The Rare Richardsitas Betsch (Collembola, Symphypleona, Sminthuridae): A New Species from Australia with Comments on the Genus and on the Sminthurinae




Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Taxonomic Summary and Genus Diagnosis
- Order Symphyleona Börner, 1901 [29]
- Superfamily Sminthuroidea Bretfeld, 1994 [30]
- Family Sminthuridae Lubbock, 1862 [1]
- Subfamily Sminthurinae Lubbock, 1862 [1]
- Genus Richardsitas Betsch, 1975 [15]
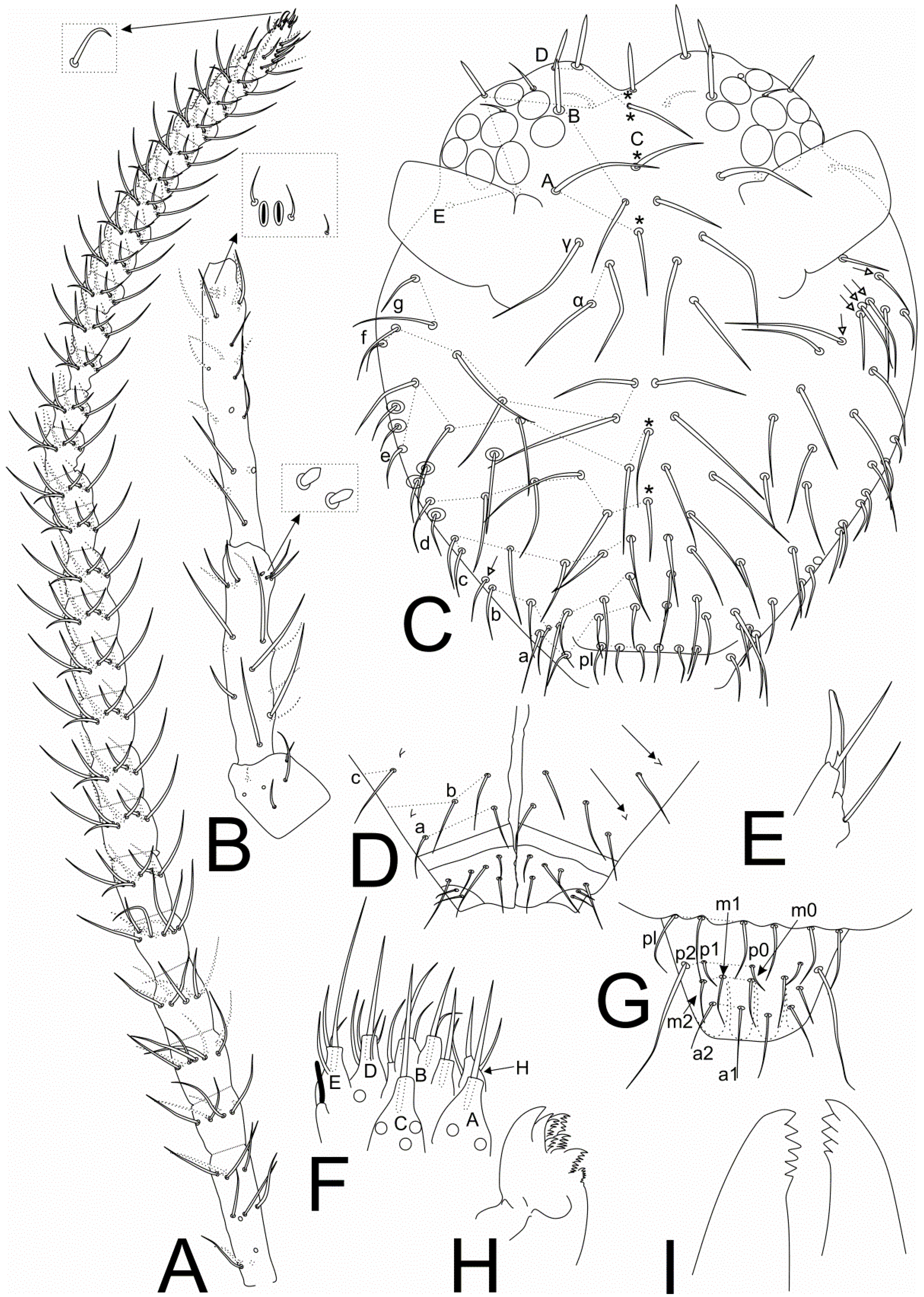
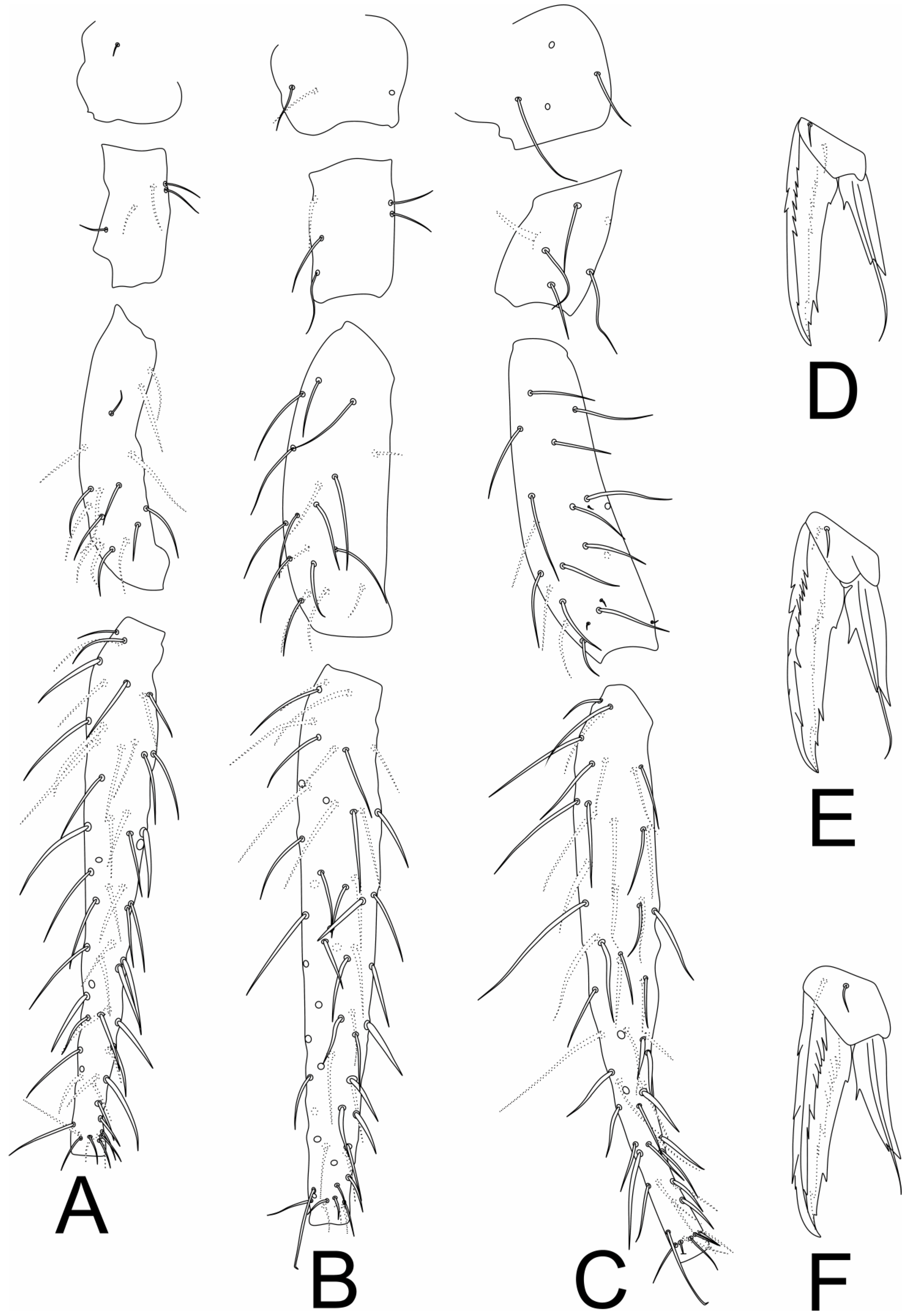
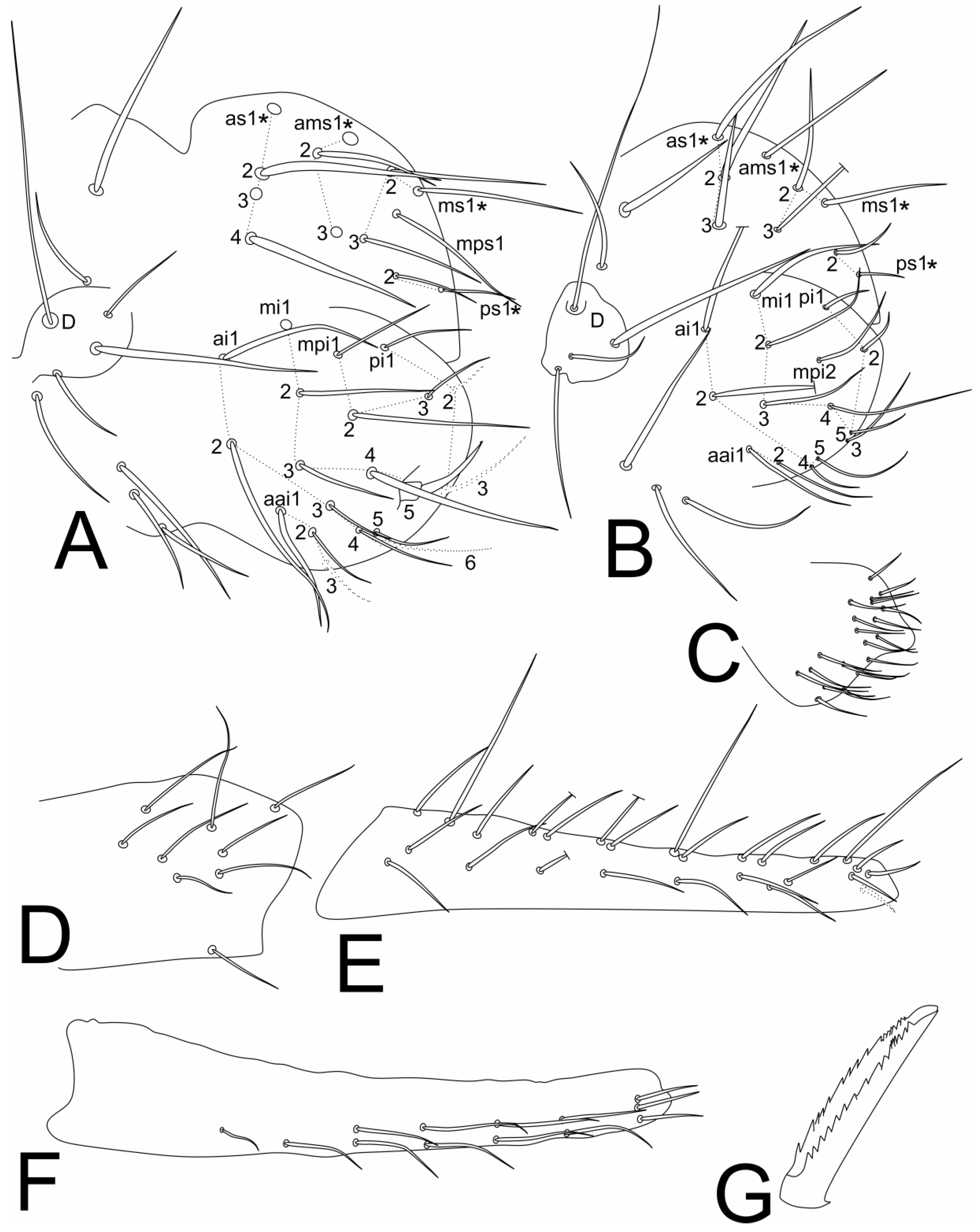
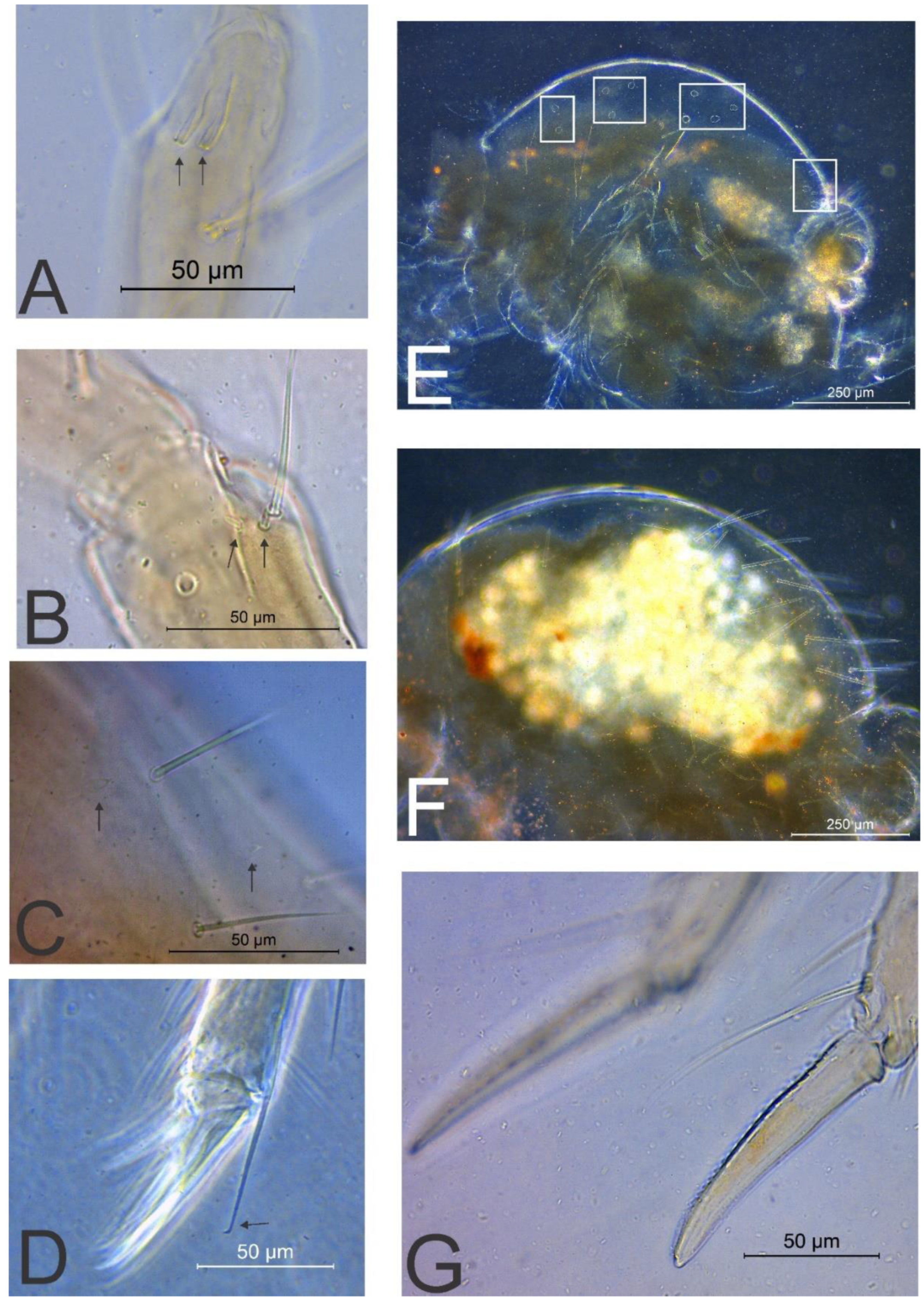
3.2. Richardsitas subferoleum sp. Nov.
3.3. Identification Key and Distribution of Richardsitas Species
- Mucronal chaeta absent; Ant IV with 28 subsegments; Ant II with 2 dorso-apical short sensilla; males with 16–17 short candle-shaped chaetae distributed in 4 zones of dorsal large abdomen … Richardistas subferoleum sp. nov.; Australia‒Mucronal chaeta