Male-biased Adult Production of the Striped Fruit Fly, Zeugodacus scutellata, by Feeding dsRNA Specific to Transformer-2

Department of Plant Medicals, Andong National University, Andong 36729, Korea
*
Author to whom correspondence should be addressed.
Insects 2020, 11(4), 211; https://doi.org/10.3390/insects11040211
Submission received: 3 March 2020
/
Revised: 20 March 2020
/
Accepted: 27 March 2020
/
Published: 28 March 2020
Abstract
:Sterile insect release technique (SIT) is effective for eradicating quarantine insects including various tephritid fruit flies. When SIT is used for fruit flies, it is challenging to remove females from sterile males due to oviposition-associated piercing damage. This study developed a sex transition technique by feeding double-stranded RNA (dsRNA) specific to a sex-determining gene, Transformer-2 (Zs-Tra2) of the striped fruit fly, Zeugodacus scutellata. Zs-Tra2 is homologous to other fruit fly orthologs. It is highly expressed in female adults. RNA interference (RNAi) of Zs-Tra2 by injecting or feeding its specific dsRNA to larvae significantly increased male ratio. Recombinant Escherichia coli cells expressing dsRNA specific to Zs-Tra2 were prepared and used to feed larvae to suppress Zs-Tra2 gene expression levels. When these recombinant bacteria were fed to larvae during the entire feeding stage, the test population was significantly male-biased. Some females treated with such recombinant E. coli exhibited mosaic morphological characters such as the presence of male-specific abdominal setae in females. This study proposes a novel technique by feeding dsRNA specific to Transformer-2 to reduce female production during mass-rearing of tephritid males for SIT.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
A number of tephritid fruit flies are designated as quarantine pests in many countries [1]. These fruit flies are classified into the Tephritoidea superfamily of about 7300 species [2] with two possible origins based on molecular phylogenetic analysis [3]. In Tephritoidea, the largest family is Tephritidae, which possesses more than 4400 species [4,5]. In Korea, 90 tephritid species have been identified, of which two species (Zeugodacus depressa and Z. scutellata) are regarded as insect pests [6].
The striped fruit fly, Z. scutellata, occurs in Asian countries such as Korea, Japan, Taiwan, China, Bhutan, Thailand, India, and Malaysia [7]. Its outbreak can cause more than 70% damage to pumpkin flowers, thus posing serious threat to crop production [8]. Chinese cucumber (Trichosanthes kirilowii) is the main wild host of Z. scutellata. Its alternative hosts include wild gourds (Diplocyclos palmatus, Zehneria liukiuensis, and Trichosanthes spp.) [9,10,11]. Although chemical and biological control tactics have been developed to control fruit fly populations, sterile insect release technique (SIT) is known to be effective in eradicating the fruit flies [12].
SIT is a genetic control method of specific target insect species using a massive release of sterile males to compete with wild males in mating with females. In addition, females mated with sterile males will produce no offspring [13]. In a SIT program, one obstacle is sex sorting after mass-rearing to release only males [14]. A genetic sexing line using chromosomal translocating technology was generated to facilitate the sorting of females by exhibiting different colored pupae of the melon fly, Z. cucurbitae, in which brown pupae are males while white pupae are females [15]. For Mediterranean fruit flies, Ceratitis capitata, a transgenic approach has been used to develop an effective genetic sexing line with females killed at the early embryonic stage by adding transgenic apoptotic genes controlled by female-specific gene expression [16]. However, classical genetic or transgenic sexing systems have practical limitations, such as the extra labor needed for sorting pupae or genetically modified organism issue. To overcome these obstacles, a novel molecular approach focusing on manipulation of the sex determination system of insects is needed.
Like sex determination systems known in other animals, insect sex is determined by sex chromosomes. However, to be differentiated into males and females, chromosomal information is interpreted by sex-determining genes of doublesex (Dsx) for expressing sex-specific morphological characters and fruitless (Fru) for expressing sex-specific behavior. These genes have been well characterized in Drosophila [17]. Briefly, X-linked signal at XX females can activate Sex lethal (Sxl) which is a sex determination master gene. Subsequent functional Sxl protein product can help make a functional transformer (Tra) protein by preventing alternative splicing generating premature stop codon. The functional Tra protein can bind to transformer-2 (Tra-2), an RNA-binding protein. This complex allows female-specific alternative splicing of Dsx mRNA. In contrast, males without an X-linked signal cannot form a functional Sxl protein. Thus, they will fail to make functional Tra proteins, leading to no Tra/Tra-2 protein complex and male-specific alternative splicing of Dsx mRNA. Tephritids such as C. capitate have been well studied. They do not have the Sxl. However, Dominant Male Determiner (M) on the Y chromosome is presumed to inhibit Tra protein which in turn prevents the formation of female-specific Tra mRNA [18]. Tra-2 is required for female development because the RNA interference (RNAi) of Tra-2 expression in XX females can lead to phenotypic males of C. capitata [19]. When this RNAi technique against Tra-2 expression was applied to eggs of the Oriental fruit fly, Bactrocera dorsalis, it produced almost 100% males [20]. Similar approach has been applied to the pumpkin fruit fly (Z. tau), resulting in male-biased sex ratio and some intersexes [21]. However, egg injections of double-strand RNA are a practical limitation to produce males only in mass-rearing for SIT.
To overcome such a practical obstacle of RNAi strategy, this study used a feeding technique for dsRNA against Tra-2 identified from Z. scutellata (Zs-Tra2). To analyze its function, RNAi was performed on young larvae by injecting dsRNA specific to Zs-Tra2. To be practical, dsRNA was fed to Z. scutellata to determine its RNAi efficiency. To continuously and economically feed dsRNA, a recombinant Escherichia coli expressing dsRNA specific to Zs-Tra2 was constructed and supplied to feed for larvae. This study provides a novel technology for the production of male-biased fruit flies for SIT.
2. Materials and Methods
2.1. Insect Rearing and Morphological Characters for Sex Determination
Z. scutellata larvae were collected from a pumpkin field in Andong, Korea and reared on a larval artificial diet consisting of 1 g agar, 5 g pumpkin powder, 5 g yeast, 4 g wheat bran, 5 g sugar, 4 g casein, 1 g citric acid, 0.5 g sodium benzoate, 0.1 g methyl-p-hydroxybenzoate, and 0.02 g streptomycin in 73.38 mL water. Laboratory conditions were: temperature of 26 °C, relative humidity of 70–75%, and photoperiod of 14:10 h (L:D). Under these laboratory conditions, larvae underwent three instars (L1–L3) for approximately 4.5 days. Fresh pumpkin stems and flowers were provided in adult cages for oviposition. Adults were fed an adult artificial diet (2 g sucrose, 1 g yeast, and 1 g casein). Laid eggs were collected and transferred to larval artificial diet. For pupation, a larval growth box was provided with sand. Pupae were kept in the box under laboratory conditions until adults emerged. Female and male characters of adults were externally determined with oviposition and abdominal setae, respectively (Figure S1). Internally, ovary and testis were observed to clarify the sex.
2.2. Sequence Analysis
Tra-2 sequence alignment and similarity analysis was performed using the BLAST program on the NCBI website (http://www.ncbi.nlm.nih.gov). Phylogenetic tree of Tra-2 was constructed using MEGA6 program. Bootstrapping values were obtained with 1000 repetitions to support branch and gene clusters. Pfam (http://pfam.xfam.org) was used to predict protein domain structure. Exon and intron region of Tra-2 were predicted using the Spidey program (http://www.ncbi.nlm.nih.gov/spidey/).
2.3. RNA Extraction and cDNA Preparation
Total RNAs were extracted from all developmental stages (30 eggs, 10 young larvae (L1–L2), 5 L3 larvae, one pupa, or one adult) using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instruction. Extracted RNA was treated with DNase to remove genomic DNA (gDNA) contamination and confirmed no gDNA by no PCR product using the extracted RNA as template. After RNA extraction, RNA was resuspended in nuclease-free water and quantified using a spectrophotometer (NanoDrop, Thermo Scientific, Wilmington, DE, USA). Extracted RNA (1 µg) was then used for cDNA synthesis with RT PreMix (Intron Biotechnology, Seoul, Korea) containing oligo dT primer according to the manufacturer’s instruction.
2.4. RT-PCR and RT-qPCR
RT-PCR was conducted using DNA Taq polymerase (GeneALL, Seoul, Korea) with an initial heat treatment at 94 °C for 2 min followed by 35 cycles of DNA denaturation at 94 °C for 1 min, primer annealing at 50 °C, extension at 72 °C for 1 min. A final chain extension step was performed at 72 °C for 10 min. All gene expression levels in this study were determined using a real-time PCR machine (Step One Plus Real-Time PCR System, Applied Biosystem, Singapore) under the guideline of Bustin et al. [22]. RT-qPCR was conducted with a reaction volume of 20 µL containing 10 µL of Power SYBR Green PCR Master Mix (Thermo Scientific Korea), 5 µL of cDNA template (50 ng), 1 µL of forward primer, and 1 µL of reverse primer (Table S1). After 10 min of an initial denaturation step, qPCR was performed with 40 cycles of denaturation at 94 °C for 1 min, annealing at 50 °C for 1 min, and extension at 72 °C for 1 min. Fluorescence emitted from the newly synthesized PCR products at every cycle was monitored to quantify PCR products. Melting curves of PCR products were analyzed to confirm single product. Expression level of actin as reference was used to normalize target gene expression level after different treatments. Quantitative analysis was performed using the comparative CT (2−ΔΔCT) method [23].
