
Genome-Wide Identification of the SPP/SPPL Gene Family and BnaSPPL4 Regulating Male Fertility in Rapeseed (Brassica napus L.)

Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of SPP/SPPL Family Members
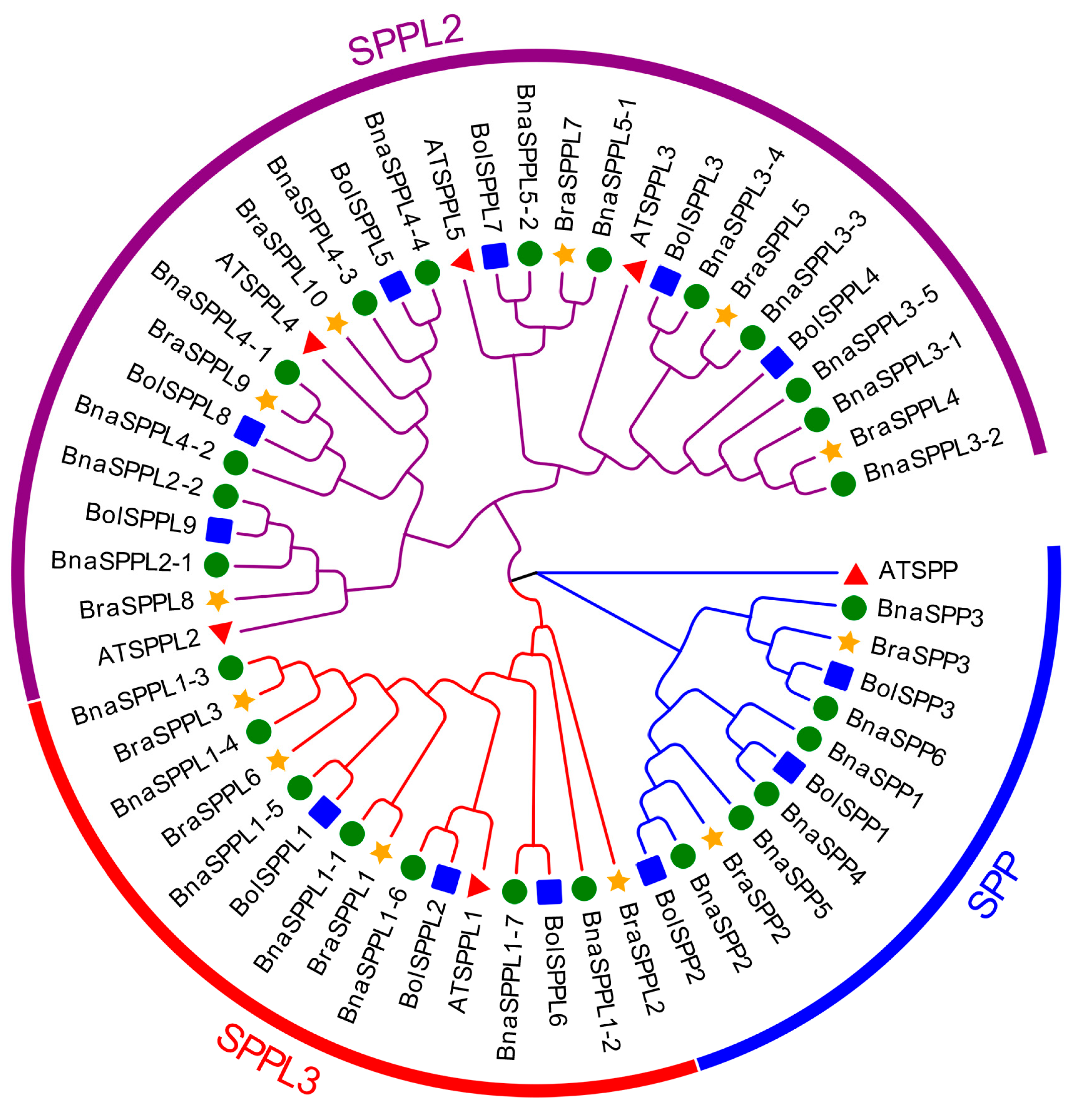
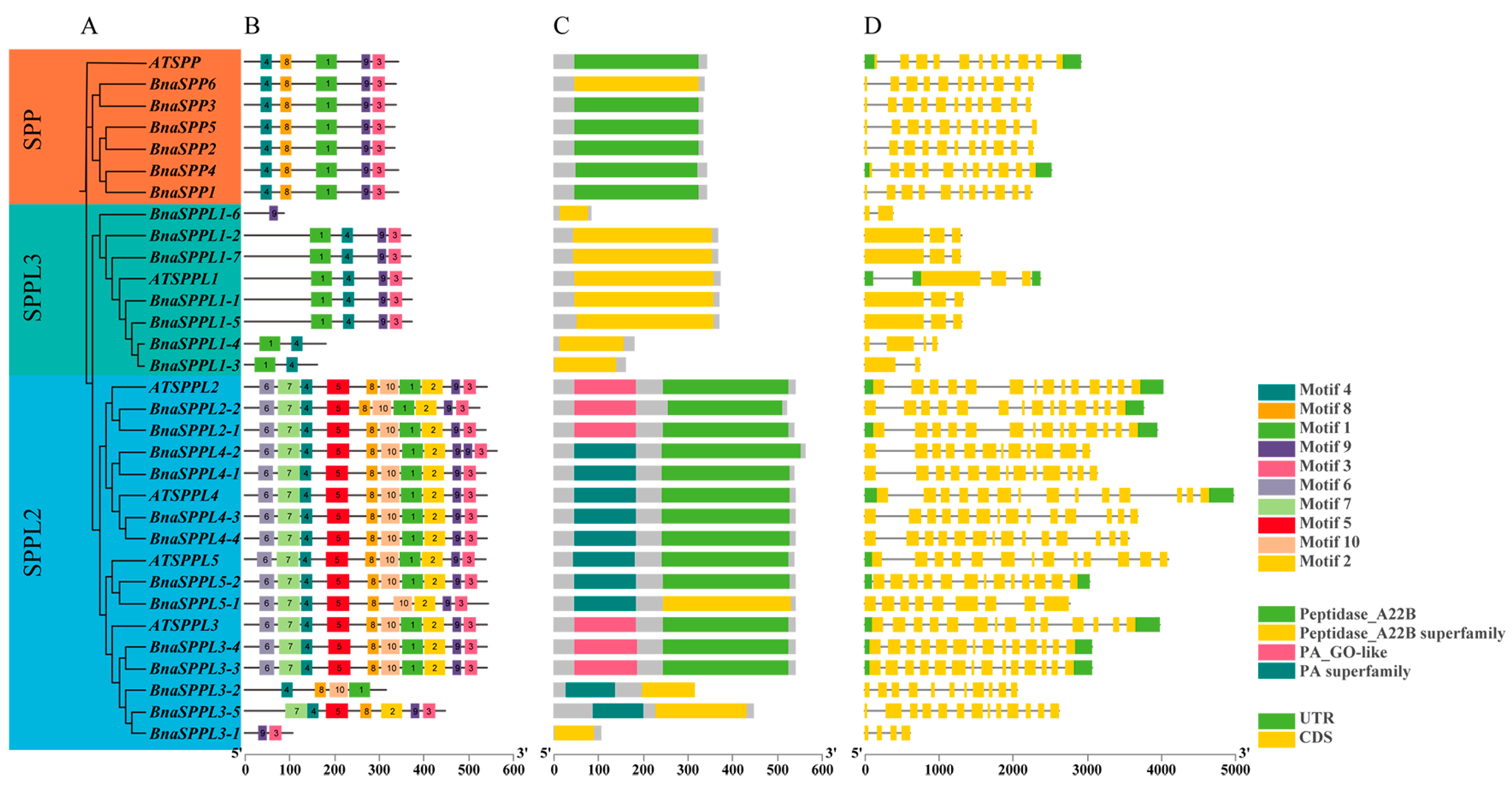
2.2. Phylogenetic and Gene Structure Characterization of BnaSPP/SPPLs
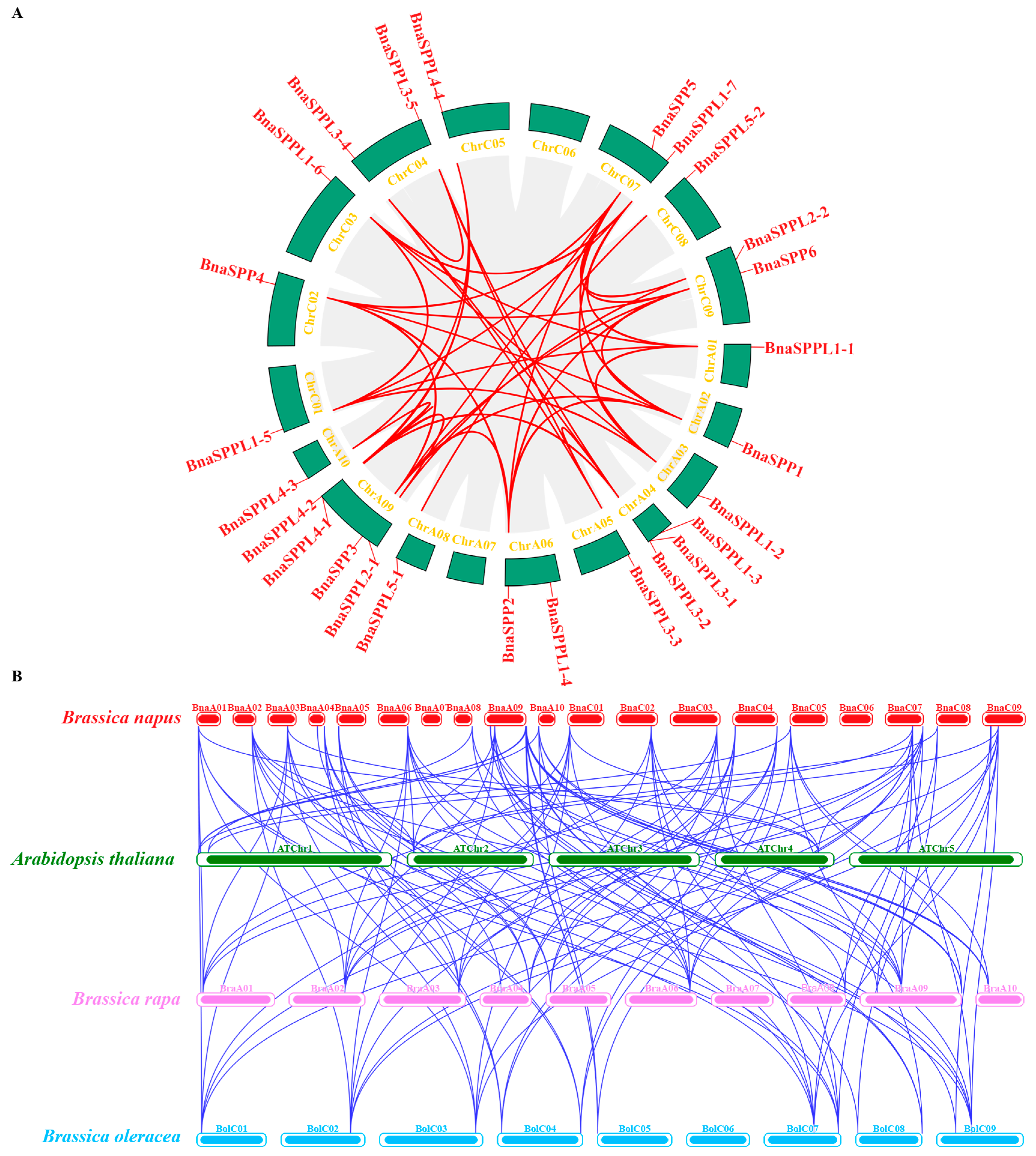
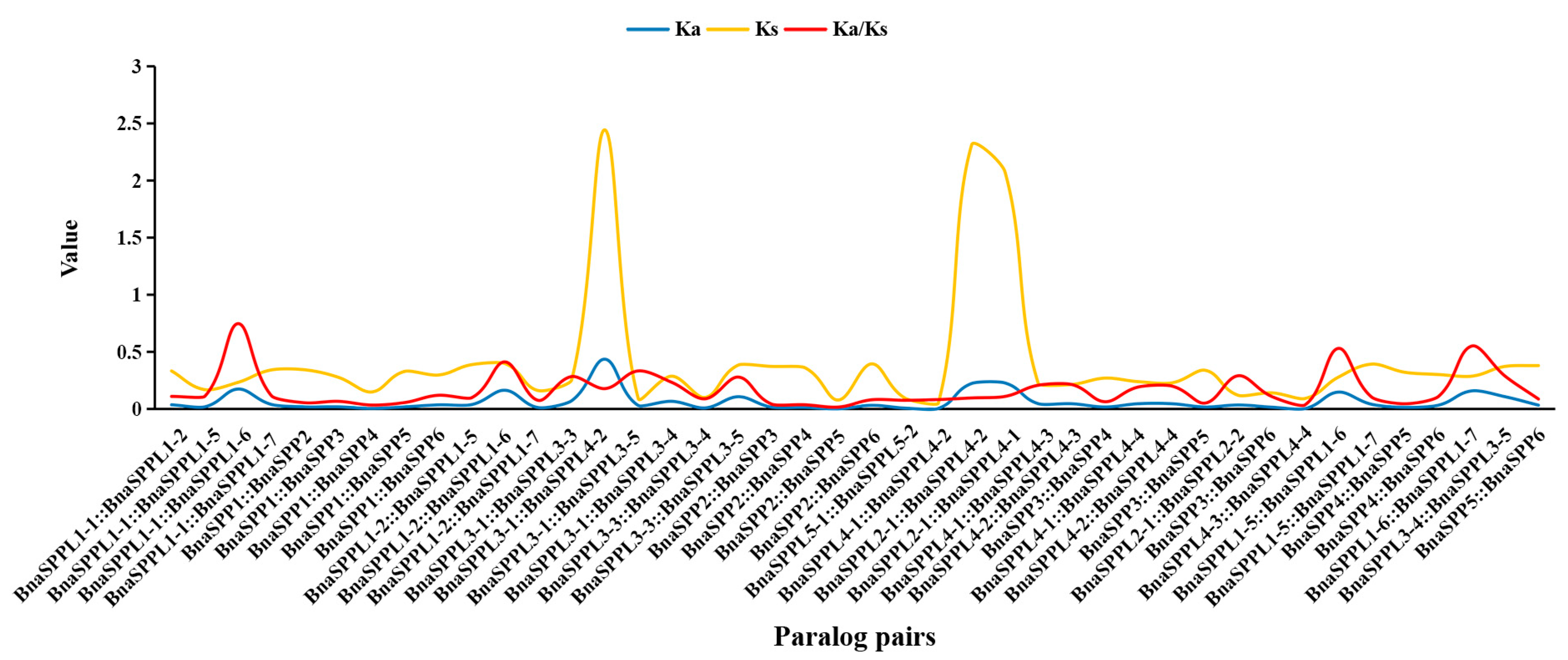
2.3. Chromosomal Localization and Collinearity Analysis of BnaSPP/SPPLs
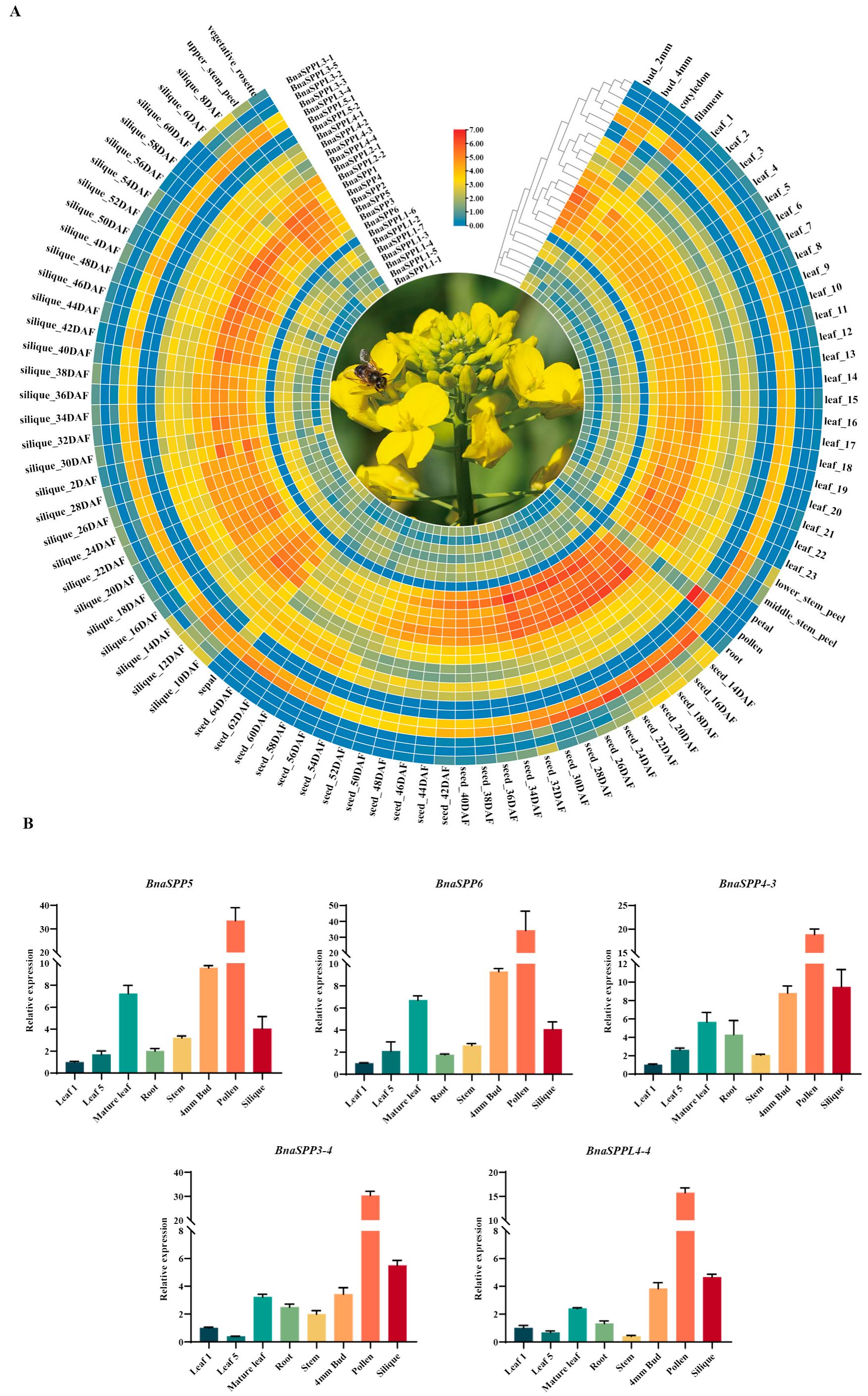
2.4. Analyzing the Expression Pattern of BnaSPP/SPPLs in Different Tissues
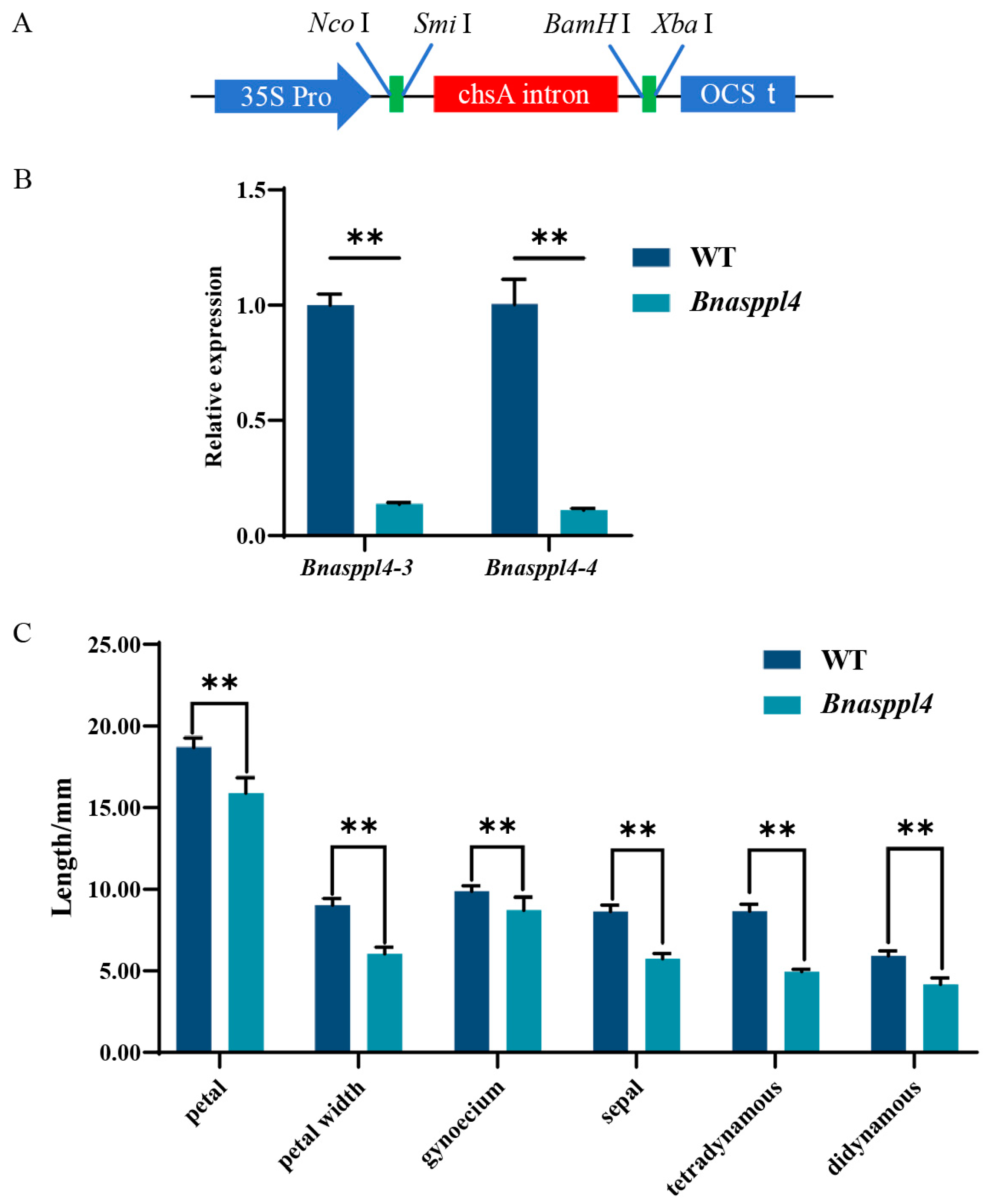
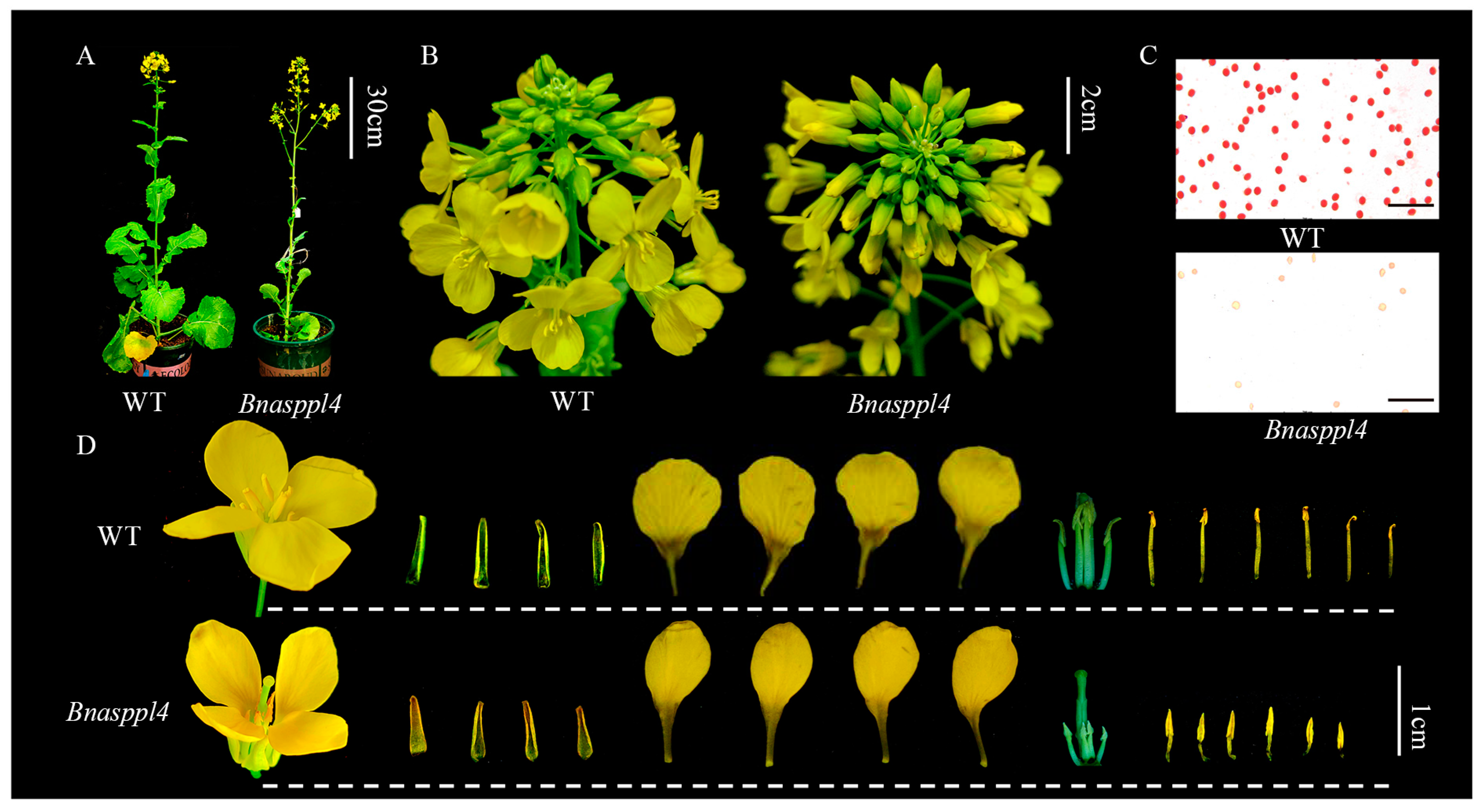
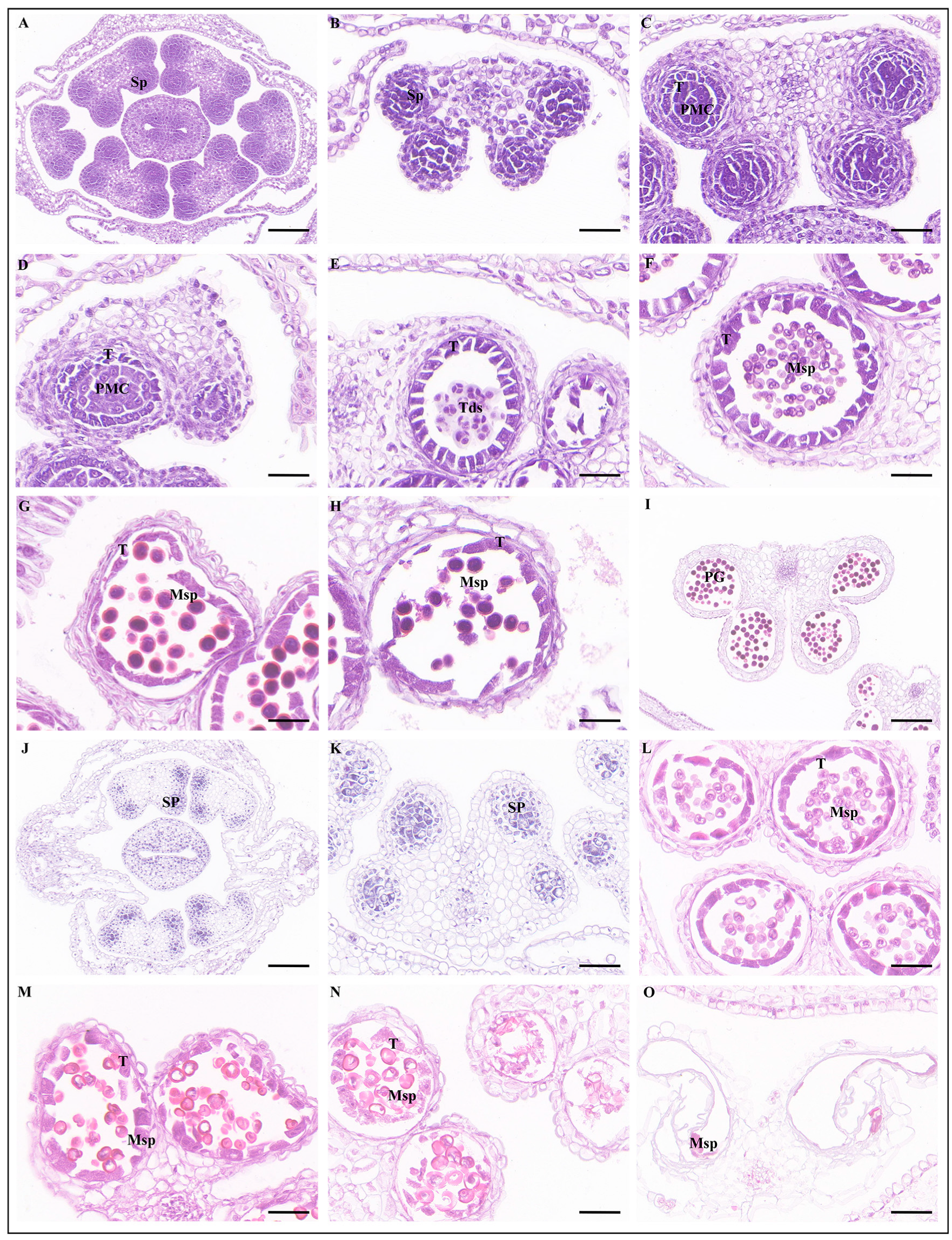
2.5. BnaSPPL4 Regulates Male Fertility in Rapeseed
3. Discussion
4. Materials and Methods
4.1. Identification of SPP/SPPL Family Members
4.2. Phylogenetic Analysis of Members of the SPP/SPPL Family
4.3. Chromosomal Localization and Covariance Analysis of BnaSPP/SPPLs
4.4. Expression Pattern Analysis of BnaSPP/SPPLs
4.5. Plasmid Construction and Genetic Transformation of Rapeseed
4.6. Phenotypic and Paraffin Section Observations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weihofen, A.; Binns, K.; Lemberg, M.K.; Ashman, K.; Martoglio, B. Identification of signal peptide peptidase, a presenilin-type aspartic protease. Science 2002, 296, 2215–2218. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, A.P.; Moliaka, Y.K.; Korovaitseva, G.I.; Rogaev, E.I. Novel class of polytopic proteins with domains associated with putative protease activity. Biochem.