Moitra, P.; Bhagat, D.; Kamble, V.B.; Umarji, A.M.; Pratap, R.; Bhattacharya, S. First example of engineered β-cyclodextrinylated MEMS devices for volatile pheromone sensing of olive fruit pests. Biosens. Bioelectron. 2021, 173, 112728. [Google Scholar] [CrossRef]
Figure 1.
Worldwide distribution of species belonging to the Anastrepha, Bactrocera, Ceratitis, Rhagoletis and Zeugodacus genera. The map is based on the currently available distribution data of species belonging to each genus retrieved from the EPPO Global Database (https://gd.eppo.int) (accessed on 13 November 2020) and integrated with information from CABI (https://www.cabi.org/isc/) (accessed on 13 November 2020) and the literature. In brackets, the total number of living species (obtained from Catalogue of Life: 2019 Annual Checklist [40] and [39]) for each genus and the original geographic range are reported. Lists of the major pests within each genus are also indicated. Background map was retrieved from https://freevectormaps.com/world-maps/WRLD-EPS-01-0011?ref=atr (accessed on 29 July 2020). Information about the status of invasion is reported in more details for the species A. ludens, B. dorsalis and C. capitata, for which yellow and green dots indicate transient presence or achieved eradication, respectively.
Figure 1.
Worldwide distribution of species belonging to the Anastrepha, Bactrocera, Ceratitis, Rhagoletis and Zeugodacus genera. The map is based on the currently available distribution data of species belonging to each genus retrieved from the EPPO Global Database (https://gd.eppo.int) (accessed on 13 November 2020) and integrated with information from CABI (https://www.cabi.org/isc/) (accessed on 13 November 2020) and the literature. In brackets, the total number of living species (obtained from Catalogue of Life: 2019 Annual Checklist [40] and [39]) for each genus and the original geographic range are reported. Lists of the major pests within each genus are also indicated. Background map was retrieved from https://freevectormaps.com/world-maps/WRLD-EPS-01-0011?ref=atr (accessed on 29 July 2020). Information about the status of invasion is reported in more details for the species A. ludens, B. dorsalis and C. capitata, for which yellow and green dots indicate transient presence or achieved eradication, respectively.

Figure 2.
Diagram showing the main steps of mating behaviour in tephritids and the signals transmitted and perceived. Lekking behaviour is common in Tephritidae, but notably absent in Rhagoletis species. Similarly, pre-mating trophallaxis is common, but this phenomenon is known to occur also during and post mating [47]. HMP, host-marking pheromone.
Figure 2.
Diagram showing the main steps of mating behaviour in tephritids and the signals transmitted and perceived. Lekking behaviour is common in Tephritidae, but notably absent in Rhagoletis species. Similarly, pre-mating trophallaxis is common, but this phenomenon is known to occur also during and post mating [47]. HMP, host-marking pheromone.

Figure 3.
Tissues involved in semiochemicals’ production in tephritids. Diagrammatic representation of a male (left) and female (right) generalized tephritid showing the tissues/body compartments involved in pheromone production. Reproductive tracts and rectum are shown in upper left (male) and right (female) boxes. Bottom left and right boxes show the abdomen of a calling male, showing the expanded rectal epithelium (pae, pleural abdominal expansion), and the ovipositor of a female. Abbreviations: Male reproductive system: tes (testes), ves (vas deferens), acg (accessory glands), ejd (ejaculatory duct), eja (ejaculatory apodeme), aed (aedeagus). Female reproductive system: ova (ovaries), ovr (ovariole), ovd (oviduct), spm (spermathecae), acg (accessory glands), cgd (colleterial gland duct), ovp (ovipositor). Rectum: rev (rectal valve), rep (rectal papilla), res (reservoir containing pheromone), ant (anal tube).
Figure 3.
Tissues involved in semiochemicals’ production in tephritids. Diagrammatic representation of a male (left) and female (right) generalized tephritid showing the tissues/body compartments involved in pheromone production. Reproductive tracts and rectum are shown in upper left (male) and right (female) boxes. Bottom left and right boxes show the abdomen of a calling male, showing the expanded rectal epithelium (pae, pleural abdominal expansion), and the ovipositor of a female. Abbreviations: Male reproductive system: tes (testes), ves (vas deferens), acg (accessory glands), ejd (ejaculatory duct), eja (ejaculatory apodeme), aed (aedeagus). Female reproductive system: ova (ovaries), ovr (ovariole), ovd (oviduct), spm (spermathecae), acg (accessory glands), cgd (colleterial gland duct), ovp (ovipositor). Rectum: rev (rectal valve), rep (rectal papilla), res (reservoir containing pheromone), ant (anal tube).

