Transcriptomics-Based Approach Identifies Spinosad-Associated Targets in the Colorado Potato Beetle, Leptinotarsa decemlineata

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Isolation
2.3. Large RNA Library Construction and Sequencing
2.4. Small RNA Library Construction and Sequencing
2.5. Synthesis of cDNA
2.6. qRT-PCR Amplification of Spinosad-Associated Transcripts Identified via NGS
2.7. Functional Assessment of Spinosad-Modulated mRNAs
3. Results
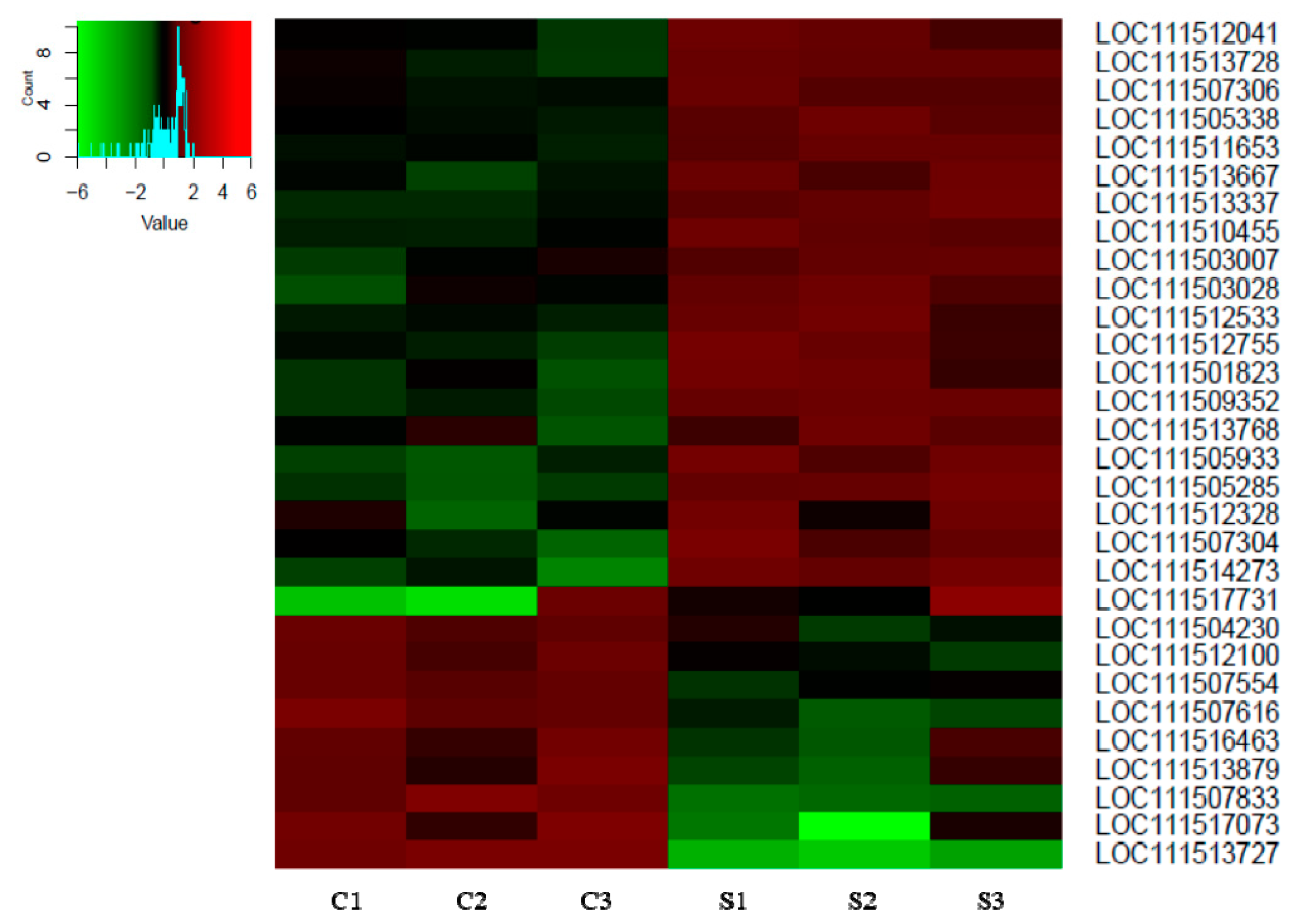
3.1. mRNA Expression in Spinosad-Exposed L. decemlineata by NGS
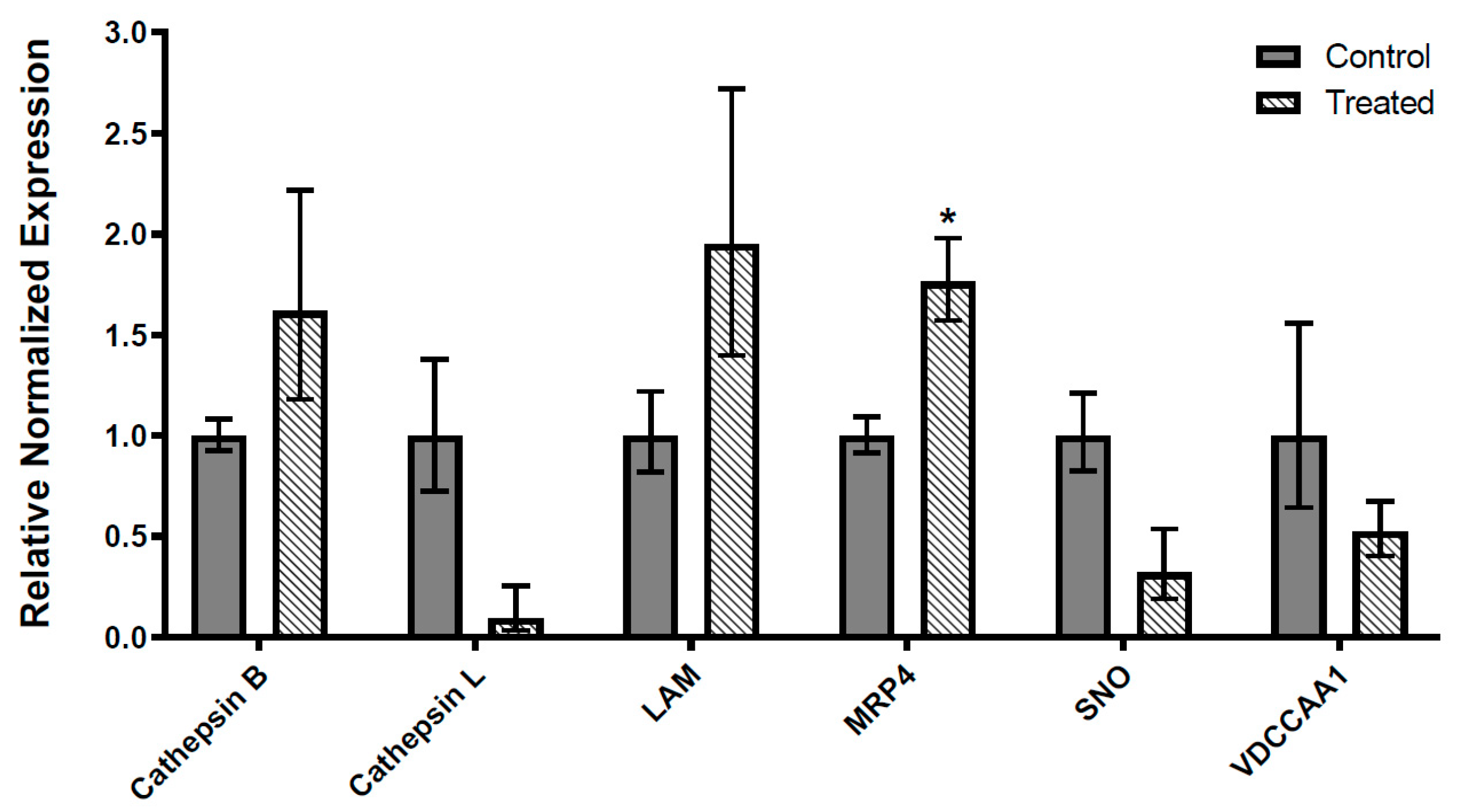
3.2. qRT-PCR-Based Expression of Transcripts Modulated by Spinosad in L. decemlineata as Determined by NGS
3.3. miRNA Expression in Control and Spinosad-Exposed Insects via NGS
3.4. Functional Classification of Differentially Expressed mRNA Transcripts Following Spinosad Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weber, D. Colorado beetle: Pest on the move. Pestic. Outlook 2003, 14, 256–259. [Google Scholar] [CrossRef]
- Hare, D.J. Impact of defoliation by the Colorado potato beetle on potato yields. J. Econ. Entomol. 1980, 73, 369–373. [Google Scholar] [CrossRef]
- Weber, D.C.; Ferro, D.N.; Buonaccorsi, J.; Hazzard, R.V. Disrupting spring colonization of Colorado potato beetle to nonrotated potato fields. Entomol. Exp. Appl. 1994, 73, 39–50. [Google Scholar] [CrossRef]
- Weisz, R.; Smilowitz, Z.; Christ, B. Distance, rotation, and border crops affect Colorado potato beetle (Coleoptera, Chrysomelidae) colonization and population-density and early blight (Alternaria-solani) severity in rotated potato fields. J. Econ. Entomol. 1994, 87, 723–729. [Google Scholar] [CrossRef]
- Wraight, S.P.; Ramos, M.E. Synergistic interaction between Beauveria bassiana- and Bacillus thuringiensis tenebrionis-based biopesticides applied against field populations of Colorado potato beetle larvae. J. Invertebr. Pathol. 2005, 90, 139–150. [Google Scholar] [CrossRef]
- Guo, W.C.; Wang, Z.A.; Luo, X.L.; Jin, X.; Chang, J.; He, J.; Tu, E.X.; Tian, Y.C.; Si, H.J.; Wu, J.H. Development of selectable marker-free transgenic potato plants expressing cry3A against the Colorado potato beetle (Leptinotarsa decemlineata Say). Pest Manag. Sci. 2016, 72, 497–504. [Google Scholar] [CrossRef]
- Scott, I.M.; Tolman, J.H.; MacArthur, D.C. Insecticide resistance and cross-resistance development in Colorado potato beetle Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae) populations in Canada 2008–2011. Pest Manag. Sci. 2015, 71, 712–721. [Google Scholar] [CrossRef]
- Szendrei, Z.; Grafius, E.; Byrne, A.; Ziegler, A. Resistance to neonicotinoid insecticides in field populations of the Colorado potato beetle (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2012, 68, 941–946. [Google Scholar] [CrossRef]