2.5. Preparation of Double Stranded RNA (dsRNA) with In Vitro Transcription
Template DNA was amplified with gene-specific primers (Table S1) containing T7 promoter sequence (5′-TAATACGACTCACTATAGGGAGA-3′) at their 5′ ends. dsRNA was prepared using a Megascript RNAi Kit (Ambion, Austin, TX, USA) according to the manufacturer’s instruction. The resulting PCR product was used to in vitro to synthesize dsRNA specific to Zs-Tra2 using T7 RNA polymerase with NTP mixture at 37 °C for 4 h. The synthesized dsRNA was purified with a filter cartridge (10051G2, Thermo Fisher Scientific, Vilnius, Lithuania: 10 kDa cut-off size) and resuspended with deionized water.
2.6. Injection or Feeding Treatment of dsRNA to Young Larvae
For the dsRNA injection, 0.3 µL (500 ng/μL) of purified dsRNA was injected to early L1 (<12 h) larval hemocoel with a microinjector (PV830, World Precision Instruments, Sarasota, FL, USA). Treated larvae were then provided with larval artificial diet ad libitum and reared under laboratory conditions until pupation. As dsRNA control (dsCON), dsRNA specific to a viral gene, CpBV-ORF302, was prepared using the method of Park and Kim [24].
For feeding dsRNA, 10 µL (500 ng/μL) of purified dsRNA was mixed with 250 mg of artificial diet. The treated diet was then used to feed five larvae at early L1. At 48 h after feeding, treated larvae were fed with untreated fresh artificial diet and maintained until pupation. The control larvae were fed diet mixed with dsCON.
2.7. Preparation of dsRNA with Recombinant Bacteria
L4440 vector was kindly provided by Seung Jae Lee (Pohang University of Science and Technology, Pohang, Korea). Multiple cloning sites were flanked at both ends with T7 promoters in an inverted orientation. PCR primers used in in vitro dsRNA preparation were used to amplify DNA fragments followed by digestion with restriction enzymes of Hind III and Xba I. Digested DNA fragments were ligated to the L4440 vector. A recombinant vector was used to transform competent cells of Escherichia coli HT115 (DE3) lacking RNase III by electroporation. Postive clones were selected on ampicillin Luria–Bertani (LB) medium containing 100 μg/mL ampicillin (AMP) and confirmed by sequencing the insert fragment.
To produce dsRNA from recombinant bacteria, a single colony was picked and inoculated to AMP-LB medium followed by incubation at 37 °C with shaking (250 rpm) for 16 h. To the cultured broth (5 mL), 500 mL of fresh LB medium containing 100 μg/mL ampicillin was then added followed by incubation at 37 °C until bacteria reached their exponential growth phase with OD600 = 0.6~0.7. Expression of T7 polymerase was induced by adding 0.4 mM (final concentration) of IPTG. Bacteria were then incubated at 37 °C for 4 h. These cultured cells were harvested by centrifugation at 7000× g for 30 min.
2.8. Quantification of dsRNA Produced from Recombinant Bacteria
Total bacterial RNA was extracted using RNA extraction mini kit (Qiagen Korea, Seoul, Korea). The dsRNA was confirmed by electrophoresis on 1% agarose gel. Quantification of dsRNA amounts of recombinant bacteria followed the method of Kim et al. [25]. A standard curve was generated with known amounts of purified dsRNA synthesized by in vitro transcription described above. RNA band intensity was quantified with an image analyzer (Image Lab Software, Bio-Rad Korea, Seoul, Korea).
2.9. Pretreatment of Recombinant Bacteria Using Heat and Sonication