Figure 4.
Overlap among chemicals identified in the headspace emissions of tephritids. Venn diagrams (http://bioinformatics.psb.ugent.be/webtools/Venn/) (accessed on 10 March 2021) illustrating the number of common and unique chemicals found among the compared species in the (A) Ceratitis, (B) Anastrepha, and (C) Bactrocera genera. (D) For the genus Zeugodacus, the number of compounds emitted by Z. cucurbitae are reported. Pie charts represent the proportion in chemical classes of the total number of compounds (in parenthesis) isolated for each species.
Figure 4.
Overlap among chemicals identified in the headspace emissions of tephritids. Venn diagrams (http://bioinformatics.psb.ugent.be/webtools/Venn/) (accessed on 10 March 2021) illustrating the number of common and unique chemicals found among the compared species in the (A) Ceratitis, (B) Anastrepha, and (C) Bactrocera genera. (D) For the genus Zeugodacus, the number of compounds emitted by Z. cucurbitae are reported. Pie charts represent the proportion in chemical classes of the total number of compounds (in parenthesis) isolated for each species.

Figure 5.
Comparison among the chemicals identified in the headspace emissions of monophagous and oligophagous tephritid species. UpSet plot showing unique and overlapping chemicals across five species belonging to the Zeugodacus, Bactrocera, Anastrepha and Rhagoletis genera. The intersection matrix is sorted in descending order. Connected dots represent intersections of overlapping chemicals and horizontal bars show the total number of compounds identified in each species headspace. The plot was generated using the UpSetR package in R [255].
Figure 5.
Comparison among the chemicals identified in the headspace emissions of monophagous and oligophagous tephritid species. UpSet plot showing unique and overlapping chemicals across five species belonging to the Zeugodacus, Bactrocera, Anastrepha and Rhagoletis genera. The intersection matrix is sorted in descending order. Connected dots represent intersections of overlapping chemicals and horizontal bars show the total number of compounds identified in each species headspace. The plot was generated using the UpSetR package in R [255].

Figure 6.
Comparison among the chemicals identified in the headspace emissions of polyphagous tephritid species. UpSet plot showing unique and overlapping chemicals across five species belonging to the Bactrocera, Anastrepha and Ceratitis genera. The intersection matrix is sorted in descending order. Different genera are indicated in different colors. Connected dots represent intersections of overlapping chemicals and horizontal bars show the total number of compounds identified in each species headspace. The plot was generated using the UpSetR package in R [255].
Figure 6.
Comparison among the chemicals identified in the headspace emissions of polyphagous tephritid species. UpSet plot showing unique and overlapping chemicals across five species belonging to the Bactrocera, Anastrepha and Ceratitis genera. The intersection matrix is sorted in descending order. Different genera are indicated in different colors. Connected dots represent intersections of overlapping chemicals and horizontal bars show the total number of compounds identified in each species headspace. The plot was generated using the UpSetR package in R [255].


