First Insights on Early Host Plants and Dispersal Behavior of Halyomorpha halys (Hemiptera: Pentatomidae) from Overwintering to Crop Colonization

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sites and Study Design
2.2. Sampling by Pheromone Traps
2.3. Sampling on Host Plants
2.4. Immunomarking-Capture Technique
3. Results
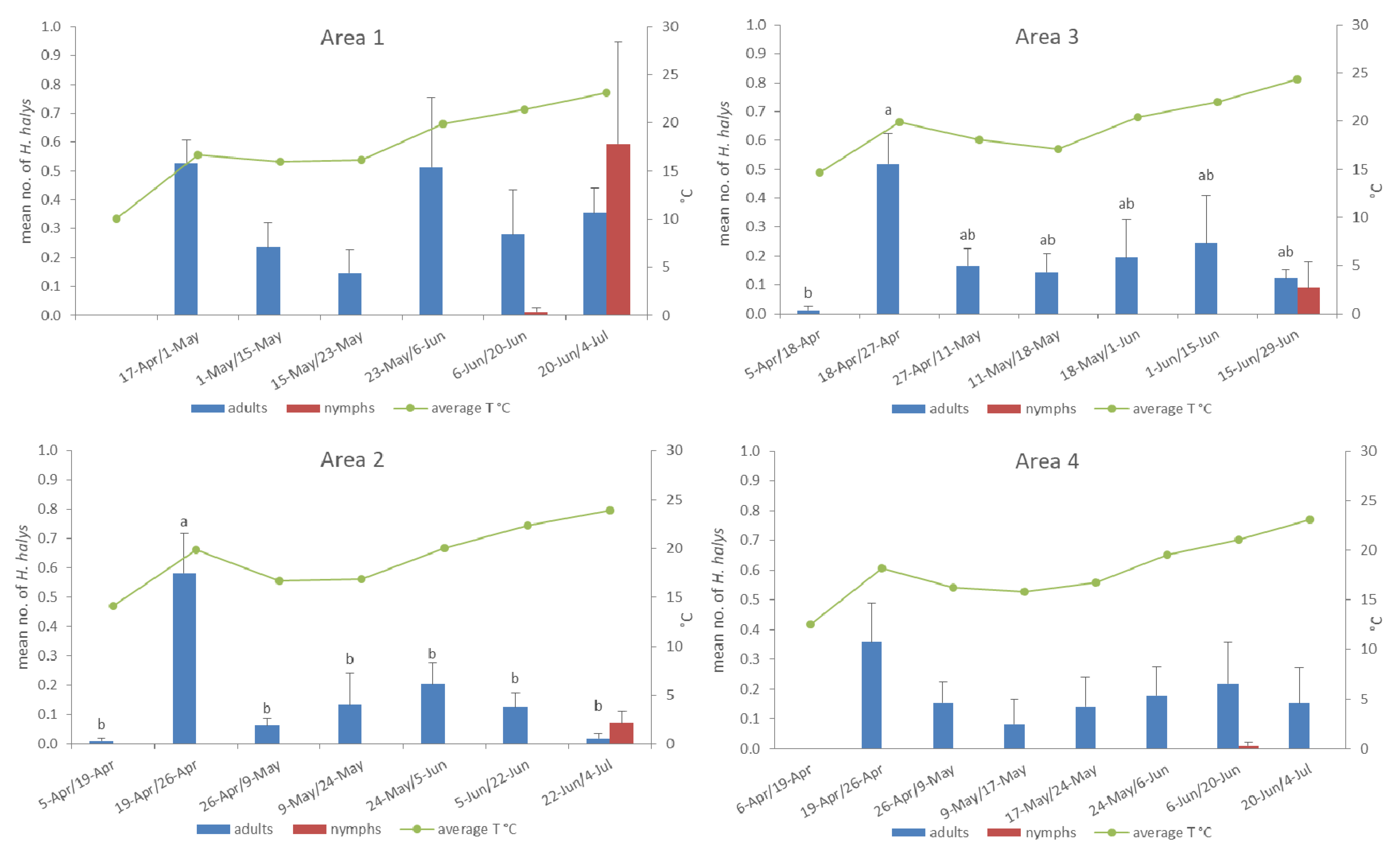
3.1. Population Dynamics after Overwintering Assessed by Pheromone Traps