present; Ant IV with 30 subsegments; Ant II lacking modified short sensilla; males with about 26 or more short candle-shaped or spine-like chaetae distributed in 3–4 zones of the dorsal large abdomen … 2
- Males with about 26 short candle-shaped chaetae distributed in 3 zones of the dorsal large abdomen … R. griveaudi Betsch, 1977 [16]; Madagascar‒Males with about 58 short candle-shaped or spine-like chaetae distributed in 4 zones of dorsal large abdomen … R. najtae Betsch, 1975 [15]; Madagascar
4. Discussion
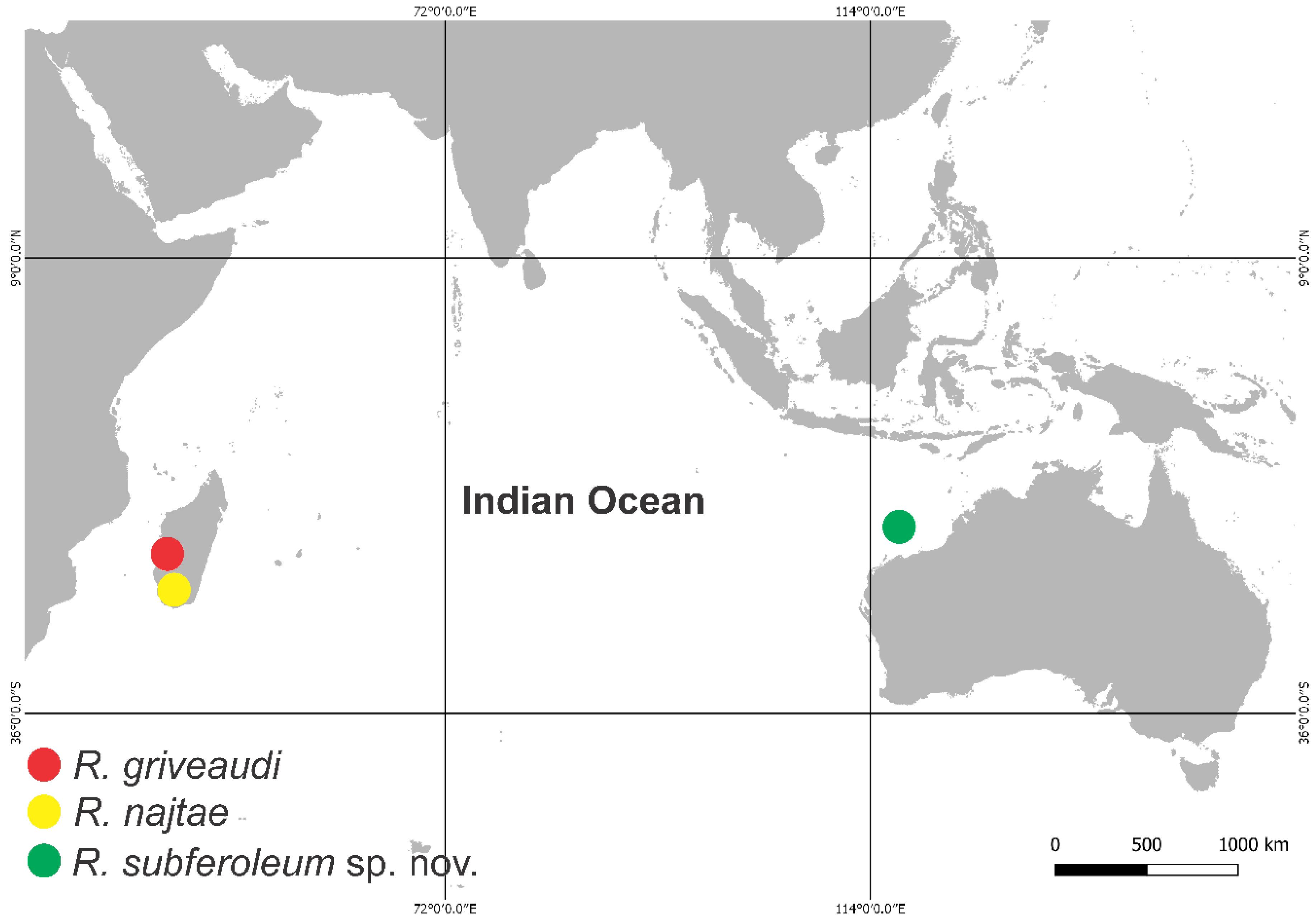
4.1. Remarks on the Distribution and Morphology of the Richardsitas Species
4.2. Remarks on Some Sminthurinae Genera
4.3. Identification Key and Distribution of Extant Sminthurinae Genera
- Metatrochanteral spine absent ... 2–Metatrochanteral spine present ... 4
- Capitate tenent-hairs absent on tibiotarsi ... Sminthurus Latreille, 1802 [19]; Holarctic*–Capitate tenent-hairs present on tibiotarsi ... 3