After overexpression and re-suspending bacterial cells in distilled water, two sequential pre-treatments were applied before feeding larvae. Bacterial cells were killed by heat treatment at 95 °C for 10 min. Bacterial cell walls were then disrupted with an ultrasonicator (Bandelin Sonoplus, Berlin, Germany) at 95% intensity with 10 cycles of 10 min burst. Bacterial viability was assessed by plating 100 μL of treated bacterial sample on LB plate containing ampicillin (100 μg/mL) to confirm no bacterial growth after heat treatment.
2.10. Feeding Assay of Recombinant Bacteria Expressing dsRNA
Recombinant bacteria (10 µL, 6.5 × 10 8 cells/mL) were mixed with 50 mg of larval artificial diet. Treated diet was used to feed five larvae at early L1. Every 48 h, the diet was replaced with a freshly treated diet. As a control, non-recombinant HT115 bacteria were used to treat the diet. Each treatment was replicated three times. For each replication, 100 larvae were used.
2.11. Statistical Analysis
All studies were performed with three independent replications. Results are presented as mean ± standard deviation and plotted with Sigma plot (Systat Software, San Jose, CA, USA). Means were compared by least square difference (LSD) test of one-way analysis of variance (ANOVA) using SAS program [26]. Significance was discriminated at Type I error = 0.05.
3. Results
3.1. Morphological Characters of Female and Male Z. scutellata Adults
Female adults had significantly (p < 0.05) longer body length than males (Figure S1A) even when the ovipositor was not included. Both female and male Z. scutellata adults had six visible abdominal segments. Males had 12–18 long pleural setae on the third abdominal segment (Figure S1B). Reproductive organs such as ovary and testis were recognized just after adult emergence (Figure S1C). In females, a pair of ovaries were connected to oviducts ending with ovipositor while a pair of testes were connected to vas deferens which were linked to the ejaculatory duct along with the accessory gland in males.
3.2. Prediction of Transformer-2 (Zs-Tra2) of Z. Scutellata and Expression Profile
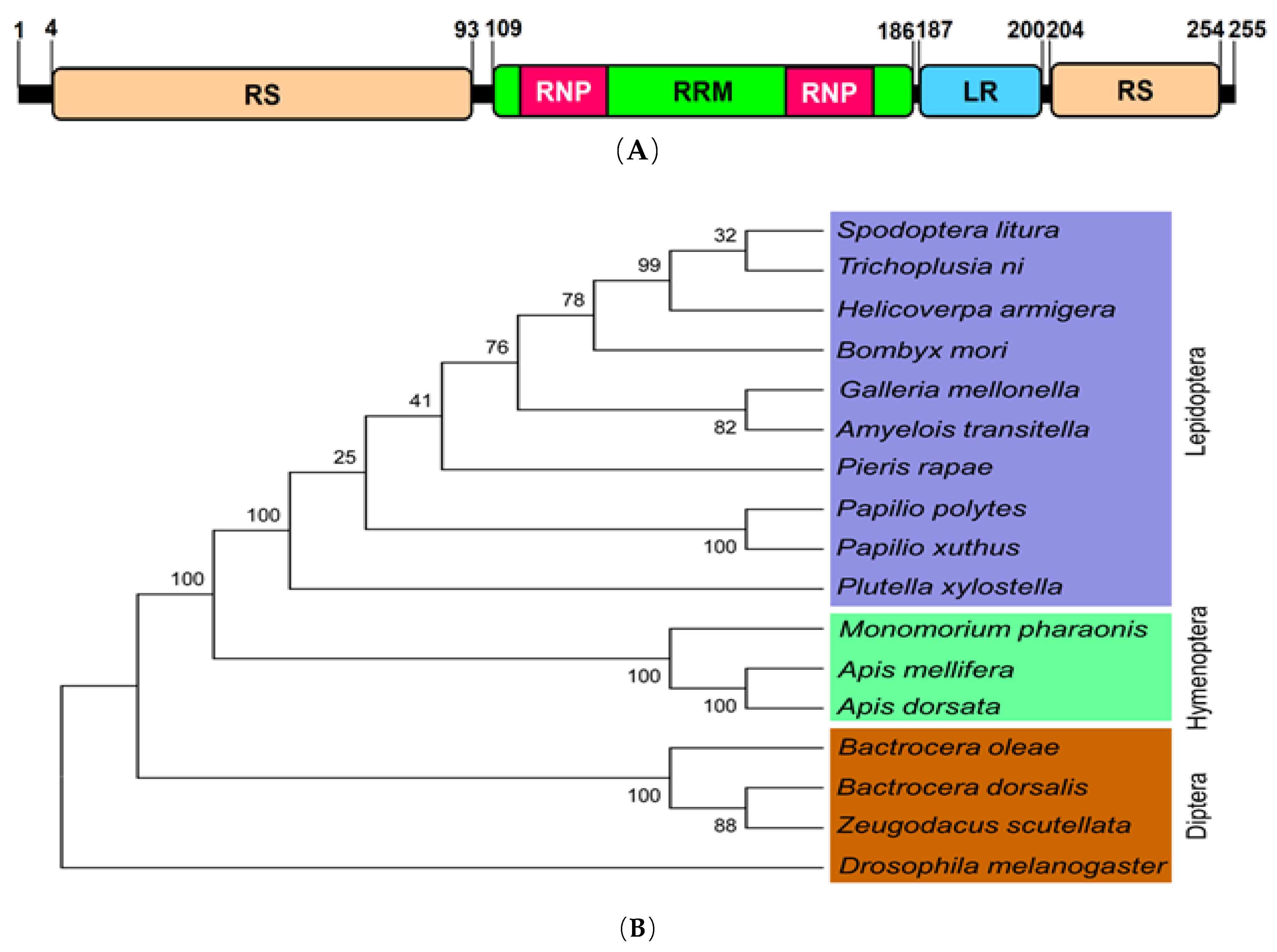
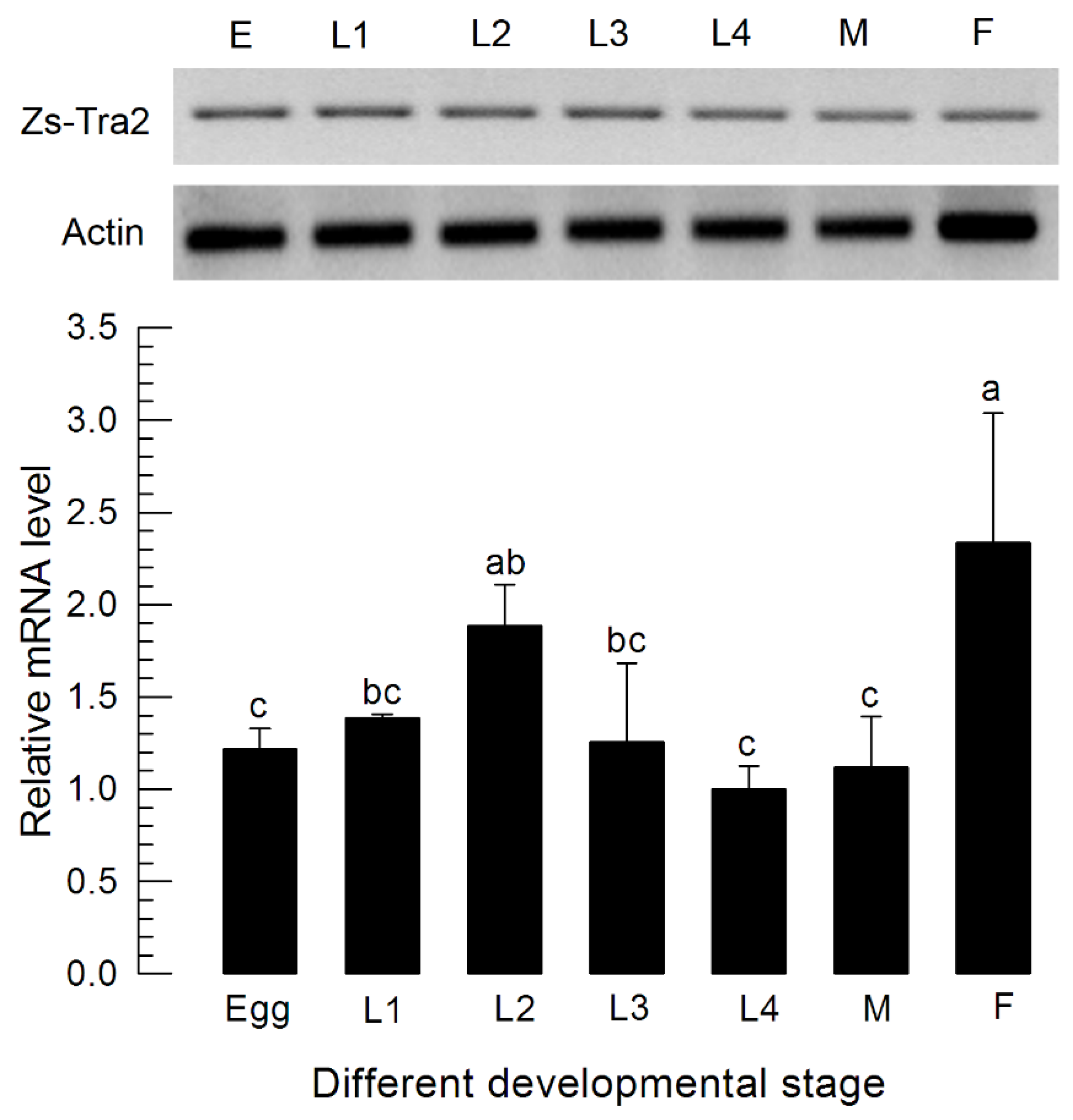
Zs-Tra2 sequence was obtained from GenBank with accession number of MT103663. Zs-Tra2 protein was predicted to possess signature motifs such as arginine/serine-rich domain (RS), ribonucleoprotein region (RNP), RNA recognition motif (RRM), and a linker region (LR) between RRM and RS (Figure 1A). Phylogenetic analysis indicated that Zs-Tra2 was clustered with other dipteran Tra-2 genes, in which it shared the highest homology (88%) with that of B. dorsalis (Figure 1B). Expression patterns of Zs-Tra2 in different developmental stages of Z. scutellata were analyzed (Figure 2). Zs-Tra2 was expressed in all developmental stage from eggs to adults with high expression levels in females. In females, it was expressed in all three body parts, such as head, thorax, and abdomen (data not shown).
3.3. Male-Biased Sex Ratio after RNAi of Zs-Tra2 Expression
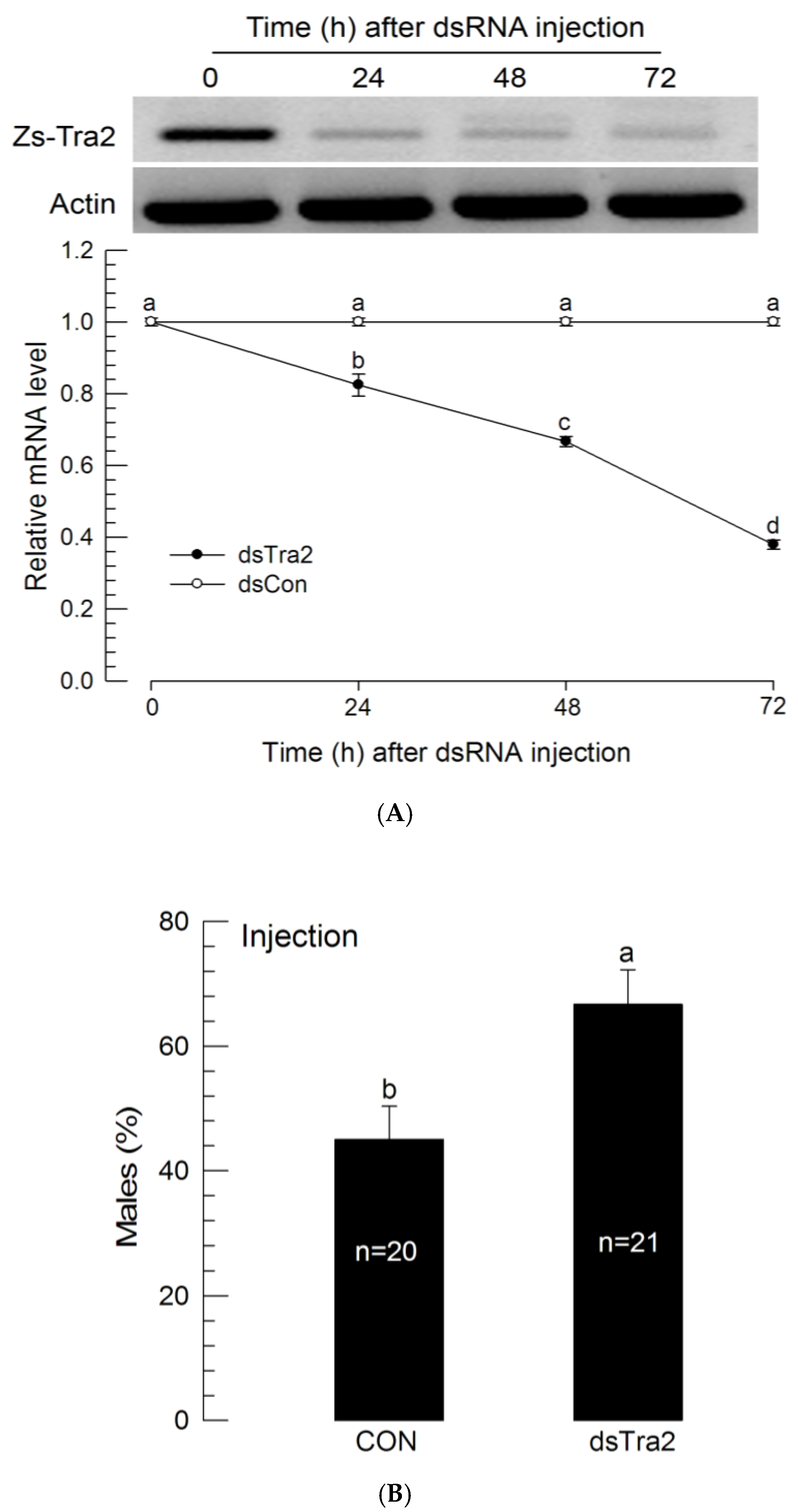
To determine the function of Zs-Tra2 in sex determination, Zs-Tra2 expression was silenced by RNAi (Figure 3). To silence Zs-Tra2 expression, its specific dsRNA was injected to first instar larvae which significantly (p < 0.05) reduced its mRNA level at 24 h after injection. The microinjection gave little injection damage (<1%) on the larval survival. The RNAi effect was maintained for at least 72 h after injection (Figure 3A). Under this condition, RNAi-treated larvae had 63% of males while control larvae had only 41% males (Figure 3B).