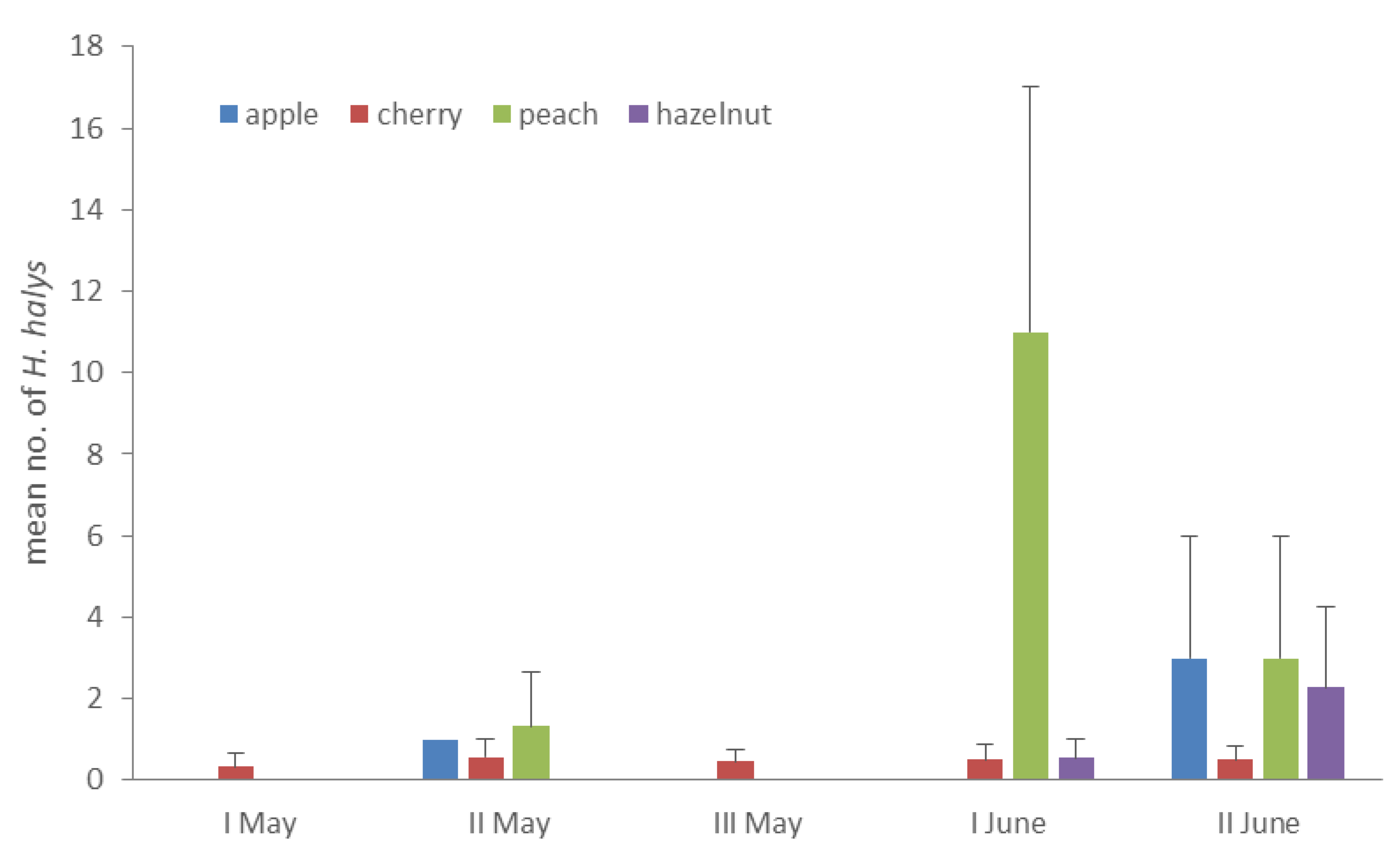
3.2. Early Colonization and Abundance on Host Plants
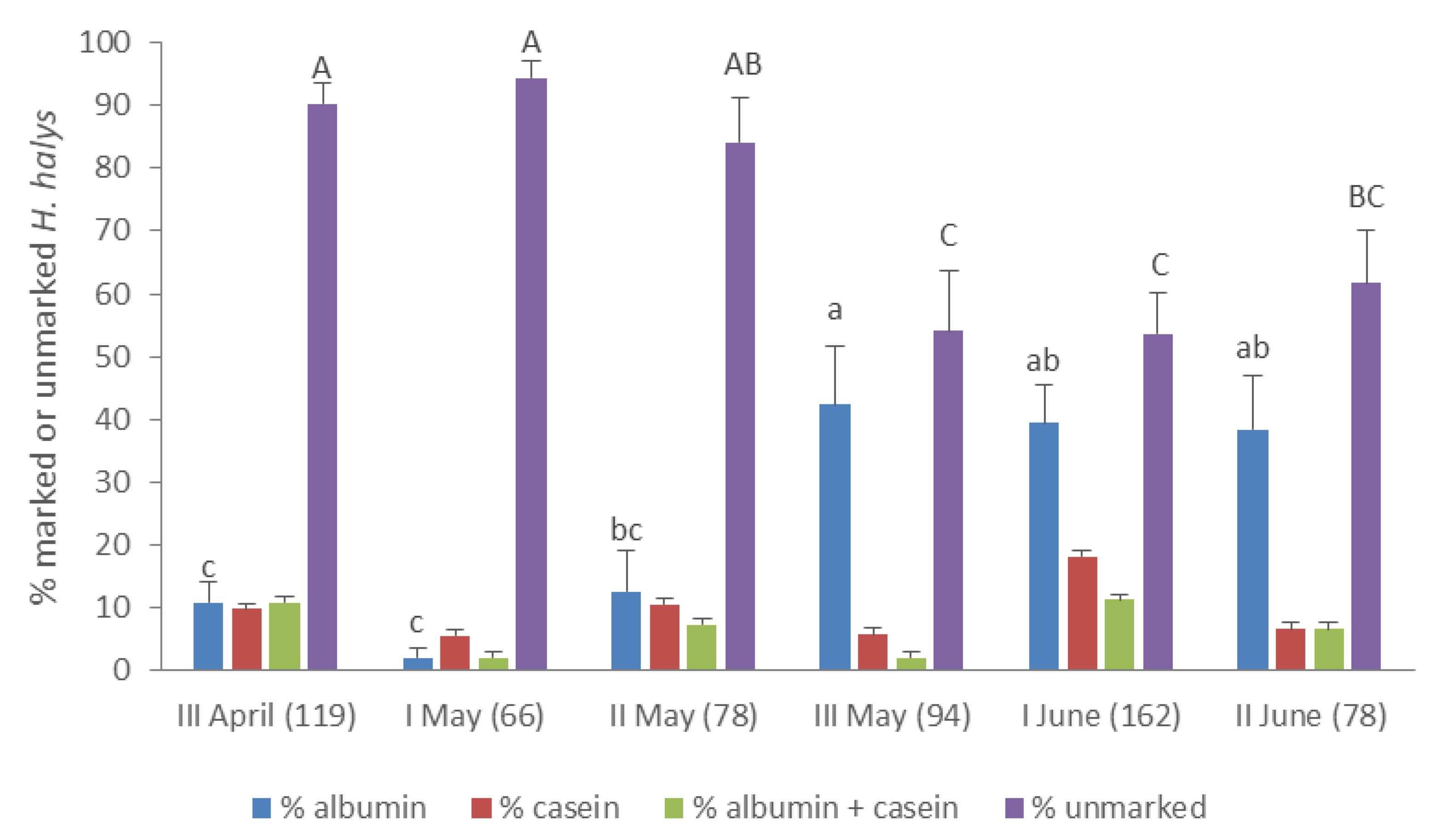
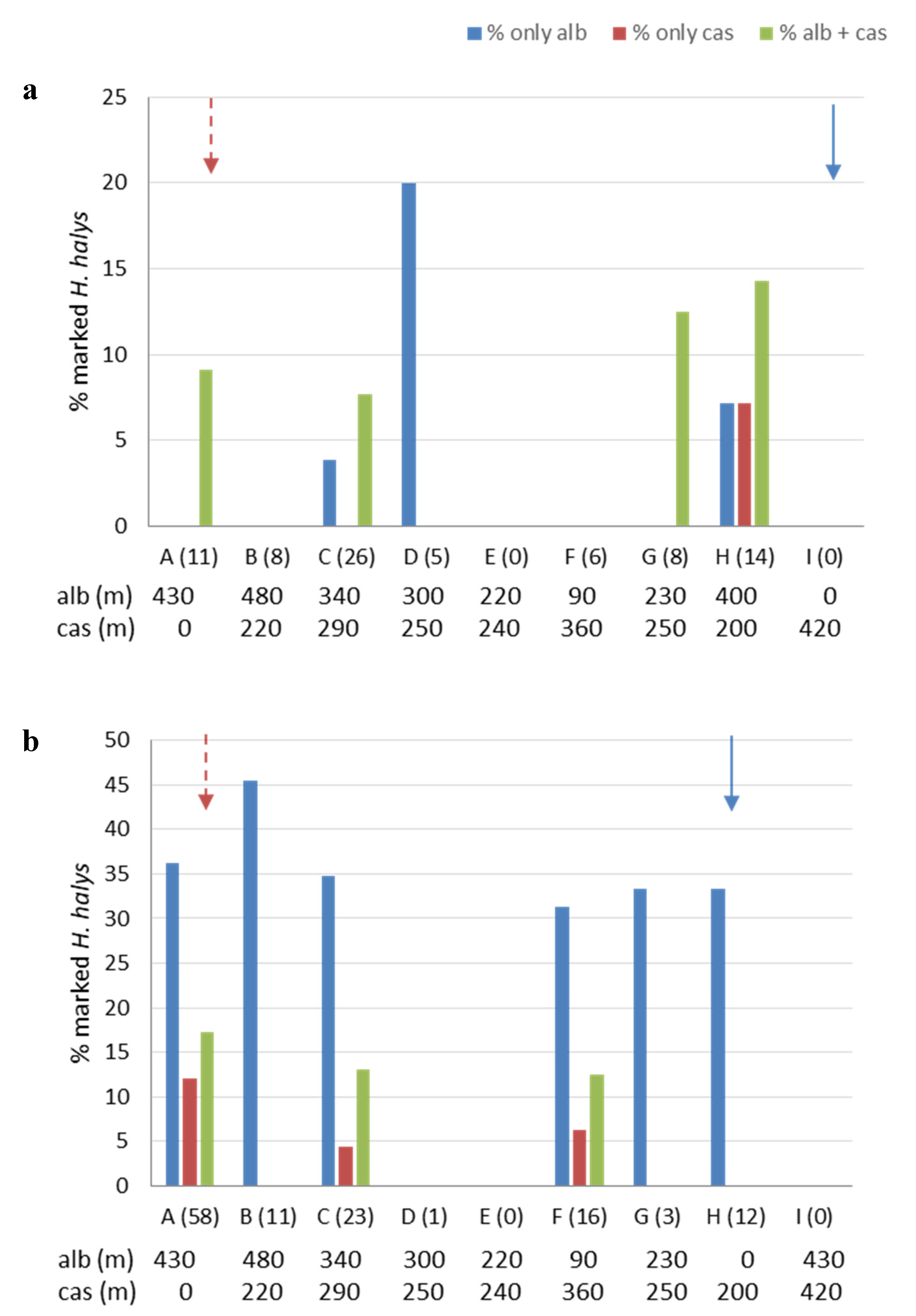
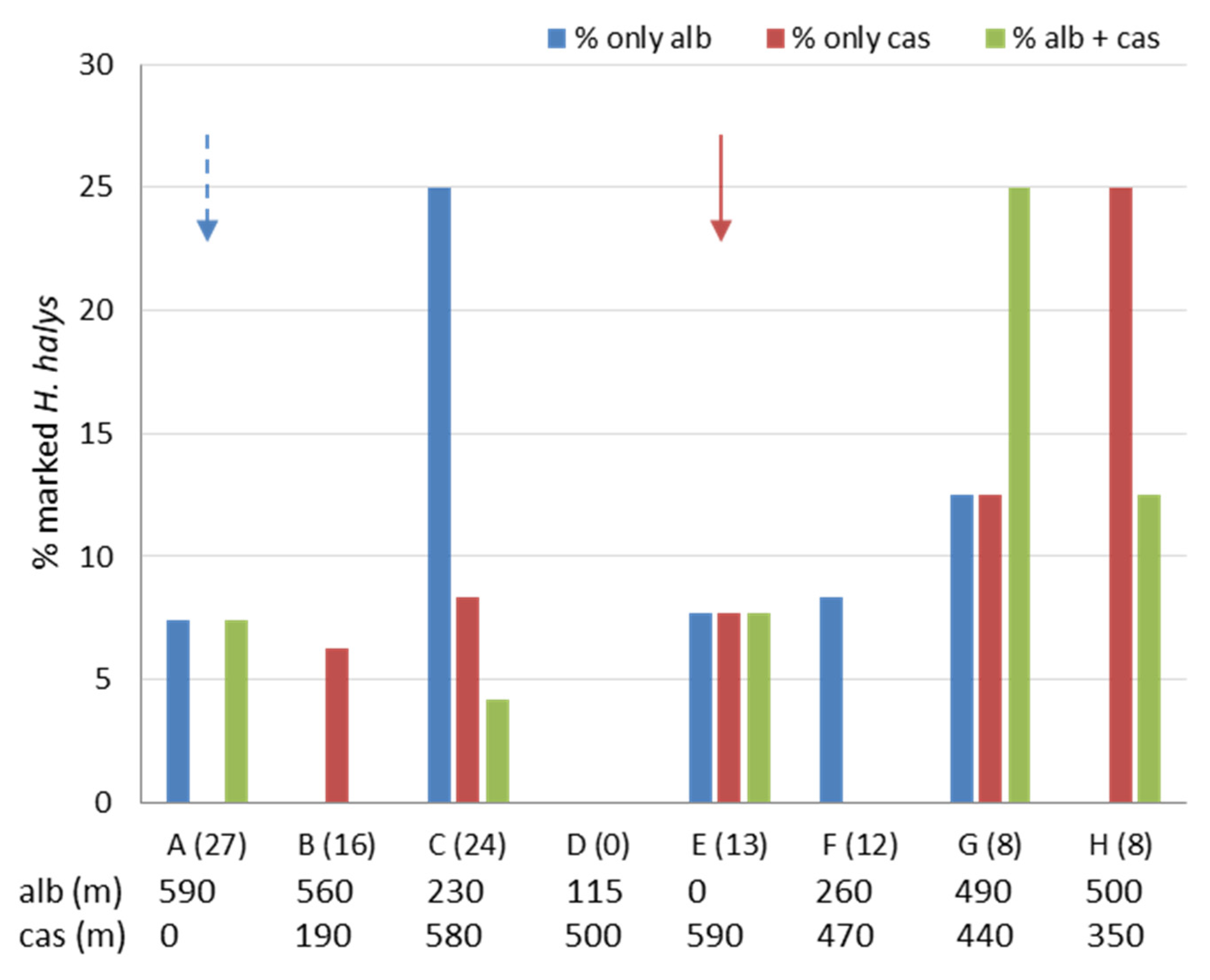