-Mosc. 2002, 67, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Hutton, M.; Nyborg, A.; Baker, M.; Jansen, K.; Golde, T.E. Identification of a novel family of presenilin homologues. Hum. Mol. Genet 2002, 11, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Mentrup, T.; Fluhrer, R.; Schröder, B. Latest emerging functions of SPP/SPPL intramembrane proteases. Eur. J. Cell Biol. 2017, 96, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Jurisch-Yaksi, N.; Sannerud, R.; Annaert, W. A fast growing spectrum of biological functions of γ-secretase in development and disease. Biochim. Biophys. Acta 2013, 1828, 2815–2827. [Google Scholar] [CrossRef] [PubMed]
- Fluhrer, R.; Steiner, H.; Haass, C. Intramembrane proteolysis by signal peptide peptidases: A comparative discussion of GXGD-type aspartyl proteases. J. Biol. Chem. 2009, 284, 13975–13979. [Google Scholar] [CrossRef] [PubMed]
- Krawitz, P.; Haffner, C.; Fluhrer, R.; Steiner, H.; Schmid, B.; Haass, C. Differential localization and identification of a critical aspartate suggest non-redundant proteolytic functions of the presenilin homologues SPPL2b and SPPL3. J. Biol. Chem. 2005, 280, 39515–39523. [Google Scholar] [CrossRef] [PubMed]
- Casso, D.J.; Liu, S.; Biehs, B.; Kornberg, T.B. Expression and characterization of Drosophila signal peptide peptidase-like (sppL), a gene that encodes an intramembrane protease. PLoS ONE 2012, 7, e33827. [Google Scholar] [CrossRef] [PubMed]
- Nyborg, A.C.; Ladd, T.B.; Jansen, K.; Kukar, T.; Golde, T.E. Intramembrane proteolytic cleavage by human signal peptide peptidase like 3 and malaria signal peptide peptidase. FASEB J. 2006, 20, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Asakura, T.; Uemura, T.; Ueda, T.; Terauchi, K.; Misaka, T.; Abe, K. Signal peptide peptidase and its homologs in Arabidopsis thaliana--plant tissue-specific expression and distinct subcellular localization. FEBS