- Female’s subanal appendage short, chaeta-like, oval or leaf-like; male’s genital plate with normal granules ... Spatulosminthurus Betsch and Betsch-Pinot, 1984 [21]; Palaearctic–Female’s subanal appendage long and chaeta-like; male’s genital plate with short cuticular points … Janusius Bretfeld, 2010 [34]; Holarctic
- D bothriotrichium ciliate ... Austrosminthurus Delamare Deboutteville and Massoud, 1963** [17]; Argentina–D bothriotrichium smooth ... 5
- A single neosminthuroid chaeta present on parafurcal area; males with two horn-like chaetae on apical Ant III ... Keratosminthurus Zeppelini, 2020 [9]; Brazil–Neosminthuroid chaeta on parafurcal area absent; males lacking horn-like chaetae on apical Ant III ... 6
- Male’s dorsal large abdomen with 3–4 zones of short candle-shaped or spine-like chaetae; one capitate tenent-hair present on tibiotarsi II–III ... Richardsitas Betsch, 1975 [15]; Madagascar, Australia–Male’s dorsal large abdomen without zones of modified chaetae; tibiotarsi without tenent-hairs ... 7
- Postantennal chaeta present; dorsal head and large abdomen with long rough often blunt chaetae; large abdomen posteriorly with one pair of cuticular glands ... Allacma Börner, 1906 [36]; Holarctic–Postantennal chaeta absent; dorsal head and large abdomen chaetotaxy with regular and/or spine-like chaetae; large abdomen posteriorly without cuticular glands ... 8
- Large abdomen dorso-posterior spines present ... 9–Large abdomen dorso-posterior spines absent ... 10
- Ant IV with 15–18 subsegments; head and bothriotrichia areas of cuticle with complex girandole-like granules ... Caprainea Dallai, 1970 [37]; Palaearctic–Ant IV mostly with 9–14 subsegments, rarely with 15; head and body cuticle without remarkable different granules … Pararrhopalites Bonet and Tellez, 1947 [22]; Holotropical
- Ant IV with 13–15 subsegments; males with a well-developed clasping organ on ventral Abd VI ... Novokatianna Salmon, 1944 [20]; New Zealand–Ant IV with 18 or more subsegments; males devoid of a clasping organ on ventral Abd VI ... 11
- Eyes and body pigment absent; cave species ... Galeriella Ćurčić and Lučić, 2007 [18]; Bosnia-Herzegovina–8+8 eyes present, specimens pigmented; surface species ... Temeritas Richards, 1963 [17]; Holotropical
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lubbock, J. Notes on the Thysanura pt. I-Sminthuridae. Trans. Linn. Soc. Lond. 1862, 23, 429–448. [Google Scholar] [CrossRef]
- Bretfeld, G. Synopses on Palaearctic Collembola, Volume 2. Symphypleona. Abh. Ber. Naturkundemus. Görlitz 1999, 71, 1–318. [Google Scholar]
- Cipola, N.G.; Silva, D.D.; Bellini, B.C. Class Collembola. In Keys to Neotropical Hexapoda-Thorp and Covich’s Freshwater Invertebrates, 4th ed.; Hamada, N., Thorp, J.H., Rogers, D.C., Eds.; Elsevier Academic Press: London, UK, 2018; Volume 3, pp. 11–55. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 20 May 2020).