- Huseth, A.S.; Groves, R.L. Effect of insecticide management history on emergence phenology and neonicotinoid resistance in Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2013, 106, 2491–2505. [Google Scholar] [CrossRef]
- Mota-Sanchez, D.; Hollingworth, R.M.; Grafius, E.J.; Moyer, D.D. Resistance and cross-resistance to neonicotinoid insecticides and spinosad in the Colorado potato beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2006, 62, 30–37. [Google Scholar] [CrossRef]
- Hertlein, M.B.; Mavrotas, C.; Jousseaume, C.; Lysandrou, M.; Thompson, G.D.; Jany, W.; Ritchie, S.A. A review of spinosad as a natural product for larval mosquito control. J. Am. Mosq. Control Assoc. 2010, 26, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.R.; Rodrigues, A.R.; Silva, W.M.; Silva, T.B.; Silva, V.R.; Guedes, R.N.; Siqueira, H.A. Spinosad and the tomato borer Tuta absoluta: A bioinsecticide, an invasive pest threat, and high insecticide resistance. PLoS ONE 2014, 9, e103235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayyed, A.H.; Saeed, S.; Noor-Ul-Ane, M.; Crickmore, N. Genetic, biochemical, and physiological characterization of spinosad resistance in Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2008, 101, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Shad, S.A.; Sayyed, A.H.; Fazal, S.; Saleem, M.A.; Zaka, S.M.; Ali, M. Field evolved resistance to carbamates, organophosphates, pyrethroids, and new chemistry insecticides in Spodoptera litura Fab. (Lepidoptera: Noctuidae). J. Pest Sci. 2012, 85, 153–162. [Google Scholar] [CrossRef]
- Okuma, D.M.; Bernardi, D.; Horikoshi, R.J.; Bernardi, O.; Silva, A.P.; Omoto, C. Inheritance and fitness costs of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to spinosad in Brazil. Pest Manag. Sci. 2018, 74, 1441–1448. [Google Scholar] [CrossRef]
- Perry, T.; McKenzie, J.A.; Batterham, P.A. Dalpha6 knockout strain of Drosophila melanogaster confers a high level of resistance to spinosad. Insect Biochem. Mol. Biol. 2007, 37, 184–188. [Google Scholar] [CrossRef]
- Yin, X.H.; Wu, Q.J.; Zhang, Y.J.; Long, Y.H.; Wu, X.M.; Li, R.Y.; Wang, M.; Tian, X.L.; Jiao, X.G. Analysis of persistent changes to γ-aminobutyric acid receptor gene expression in Plutella xylostella subjected to sublethal amounts of spinosad. Genet. Mol. Res. 2016, 15, gmr.15038782. [Google Scholar] [CrossRef]
- Yang, M.; Hao, Y.; Gao, J.; Zhang, Y.; Xu, W.; Tao, L. Spinosad induces autophagy of Spodoptera frugiperda Sf9 cells and the activation of AMPK/mTOR signaling pathway. Comp. Biochem. Physiol. 2017, C195, 52–59. [Google Scholar] [CrossRef]