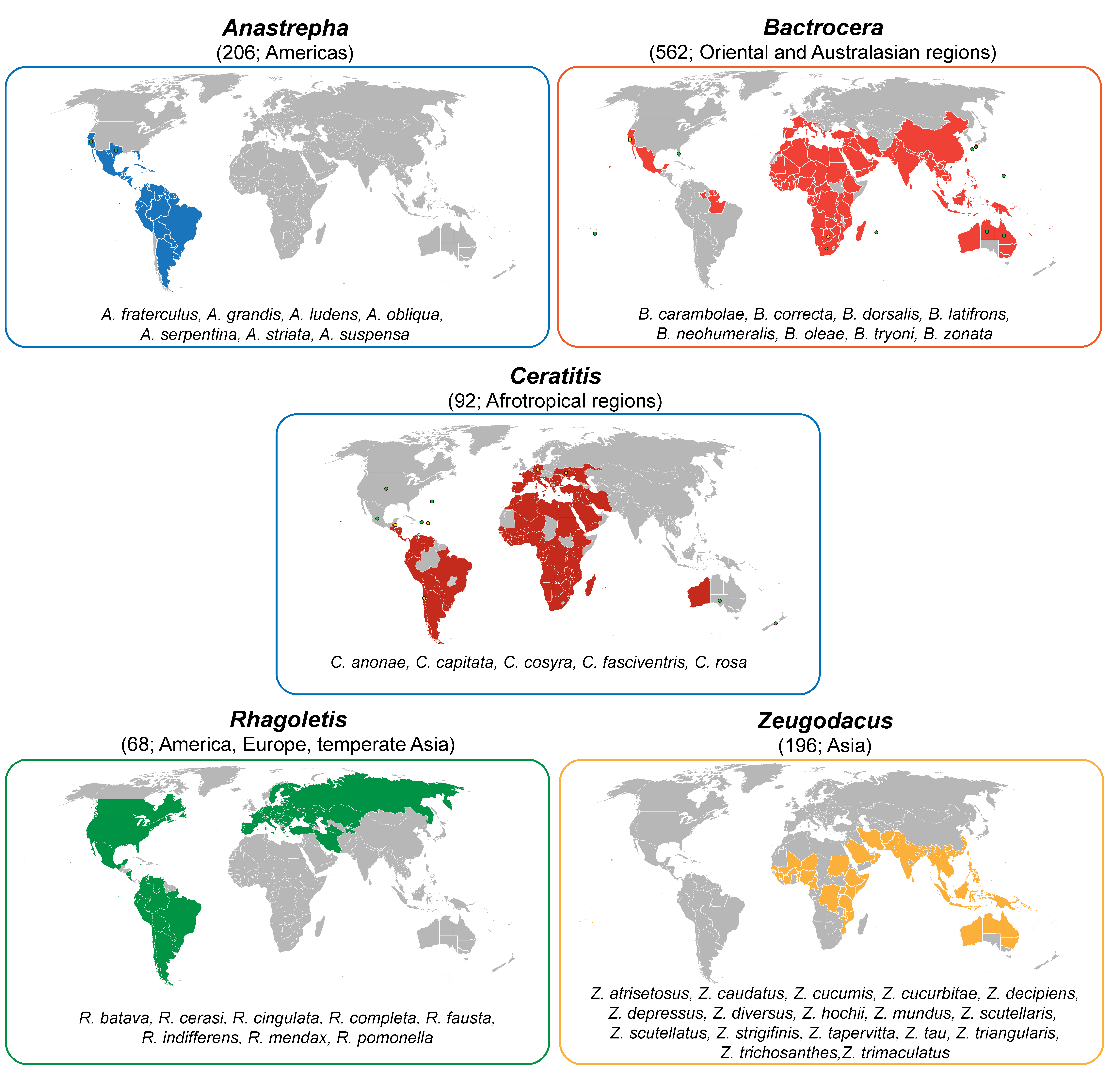{kind=link}
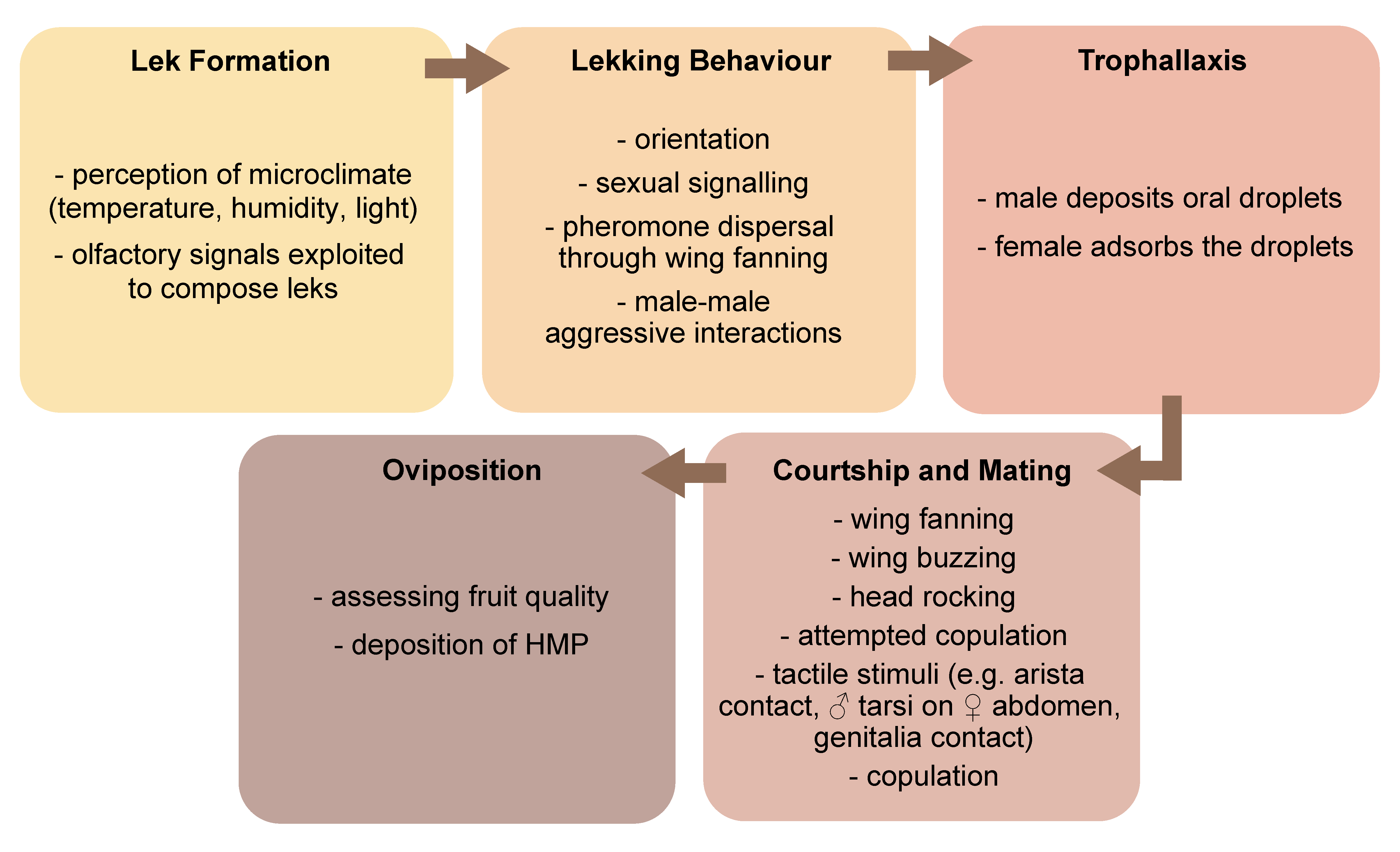{kind=link}
{kind=link}
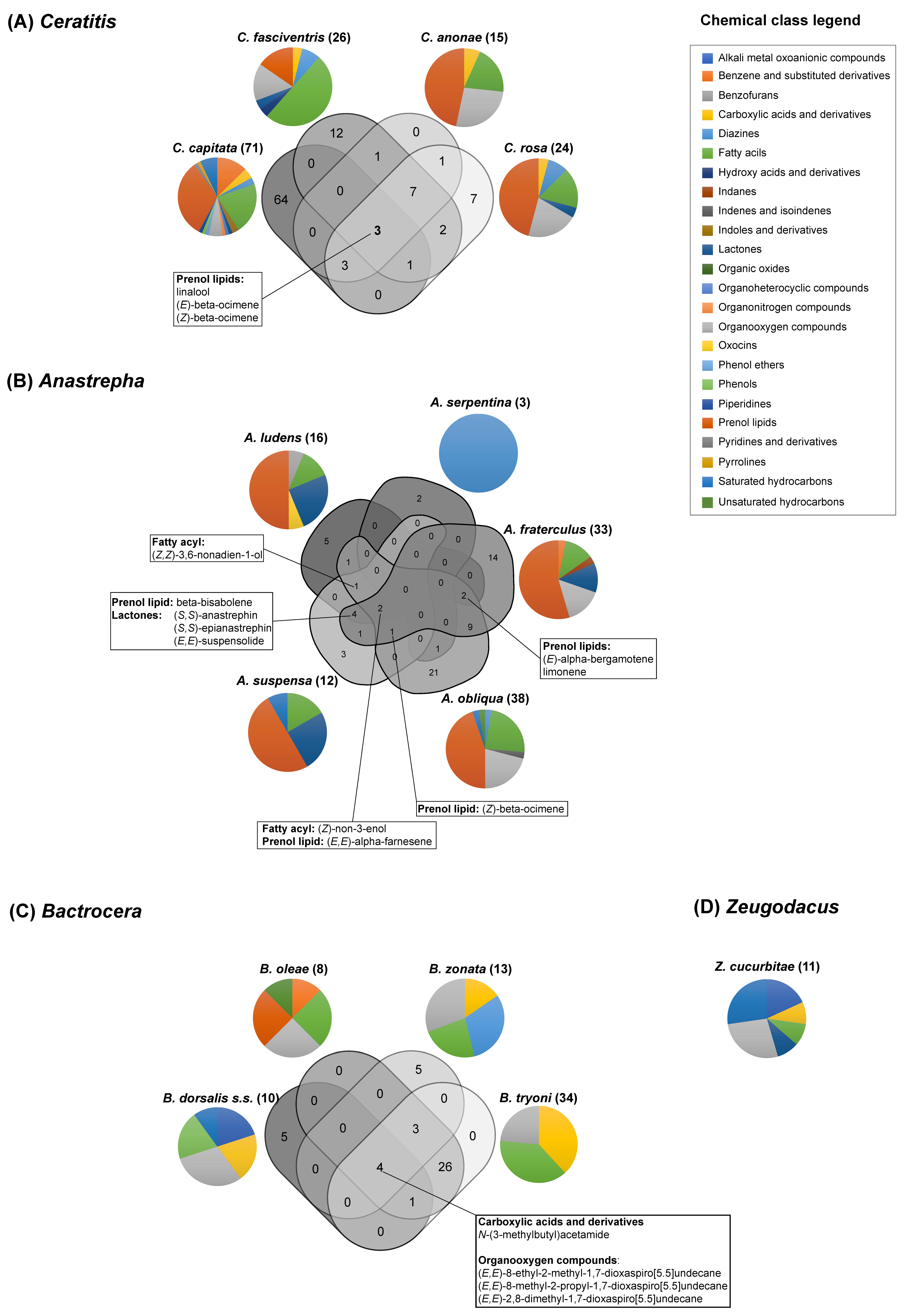{kind=link}
{kind=link}
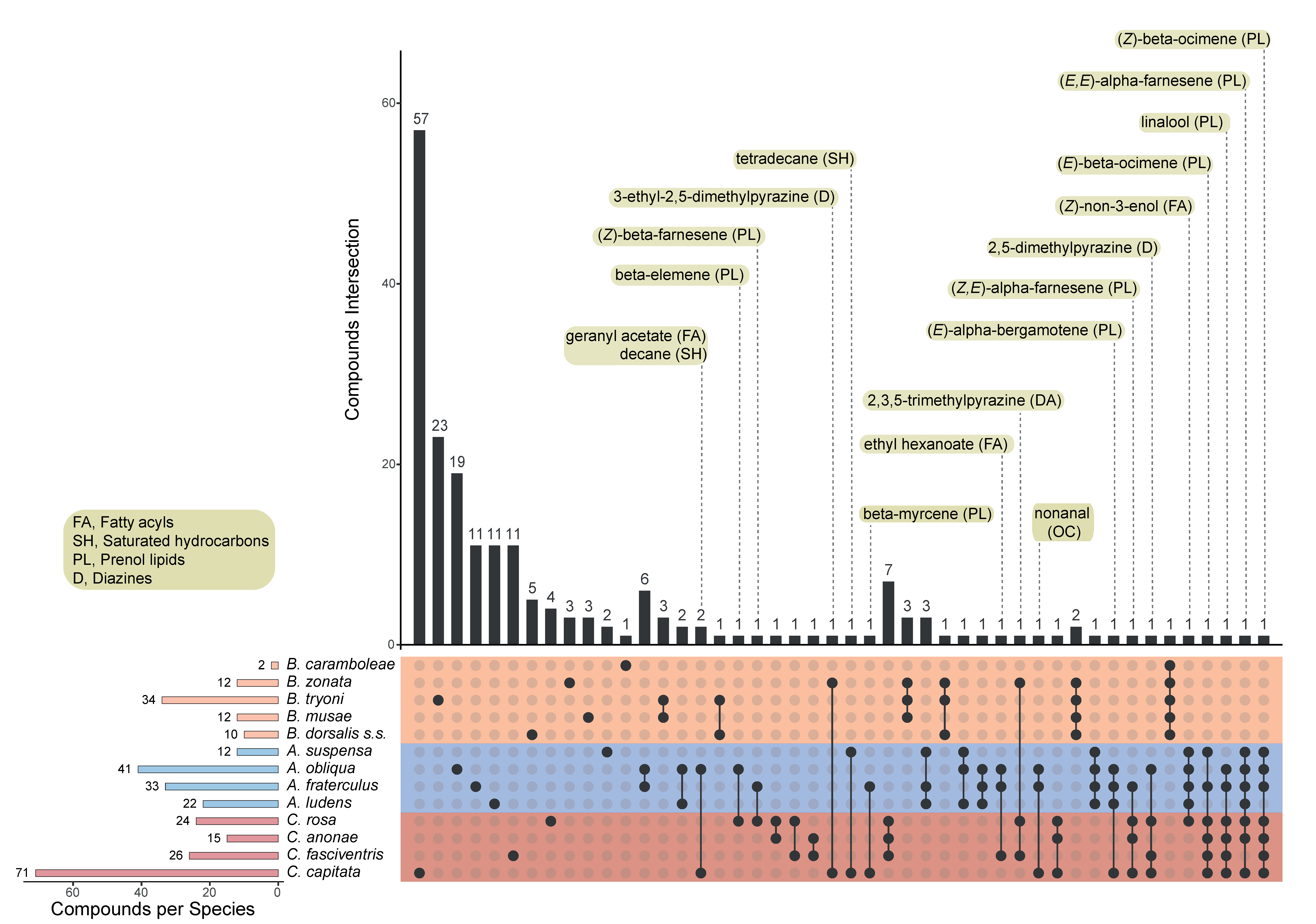{kind=link}
Table 1.
List of tephritid species for which volatile pheromone has been chemically analysed from the headspace.
Table 1.
List of tephritid species for which volatile pheromone has been chemically analysed from the headspace.
| Genus | Species | Male/Female-Borne | References |
|---|---|---|---|
| Anastrepha | A. fraterculus | Male | [175,176,177,178,179] |
| A. ludens | Male | [87,180,181,182,183,184,185,186] | |
| A. obliqua | Male | [187,188,189,190,191,192] | |
| A. serpentina | Male | [193] | |
| A. suspensa | Male | [149,180,185,194,195,196,197,198,199,200,201,202,203] | |