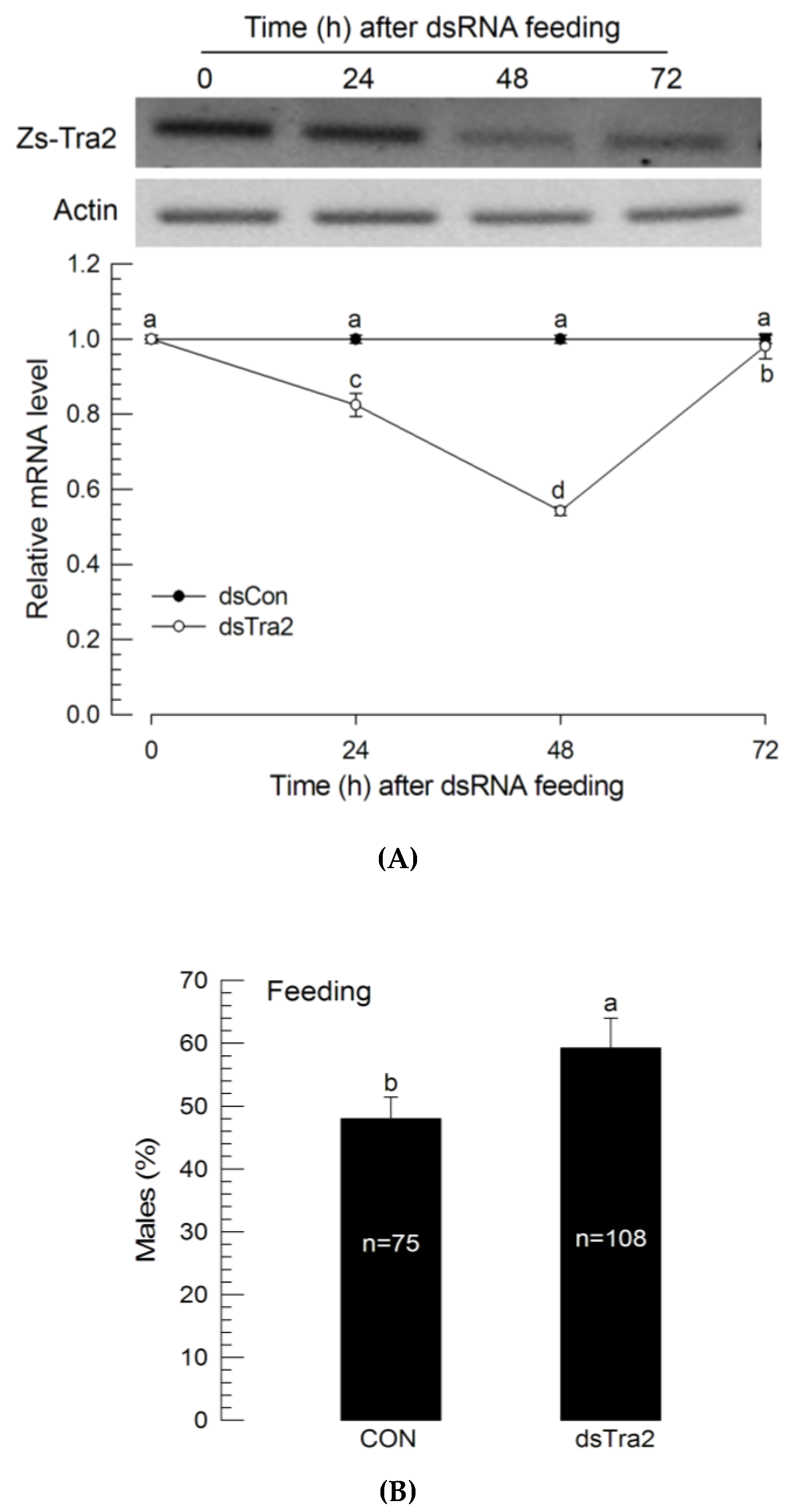
To determine whether RNAi effect could be achieved by oral administration, dsRNA was delivered via feeding after mixing dsRNA with the larval diet (Figure 4A). L1 larvae were treated for 24 h with a diet containing dsRNA. The Zs-Tra2 expression level was significantly (p < 0.05) decreased for two days after the feeding treatment. However, it then increased back to the level of untreated larvae. RNAi feeding also resulted in a male-biased sex ratio (Figure 4B).
3.4. Construction of Recombinant E. coli Expressing dsRNA Specific to Zs-Tra2 and Its Effect on Sex Transformation
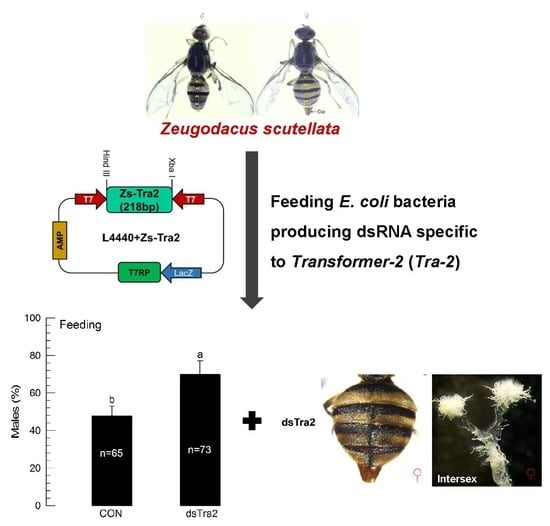
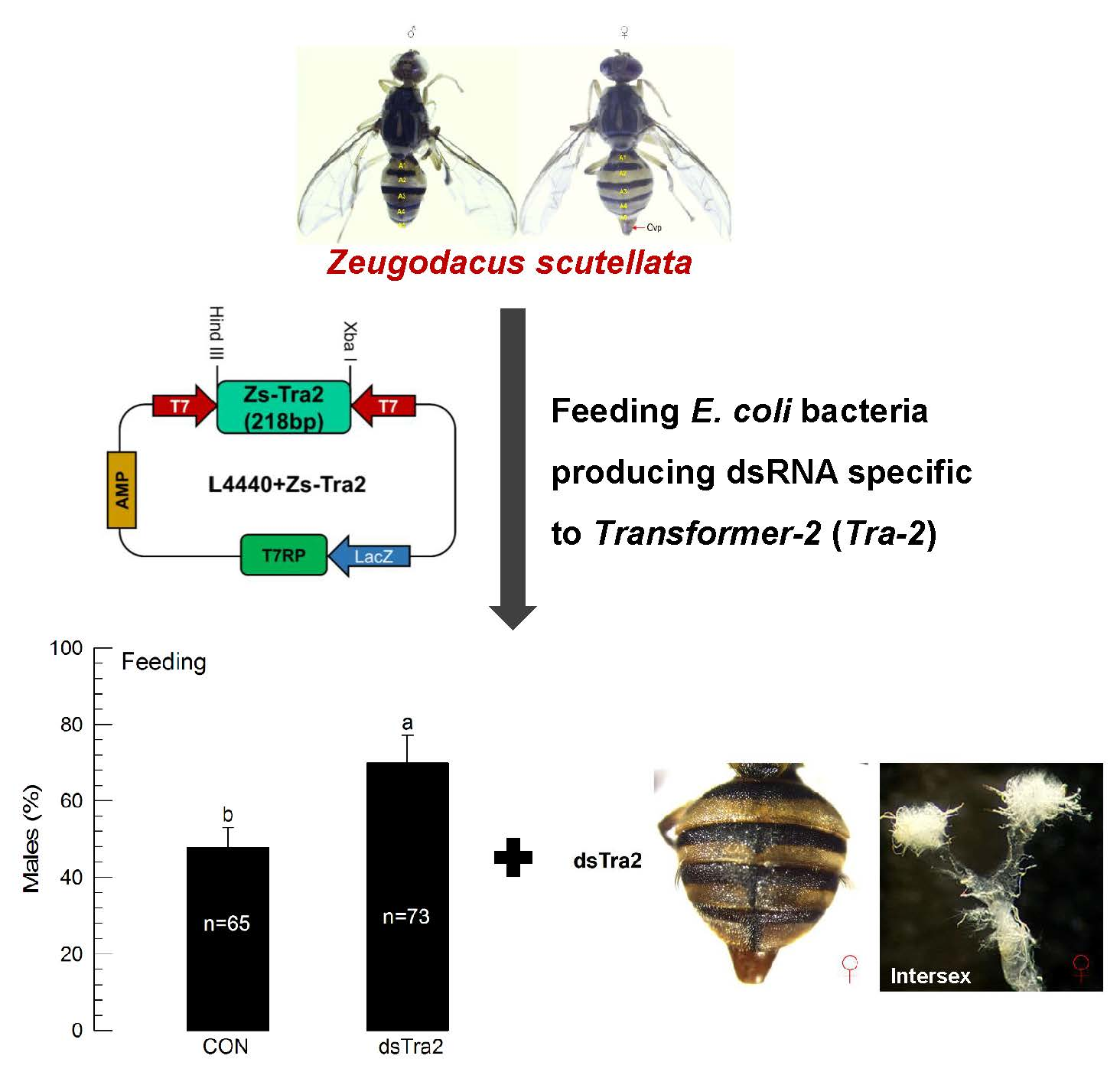
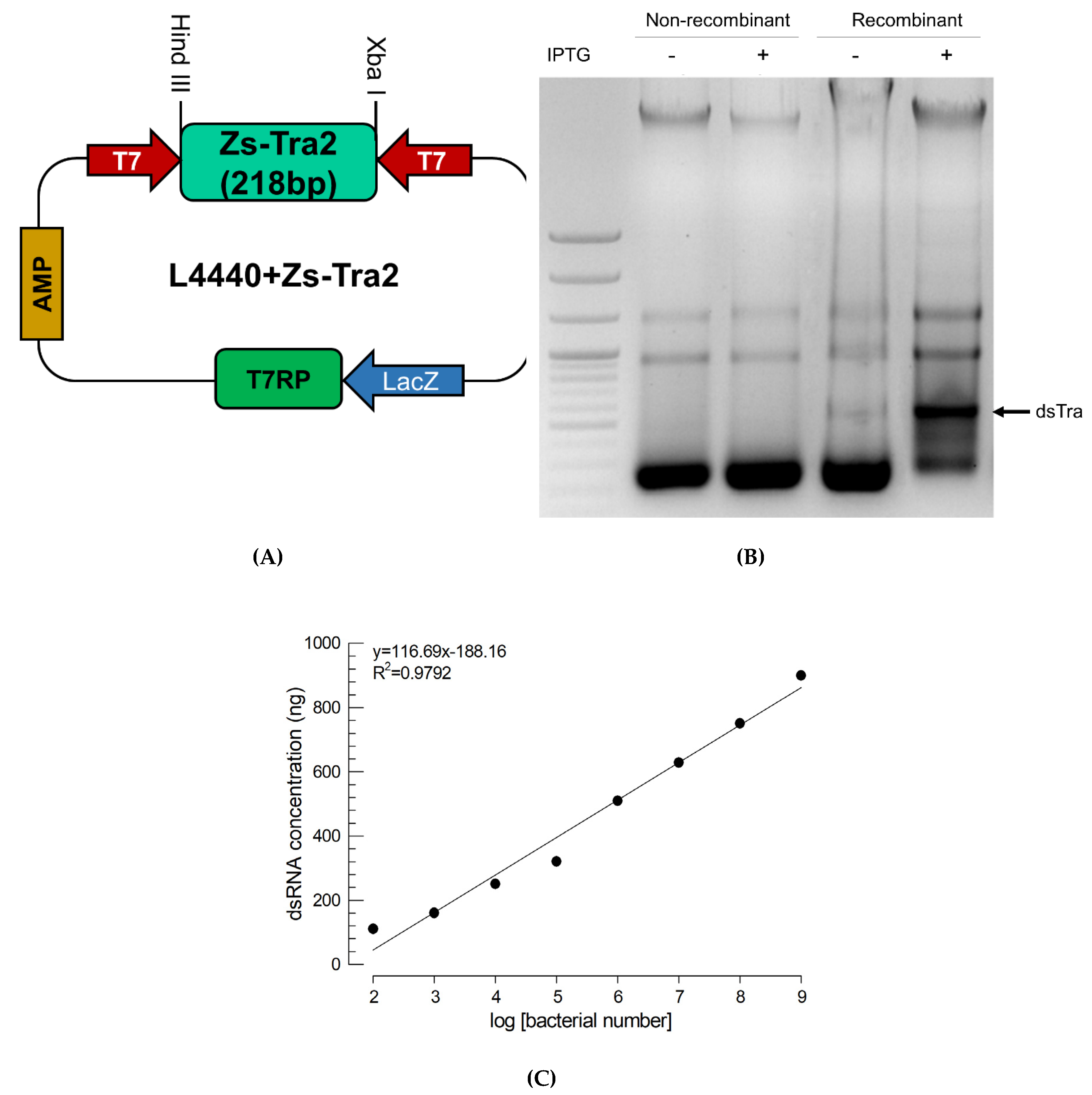
To express dsRNA in E. coli, a partial (218 bp) sequence of Zs-Tra2 was inserted between opposite T7 promoters (Figure 5A). The recombinant L4440 expression vector was then used to transform E. coli and the dsRNA was over-expressed by adding IPTG inducer (Figure 5B). Extracted RNA from recombinant bacteria contained dsRNA specific to Zs-Tra2. The amount of dsRNA produced from such recombinant E. coli was linearly proportional to bacterial density (Figure 5C).
3.5. Effect of Feeding Recombinant E. coli on Sex Ratio of Z. scutellata
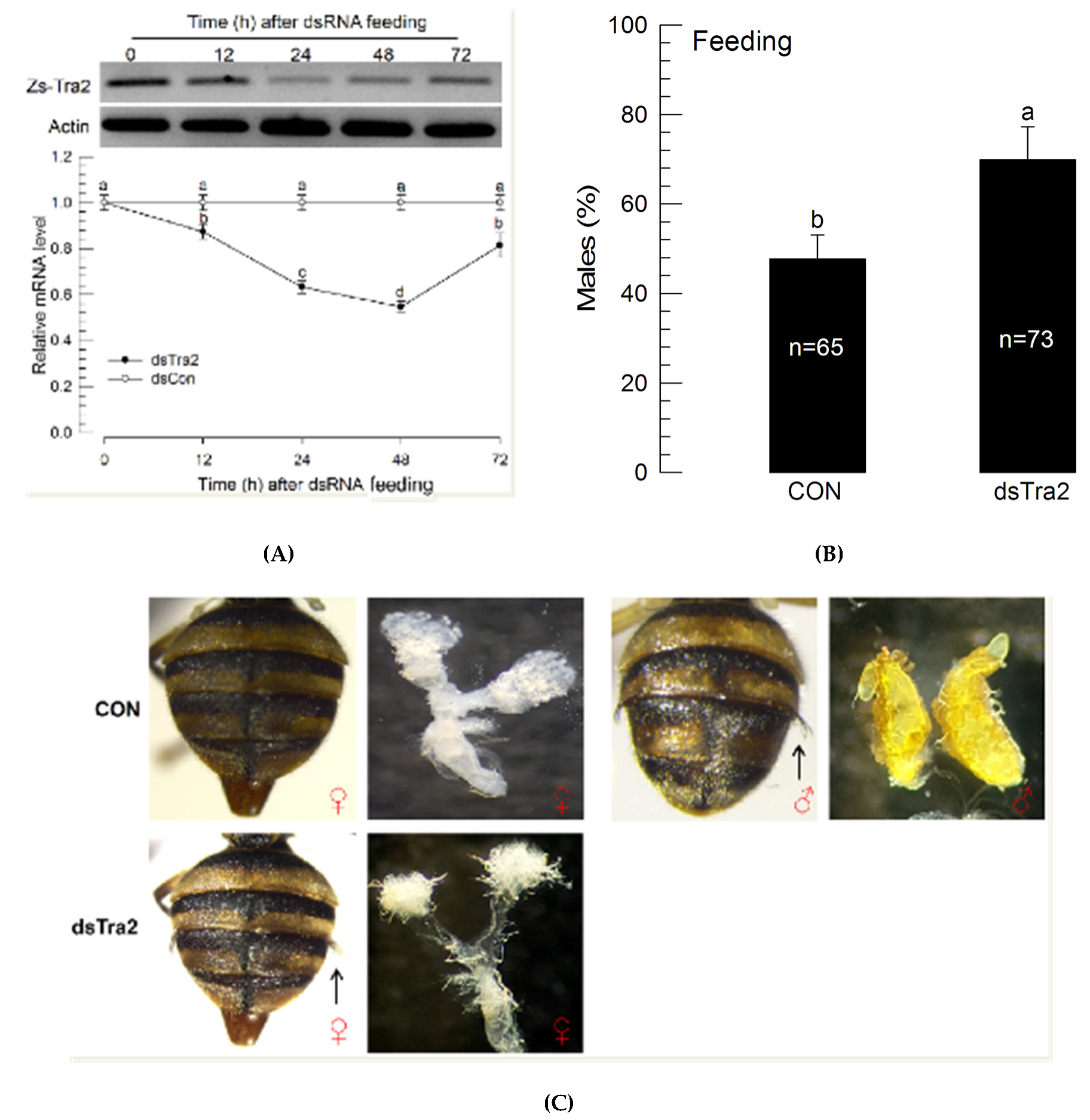
Feeding recombinant bacteria significantly (p < 0.05) reduced mRNA levels of Zs-Tra2 (Figure 6A). This RNAi effect resulted in a male-biased sex ratio in adults (Figure 6B). Compared to the control, RNAi-treated flies had more males by over 30%. Interestingly, some females (containing ovipositor) in the RNAi-treated group developed male-characteristic abdominal setae (Figure 6C). The intersex-like individuals had the ovary.
4. Discussion
This study developed a novel sexing system by feeding dsRNA against Z. scutellata, a serious insect pest of pumpkin crops [8]. Its larvae feed pumpkin flowers within stamen which prevents their exposure to chemical insecticides [27]. Although a mixture treatment of male lure (raspberry ketone or cuelure) with insecticides has been effective for controlling its population [28], SIT has been regarded as one of the most effective control tactics against this insect pest [29]. An obstacle for SIT is the tedious sorting out of females to be removed. To develop an effective sexing system, a comprehensive understanding of sexual differentiation in Z. scutellata offers an opportunity to promote the development of novel sexing strategies and genetic control techniques to fight this pest. In this regard, the altering sex determination system was exploited by RNAi in this study.