- Yang, M.; Wang, B.; Gao, J.; Zhang, Y.; Xu, W.; Tao, L. Spinosad induces programmed cell death involves mitochondrial dysfunction and cytochrome C release in Spodoptera frugiperda Sf9 cells. Chemosphere 2017, 169, 155–161. [Google Scholar] [CrossRef]
- Sagri, E.; Reczko, M.; Gregoriou, M.E.; Tsoumani, K.T.; Zygouridis, N.E.; Salpea, K.D.; Zalom, F.G.; Ragoussis, J.; Mathiopoulos, K.D. Olive fly transcriptomics analysis implicates energy metabolism genes in spinosad resistance. BMC Genom. 2008, 15, 714. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.H.; Lu, W.P.; Guo, W.C.; Xia, Z.H.; Fu, W.J.; Li, G.Q. Chlorantraniliprole susceptibility in Leptinotarsa decemlineata in the north Xinjiang Uygur autonomous region in China. J. Econ. Entomol. 2012, 105, 549–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumas, P.; Sambou, M.; Gaudet, J.D.; Morin, M.D.; Moffat, C.E.; Boquel, S.; Jr Morin, P. Differential expression of transcripts with potential relevance to chlorantraniliprole response in the Colorado potato beetle, Leptinotarsa decemlineata. Arch. Insect Biochem. Physiol. 2020, 103, e21642. [Google Scholar] [CrossRef] [PubMed]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Wysoker, A.; Tibbetts, K.; Fennell, T. Picard Tools Version 2.5. 2014. Available online: http://broadinstitute.github.io/picard (accessed on 12 February 2019).
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Patcher, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Xu, H.; Leung, R.K.K. An optimized protocol for generation and analysis of Ion proton sequencing reads for RNA-Seq. BMC Genom. 2016, 17, 403. [Google Scholar] [CrossRef] [Green Version]
- Grieve, S.; Wajnberg, G.; Lees, M.; Chacko, S.; Weir, J.; Crapoulet, N.; Reinman, T. TAZ functions as tumor suppressor in multiple myeloma by downregulating MYC. Blood Adv. 2019, 3, 3613–3625. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 12 February 2019).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalsi, M.; Palli, S.R. Transcription factor cap n collar C regulates multiple cytochrome P450 genes conferring adaptation to potato plant allelochemicals and resistance to imidacloprid in Leptinotarsa decemlineata (Say). Insect Biochem. Mol. Biol. 2017, 83, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.W.; Li, Y.Z.; Li, G.Q.; Wan, P.J.; Li, C. Identification of cuticular protein genes in the colorado potato beetle Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2019, 112, 912–923. [Google Scholar] [CrossRef]
- Clements, J.; Sanchez-Sedillo, B.; Bradfield, C.A.; Groves, R.L. Transcriptomic analysis reveals similarities in genetic activation of detoxification mechanisms resulting from imidacloprid and chlorothalonil exposure. PLoS ONE 2018, 13, e0205881. [Google Scholar] [CrossRef] [Green Version]
- Hemingway, J.; Hawkes, N.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef]