- Betsch, J.M. Éléments pour une monographie des Collemboles Symplyplêones (Hexapodes, Aptérygotes). Mém. Mus. Natl. Hist. Nat. Sér. A Zool. 1980, 116, 1–227. [Google Scholar]
- Christiansen, K.; Pike, E. Cretaceous Collembola (Arthropoda, Hexapoda) from the Upper Cretaceous of Canada. Cretac. Res. 2002, 23, 165–188. [Google Scholar] [CrossRef]
- Christiansen, K.; Nascimbene, P. Collembola (Arthropoda, Hexapoda) from the mid Cretaceous of Myanmar (Burma). Cretac. Res. 2006, 27, 318–363. [Google Scholar] [CrossRef]
- Sánchez-García, A.; Engel, M.S. Long-term stasis in a diverse fauna of Early Cretaceous springtails (Collembola: Symphypleona). J. Syst. Palaeontol. 2016, 15, 513–537. [Google Scholar] [CrossRef]
- Zeppelini, D.; Brito, R.A.; Zampaulo, R.; Lima, E.C. A new highly dimorphic genus of Sminthuridae (Collembola: Symphypleona) from Brazil. Zootaxa 2020, 4729, 25–46. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Naturæ perR tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis, 10th ed.; Impensis Direct Laurentii Salvii: Stockholm, Sweden, 1758; pp. 1–824. [Google Scholar] [CrossRef] [Green Version]
- Womersley, H. Notes on some Australian Collembola. Stylops 1934, 3, 244–246. [Google Scholar] [CrossRef]
- Womersley, H. Primitive insects of South Australia: Silverfish, Springtails, and their Allies. In Handbook of the Flora and Fauna of South Australia, 1st ed.; Government Printer: Adelaide, Australia, 1939; pp. 1–322. [Google Scholar]
- Greenslade, P. The rare collembolan genus, Temeritas (Symphypleona: Sminthuridae), in Southern Australia: Systematics, distribution and conservation status. Vic. Nat. 2013, 130, 45–49. [Google Scholar]
- Greenslade, P. Collembola. In Zoological Catalogue of Australia, 1st ed.; Walton, D.W., Ed.; CSIRO: Melbourne, Australia, 1994; Volume 22, pp. 19–184. [Google Scholar]
- Betsch, J.M. Étude des Collemboles de Madagascar IV—Deux nouveaux genres de Symphypléones à dimorphisme sexuel important: Parabourletiella et Richardsitas. Rev. Ecol. Biol. Sol. 1975, 12, 477–485. [Google Scholar]
- Betsch, J.M. Étude des Collemboles de Madagascar V—Sur deux Symphypléones de la forét séche en secteur bioclimatique subaride. Bull. Soc. Entomol. Fr. 1977, 82, 119–125. [Google Scholar]
- Delamare-Deboutteville, C.; Massoud, Z. Collemboles symphypléones. In Biologie de L’Amérique Australe, 1st ed.; Delamare-Deboutteville, C., Rapoport, E., Eds.; Éditions du CNRS: Paris, France, 1963; Volume 2, pp. 169–289. [Google Scholar]
- Ćurčić, B.P.M.; Lučić, L.R.; Tomić, V.T.; Makarov, S.E.; Karaman, I.M. Galeriella liciniana, a new cave genus and species of springtails (Collembola, Sminthuridae) from Herzegovina. Arch. Biol. Sci. 2007, 59, 63–64. [Google Scholar] [CrossRef]
- Latreille, P.A. Histoire naturelle, générale et particulière des Crustacés et des Insectes III. F. Dufart Paris 1802, 15, 69–83. [Google Scholar]
- Salmon, J.T. New Genera, species and records of New Zealand Collembola and a discussion of Entomobrya atrocincta Schött. R. Dominion Mus. 1944, 1, 123–182. [Google Scholar]
- Betsch, J.M.; Betsch-Pinot, M.C. Contribution à l’étude des Sminthurus (Collembola, Symphypleona). Ann. Soc. R. Zool. Belg. 1984, 114, 71–81. [Google Scholar]
- Bonet, F.; Tellez, C. Un Nuevo Genero de Esminturidos (Collembola). Rev. Soc. Mex. Hist. Nat. 1947, 8, 193–203. [Google Scholar]
