Isolation and Identification of a Strain of Aspergillus Tubingensis With Deoxynivalenol Biotransformation Capability

Abstract
:1. Introduction
2. Results and Discussion
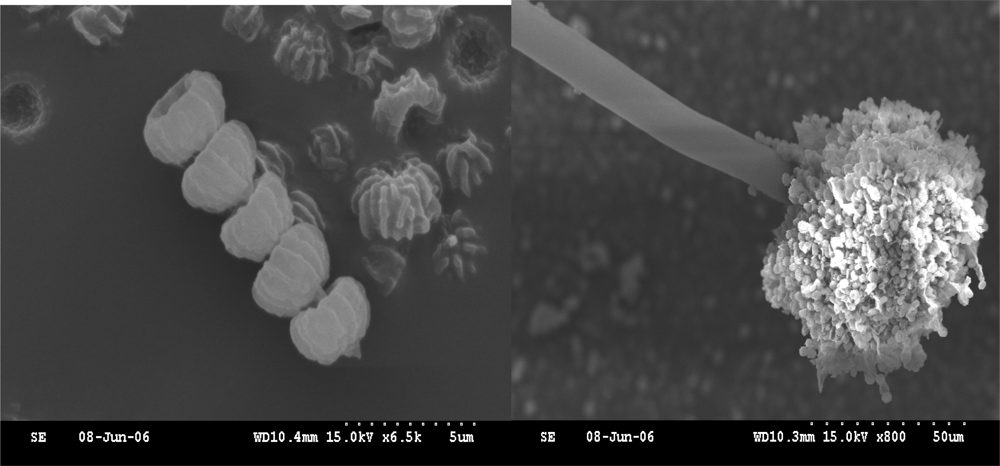
2.1. Isolation of NJA-1
2.2. Identification of NJA-1
2.3. The effect of transformation
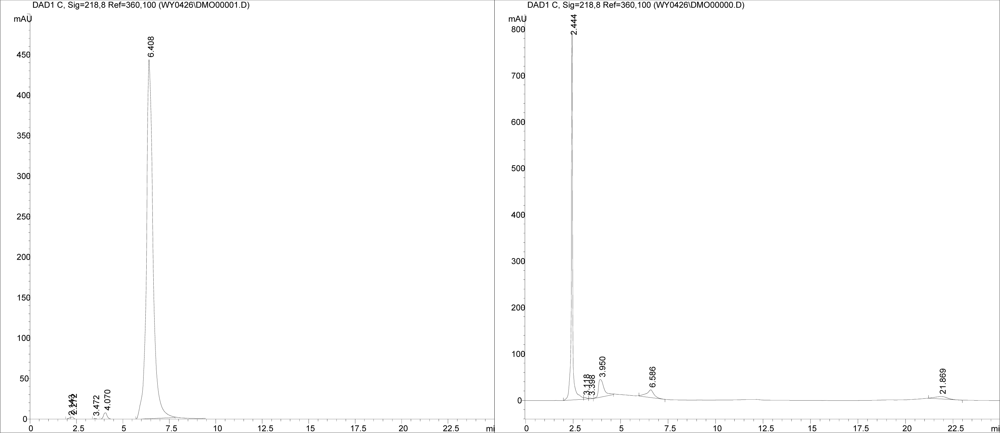
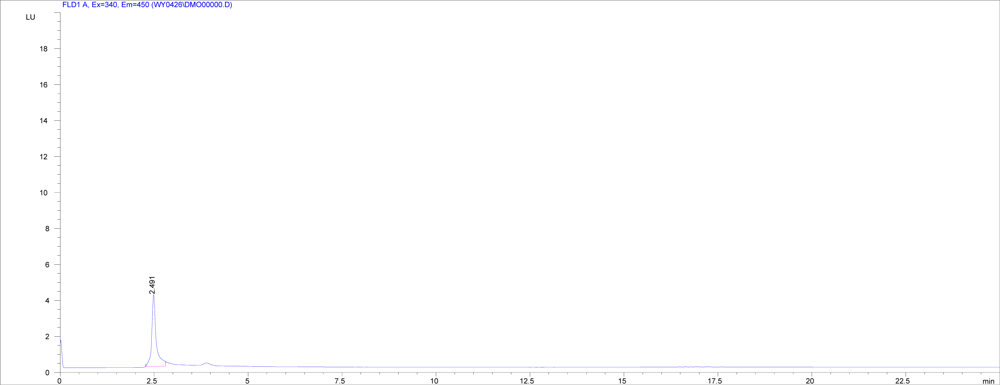
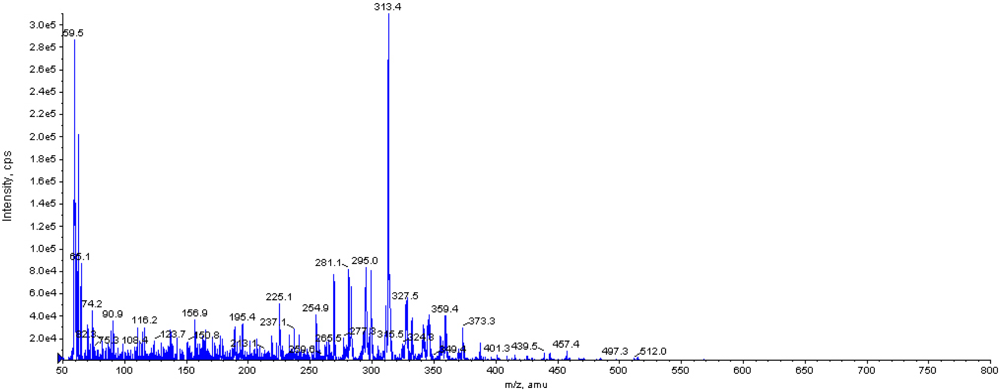
2.4. Screening for DON biotransformation product by NJA-1
3. Experimental Section
3.1. Sampling
3.2. Culture media
3.3. Purification and isolation
3.4. Identification of NJA-1
3.4.1. The observation of conventional morphology and scanning electron microscope
3.4.2. Identification of NJA-1 with PCR
3.5 The effect of DON transformation
3.6 Sscreening for DON biotransformation
4. Conclusions
Acknowledgments
References
- Pieters, MN; Freijer, J; Baars, BJ; Fiolet, DC; van, KJ; Slob, W. Risk assessment of deoxynivalenol in food: concentration limits, exposure and effects. Adv. Exp. Med. Biol 2002, 504, 235–248. [Google Scholar]
- Trucksess, MW; Thomas, F; Young, K; Stack, ME; Fulgueras, WJ; Page, SW. Survey of deoxynivalenol in U.S. 1993 wheat and barley crops by enzyme-linked immunosorbent assay. J. AOAC Int 1995, 78, 631–636. [Google Scholar]
- Tanaka, T; Hasegawa, A; Matsuki, Y; Ueno, Y. A survey of the occurrence of nivalenol, deoxynivalenol and zearalenone in food stuffs and health foods in Japan. Food Addit. Contam 1985, 2, 259–265. [Google Scholar]
- Tanaka, T; Yamamoto, S; Hasegawa, A; Aoki, N; Besling, JR; Sugiura, Y; Ueno, Y. A survey of the natural occurrence of Fusarium mycotoxins, deoxynivalenol, nivalenol and zearalenone, in cereals harvested in the Netherlands. Mycopathologia 1990, 110, 19–22. [Google Scholar]
- Papadopoulou-Bouraoui, A; Vrabcheva, T; Valzacchi, S; Stroka, J; Anklam, E. Screening survey of deoxynivalenol in beer from the European market by an enzyme-linked immunosorbent assay. Food Addit. Contam 2004, 21, 607–617. [Google Scholar]