- Gong, Y.J.; Wang, Z.H.; Shi, B.C.; Kang, Z.J.; Zhu, L.; Jin, G.H.; Weig, S.J. Correlation between pesticide resistance and enzyme activity in the diamondback moth, Plutella xylostella. J. Insect Sci. 2013, 13, 135. [Google Scholar] [CrossRef] [Green Version]
- Lü, F.G.; Fu, K.Y.; Li, Q.; Guo, W.C.; Ahmat, T.; Li, G.Q. Identification of carboxylesterase genes and their expression profiles in the Colorado potato beetle Leptinotarsa decemlineata treated with fipronil and cyhalothrin. Pestic. Biochem. Physiol. 2015, 122, 86–95. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, M.; Ma, Z.; You, C.; Gao, X.; Shi, X. Esterase-mediated spinosad resistance in house flies Musca domestica (Diptera: Muscidae). Ecotoxicology 2020, 29, 35–44. [Google Scholar] [CrossRef]
- Dermauw, W.; Van Leeuwen, T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance. Insect Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.L.; Zhang, M.; Wang, K.; Qiao, X.F.; Chen, M.H. Molecular cloning, expression pattern of multidrup resistance associated protein 1 (MRP1, ABCC1) gene, and the synergistic effects of verapamil on toxicity of two insecticides in the bird cherry-oat aphid. Arch. Insect Biochem. Physiol. 2016, 92, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Gaddelapati, S.C.; Kalsi, M.; Roy, A.; Palli, S.R. Cap ‘n’ collar C regulates genes responsible for imidacloprid resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Insect Biochem. Mol. Biol. 2018, 99, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.; Olson, J.M.; Sanchez-Sedillo, B.; Bradford, B.; Groves, R.L. Changes in emergence phenology, fatty acid composition, and xenobiotic-metabolizing enzyme expression is associated with increased insecticide resistance in the Colorado potato beetle. Arch. Insect Biochem. Physiol. 2020, 103, e21630. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Yang, M.; Gao, J.; Zhang, Y.; Tao, L. Oxidative stress and DNA damage induced by spinosad exposure in Spodoptera frugiperda Sf9 cells. Food Agric. Immunol. 2018, 29, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Piner, P.; Üner, N. Oxidative stress and apoptosis was induced by bio-insecticide spinosad in the liver of Oreochromis niloticus. Environ. Toxicol. Pharmacol. 2013, 36, 956–963. [Google Scholar] [CrossRef]
- Arakane, Y.; Muthukrishnan, S. Insect chitinase and chitinase-like proteins. Cell Mol. Life Sci. 2010, 67, 201–216. [Google Scholar] [CrossRef]
- Shaurub, E.S.H.; El Aziz, N.M.A. Biochemical effects of lambda cyhalothrin and lufenuron on Culex pipiens L. (Diptera: Culicidae). Int. J. Mosq. Res. 2015, 2, 122–126. [Google Scholar]