High expression levels of Zs-Tra2 were required for female sex differentiation in Z. scutellata. Zs-Tra2 was expressed in all developmental stages. It was also expressed in both male and female adults. However, it was expressed in much higher levels in females than in males. Tra-2 protein and RNA-binding protein 1 are associated with Tra protein to form a splicing regulatory complex that mediates the splicing of its targets, Dsx and Fru mRNAs in Drosophila melanogaster [30]. Tra-2 belongs to the family of RNA binding proteins containing an RNA-recognition motif (‘RRM’) domain which plays crucial role in regulating mRNA splicing of Dsx mRNA into a female type variant [31,32]. Without Tra-2, Dsx mRNA is spliced into a male-specific Dsx transcript variant. In D. melanogaster, two mRNA variants of Tra-2 are expressed in somatic tissues of both sexes. The other two variants are present only in male germline [33]. Somatic Tra-2 products mediate the alternative splicing of Dsx mRNA while two male Tra-2 proteins are functioning in spermatogenesis [34,35,36]. In tephritid fruit flies, mRNA variants of Tra-2 have been reported. For example, three mRNA variants of Tra-2 have been identified in Z. tau, of which at least one is expressed in both sexes and the other is specific to the embryo or germline [21]. Although Zs-Tra2 mRNA variants were not assessed in our current study, its expression profile in males suggested the presence of variants of the Zs-Tra2 transcript.
The expression of RNAi specific to Zs-Tra2 resulted in a male-biased sex ratio in Z. scutellata. A microinjection of dsRNA to L1 larvae was effective in increasing its male production. Feeding dsRNA to growing larvae also increased its male ratio. In various fruit flies other than Drosophila, Tra-2 genes have been identified. They play crucial roles in the regulation of female-specific splicing of Dsx pre-mRNA. These include 12 Anastrepha species [37,38], Bactrocera oleae [39], B. tryoni, B. jarvisi [40], B. dorsalis [20], B. tau [21], and C. capitata [19]. The knocking down of Tra-2 expression in these tephritid fruit flies can interrupt the splicing pattern of Dsx pre-mRNA of females to differentiate females into male phenotypes [41,42]. Compared to these previous studies using a dsRNA injection in an early embryo, our current result of RNAi was obtained by injection or the feeding of dsRNA to growing larvae. Injecting dsRNA into an early embryo is likely to be more effective in producing males than injecting or feeding dsRNA to young larvae. In B. dorsalis, nearly 100% males were obtained with embryonic RNAi against Tra-2 [20]. In Z. tau, dsRNA injection to embryo has produced 72.4% males while the control has produced only 38.6% males [21]. This indicated an increase of 187.6% in male production. In contrast, dsRNA injections into larvae produced only a 153.7% increase in male production compared to control injection. The feeding of dsRNA resulted in much less efficiency (125.0% increase). The lower efficiency of feeding dsRNA might be explained by a lower amount of dsRNA targeting mRNA compared to injecting dsRNA because of the environmental RNAi. In general, the efficiency of RNAi depends on the amount of exogenous dsRNA application [43]. Thus, injecting dsRNA into embryos is more effective than dsRNA treatment against larvae. However, injecting dsRNA to young embryos or larvae is not practical for mass-rearing to perform SIT. Thus, feeding dsRNA to larvae would be ideal to increase male sex ratio.
Recombinant E. coli was constructed to produce dsRNA specific to Zs-Tra2. The amount of dsRNA produced was proportional to the bacterial cell number. Based on the regression equation, the feeding treatment with recombinant bacteria used 12.5 μg of dsRNA in a 250 mg diet. This dose of dsRNA was 2.5 times higher than feeding dsRNA prepared by in vitro transcription. The injection of dsRNA into young larvae used only 15 ng dsRNA per larva. This can be explained by the difficulty in the release of dsRNA from the bacteria in the insect gut. To facilitate dsRNA release from bacteria, this study pre-treated bacteria with heat or ultra-sonication to disrupt the bacterial cell walls and membranes. While there was no significant difference in insecticidal activity between live and heat-killed bacteria, a pretreatment of sonication can significantly enhance the ability to kill Spodoptera exigua insects [25]. The degradation of dsRNA in the gut lumen likely occurs by dsRNase attacks originating from the midgut, as seen in several insects including Bombyx mori [43] and Lygus lineolaris [44] or from salivary secretion during feeding in a sucking insect, Acyrthosiphon pisum [45]. Thus, the actual number of dsRNA molecules released from the transformed E. coli might be remarkably reduced due to the limitation of dsRNA release and the degrading factor in the gut of Z. scutellata. The sex transformation technique by feeding dsRNA was supported by the development of intersex individuals detected in some females. This might be explained by the mosaic expression of male- or female-type Dsx expression by fluctuating Zs-Tra2 mRNA levels among different tissues. These results indicated that the dsRNA feeding technique was effective in suppressing Zs-Tra2 expression to alter sex ratio biased to males. In particular, recombinant bacteria expressing the dsRNA are effective and economical for a mass rearing system to perform SIT. Furthermore, more than 80% of RNAi-generated males could mate with normal females in a study on B. dorsalis [20], suggesting that feeding dsRNA is an ideal approach in a genetic sexing system to achieve male-biased production in SIT.
5. Conclusions
This study aimed to develop a technique for the sex transformation of Z. scutellata using sex determination system. A certain level of Zs-Tra2 expression was required for female sex differentiation. Its expression level was suppressed by RNAi specific to Zs-Tra2 by feeding dsRNA during the larval period. Feeding recombinant E. coli expressing dsRNA was also effective in suppressing Zs-Tra2 expression and resulted in a male-biased sex ratio. This study introduces a novel technique for sex transformation by feeding dsRNA, which would be applicable for the mass rearing of quarantine fruit flies to be eradicated by SIT.
Supplementary Materials
The following is available online at https://www.mdpi.com/2075-4450/11/4/211/s1, Figure S1, Sexual morphs of Z. scutellata adults. Table S1, Primers used in this study for RT-qPCR.
Author Contributions
Conceptualization, Y.K.; Methodology, M.A.A.B. and M.V.; Formal Analysis, M.A.A.B.; Writing-Original Draft Preparation, M.A.A.B.; Writing-Review and Editing, Y.K.; Funding Acquisition, Y.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries (IPET) through Exporting Promotion Technology Development Program funded by the Ministry of Agriculture, Food and Rural Affairs (MAFRA) (316015-04).
Acknowledgments
We appreciate Youngim Song (Andong National University) for supplying materials and purchasing chemicals.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vargas, R.I.; Piñero, J.C.; Leblanc, L. An overview of pest species of Bactrocera fruit flies (Diptera: Tephritidae) and the integration of biopesticides with other biological approaches for their management with a focus on the Pacific region. Insects 2015, 6, 297–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korneyev, V. Phylogenetic Relationships among the Families of the Superfamily Tephritoidea. In Fruit Flies (Tephritidae). Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC: Boca Raton, FL, USA, 1999; pp. 3–22. [Google Scholar]
- Han, H.; Ro, K.-E. Molecular phylogeny of the superfamily Tephritoidea (Insecta: Diptera): New evidence from the mitochondrial 12S, 16S, and COII genes. Mol. Phylogenetics Evol. 2005, 34, 416–430. [Google Scholar] [CrossRef] [PubMed]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International/ACIAR: Boston, MA, USA, 1992. [Google Scholar]
- Schutze, M.K.; Virgilio, M.; Norrbom, A.; Clarke, A.R. Tephritid integrative taxonomy: Where we are now, with a focus on the resolution of three tropical fruit fly species complexes. Annu. Rev. Entomol. 2017, 62, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, D.-S. Integrated pest management against Bactrocera fruit flies. Korean J. Appl. Entomol. 2016, 55, 359–376. [Google Scholar] [CrossRef]
- Han, M.J.; Lee, S.H.; Ahn, S.B.; Choi, J.Y.; Choi, K.M. Distribution, damage and host plants of pumpkin fruit fly, Paradacus depressa (Shiraki). RDA J. Agri. Sci. 1994, 36, 346–350. [Google Scholar]
- Kim, Y.-P.; Jeon, S.-W.; Lee, S.-G.; Kim, K.-H.; Choi, N.-J.; Hwang, C.-Y. Seasonal occurrence and damage of Bactrocera scutellata (Diptera: Tephritidae) in Jeonbuk province. Korean J. Appl. Entomol. 2010, 49, 299–304. [Google Scholar] [CrossRef]
- Shiraki, T. Fruit flies of the Ryukyu islands. Bull. United States Natl. Mus. 1968, 263, 1–104. [Google Scholar] [CrossRef]