- Pestka, JJ; Smolinski, AT. Deoxynivalenol: toxicology and potential effects on humans. J. Toxicol. Environ. Health B Crit. Rev 2005, 8, 39–69. [Google Scholar]
- Overnes, G; Matre, T; Sivertsen, T; Larsen, HJ; Langseth, W; Reitan, LJ; Jansen, JH. Effects of diets with graded levels of naturally deoxynivalenol-contaminated oats on immune response in growing pigs. Zentralbl. Veterinarmed. A 1997, 44, 539–550. [Google Scholar]
- Rotter, BA; Prelusky, DB; Pestka, JJ. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar]
- Drochner, W; Schollenberger, M; Piepho, HP; Gotz, S; Lauber, U; Tafaj, M; Klobasa, F; Weiler, U; Claus, R; Steffl, M. Serum IgA-promoting effects induced by feed loads containing isolated deoxynivalenol (DON) in growing piglets. J. Toxicol. Environ. Health A 2004, 67, 1051–1067. [Google Scholar]
- Johnson, PJ; Casteel, SW; Messer, NT. Effect of feeding deoxynivalenol (vomitoxin)-contaminated barley to horses. J. Vet. Diagn. Invest 1997, 9, 219–221. [Google Scholar]
- Prelusky, DB. The effect of low-level deoxynivalenol on neurotransmitter levels measured in pig cerebral spinal fluid. J. Environ. Sci. Health B 1993, 28, 731–761. [Google Scholar]
- Pieters, MN; Bakker, M; Slob, W. Reduced intake of deoxynivalenol in The Netherlands: A risk assessment update. Toxicol. Lett 2004, 153, 145–153. [Google Scholar]
- Bretz, M; Beyer, M; Cramer, B; Knecht, A; Humpf, HU. Thermal degradation of the Fusarium mycotoxin deoxynivalenol. J. Agric. Food Chem 2006, 54, 6445–6451. [Google Scholar]
- Lauren, DR; Smith, WA. Stability of the fusarium mycotoxins nivalenol, deoxynivalenol and zearalenone in ground maize under typical cooking environments. Food Addit. Contam 2001, 18, 1011–1016. [Google Scholar]
- O'Neill, K; Damoglou, AP; Patterson, MF. The stability of deoxynivalenol and 3-acetyl deoxynivalenol to gamma irradiation. Food Addit. Contam 1993, 10, 209–215. [Google Scholar]
- Yumbe-Guevara, BE; Imoto, T; Yoshizawa, T. Effects of heating procedures on deoxynivalenol, nivalenol and zearalenone levels in naturally contaminated barley and wheat. Food Addit. Contam 2003, 20, 1132–1140. [Google Scholar]