J. 2008, 275, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Friedmann, E.; Lemberg, M.K.; Weihofen, A.; Dev, K.K.; Dengler, U.; Rovelli, G.; Martoglio, B. Consensus analysis of signal peptide peptidase and homologous human aspartic proteases reveals opposite topology of catalytic domains compared with presenilins. J. Biol. Chem. 2004, 279, 50790–50798. [Google Scholar] [CrossRef]
- Haass, C. Take five--BACE and the gamma-secretase quartet conduct Alzheimer’s amyloid beta-peptide generation. EMBO J. 2004, 23, 483–488. [Google Scholar] [CrossRef]
- Fukumori, A.; Fluhrer, R.; Steiner, H.; Haass, C. Three-amino acid spacing of presenilin endoproteolysis suggests a general stepwise cleavage of gamma-secretase-mediated intramembrane proteolysis. J. Neurosci. 2010, 30, 7853–7862. [Google Scholar] [CrossRef]
- Nyborg, A.C.; Jansen, K.; Ladd, T.B.; Fauq, A.; Golde, T.E. A signal peptide peptidase (SPP) reporter activity assay based on the cleavage of type II membrane protein substrates provides further evidence for an inverted orientation of the SPP active site relative to presenilin. J. Biol. Chem. 2004, 279, 43148–43156. [Google Scholar] [CrossRef]
- De Strooper, B.; Annaert, W.; Cupers, P.; Saftig, P.; Craessaerts, K.; Mumm, J.S.; Schroeter, E.H.; Schrijvers, V.; Wolfe, M.S.; Ray, W.J.; et al. A presenilin-1-dependent gamma-secretase-like protease mediates release of Notch intracellular domain. Nature 1999, 398, 518–522. [Google Scholar] [CrossRef]
- De Strooper, B.; Saftig, P.; Craessaerts, K.; Vanderstichele, H.; Guhde, G.; Annaert, W.; Von Figura, K.; Van Leuven, F. Deficiency of presenilin-1 inhibits the normal cleavage of amyloid precursor protein. Nature 1998, 391, 387–390. [Google Scholar] [CrossRef]