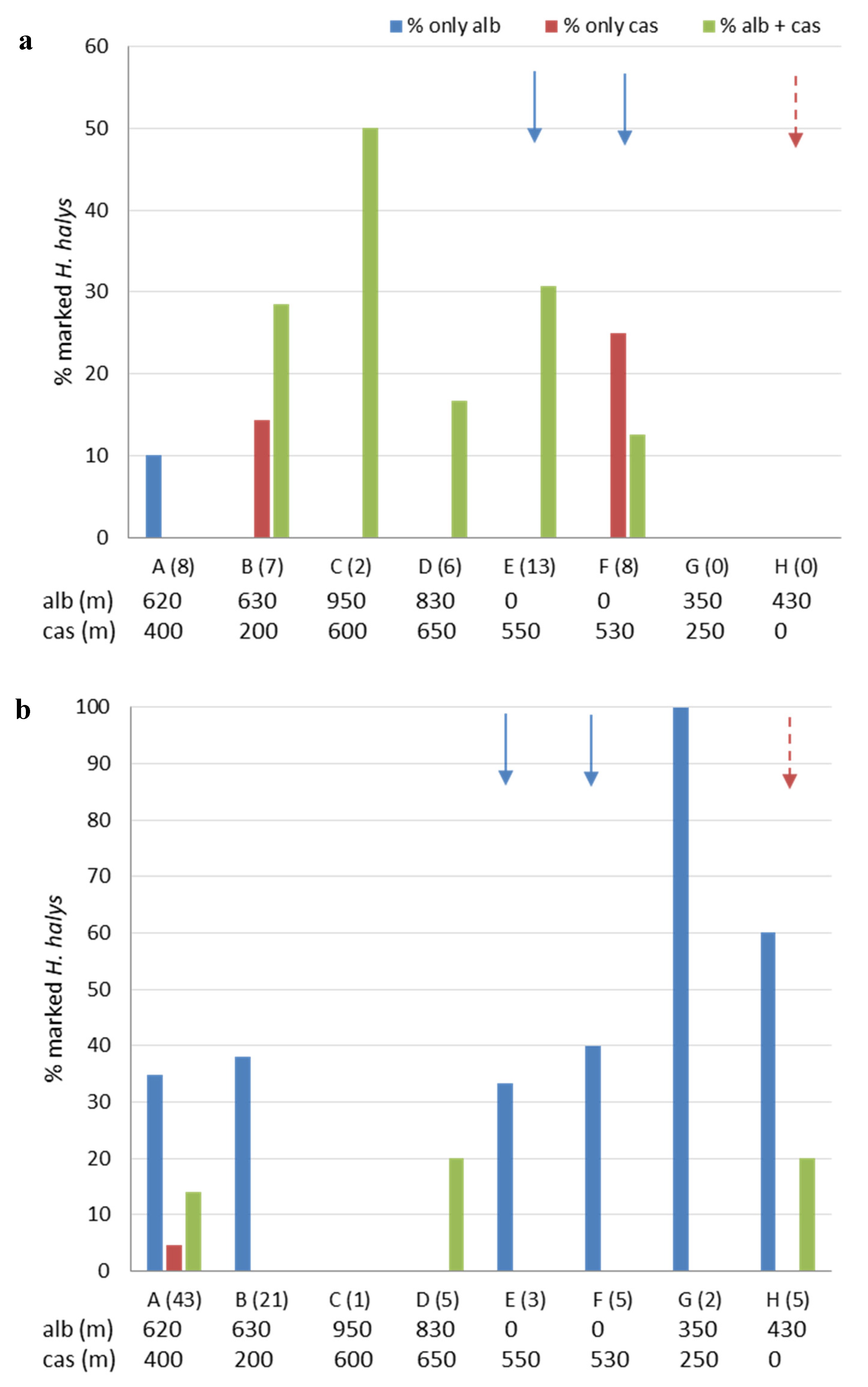
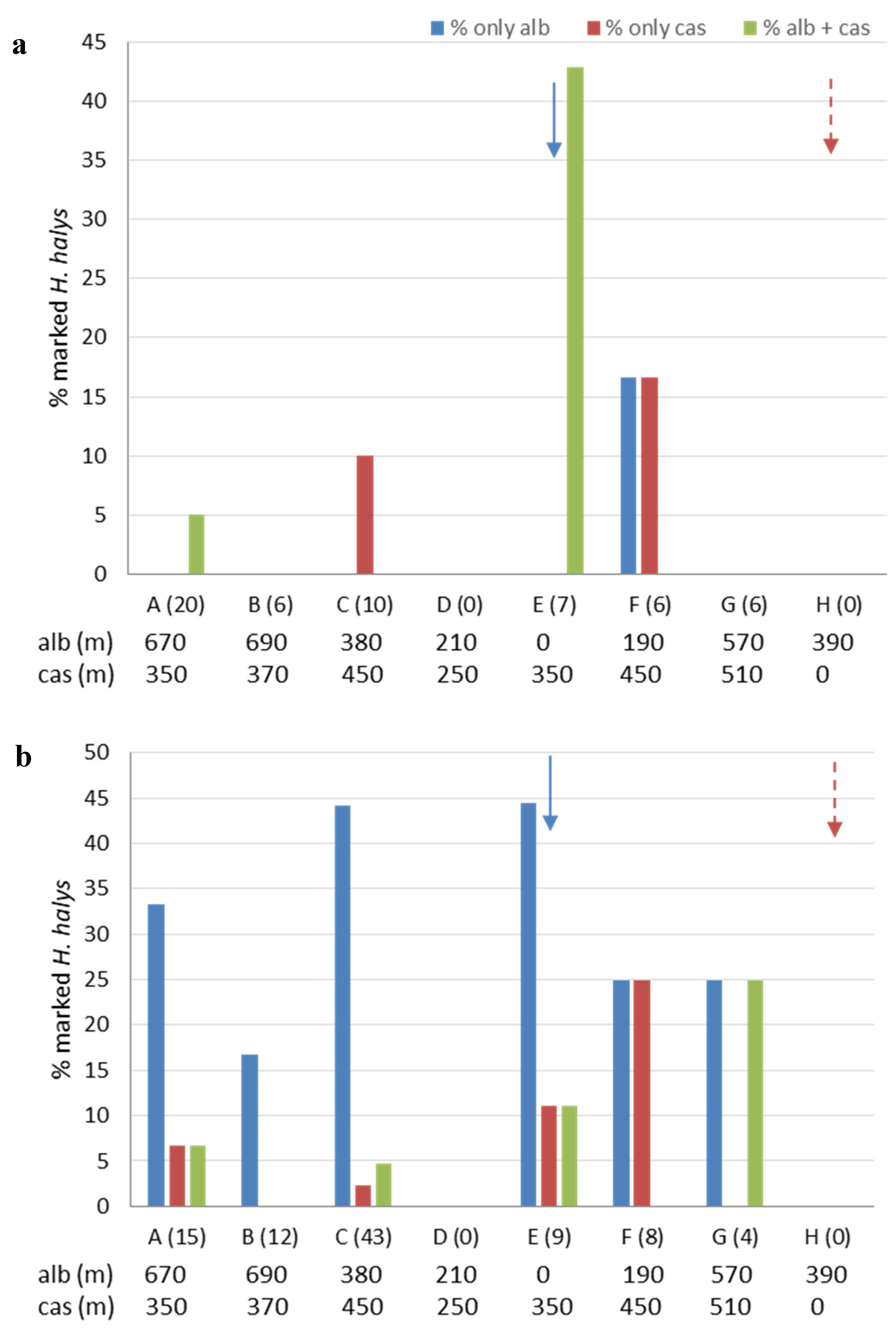
3.3. Movement and Crop Colonization Tracked with Immunomarking-Capture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leskey, T.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halyomorpha halys (Brown Marmorated Stink Bug). CABI Invasive Species Compendium. Available online: https://www.cabi.org/isc/datasheet/27377#REF-DDB-183911 (accessed on 5 August 2020).