- Zhang, J.; Sun, T.; Sun, Z.; Li, H.; Qi, X.; Zhong, G.; Yi, X. Azadirachtin acting as a hazardous compound to induce multiple detrimental effects in Drosophila melanogaster. J. Hazard. Mater. 2018, 359, 338–347. [Google Scholar] [CrossRef]
- Dias, R.O.; Cardoso, C.; Pimentel, A.C.; Damasceno, T.F.; Ferreira, C.; Terra, W.R. The roles of mucus-forming mucins, peritrophins and peritrophins with mucin domains in the insect midgut. Insect Mol. Biol. 2018, 27, 46–60. [Google Scholar] [CrossRef]
- Sandoval-Mojica, A.F.; Scharf, M.E. Silencing gut genes associated with the peritrophic matrix of Reticulitermes flavipes (Blattodea: Rhinotermitidae) increases susceptibility to termiticides. Insect Mol. Biol. 2016, 25, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Duan, D.; Zheng, R.; Lin, S.; Chen, Y.; Tian, H.; Zhao, J.; Tian, S.; Wei, H.; Gu, X. Modulation of juvenile hormone esterase gene expression against development of Plutella xylostella (Lepidoptera: Plutellidae). J. Econ. Entomol. 2016, 109, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Tian, S.; Ren, K.; Chen, Y.; Lin, S.; Chen, Y.; Tian, H.; Zhao, J.; Wang, C.; Wei, H.; et al. Effect of treatment with 3-octylthio-1,1,1-trifluoropropan-2-one in the diamondback moth (Lepidoptera: Plutellidae) to the toxicity of diafenthiuron, indoxacarb, and Bacillus thuringiensis. J. Econ. Entomol. 2020, 16, toaa041. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Congiu, L.; Lindström, L.; Piiroinen, S.; Vidotto, M.; Grapputo, A. Sequencing, De Novo assembly and annotation of the Colorado Potato Beetle, Leptinotarsa decemlineata, Transcriptome. PLoS ONE 2014, 9, e86012. [Google Scholar] [CrossRef] [Green Version]
- Morin, M.D.; Lyons, P.J.; Crapoulet, N.; Boquel, S.; Morin, P.J. Identification of differentially expressed miRNAs in colorado potato beetles (Leptinotarsa decemlineata (Say)) exposed to imidacloprid. Int. J. Mol. Sci. 2017, 18, 2728. [Google Scholar] [CrossRef] [Green Version]
- Puinean, A.M.; Foster, S.P.; Oliphant, L.; Denholm, I.; Field, L.M.; Millar, N.S.; Williamson, M.S.; Bass, C. Amplification of a cytochrome P450 gene is associated with resistance to neonicotinoid insecticides in the aphid Myzus persicae. PLoS Genet. 2010, 6, e1000999. [Google Scholar] [CrossRef] [Green Version]
- Peng, T.; Pan, Y.; Gao, X.; Xi, J.; Zhang, L.; Ma, K.; Wu, Y.; Zhang, J.; Shang, Q. Reduced abundance of the CYP6CY3-targeting let-7 and miR-100 miRNAs accounts for host adaptation of Myzus persicae nicotianae. Insect Biochem. Mol. Biol. 2016, 75, 89–97. [Google Scholar] [CrossRef]
- Wan, P.J.; Shi, X.Q.; Kong, Y.; Zhou, L.T.; Guo, W.C.; Ahmat, T.; Li, G.Q. Identification of cytochrome P450 monooxygenase genes and their expression profiles in cyhalothrin-treated Colorado potato beetle, Leptinotarsa decemlineata. Pestic. Biochem. Physiol. 2013, 107, 360–368. [Google Scholar] [CrossRef]