- Zeppelini, D.; Brito, R.A. Two new species of Pararrhopalites (Collembola: Symphypleona: Sminthuridae) in Brazil. Fla. Entomol. 2014, 97, 1733–1744. [Google Scholar] [CrossRef]
- Medeiros, G.S.; Bellini, B.C. Two new species of Temeritas Richards (Collembola, Symphypleona, Sminthuridae) from Neotropical Region, with coments on the genus. Zootaxa 2019, 4586, 536–552. [Google Scholar] [CrossRef]
- Betsch, J.M.; Waller, A. Chaetotaxic nomenclature of the head, thorax and abdomen in Symphypleona (Insecta, Collembola). Acta Zool. Fenn. 1994, 195, 5–12. [Google Scholar]
- Betsch, J.M. An ontogenetically focused chaetotaxial scheme in Symphypleona (collembolan): The 6th abdominal segment. Pedobiologia 1997, 41, 13–18. [Google Scholar]
- Fjellberg, A. The Labial Palp in Collembola. Zool. Anz. 1999, 237, 309–330. [Google Scholar]
- Cipola, N.G.; Morais, J.W.; Bellini, B.C. A new species of Seira (Collembola: Entomobryidae: Seirini) from Northern Brazil, with the addition of new chaetotaxic characters. Zoologia 2014, 31, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Börner, C. Neue Collembolenformen und zur Nomenclatur der Collembola Lubbock. Zool. Anz. 1901, 24, 696–712. [Google Scholar]
- Bretfeld, G. The chaetotaxy of the small abdomen of the Symphypleona (Insecta, Collembola) and its phylogenetic interpretation. Acta Zool. Fenn. 1994, 195, 13–17. [Google Scholar]
- Lynch, A.J.J.; Beeton, R.J.S.; Greenslade, P. The conservation significance of the biota of Barrow Island, Western Australia. J. R. Soc. West. Aust. 2019, 102, 98–133. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Christiansen, K.A.; Bellinger, P. Biogeography of Hawaiian Collembola: The simple principles and complex reality. Orient. Insects 1994, 28, 309–351. [Google Scholar] [CrossRef]
- Bretfeld, G. Fifth report on Symphypleona from Russia, and also a review of Deuterosminthurus kaplini Martynova, 1979 from Turkmenistan (Insecta, Collembola). Soil Org. 2010, 82, 301–316. [Google Scholar]
- Bernard, C.E.; Wynne, J.J. Disparrhopalites naasaveqw n. sp. from caves at Wupatki National Monument, Arizona, synonymy of Dietersminthurus Palacios-Vargas, Cuéllar & Vázquez, 1998 with Disparrhopalites Stach, 1956 and compositionof Songhaicinae (Collembola: Sminthuridae). Zootaxa 2017, 4319, 77–90. [Google Scholar] [CrossRef]
- Börner, C. Das System der Collembolen nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Mit. Natur. Mus. Hamburg 1906, 23, 147–188. [Google Scholar]
- Dallai, R. Investigations on Collembola: 10. Examination of the cuticle in some species of the tribe Sminthurini Börner, 1913, by means of the scanning electron microscope. Monit. Zool. Ital. 1970, 4, 41–53. [Google Scholar]
- Dányi, L.; Park, K.H. A new species of Janusius Bretfeld, 2010 (Collembola, Sminthuridae) from the Korean Peninsula, with a key to the species of the genus. Zootaxa 2016, 4154, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Baijal, H.H.N.; Agarwal, N. On new species of the genus Pararrhopalites Bonet and Telbez (Collembola: Arrhopalitini) from India. Zool. Anz. 1972, 189, 94–96. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Colour | Ant IV Subsegments | Ant II ap. Sens | Dorsal Abdominal Zones of Modified Chaetae * | Heterogeneity of Abdominal Spines * | Number of Abdominal Spines * | Dens Dorsal (Posterior) Chaetae | Mucronal Chaeta |