- Widestrand, J; Pettersson, H. Effect of time, temperature and solvent on the stability of T-2 toxin, HT-2 toxin, deoxynivalenol and nivalenol calibrants. Food Addit. Contam 2001, 18, 987–992. [Google Scholar]
- Doll, S; Danicke, S; Valenta, H; Flachowsky, G. In vitro studies on the evaluation of mycotoxin detoxifying agents for their efficacy on deoxynivalenol and zearalenone. Arch. Anim. Nutr 2004, 58, 311–324. [Google Scholar]
- Niderkorn, V; Morgavi, DP; Pujos, E; Tissandier, A; Boudra, H. Screening of fermentative bacteria for their ability to bind and biotransform deoxynivalenol, zearalenone and fumonisins in an in vitro simulated corn silage model. Food Addit. Contam 2007, 24, 406–415. [Google Scholar]
- Poppenberger, B; Berthiller, F; Lucyshyn, D; Sieberer, T; Schuhmacher, R; Krska, R; Kuchler, K; Glossl, J; Luschnig, C; Adam, G. Detoxification of the Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem 2003, 278, 47905–47914. [Google Scholar]
- Abolmaali, S; Mitterbauer, R; Spadiut, O; Peruci, M; Weindorfer, H; Lucyshyn, D; Ellersdorfer, G; Lemmens, M; Moll, WD; Adam, G. Engineered bakers yeast as a sensitive bioassay indicator organism for the trichothecene toxin deoxynivalenol. J. Microbiol. Method 2008, 72, 306–312. [Google Scholar]
- Avantaggiato, G; Havenaar, R; Visconti, A. Evaluation of the intestinal absorption of deoxynivalenol and nivalenol by an in vitro gastrointestinal model, and the binding efficacy of activated carbon and other adsorbent materials. Food Chem. Toxicol 2004, 42, 817–824. [Google Scholar]
- Fuchs, E; Binder, EM; Heidler, D; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam 2002, 19, 379–386. [Google Scholar]
- Shima, J; Takase, S; Takahashi, Y; Iwai, Y; Fujimoto, H; Yamazaki, M; Ochi, K. Novel detoxification of the trichothecene mycotoxin deoxynivalenol by a soil bacterium isolated by enrichment culture. Appl. Environ. Microbiol 1997, 63, 3825–3830. [Google Scholar]