- Chen, C.Y.; Malchus, N.S.; Hehn, B.; Stelzer, W.; Avci, D.; Langosch, D.; Lemberg, M.K. Signal peptide peptidase functions in ERAD to cleave the unfolded protein response regulator XBP1u. EMBO J. 2014, 33, 2492–2506. [Google Scholar] [CrossRef]
- Bat-Ochir, C.; Kwak, J.Y.; Koh, S.K.; Jeon, M.H.; Chung, D.; Lee, Y.W.; Chae, S.K. The signal peptide peptidase SppA is involved in sterol regulatory element-binding protein cleavage and hypoxia adaptation in Aspergillus nidulans. Mol. Microbiol. 2016, 100, 635–655. [Google Scholar] [CrossRef]
- Fluhrer, R.; Grammer, G.; Israel, L.; Condron, M.M.; Haffner, C.; Friedmann, E.; Böhland, C.; Imhof, A.; Martoglio, B.; Teplow, D.B.; et al. A gamma-secretase-like intramembrane cleavage of TNFalpha by the GxGD aspartyl protease SPPL2b. Nat. Cell Biol. 2006, 8, 894–896. [Google Scholar] [CrossRef]
- Friedmann, E.; Hauben, E.; Maylandt, K.; Schleeger, S.; Vreugde, S.; Lichtenthaler, S.F.; Kuhn, P.H.; Stauffer, D.; Rovelli, G.; Martoglio, B. SPPL2a and SPPL2b promote intramembrane proteolysis of TNFalpha in activated dendritic cells to trigger IL-12 production. Nat. Cell Biol. 2006, 8, 843–848. [Google Scholar] [CrossRef]
- Schneppenheim, J.; Dressel, R.; Hüttl, S.; Lüllmann-Rauch, R.; Engelke, M.; Dittmann, K.; Wienands, J.; Eskelinen, E.L.; Hermans-Borgmeyer, I.; Fluhrer, R.; et al. The intramembrane protease SPPL2a promotes B cell development and controls endosomal traffic by cleavage of the invariant chain. J. Exp. Med. 2013, 210, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Schneppenheim, J.; Hüttl, S.; Kruchen, A.; Fluhrer, R.; Müller, I.; Saftig, P.; Schneppenheim, R.; Martin, C.L.; Schröder, B. Signal-peptide-peptidase-like 2a is required for CD74 intramembrane proteolysis in human B cells. Biochem. Biophys. Res. Commun. 