- Miyatake, T.; Kuba, H.; Yukawa, J. Seasonal occurrence of Bactrocera scutellata (Diptera: Tephritidae), a cecidophage of stem galls produced by Lasioptera sp. (Diptera: Cecidomyiidae) on wild gourds (Cucurbitaceae). Ann. Entomol. Soc. Am. 2000, 93, 1274–1279. [Google Scholar] [CrossRef]
- Ohno, S.; Haraguchi, D.; Kohama, T. New host and distribution records of the fruit fly, Bactrocera scutellata (Hendel) (Diptera: Tephritidae), in Southwestern Japan, and a case of infestation of the species on cucumber fruits at Okinawa Island. Jpn. J. Entomol. 2006, 9, 7–9. [Google Scholar]
- Enkerlin, W.R. Impact of fruit fly control programs using the sterile insect technique. In Sterile Insect Technique Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Berlin, Germany, 2005; pp. 651–676. [Google Scholar]
- Knipling, E.F. Possibilities of Insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Rendón, P.; McInnis, D.; Lance, D.; Stewart, J. Medfly (Diptera: Tephritidae) genetic sexing: Large-scale field comparison of males-only and bisexual sterile fly releases in Guatemala. J. Econ. Entomol. 2004, 97, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- McInnis, D.; Tam, S.; Lim, R.; Komatsu, J.; Kurashima, R.; Albrecht, C. Development of a pupal color-based genetic sexing strain of the melon fly, Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2004, 97, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Ogaugwu, C.E.; Schetelig, M.F.; Wimmer, E.A. Transgenic sexing system for Ceratitis capitata (Diptera: Tephritidae) based on female-specific embryonic lethality. Insect Biochem. Mol. Boil. 2013, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cline, T.W. The Drosophila sex determination signal: How do flies count to two? Trends Genet. 1993, 9, 385–390. [Google Scholar] [CrossRef]
- Sharma, A.; Heinze, S.D.; Wu, Y.; Kohlbrenner, T.; Morilla, I.; Brunner, C.; Wimmer, E.A.; Van De Zande, L.; Robinson, M.D.; Beukeboom, L.W.; et al. Male sex in houseflies is determined byMdmd, a paralog of the generic splice factor geneCWC22. Science 2017, 356, 642–645. [Google Scholar] [CrossRef]
- Salvemini, M.; Robertson, M.; Aronson, B.; Atkinson, P.; Polito, L.C.; Saccone, G. Ceratitis capitata transformer-2 gene is required to establish and maintain the autoregulation of Cctra, the master gene for female sex determination. Int. J. Dev. Boil. 2009, 53, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Wu, Q.; Li, J.-W.; Zhang, G.; Wan, F.-H. RNAi-mediated knock-down of transformer and transformer 2 to generate male-only progeny in the Oriental fruit fly, Bactrocera dorsalis (Hendel). PLoS ONE 2015, 10, e0128892. [Google Scholar] [CrossRef] [Green Version]
- Thongsaiklaing, T.; Nipitwattanaphon, M.; Ngernsiri, L. The transformer2 gene of the pumpkin fruit fly, Bactrocera tau (Walker), functions in sex determination, male fertility and testis development. Insect Mol. Boil. 2018, 27, 766–779. [Google Scholar] [CrossRef]
- Bustin, S.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Park, B.; Kim, Y. Transient transcription of a putative RNase containing BEN domain encoded in Cotesia plutellae bracovirus induces an immunosuppression of the diamondback moth, Plutella xylostella. J. Invertebr. Pathol. 2010, 105, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Park, Y.; Kim, Y. A transformed bacterium expressing double-stranded RNA specific to integrin β1 enhances Bt toxin efficacy against a polyphagous insect pest, Spodoptera exigua. PLoS ONE 2015, 10, e0132631. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT User’s Guide; SAS Institute: Cary, NC, USA, 1989. [Google Scholar]
- Jeon, S.-W.; Kang, T.-J.; Cho, M.-R.; Kim, K.-H.; Lee, S.G.; Kim, J.S.; Park, H.W. Adult longevity and life table analysis of striped fruit fly, Bactrocera scutellata (Hendel) (Diptera:Tephritidae). Korean J. Appl. Entomol. 2012, 51, 485–488. [Google Scholar] [CrossRef]
- Kim, K.; Kim, M.; Kwon, G.; Kim, Y. Technologies required for development of trap-based MAT control against the striped fruit fly, Bactrocera scutellata. Korean J. Appl. Entomol. 2017, 56, 51–60. [Google Scholar] [CrossRef]
- Kim, Y.; Al Baki, M.A.; Kwon, G. Technique to generate sterile males of striped fruit flies, Zeugodacus scutellata, using electron beam irradiation and their application to genetic control. Korean J. Appl. Entomol. 2020, 59, 1–10. [Google Scholar]
- Amrein, H.; Maniatis, T.; Nöthiger, R. Alternatively spliced transcripts of the sex-determining gene tra-2 of Drosophila encode functional proteins of different size. EMBO J. 1990, 9, 3619–3629. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.S.; Wolfner, M.F. A molecular analysis of doublesex, a bifunctional gene that controls both male and female sexual differentiation in Drosophila melanogaster. Genome Res. 1988, 2, 477–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshijima, K.; Inoue, K.; Higuchi, I.; Sakamoto, H.; Shimura, Y. Control of doublesex alternative splicing by transformer and transformer-2 in Drosophila. Science 1991, 252, 833–836. [Google Scholar] [CrossRef]
- Mattox, W.; Palmer, M.J.; Baker, B.S. Alternative splicing of the sex determination gene transformer-2 is sex-specific in the germ line but not in the soma. Genes Dev. 1990, 4, 789–805. [Google Scholar] [CrossRef] [Green Version]
- Mattox, W.; Baker, B.S. Autoregulation of the splicing of transcripts from the transformer-2 gene of Drosophila. Genes Dev. 1991, 5, 786–796. [Google Scholar] [CrossRef] [Green Version]
- Mattox, W.; McGuffin, M.E.; Baker, B.S. A negative feedback mechanism revealed by functional analysis of the alternative isoforms of the Drosophila splicing regulator transformer-2. Genetics 1996, 143, 303–314. [Google Scholar] [PubMed]
- McGuffin, M.E.; Chandler, D.; Somaiya, D.; Dauwalder, B.; Mattox, W. Autoregulation of transformer-2 alternative splicing is necessary for normal male fertility in Drosophila. Genetics 1998, 149, 1477–1486. [Google Scholar] [PubMed]
- Sarno, F.; Ruiz, M.F.; Eirin-Lopez, J.M.; Perondini, A.; Selivon, D.; Sánchez, L. The gene transformer-2 of Anastrepha fruit flies (Diptera, Tephritidae) and its evolution in insects. BMC Evol. Boil. 2010, 10, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schetelig, M.F.; Milano, A.; Saccone, G.; Handler, A.M. Male only progeny in Anastrepha suspensa by RNAi-induced sex reversion of chromosomal females. Insect Biochem. Mol. Boil. 2012, 42, 51–57. [Google Scholar] [CrossRef]
- Lagos, D.; Koukidou, M.; Savakis, C.; Komitopoulou, K. The transformer gene in Bactrocera oleae: The genetic switch that determines its sex fate. Insect Mol. Boil. 2007, 16, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Morrow, J.L.; Riegler, M.; Frommer, M.; Shearman, D. Expression patterns of sex-determination genes in single male and female embryos of two Bactrocera fruit fly species during early development. Insect Mol. Boil. 2014, 23, 754–767. [Google Scholar] [CrossRef]
- Pane, A.; Salvemini, M.; Delli Bovi, P.; Polito, C.; Saccone, G. Faculty of 1000 evaluation for The transformer gene in Ceratitis capitata provides a genetic basis for selecting and remembering the sexual fate. Development 2002, 129, 3715–3725. [Google Scholar]
- Hediger, M.; Siegenthaler, C.; Moser, M.; Bopp, D. The transformer2 gene in Musca domestica is required for selecting and maintaining the female pathway of development. Dev. Genes Evol. 2005, 215, 165–176. [Google Scholar]
- Liu, J.; Smagghe, G.; Swevers, L. Transcriptional response of BmToll9-1 and RNAi machinery genes to exogenous dsRNA in the midgut of Bombyx mori. J. Insect Physiol. 2013, 59, 646–654. [Google Scholar] [CrossRef]
- Allen, M.; Walker, W.B.; Iii, W.B.W. Saliva of Lygus lineolaris digests double stranded ribonucleic acids. J. Insect Physiol. 2012, 58, 391–396. [Google Scholar] [CrossRef]
- Christiaens, O.; Swevers, L.; Smagghe, G. DsRNA degradation in the pea aphid (Acyrthosiphon pisum) associated with lack of response in RNAi feeding and injection assay. Peptides 2014, 53, 307–314. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecular characters of Transformer-2 gene (Zs-Tra2) of Zeugodacus scutellata. (A) Functional domains and motifs were predicted with Pfam (https://www.pfam.xfam.org/): Arginine/serine rich domain (‘RS’), RNA recognition motif (‘RRM’), ribonucleoprotein region (‘RNP’), and linker region (LR). (B) Phylogenetic analysis of Zs-Tra2 with other insect orthologs. Amino acid sequences of Transformer-2 were retrieved from GenBank with accession numbers of XP_012553366.1 for Bombyx mori, NP_001252514.1 for Apis mellifera, XP_021199210.1 for Helicoverpa armigera, XP_022838015.1 for Spodoptera litura, XP_006623037.1 for Apis dorsata, XP_026762058.1 for Galleria mellonella, XP_012539181.1 for Monomorium pharaonis, CAD67988.1 for Bactrocera oleae, ALJ94044.1 for Bactrocera dorsalis, XP_013197999.1 for Amyelois transitella, XP_013145601.1 for Papilio polytes, XP_013170155.1 for Papilio xuthus, XP_011560377.1 for Plutella xylostella, XP_022120857.1 for Pieris rapae, XP_026724961.1 for Trichoplusia ni, and AAF58232.2 for Drosophila melanogaster. Amino acid sequences were aligned with MEGA6 program. Bootstrapping values were obtained with 1000 repetitions to support branch and clustering.