- Maistrello, L.; Vaccari, G.; Caruso, S.; Costi, E.; Bortolini, S.; Macavei, L.; Foca, G.; Ulrici, A.; Bortolotti, P.P.; Nannini, R.; et al. Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in northern Italy. J. Pest Sci. 2017, 90, 1231–1244. [Google Scholar] [CrossRef]
- Bosco, L.; Moraglio, S.T.; Tavella, L. Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J. Pest Sci. 2017, 91, 661–670. [Google Scholar] [CrossRef]
- Kuhar, T.P.; Kamminga, K. Review of the chemical control research on Halyomorpha halys in the USA. J. Pest Sci. 2017, 90, 1021–1031. [Google Scholar] [CrossRef]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Rijal, J.; Gyawaly, S. Characterizing brown marmorated stink bug injury in almond, a new host crop in California. Insects 2018, 9, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acebes-Doria, A.; Leskey, T.C.; Bergh, J.C. Host plant effects on Halyomorpha halys (Hemiptera: Pentatomidae) nymphal development and survivorship. Environ. Entomol. 2016, 45, 663–670. [Google Scholar] [CrossRef]
- Lee, D.H.; Leskey, T.C. Flight behavior of foraging and overwintering brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). Bull. Entomol. Res. 2015, 105, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Wiman, N.G.; Walton, V.M.; Shearer, P.W.; Rondon, S.I.; Lee, J.C. Factors affecting flight capacity of brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). J. Pest Sci. 2014, 88, 37–47. [Google Scholar] [CrossRef]
- Lee, D.; Nielsen, A.L.; Leskey, T.C. Dispersal capacity and behavior of nymphal stages of Halyomorpha halys (Hemiptera: Pentatomidae) evaluated under laboratory and field conditions. J. Insect Behav. 2014, 27, 639–651. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Jones, V.P.; Nielsen, A.L. Utilizing immunomarking techniques to track Halyomorpha halys (Hemiptera: Pentatomidae) movement and distribution within a peach orchard. Peer J 2016, 4, e1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venugopal, P.D.; Dively, G.P.; Lamp, W.O. Spatiotemporal dynamics of the invasive Halyomorpha halys (Hemiptera: Pentatomidae) in and between adjacent corn and soybean fields. J. Econ. Entomol. 2015, 108, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Leskey, T.; Hamilton, G.; Nielsen, A.L.; Polk, D.; Rodriguez-Saona, C.; Bergh, J.C.; Herbert, D.A.; Kuhar, T.P.; Pfeiffer, D.; Dively, G.; et al. Pest status of the brown marmorated stink bug, Halyomorpha halys in the USA. Outlooks Pest Manag. 2012, 23, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Cullum, J.P.; Anderson, J.L.; Daugherty, J.L.; Beckett, L.M.; Leskey, T.C. Characterization of overwintering sites of the invasive brown marmorated stink bug in natural landscapes using human surveyors and detector canines. PLoS ONE 2014, 9, e91575. [Google Scholar] [CrossRef] [PubMed]
- Hancock, T.J.; Lee, D.H.; Bergh, J.C.; Morrison, W.R., III; Leskey, T.C. Presence of the invasive brown marmorated stink bug Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) on home exteriors during the autumn dispersal period: Results generated by citizen scientists. Agric. For. Entomol. 2019, 21, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Panizzi, A.R. Wild hosts of pentatomids: Ecological significance and role in their pest status on crops. Annu. Rev. Entomol. 1997, 42, 99–122. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.C.; Morrison, W.R., III; Khrimian, A.; Rice, K.B.; Leskey, T.C.; Rodriguez-Saona, C.; Nielsen, A.L.; Blaauw, B.R. Chemical ecology of Halyomorpha halys: Discoveries and applications. J. Pest Sci. 2017, 90, 989–1008. [Google Scholar] [CrossRef]
- Morrison, W.R., III; Milonas, P.; Kapantaidaki, D.E.; Cesari, M.; Bella, E.D.; Guidetti, R.; Haye, T.; Maistrello, L.; Moraglio, S.T.; Piemontese, L.; et al. Attraction of Halyomorpha halys (Hemiptera: Pentatomidae) haplotypes in North America and Europe to baited traps. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Morrison, W.R., III; Mathews, C.; Leskey, T.C.; Nielsen, A.L. Measuring host plant selection and retention of Halyomorpha halys by a trap crop. Entomol. Exp. Appl. 2017, 163, 197–208. [Google Scholar] [CrossRef]
- Jones, V.P.; Hagler, J.R.; Brunner, J.F.; Baker, C.C.; Wilburn, T.D. An inexpensive immunomarking technique for studying movement patterns of naturally occurring insect populations. Environ. Entomol. 2006, 35, 827–836. [Google Scholar] [CrossRef]