2014, 451, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Beisner, D.R.; Langerak, P.; Parker, A.E.; Dahlberg, C.; Otero, F.J.; Sutton, S.E.; Poirot, L.; Barnes, W.; Young, M.A.; Niessen, S.; et al. The intramembrane protease Sppl2a is required for B cell and DC development and survival via cleavage of the invariant chain. J. Exp. Med. 2013, 210, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, H.; Yabas, M.; Short, A.; Miosge, L.; Barthel, N.; Teh, C.E.; Roots, C.M.; Bull, K.R.; Jeelall, Y.; Horikawa, K.; et al. B cell survival, surface BCR and BAFFR expression, CD74 metabolism, and CD8- dendritic cells require the intramembrane endopeptidase SPPL2A. J. Exp. Med. 2013, 210, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Fluhrer, R.; Reiss, K.; Kremmer, E.; Saftig, P.; Haass, C. Regulated intramembrane proteolysis of Bri2 (Itm2b) by ADAM10 and SPPL2a/SPPL2b. J. Biol. Chem. 2008, 283, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, P.H.; Voss, M.; Haug-Kröper, M.; Schröder, B.; Schepers, U.; Bräse, S.; Haass, C.; Lichtenthaler, S.F.; Fluhrer, R. Secretome analysis identifies novel signal Peptide peptidase-like 3 (Sppl3) substrates and reveals a role of Sppl3 in multiple Golgi glycosylation pathways. Mol. Cell. Proteomics 2015, 14, 1584–1598. [Google Scholar] [CrossRef] [PubMed]
- Voss, M.; Künzel, U.; Higel, F.; Kuhn, P.H.; Colombo, A.; Fukumori, A.; Haug-Kröper, M.; Klier, B.; Grammer, G.; Seidl, A.; et al. Shedding of glycan-modifying enzymes by signal peptide peptidase-like 3 (SPPL3) regulates cellular N-glycosylation. EMBO J. 2014, 33, 2890–2905. [Google Scholar] [CrossRef]