|---|---|---|---|---|---|---|---|---|
| R. griveaudi [5,16] | pink to red | 30 | - | 3 | + | 26 | ? | + |
| R. najtae [5,15] | red | 30 | - | 4 | + | 58 | 26 | + |
| R. subferoleum sp. nov. | pink | 28 | + | 4 | - | 16–17 | 26 | - |
| Genera | Ant IV Subsegments | Eyes | Head Sexual Dimorphism | Post Antennal Chaeta | Abdominal Dorso-Anterior Spines | Abdominal Dorso-Posterior Spines | Dorsal Large Abdomen Sexual dimorphism | D Bothriotrichium | Neosminthuroid Chaetae on Large abdomen | Metatrochanteral Spine | Capitate Tenent-Hairs | Ungual Tunica | Dens Ventral Chaetae | Mucronal Chaeta |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Galeriella ∆ [18] | 32 | - | - | - | - | - | - | Smooth | - | + | - | - | ? | - |
| Keratosminthurus ∆ [9] | 18–20 | + | + | - | - | - | - | Smooth | + | + | - | +/- | 12–13 | - |
| Richardsitas ∆ [5,15,16] | 28–30 | + | - | - | + | + | + | Smooth | - | + | + | - | 13 | +/- |
| Temeritas ∆ [5,17,24] | 18–46 | + | - | - | +/- | - | -* | Smooth | - | + | - | +/- | 13 | +/- |
| Allacma [2,36] | 12–15 | + | - | + | - | - | - | Smooth | - | + | - | +/- | 11–15 | + |
| Austrosminthurus [5,17] *** | ? | + | -? | - | ? | - | -? | Ciliate | -? | + | - | + | 13 | + |
| Caprainea [2,37] | 15–18 | + | - | - | + | + | - | Smooth | - | + | - | + | About 13 | +/- |
| Janusius [34,38] | 18–26 | + | +/- | +/- | - | - | - | Smooth | - | - | + | - | 9–15 | +/- |
| Novokatianna [5,20] | 13–15 | + | - | - | - | - | - | Smooth | - | + | - | ? | ? | + |
| Pararrhopalites [2,22,23] ** | 9–15 | +/- | - | - | +/- | + | - | Smooth | - | + | - | +/- | 7–14 | +/- |
| Sminthurus [2,5] | 14–30 | + | +/- | +/- | - | - | - | Smooth | - | - | - | +/- | About 15 | +/- |
| Spatulosminthurus [2,21] | 14–25 | + | +/- | +/- | - | - | - | Smooth | - | - | + | +/- | 13–15 | +/- |
| Archeallacma † [8] | 14–15 | + | ? | ? | - | - | ? | ? | ? | ? | - | + | ? | - |
| Brevimucronus † [6] | - | + | ? | ? | ? | ? | ? | ? | ? | -? | + | - | ? | -? |
| Grinnellia † [7] | 10 | + | -? | -? | + | + | ? | Smooth | -? | + | - | +/- | ? | -? |
| Katiannasminthurus † [8] | 14 | + | ? | ? | + | + | ? | ? | -? | ? | - | + | ? | - |
| Mucrovirga † [7] | 9–10 | + | ? | ? | +? | +? | ? | ? | ? | ? | + | - | ? | -? |
| Sminthurconus † [7] | 12 | + | ? | ? | + | + | ? | ? | ? | + | -? | - | ? | -? |
| Sminthuricinus † [7] | 11–12 | + | ? | ? | + | + | ? | Smooth? | ? | + | + | - | ? | -? |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medeiros, G.d.S.; Greenslade, P.; Bellini, B.C. The Rare Richardsitas Betsch (Collembola, Symphypleona, Sminthuridae): A New Species from Australia with Comments on the Genus and on the Sminthurinae. Insects 2020, 11, 519. https://doi.org/10.3390/insects11080519
Medeiros GdS, Greenslade P, Bellini BC. The Rare Richardsitas Betsch (Collembola, Symphypleona, Sminthuridae): A New Species from Australia with Comments on the Genus and on the Sminthurinae. Insects. 2020; 11(8):519. https://doi.org/10.3390/insects11080519
Chicago/Turabian StyleMedeiros, Gleyce da S., Penelope Greenslade, and Bruno C. Bellini. 2020. "The Rare Richardsitas Betsch (Collembola, Symphypleona, Sminthuridae): A New Species from Australia with Comments on the Genus and on the Sminthurinae" Insects 11, no. 8: 519. https://doi.org/10.3390/insects11080519