- Lessio, F.; Tota, F.; Alma, A. Tracking the dispersion of Scaphoideus titanus Ball (Hemiptera: Cicadellidae) from wild to cultivated grapevine: Use of a novel mark–capture technique. Bull. Entomol. Res. 2014, 104, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Hagler, J.R. Super Mark It! A review of the protein immunomarking technique. Ann. Entomol. Soc. Am. 2019, 112, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Wallner, A.M.; Hamilton, G.C.; Nielsen, A.L.; Hahn, N.; Green, E.J.; Rodriguez-Saona, C.R. Landscape factors facilitating the invasive dynamics and distribution of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), after arrival in the United States. PLoS ONE 2014, 9, e95691. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; Acebes-Doria, A.L.; Rice, K.B.; Short, B.D.; Adams, C.G.; Gut, L.J.; Leskey, T.C. Estimating monitoring trap plume reach and trapping area for nymphal and adult Halyomorpha halys (Hemiptera: Pentatomidae) in crop and non-crop habitats. Environ. Entomol. 2019, 48, 1104–1112. [Google Scholar] [CrossRef] [Green Version]
- Morrison, W.R., III; Allen, M.; Leskey, T.C. Behavioural response of the invasive Halyomorpha halys (Hemiptera: Pentatomidae) to host plant stimuli augmented with semiochemicals in the field. Agric. For. Entomol. 2018, 20, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Bergh, J.C.; Morrison, W.R., III; Joseph, S.V.; Leskey, T.C. Characterizing spring emergence of adult Halyomorpha halys using experimental overwintering shelters and commercial pheromone traps. Entomol. Exp. Appl. 2017, 162, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Morrison, W.R., III; Acebes-Doria, A.; Ogburn, E.; Kuhar, T.P.; Walgenbach, J.F.; Bergh, J.C.; Nottingham, L.; Dimeglio, A.; Hipkins, P.; Leskey, T.C. Behavioral response of the brown marmorated stink bug (Hemiptera: Pentatomidae) to semiochemicals deployed inside and outside anthropogenic structures during the overwintering period. J. Econ. Entomol. 2017, 110, 1002–1009. [Google Scholar] [CrossRef]
- Costi, E.; Haye, T.; Maistrello, L. Biological parameters of the invasive brown marmorated stink bug, Halyomorpha halys, in southern Europe. J. Pest Sci. 2017, 90, 1059–1067. [Google Scholar] [CrossRef]
- Martinson, H.M.; Venugopal, P.D.; Bergmann, E.J.; Shrewsbury, P.M.; Raupp, M.J. Fruit availability influences the seasonal abundance of invasive stink bugs in ornamental tree nurseries. J. Pest Sci. 2015, 88, 461–468. [Google Scholar] [CrossRef]
- Formella, A.; Dorman, S.J.; Taylor, S.V.; Kuhar, T.P. Effects of aggregation lure and tree species on Halyomorpha halys (Hemiptera: Pentatomidae) seasonal oviposition. J. Econ. Entomol. 2019, 113, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Hedstrom, C.S.; Shearer, P.W.; Miller, J.C.; Walton, V.M. The effects of kernel feeding by Halyomorpha halys (Hemiptera: Pentatomidae) on commercial hazelnuts. J. Econ. Entomol. 2014, 107, 1858–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathews, C.; Blaauw, B.; Dively, G.; Kotcon, J.; Moore, J.L.; Ogburn, E.; Pfeiffer, D.G.; Trope, T.; Walgenbach, J.F.; Welty, C.; et al. Evaluating a polyculture trap crop for organic management of Halyomorpha halys and native stink bugs in peppers. J. Pest Sci. 2017, 90, 1245–1255. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area Characteristics | Sampling Period (No. of Samplings) | Trap Details | |||||||
|---|---|---|---|---|---|---|---|---|---|
| # | Location | Surface Area (ha) | Forest (ha) | # | Surrounding Crops/Tree Rows/Forest | Min. Distance (m) from | |||
| Water | Road | Build. | |||||||
| 1 | Antignano (AT) N: 44°50′ E: 8°08′ 260 m a.s.l. | 13.6 | 61.4 S-SE-SW | traps: 17 Apr–4 Jul (6) plants: 23 Apr–18 Jun (5) | 1 | forest | 338 | 106 | 81 |
| 2 | hazelnut, forest | 114 | 167 | 185 | |||||
| 3 | hazelnut | 218 | 37 | 50 | |||||
| 4 | hazelnut, meadow, wheat | 533 | 36 | 101 | |||||
| 5 | vineyard, maize, apple | 501 | 80 | 55 | |||||
| 6 | maize, meadow | 430 | 44 | 35 | |||||
| 2 | Frossasco (TO) N: 44°56′ E: 7°21′ 376 m a.s.l. | 28.1 | 30.9 N; middle | traps: 5 Apr–4 Jul (7) plants: 20 Apr–22 Jun (6) | 1 | hazelnut, meadow | 888 | 538 | 135 |
| 2 | wheat, meadow, forest | 1002 | 604 | 277 | |||||
| 3 | meadow, ryegrass, forest | 841 | 1080 | 317 | |||||
| 4 | meadow, ryegrass, forest | 603 | 1140 | 243 | |||||
| 5 | maize | 486 | 967 | 113 | |||||
| 6 | maize | 446 | 753 | 195 | |||||
| 3 | Moncestino (AL) N: 45°9′ E: 8°09′ 287 m a.s.l. | 48.1 | 147.4S-SW | traps: 5 Apr–29 Jun (7) plants: 18 Apr–14 Jun (6) | 1 | hazelnut, poplar, sunflower | 317 | 111 | 235 |
| 2 | maize, wheat, poplar, sorghum | 516 | 229 | 193 | |||||
| 3 | maize | 134 | 234 | 110 | |||||
| 4 | maize | 245 | 33 | 174 | |||||
| 5 | forest | 440 | 41 | 168 | |||||
| 6 | maize, forest | 507 | 202 | 315 | |||||
| 4 | Narzole (CN) N: 44°36′ E: 7°52′ 325 m a.s.l. | 50.5 | 86.6 E-NE-N; SW | traps: 6 Apr–4 Jul (8) plants: 19 Apr–22 Jun (7) | 1 | hazelnut, alfalfa | 1 | 108 | 209 |
| 2 | hazelnut, forest | 118 | 277 | 284 | |||||
| 3 | hazelnut, hybrid planes | 15 | 844 | 161 | |||||
| 4 | forest, hazelnut, maples | 43 | 862 | 50 | |||||
| 5 | hazelnut, forest, maples | 5 | 838 | 235 | |||||