- Voss, M.; Schröder, B.; Fluhrer, R. Mechanism, specificity, and physiology of signal peptide peptidase (SPP) and SPP-like proteases. Biochim. Biophys. Acta 2013, 1828, 2828–2839. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, J.; Mentrup, T.; Heidasch, R.; Müller, S.A.; Biswas, U.; Meyer, R.; Papadopoulou, A.A.; Dederer, V.; Haug-Kröper, M.; Adamski, V.; et al. The intramembrane protease SPPL2c promotes male germ cell development by cleaving phospholamban. EMBO Rep. 2019, 20, e46449. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, A.A.; Müller, S.A.; Mentrup, T.; Shmueli, M.D.; Niemeyer, J.; Haug-Kröper, M.; von Blume, J.; Mayerhofer, A.; Feederle, R.; Schröder, B.; et al. Signal peptide peptidase-like 2c impairs vesicular transport and cleaves SNARE proteins. EMBO Rep. 2019, 20, e46451. [Google Scholar] [CrossRef]
- Han, S.; Green, L.; Schnell, D.J. The signal peptide peptidase is required for pollen function in Arabidopsis. Plant Physiol. 2009, 149, 1289–1301. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef]
- Letian, C.; Yao-Guang, L. Male Sterility and Fertility Restoration in Crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef]
- Virmani, S.S.; Ilyas-Ahmed, M. Environment-sensitive genic male sterility (EGMS) in crops. Adv. Agron. 2001, 72, 139–195. [Google Scholar] [CrossRef]
- Adrain, C.; Freeman, M. New lives for old: Evolution of pseudoenzyme function illustrated by iRhoms. Nat. Rev. Mol. Cell Biol. 2012, 13, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Urny, J.; Hermans-Borgmeyer, I.; Gercken, G.; Schaller, H.C. Expression of the presenilin-like signal peptide peptidase (SPP) in mouse adult brain and during development. Gene Expr. Patterns 2003, 3, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Sato, T.; Wolfe, M.S. A C-terminal region of signal peptide peptidase defines a functional domain for intramembrane aspartic protease catalysis. J. Biol. Chem. 2007, 282, 20172–20179. [Google Scholar] [CrossRef] [PubMed]