| Bactrocera | B. carambolae | Male | [72] |
| B. dorsalis s.s. | Male/Female | [68,204,205,206] | |
| B. musae | Male/Female | [76] | |
| B. oleae | Male/Female | [80,174,207,208] | |
| B. tryoni | Male/Female | [209,210,211,212] | |
| B. zonata | Male/Female | [213] | |
| Zeugodacus | Z. cucurbitae | Male/Female | [204,205] |
| Ceratitis | C. anonae | Male | [214] |
| C. capitata | Male | [166,173,215,216,217,218,219,220,221,222,223,224,225] | |
| C. fasciventris | Male | [214] | |
| C. rosa | Male | [214] | |
| Rhagoletis | R. batava | Male | [61] |
| R. cerasi | Male | [226] |
Table 2.
List of tephritid species for which pheromones have been derived from rectal glands extracts.
Table 2.
List of tephritid species for which pheromones have been derived from rectal glands extracts.
| Genus | Species | Sex | References |
|---|---|---|---|
| Anastrepha | A. fraterculus | Male * | [227] |
| A. ludens | Male * | [182] | |
| A. ludens | Male * | [228] | |
| A. ludens | Male * | [183] | |
| A. ludens | Male * | [185] | |
| A. ludens | Male * | [184] | |
| A. suspensa | Male * | [203] | |
| Bactrocera | B. albistrigata | Male | [229] |
| B. cacuminata | Male | [230] | |
| B. carambolae | Male | [72] | |
| B. correcta | Male/Female | [231] | |
| B. correcta | Male | [232] | |
| B. distincta | Male | [230] | |
| B. dorsalis | Male | [68] | |
| B. dorsalis | Male | [229] | |
| B. facialis | Male | [233] | |
| B. halfordiae | Male | [234] | |
| B. kirki | Male | [233] | |
| B. kraussi | Male | [233] | |
| B. latifrons | Male | [234] | |
| B. musae | Male/Female | [76] | |
| B. neohumeralis | Male | [209] | |
| B. nigrotibialis | Male | [229] | |
| B. occipitalis | Male | [234] | |
| B. oleae | Male/Female | [79] | |
| B. oleae | Male/Female | [85] | |
| B. oleae | Female | [230] | |
| B. oleae | Male/Female | [85] | |
| B. oleae | Male | [235] | |
| B. oleae | Female | [174] | |
| B. passiflorae | Male | [233] | |
| B. tryoni | Male | [209] | |
| B. tryoni | Female * | [210] | |
| B. tryoni | Female | [211] | |
| B. tryoni | Male | [212] | |
| B. tryoni | Male/Female | [236] | |
| B. umbrosa | Male | [237] | |
| B. xanthodes | Male | [233] | |
| Zeugodacus | Z. cucumis | Male | [234] |
| Z. cucurbitae | Male | [238] | |
| Z. cucurbitae | Male | [239] | |
| Z. cucurbitae | Male | [237] | |
| Z. tau | Male | [237] | |
| Ceratitis | - | - | - |
| Rhagoletis | - | - | - |
* abdominal extracts.