- Kaplanoglu, E.; Chapman, P.; Scott, I.M.; Donly, C. Overexpression of a cytochrome P450 and a UDP-glycosyltransferase is associated with imidacloprid resistance in the Colorado potato beetle, Leptinotarsa decemlineata. Sci. Rep. 2017, 7, 1762. [Google Scholar] [CrossRef] [Green Version]
- López-Galiano, M.J.; García-Robles, I.; Ruiz-Arroyo, V.M.; Sanchís Oltra, S.; Petek, M.; Rausell, C.; Real, M.D. Colorado potato beetle chymotrypsin genes are differentially regulated in larval midgut in response to the plant defense inducer hexanoic acid or the Bacillus thuringiensis Cry3Aa toxin. J. Invertebr. Pathol. 2019, 166, 107224. [Google Scholar] [CrossRef]
- Vickruck, J.L.; Scott, I.M.; Krolikowski, S.; MacKinley, P.; Donly, C.; Hann, S.; Moffat, C.E. Canada wide evaluation of the susceptibility of Colorado potato beetle larvae to six registered insecticides—Pest management research report 2019. Agric. Agri-Food Can. 2019, 58, 69. Available online: https://phytopath.ca/wp-content/uploads/2020/04/PMRR-2019-final.pdf (accessed on 26 October 2020).



{kind=link}
{kind=link}
| Primer | Sequence | Efficiency | Temperature | |
|---|---|---|---|---|
| Cathepsin B | Fwd Rev | 5′-TTCGATGCACGAGAAAATTG-3′ | 100.3% | 57.7 °C |
| 5′-CTCATGACGGAAGTTGCTGA-3′ | ||||
| Cathepsin L-like proteinase | Fwd Rev | 5′-AACGCAGTAGGTACCGTTGG-3′ | 101.3% | 57.1 °C |
| 5′-CAGCCAAAACACCATGATTG-3′ | ||||
| Lysosomal alpha-mannosidase | Fwd Rev | 5′-GAAATTGGTTCAGCCTTGGA-3′ | 99.8% | 58.1 °C |
| 5′-ATAGCTGCGCGTCTTCATTT-3′ | ||||
| Multidrug resistance-associated protein 4 | Fwd Rev | 5′-TCCAATGTGCGAAACAACAT-3′ | 102.1% | 57.1 °C |
| 5′-GGCAAAGGGAAATTGTTTGA-3′ | ||||
| Senecionine N-oxygenase | Fwd Rev | 5′-TGTTGTGGTTTCGACGTCAT-3′ | 103.6% | 58.1 °C |
| 5′-TACGTTGAACGGAACACCAA-3′ | ||||
| Voltage-dependent calcium channel type A subunit alpha-1 | Fwd Rev | 5′-AGCATAGCCATCCATCCTTG-3′ | 100.4% | 63.2 °C |
| 5′-GAGTCTGCGGTGTAGCATGA-3′ | ||||
| α-tubulin | Fwd Rev | 5′-GAGTTCCAGACCAACTTGGT-3′ | 107.9% | 52.6 °C |
| 5′-GCCATGTACTTGCCGTGACG-3′ | ||||
| Target Accession Number | Target Name | Log2 Fold-Change | p-Value |
|---|---|---|---|
| Spinosad/Control | |||
| LOC111514273 | Peritrophin-1-like | 2.08 | 0.046 |
| LOC111505285 | Polygalacturonase-like|glycoside hydrolase family 28 | 1.88 | 0.020 |
| LOC111505933 | Cathepsin B-like | 1.83 | 0.019 |
| LOC111509352 | Lysosomal alpha-mannosidase-like | 1.65 | 0.003 |
| LOC111507304 | Venom carboxylesterase-6-like | 1.60 | 0.036 |
| LOC111501823 | Lysosomal alpha-mannosidase-like | 1.45 | 0.041 |
| LOC111512755 | Cathepsin L-like proteinase | 1.45 | 0.039 |
| LOC111513337 | GILT-like protein 1 | 1.43 | 0.023 |
| LOC111517731 | Uncharacterized protein LOC111517731 | 1.39 | 0.032 |
| LOC111513667 | Tetratricopeptide repeat protein 39B-like | 1.38 | 0.016 |