- Casso, D.J.; Tanda, S.; Biehs, B.; Martoglio, B.; Kornberg, T.B. Drosophila signal peptide peptidase is an essential protease for larval development. Genetics 2005, 170, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Mergaert, P.; Nikovics, K.; Kelemen, Z.; Maunoury, N.; Vaubert, D.; Kondorosi, A.; Kondorosi, E. A novel family in Medicago truncatula consisting of more than 300 nodule-specific genes coding for small, secreted polypeptides with conserved cysteine motifs. Plant Physiol. 2003, 132, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Moreira, M.; Halbert, J.; Valloton, D.; Velten, B.; Chen, C.; Shao, Y.; Liechti, A.; Ascenção, K.; Rummel, C.; Ovchinnikova, S.; et al. Gene expression across mammalian organ development. Nature 2019, 571, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Hamblet, C.E.; Makowski, S.L.; Tritapoe, J.M.; Pomerantz, J.L. NK Cell Maturation and Cytotoxicity Are Controlled by the Intramembrane Aspartyl Protease SPPL3. J. Immunol. 2016, 196, 2614–2626. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.M.; Kim, J.; Cho, D.Y.; Choi, D.H.; Park, W.; Huh, S.J. Exome sequencing in a breast cancer family without BRCA mutation. Radiat. Oncol. J. 2015, 33, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Matthys-Rochon, E.; Vergne, P.; Detchepare, S.; Dumas, C. Male Germ Unit Isolation from Three Tricellular Pollen Species: Brassica oleracea, Zea mays, and Triticum aestivum. Plant Physiol. 1987, 83, 464–466. [Google Scholar] [CrossRef] [PubMed]