Table 3.
List of tephritid species for which host-marking behaviour has been identified.
| Genus | Species | Chemical Identity | References |
|---|---|---|---|
| Anastrepha | A. suspensa | - | [376] |
| A. sororcula | - | [377] | |
| A. fraterculus | - | [378] | |
| A. pseudoparallela | - | [379] | |
| A. bistrigata | - | [380] | |
| A. grandis | - | [381] | |
| A. ludens | 2-(2,14-Dimethylpentadecanoylamino)pentane-dioic acid | [123,382] | |
| A. striata | - | [97] | |
| A. obliqua | - | [132] | |
| A. serpentina | - | [132] | |
| Bactrocera | - | - | - |
| Ceratitis | C. capitata | - | [111,126] |
| C. cosyra | Glutathione | [33] | |
| C. rosa | Glutamic acid | [34] | |
| Rhagoletis | R. pomonella | - | [112,374,383,384] |
| R. cerasi | N-[15(β-Glucopyranosyl)-oxy-8-hydroxypalmitoyl]-taurine | [30,385,386] | |
| R. completa | - | [135] | |
| R. fausta | - | [387] | |
| R. cingulata | - | [388] | |
| R. cornivora | - | [388] | |
| R. indifferens | - | [388] | |
| R. mendax | - | [388] | |
| R. tabellaria | - | [388] | |
| R. basiola | - | [130] | |
| R. zephyria | - | [131] | |
| R. alternata | - | [389] | |
| Zeugodacus | - | - | - |
Table 4.
List of tephritid species for which CHs have been characterised.
| Genus | Species | Developmental Stage | References |
|---|---|---|---|
| Anastrepha | A. ludens | larvae and adults | [414,415] |
| A. suspensa | larvae and adults | [414,416,417,418] | |
| A. fraterculus | larvae and adults | [90,93,289,416,417,419] | |
| A. acris | larvae | [416,417] | |
| A. obliqua | larvae and adults | [417] | |
| A. serpentina | larvae | [417] | |
| A. pickeli | larvae | [417] | |
| A. striata | larvae | [417] | |
| Bactrocera | B. dorsalis | larvae and adults | [414,420,421] |
| B. carambolae | adults | [421] | |
| B. oleae | adults | [422] | |
| B. tryoni | adults | [423] | |
| B. zonata | adults | [422] | |
| Zeugodacus | Z. cucurbitae | larvae and adults | [205,414] |
| Ceratitis | C. capitata | larvae and adults | [274,289,414,418] |
| C. fasciventris | [274,289] | ||
| C. anonae | [274,289] | ||
| C. rosa | larvae and adults | [92,274,289,414] | |
| Rhagoletis | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Scolari, F.; Valerio, F.; Benelli, G.; Papadopoulos, N.T.; Vaníčková, L. Tephritid Fruit Fly Semiochemicals: Current Knowledge and Future Perspectives. Insects 2021, 12, 408. https://doi.org/10.3390/insects12050408
AMA Style
Scolari F, Valerio F, Benelli G, Papadopoulos NT, Vaníčková L. Tephritid Fruit Fly Semiochemicals: Current Knowledge and Future Perspectives. Insects. 2021; 12(5):408. https://doi.org/10.3390/insects12050408
Chicago/Turabian StyleScolari, Francesca, Federica Valerio, Giovanni Benelli, Nikos T. Papadopoulos, and Lucie Vaníčková. 2021. "Tephritid Fruit Fly Semiochemicals: Current Knowledge and Future Perspectives" Insects 12, no. 5: 408. https://doi.org/10.3390/insects12050408
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.