| LOC111512533 | Probable chitinase 10 | 1.30 | 0.048 |
| LOC111510455 | Equilibrative nucleoside transporter 3-like | 1.28 | 0.041 |
| LOC111511653 | Uncharacterized protein LOC111511653 | 1.24 | 0.012 |
| LOC111512328 | Juvenile hormone esterase-like | 1.23 | 0.024 |
| LOC111513728 | Cathepsin L-like proteinase | 1.20 | 0.006 |
| LOC111503028 | Neutral alpha-glucosidase C-like | 1.16 | 0.028 |
| LOC111512041 | Venom carboxylesterase-6-like | 1.16 | 0.008 |
| LOC111505338 | Uncharacterized protein LOC111505338 | 1.13 | 0.035 |
| LOC111503007 | Multidrug resistance-associated protein 4-like | 1.02 | 0.007 |
| LOC111513768 | Juvenile hormone acid O-methyltransferase-like | 1.02 | 0.044 |
| LOC111507306 | Venom carboxylesterase-6-like | 1.01 | 0.006 |
| LOC111504230 | Uncharacterized protein LOC111504230 | −1.04 | 0.001 |
| LOC111516463 | Probable cytochrome P450 301a1, mitochondrial|cytochrome P450 12h2 | −1.07 | 0.008 |
| LOC111507554 | Uncharacterized protein LOC111507554 | −1.10 | 0.033 |
| LOC111512100 | Neuropeptide-like protein 31 | −1.20 | 0.050 |
| LOC111513879 | Lipase 3-like | −1.32 | 0.020 |
| LOC111507616 | Senecionine N-oxygenase-like | −1.86 | 0.025 |
| LOC111517073 | Putative nuclease HARBI1 | −2.24 | 0.020 |
| LOC111507833 | Voltage-dependent calcium channel type A subunit alpha-1-like | −2.83 | 0.033 |
| LOC111513727 | Cathepsin L-like proteinase | −5.13 | 0.005 |
| Target Accession Number | Target Name | Log2 Fold-Change | p-Value |
|---|---|---|---|
| Spinosad/Control | |||
| LOC111506250 | Cytochrome P450 9e2-like | 1.41 | 0.051 |
| LOC111510879 | Cytochrome P450 6k1-like | 0.92 | 0.069 |
| LOC111517755 | Probable cytochrome P450 6a23/cytochrome P450 | 0.91 | 0.026 |
| LOC111517753 | Probable cytochrome P450 6a23/cytochrome P450 6bj3 | 0.57 | 0.064 |
| LOC111503064 | Cytochrome P450 4g15 | 0.42 | 0.071 |
| LOC111505596 | Endocuticle structural glycoprotein SgAbd-4-like/putative cuticle protein CP6 | 0.39 | 0.067 |
| LOC111505534 | NADPH-cytochrome P450 reductase | 0.33 | 0.041 |
| LOC111503441 | Cytochrome P450 4c1-like | 0.31 | 0.022 |
| LOC111517496 | Endocuticle structural glycoprotein SgAbd-8-like | −0.30 | 0.095 |
| LOC111505902 | Cytochrome P450 CYP12a2-like/cytochrome P450 353a2 | −0.49 | 0.059 |
| LOC111509981 | Cytochrome P450 4c1-like | −1.06 | 0.076 |
| LOC111516463 | Probable cytochrome P450 301a1, mitochondrial/cytochrome P450 12h2 | −1.07 | 0.008 |
| miRNAs | Average Normalized Expression |
|---|---|
| Lde-miR-8-3p | 65,301.25 |
| Lde-miR-14-5p | 63,662.96 |
| Lde-miR-317-5p | 42,146.90 |
| Lde-miR-1-3p | 24,099.48 |
| Lde-bantam-3p | 14,830.90 |
| Lde-miR-281-5p | 12,820.94 |
| Lde-miR-1175-3p | 9564.56 |
| Lde-miR-34-3p | 8436.31 |
| Lde-miR-12-3p | 6915.56 |