Figure 1.
Molecular characters of Transformer-2 gene (Zs-Tra2) of Zeugodacus scutellata. (A) Functional domains and motifs were predicted with Pfam (https://www.pfam.xfam.org/): Arginine/serine rich domain (‘RS’), RNA recognition motif (‘RRM’), ribonucleoprotein region (‘RNP’), and linker region (LR). (B) Phylogenetic analysis of Zs-Tra2 with other insect orthologs. Amino acid sequences of Transformer-2 were retrieved from GenBank with accession numbers of XP_012553366.1 for Bombyx mori, NP_001252514.1 for Apis mellifera, XP_021199210.1 for Helicoverpa armigera, XP_022838015.1 for Spodoptera litura, XP_006623037.1 for Apis dorsata, XP_026762058.1 for Galleria mellonella, XP_012539181.1 for Monomorium pharaonis, CAD67988.1 for Bactrocera oleae, ALJ94044.1 for Bactrocera dorsalis, XP_013197999.1 for Amyelois transitella, XP_013145601.1 for Papilio polytes, XP_013170155.1 for Papilio xuthus, XP_011560377.1 for Plutella xylostella, XP_022120857.1 for Pieris rapae, XP_026724961.1 for Trichoplusia ni, and AAF58232.2 for Drosophila melanogaster. Amino acid sequences were aligned with MEGA6 program. Bootstrapping values were obtained with 1000 repetitions to support branch and clustering.

Figure 2.
Expression profile of Zs-Tra2 in different developmental stages including different larval instars (‘L1–L3’), male (‘M’) adults, and female (‘F’) adults. Actin gene was used as a reference gene to normalize target gene expression level in RT-qPCR. All treatments were independently replicated three times. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test).
Figure 2.
Expression profile of Zs-Tra2 in different developmental stages including different larval instars (‘L1–L3’), male (‘M’) adults, and female (‘F’) adults. Actin gene was used as a reference gene to normalize target gene expression level in RT-qPCR. All treatments were independently replicated three times. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test).

Figure 3.
Influence of dsRNA (‘dsTra2’) specific to Zs-Tra2 on male-biased adult production in Z. scutellata. RNAi was performed by injecting 0.03 μL (500 ng/μL) of dsRNA to newly molted L1 larvae (<12 h) by microinjection. For dsRNA control (‘dsCon’), dsRNA specific to CpBV-ORF302 was prepared with the method of Park and Kim (2010). (A) RNAi efficiency was assessed by RT-qPCR. Actin was used as a reference gene of RT-qPCR to normalize target gene expression level. Each treatment was replicated with independently prepared samples. (B) Outcome was assessed in ratio of male adults. Number indicate the number of assessed adults. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test).
Figure 3.
Influence of dsRNA (‘dsTra2’) specific to Zs-Tra2 on male-biased adult production in Z. scutellata. RNAi was performed by injecting 0.03 μL (500 ng/μL) of dsRNA to newly molted L1 larvae (<12 h) by microinjection. For dsRNA control (‘dsCon’), dsRNA specific to CpBV-ORF302 was prepared with the method of Park and Kim (2010). (A) RNAi efficiency was assessed by RT-qPCR. Actin was used as a reference gene of RT-qPCR to normalize target gene expression level. Each treatment was replicated with independently prepared samples. (B) Outcome was assessed in ratio of male adults. Number indicate the number of assessed adults. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test).

Figure 4.
Influence of dsRNA (‘dsTra2’) specific to Zs-Tra2 on male-biased adult production in Z. scutellata. RNAi was performed by feeding dsRNA (10 μg) mixed with diet (500 mg) to newly molted L1 larvae (<12 h) for 48 h. For dsRNA control (‘dsCon’), dsRNA specific to CpBV-ORF302 was prepared by the method of Park and Kim [24]. (A) RNAi efficiency assessed by RT-qPCR. Actin gene expression was used as a reference gene of RT-qPCR to normalize target gene expression level. Each treatment was replicated with independently prepared samples. (B) Outcome was assessed in ratio of male adults. Number indicates the number of assessed adults. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test).
Figure 4.
Influence of dsRNA (‘dsTra2’) specific to Zs-Tra2 on male-biased adult production in Z. scutellata. RNAi was performed by feeding dsRNA (10 μg) mixed with diet (500 mg) to newly molted L1 larvae (<12 h) for 48 h. For dsRNA control (‘dsCon’), dsRNA specific to CpBV-ORF302 was prepared by the method of Park and Kim [24]. (A) RNAi efficiency assessed by RT-qPCR. Actin gene expression was used as a reference gene of RT-qPCR to normalize target gene expression level. Each treatment was replicated with independently prepared samples. (B) Outcome was assessed in ratio of male adults. Number indicates the number of assessed adults. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test).

Figure 5.
Recombinant E. coli expressing dsRNA (dsTra2) specific to Zs-Tra2. (A) Cloning of a fragment of Zs-Tra2 into an expression vector of L4440 using Hind III and Xba I multiple cloning sites. After selecting transformed cells using ampicillin (‘AMP’) medium, T7 RNA polymerase (‘T7 RP’) was overexpressed by IPTG inducer on lactose promoter (‘LacZ’). Partial Zs-Tra2 RNA is transcribed by two opposite T7 promoters (‘T7’). (B) Confirmation of dsTra2 production specifically in recombinant E. coli under IPTG induction. (C) Regression analysis of dsTra2 amounts from bacterial cell numbers. The extracted dsRNA was quantified by band intensity based on a standard curve using known amounts of dsRNA prepared by in vitro construction method. Each point represents an average obtained from three replications.
Figure 5.
Recombinant E. coli expressing dsRNA (dsTra2) specific to Zs-Tra2. (A) Cloning of a fragment of Zs-Tra2 into an expression vector of L4440 using Hind III and Xba I multiple cloning sites. After selecting transformed cells using ampicillin (‘AMP’) medium, T7 RNA polymerase (‘T7 RP’) was overexpressed by IPTG inducer on lactose promoter (‘LacZ’). Partial Zs-Tra2 RNA is transcribed by two opposite T7 promoters (‘T7’). (B) Confirmation of dsTra2 production specifically in recombinant E. coli under IPTG induction. (C) Regression analysis of dsTra2 amounts from bacterial cell numbers. The extracted dsRNA was quantified by band intensity based on a standard curve using known amounts of dsRNA prepared by in vitro construction method. Each point represents an average obtained from three replications.

Figure 6.
Influence of recombinant E. coli expressing dsRNA (‘dsTra2’) specific to Zs-Tra2 on male-biased adult production in Z. scutellata. RNAi was performed for newly molted L1 larvae (<12 h) by feeding diet (500 mg) containing recombinant E. coli, producing 25 μg of dsTra2. Control (‘dsCON’) used non-recombinant bacteria. (A) RNAi efficiency assessed by RT-qPCR. Actin was used as a reference gene of RT-qPCR to normalize target gene expression level. Each treatment was replicated with independently prepared samples. (B) Outcome assessed in ratio of male adults. Around 300~400 newly hatched L1 larvae were used for both control and treatment. Number indicates the number of assessed adults. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test). (C) Phenotypic analysis in abdominal structures including ovipositor for female character and abdominal pleural setae (see arrows) for male character.
Figure 6.
Influence of recombinant E. coli expressing dsRNA (‘dsTra2’) specific to Zs-Tra2 on male-biased adult production in Z. scutellata. RNAi was performed for newly molted L1 larvae (<12 h) by feeding diet (500 mg) containing recombinant E. coli, producing 25 μg of dsTra2. Control (‘dsCON’) used non-recombinant bacteria. (A) RNAi efficiency assessed by RT-qPCR. Actin was used as a reference gene of RT-qPCR to normalize target gene expression level. Each treatment was replicated with independently prepared samples. (B) Outcome assessed in ratio of male adults. Around 300~400 newly hatched L1 larvae were used for both control and treatment. Number indicates the number of assessed adults. Different letters above standard deviation bars indicate significant difference among means at Type I error = 0.05 (LSD test). (C) Phenotypic analysis in abdominal structures including ovipositor for female character and abdominal pleural setae (see arrows) for male character.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Al Baki, M.A.; Vatanparast, M.; Kim, Y. Male-biased Adult Production of the Striped Fruit Fly, Zeugodacus scutellata, by Feeding dsRNA Specific to Transformer-2. Insects 2020, 11, 211. https://doi.org/10.3390/insects11040211
AMA Style
Al Baki MA, Vatanparast M, Kim Y. Male-biased Adult Production of the Striped Fruit Fly, Zeugodacus scutellata, by Feeding dsRNA Specific to Transformer-2. Insects. 2020; 11(4):211. https://doi.org/10.3390/insects11040211
Chicago/Turabian StyleAl Baki, Md. Abdullah, Mohammad Vatanparast, and Yonggyun Kim. 2020. "Male-biased Adult Production of the Striped Fruit Fly, Zeugodacus scutellata, by Feeding dsRNA Specific to Transformer-2" Insects 11, no. 4: 211. https://doi.org/10.3390/insects11040211
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.