| 6 | hazelnut, alfalfa, barley | 1 | 436 | 301 | |||||
| Area | Date | Milk | Egg White | First Capture Date | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Site/Plant | Zone a | Vol. (L) b | Trap c | Site/Plant | Zone a | Vol. (L) b | Trap c | |||
| 1 | 17 Apr | rubble, woodshed | A | 28 | 1 | - | 23 Apr | |||
| 23 Apr | - | hazelnut orchard | I | 28 | 4 | 15 May | ||||
| 15 May | peach trees | A | 14 | 1 | oak | H | 28 | 6 | 23 May | |
| 2 | 20 Apr | hazelnut orchard | A | 28 | 1 | wild hazelnut | E | 28 | 4 | 26 Apr |
| 3 | 5 Apr | rubble, | H | 14 | - | - | 18 Apr | |||
| woodshed | 14 | |||||||||
| 18 Apr | rubble, | H | 14 | - | ivy, black locust, oak, chestnut | E | 14 | 5 | 27 Apr | |
| woodshed | 14 | F | 14 | 6 | ||||||
| 4 | 6 Apr | shelter, woodshed | H | 28 | - | - | 19 Apr | |||
| 19 Apr | shelter, woodshed | H | 28 | - | ivy, oak, ash | E | 14 | 4 | 26 Apr | |
| 9 May | shelter, woodshed | H | 28 | - | - | 17 May | ||||
| 17 May | - | cherry, apple, blackthorn | E | 14 | 4 | 21 May | ||||
| Host Plants | April | May | June | Mean No. | Area of Presence | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Common Name | Family | Scientific Name | II | III | I | II | III | I | II | ||
| peach | Rosaceae | Prunus persica (L.) Batsch | - | ++ | - | +++ | ++ | 3.07 ± 2.06 | 1 | ||
| mulberry | Moraceae | Morus rubra L. | - | - | ++ | ++ | ++ | 1.63 ± 0.78 | 1, 2 | ||
| apple | Rosaceae | Malus domestica Borkh. | + | - | ++ | 1.33 ± 0.88 | 1, 2, 4 | ||||
| common ash | Oleaceae | Fraxinus excelsior L. | - | - | ++ | - | + | +++ | + | 1.24 ± 0.70 | 2, 4 |
| maple | Sapindaceae | Acer spp. | - | - | - | - | - | ++ | ++ | 0.63 ± 0.40 | 1, 2, 3, 4 |
| hybrid plane | Platanaceae | Platanus hispanica Mill. | ++ | - | - | - | - | 0.60 ± 0.60 | 4 | ||
| American pokeweed | Phytolaccaceae | Phytolacca americana L. | + | - | 0.50 ± 0.50 | 2 | |||||
| sorrel | Polygonaceae | Rumex spp. | + | - | 0.50 ± 0.50 | 3, 4 | |||||
| vetch | Fabaceae | Vicia spp. | + | - | 0.50 ± 0.50 | 4 | |||||
| hazelnut | Betulaceae | Corylus avellana L. | - | - | - | - | - | + | ++ | 0.40 ± 0.32 | 1, 2, 3, 4 |
| cherry | Rosaceae | Prunus avium (L.) L. | - | - | + | + | + | + | + | 0.34 ± 0.09 | 1, 2, 3, 4 |
| bay | Lauraceae | Laurus nobilis L. | - | - | + | 0.33 ± 0.33 | 1 | ||||
| black poplar | Salicaceae | Populus spp. | - | - | ++ | - | - | - | 0.33 ± 0.33 | 2, 3, 4 | |
| black walnut | Juglandaceae | Juglans nigra L. | + | - | - | 0.33 ± 0.33 | 2, 3 | ||||
| wild hop | Cannabaceae | Humulus lupulus L. | - | + | - | 0.33 ± 0.33 | 1, 2, 3 | ||||
| European spindletree | Celastraceae | Euonymus europaeus L. | - | + | + | + | - | + | 0.33 ± 0.12 | 2, 3 | |
| ivy | Araliaceae | Hedera helix L. | - | - | - | - | + | ++ | - | 0.30 ± 0.20 | 1, 3, 4 |
| walnut | Juglandaceae | Juglans regia L. | - | + | + | + | + | + | - | 0.29 ± 0.11 | 1, 2, 3, 4 |
| fig | Moraceae | Ficus carica L. | - | - | ++ | - | - | 0.27 ± 0.27 | 1 | ||
| false indigo-bush | Fabaceae | Amorpha fruticosa L. | - | - | + | - | - | 0.20 ± 0.20 | 3 | ||
| black locust | Fabaceae | Robinia pseudoacacia L. | - | - | + | + | + | - | + | 0.18 ± 0.08 | 1, 2, 3, 4 |
| common wheat | Poaceae | Triticum aestivum L. | - | + | - | - | - | + | 0.17 ± 0.11 | 2, 3 | |
| European elder | Adoxaceae | Sambucus nigra L. | - | + | + | - | - | - | + | 0.16 ± 0.10 | 1, 2, 3, 4 |
| bloody dogwood | Cornaceae | Cornus sanguinea L. | - | - | + | - | + | + | - | 0.14 ± 0.07 | 1, 2, 3, 4 |
| elm | Ulmaceae | Ulmus minor Mill. | - | - | - | - | + | + | 0.13 ± 0.09 | 1, 2, 3, 4 | |
| oak | Fagaceae | Quercus spp. | - | + | + | + | - | - | - | 0.11 ± 0.07 | 1, 2, 3, 4 |
| sand grape | Vitaceae | Vitis rupestris Scheele | - | - | - | + | - | 0.10 ± 0.10 | 1, 2, 3 | ||
| blackthorn | Rosaceae | Prunus spinosa L. | - | - | - | - | - | + | 0.08 ± 0.08 | 1, 2 | |
| willow | Salicaceae | Salix spp. | - | - | - | + | - | - | - | 0.07 ± 0.07 | 2, 3, 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosco, L.; Nardelli, M.; Tavella, L. First Insights on Early Host Plants and Dispersal Behavior of Halyomorpha halys (Hemiptera: Pentatomidae) from Overwintering to Crop Colonization. Insects 2020, 11, 866. https://doi.org/10.3390/insects11120866
Bosco L, Nardelli M, Tavella L. First Insights on Early Host Plants and Dispersal Behavior of Halyomorpha halys (Hemiptera: Pentatomidae) from Overwintering to Crop Colonization. Insects. 2020; 11(12):866. https://doi.org/10.3390/insects11120866
Chicago/Turabian StyleBosco, Lara, Martina Nardelli, and Luciana Tavella. 2020. "First Insights on Early Host Plants and Dispersal Behavior of Halyomorpha halys (Hemiptera: Pentatomidae) from Overwintering to Crop Colonization" Insects 11, no. 12: 866. https://doi.org/10.3390/insects11120866