| Lde-miR-13b-3p | 6217.84 |
| miRNAs | Log2 Fold-Change | p-Value |
|---|---|---|
| Spinosad/Control | ||
| Lde-miR-750-3p | 1.72 | 0.0034 |
| Lde-miR-2796-5p | 1.68 | 0.0190 |
| Lde-miR-3791-5p | 1.50 | 0.0333 |
| Lde-miR-750-5p | 1.07 | 0.0003 |
| Lde-bantam-5p | 0.99 | 0.0399 |
| Lde-miR-1175-3p | 0.92 | 0.0087 |
| Lde-miR-184-3p | 0.87 | 0.0045 |
| Lde-miR-281-5p | 0.67 | 0.0296 |
| Lde-miR-125-3p | 0.64 | 0.0263 |
| Lde-miR-9e-3-5p | −0.69 | 0.0071 |
| Lde-let-7-3p | −0.76 | 0.0267 |
| Lde-miR-8-3p | −0.76 | 0.0109 |
| Lde-miR-9a-5p | −0.97 | 0.0001 |
| Lde-miR-12-3p | −1.01 | 0.0057 |
| Target Accession Number | Target Name | GO IDs | GO Terms |
|---|---|---|---|
| LOC111501823 | Lysosomal alpha-mannosidase | P:GO:0044238; P:GO:0071704 | P:primary metabolic process; P:organic substance metabolic process |
| LOC111512533 | Uncharacterized protein LOC101463243 isoform X2 | P:GO:0006508; P:GO:0006897 | P:proteolysis; P:endocytosis |
| LOC111510455 | Equilibrative nucleoside transporter 1 | P:GO:1901642 | P:nucleoside transmembrane transport |
| LOC111503007 | Probable multidrug resistance-associated protein lethal(2)03659 | P:GO:0001666; P:GO:0006979; P:GO:0008340; P:GO:0034059; P:GO:0048190 | P:response to hypoxia; P:response to oxidative stress; P:determination of adult lifespan; P:response to anoxia; P:wing disc dorsal/ventral pattern formation |
| LOC111516463 | Probable cytochrome P450 301a1, mitochondrial | P:GO:0055114; P:GO:0007490 | P:oxidation-reduction process; P:tergite morphogenesis |
| LOC111513879 | Lipase 1 | P:GO:0006629; P:GO:0016042 | P:lipid metabolic process; P:lipid catabolic process |
| LOC111507833 | Voltage-dependent calcium channel type A subunit alpha-1 isoform X14 | P:GO:0006811 | P:ion transport |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bastarache, P.; Wajnberg, G.; Dumas, P.; Chacko, S.; Lacroix, J.; Crapoulet, N.; Moffat, C.E.; Morin, P., Jr. Transcriptomics-Based Approach Identifies Spinosad-Associated Targets in the Colorado Potato Beetle, Leptinotarsa decemlineata. Insects 2020, 11, 820. https://doi.org/10.3390/insects11110820
Bastarache P, Wajnberg G, Dumas P, Chacko S, Lacroix J, Crapoulet N, Moffat CE, Morin P Jr. Transcriptomics-Based Approach Identifies Spinosad-Associated Targets in the Colorado Potato Beetle, Leptinotarsa decemlineata. Insects. 2020; 11(11):820. https://doi.org/10.3390/insects11110820
Chicago/Turabian StyleBastarache, Pierre, Gabriel Wajnberg, Pascal Dumas, Simi Chacko, Jacynthe Lacroix, Nicolas Crapoulet, Chandra E. Moffat, and Pier Morin, Jr. 2020. "Transcriptomics-Based Approach Identifies Spinosad-Associated Targets in the Colorado Potato Beetle, Leptinotarsa decemlineata" Insects 11, no. 11: 820. https://doi.org/10.3390/insects11110820