- Volkl, A; Vogler, B; Schollenberger, M; Karlovsky, P. Microbial detoxification of mycotoxin deoxynivalenol. J. Basic Microbiol 2004, 44, 147–156. [Google Scholar]
- Fawole, MO; Oso, BA. Laboratory Manual in Microbiology; Spectrum Books Ltd: Ibadan, Nigeria, 1988; p. 14. [Google Scholar]
- Stirling, D. DNA extraction from fungi, yeast, and bacteria. Method. Mol. Biol 2003, 226, 53–54. [Google Scholar]
- Sheir-Neiss, G; Lai, MH; Morris, NR. Identification of a gene for beta-tubulin in Aspergillus nidulans. Cell 1978, 15, 639–647. [Google Scholar]
- Chang, HL; DeVries, JW; Larson, PA; Patel, HH. Rapid determination of deoxynivalenol (vomitoxin) by liquid chromatography using modified Romer column cleanup. J. Assoc. Anal. Chem 1984, 67, 52–54. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of sample | Original concentration (mg/L) | Concentration after 7d cultivation. (mg/L) | Concentration after 14d cultivation. (mg/L) | Transformation efficiency at 14d (%) |
|---|---|---|---|---|
| 1 | 40 | 20.32±0.26 | 3.28±0.02 | 91.8%* |
| 2 | 2 | 1.2±0.06 | 0.139±0.02 | 93.1%* |
| 3 | 1 | 0.56±0.16 | 0.035±0.05 | 96.5%* |
| 4 | 0.4 | 0.18±0.056 | 0.015±0.07 | 96.2%* |
| negative control | 0 | 0 | 0 | 0 |
| positive control | 0.5 | 0.498±0.003 | 0.497±0.01 | 0.6% |
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
He, C.; Fan, Y.; Liu, G.; Zhang, H. Isolation and Identification of a Strain of Aspergillus Tubingensis With Deoxynivalenol Biotransformation Capability. Int. J. Mol. Sci. 2008, 9, 2366-2375. https://doi.org/10.3390/ijms9122366
He C, Fan Y, Liu G, Zhang H. Isolation and Identification of a Strain of Aspergillus Tubingensis With Deoxynivalenol Biotransformation Capability. International Journal of Molecular Sciences. 2008; 9(12):2366-2375. https://doi.org/10.3390/ijms9122366
Chicago/Turabian StyleHe, Chenghua, Yanhong Fan, Guofang Liu, and Haibin Zhang. 2008. "Isolation and Identification of a Strain of Aspergillus Tubingensis With Deoxynivalenol Biotransformation Capability" International Journal of Molecular Sciences 9, no. 12: 2366-2375. https://doi.org/10.3390/ijms9122366