- Nicoud, Q.; Barrière, Q.; Busset, N.; Dendene, S.; Travin, D.; Bourge, M.; Le Bars, R.; Boulogne, C.; Lecroël, M.; Jenei, S.; et al. Sinorhizobium meliloti Functions Required for Resistance to Antimicrobial NCR Peptides and Bacteroid Differentiation. mBio 2021, 12, e0089521. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, R.B.; Beals, T.P.; Sanders, P.M. Anther development: Basic principles and practical applications. Plant Cell 1993, 5, 1217–1229. [Google Scholar] [CrossRef]
- Zhou, B.; Liu, Y.; Chen, Z.; Liu, D.; Wang, Y.; Zheng, J.; Liao, X.; Zhou, A.R. Comparative Transcriptome Analysis Reveals the Cause for Accumulation of Reactive Oxygen Species During Pollen Abortion in Cytoplasmic Male-Sterile Kenaf Line 722HA. Int. J. Mol. Sci. 2019, 20, 5515. [Google Scholar] [CrossRef] [PubMed]
- Uyttewaal, M.; Arnal, N.; Quadrado, M.; Martin-Canadell, A.; Vrielynck, N.; Hiard, S.; Gherbi, H.; Bendahmane, A.; Budar, F.; Mireau, H. Characterization of Raphanus sativus pentatricopeptide repeat proteins encoded by the fertility restorer locus for Ogura cytoplasmic male sterility. Plant Cell 2008, 20, 3331–3345. [Google Scholar] [CrossRef] [PubMed]
- Jing, B.; Heng, S.; Tong, D.; Wan, Z.; Fu, T.; Tu, J.; Ma, C.; Yi, B.; Wen, J.; Shen, J. A male sterility-associated cytotoxic protein ORF288 in Brassica juncea causes aborted pollen development. J. Exp. Bot. 2012, 63, 1285–1295. [Google Scholar] [CrossRef]
- Singh, M.; Brown, G.G. Characterization of expression of a mitochondrial gene region associated with the Brassica “Polima” CMS: Developmental influences. Curr. Genet. 1993, 24, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Guo, Y.; Ge, J.; Hu, Y.; Dong, J.; Dong, Z. Characterization of a new temperature-sensitive male sterile line SP2S in rapeseed (Brassica napus L.). Euphytica 2015, 206, 473–485. [Google Scholar] [CrossRef]
- Juan, G.; Yingfen, G.; Cheng-Yu, Y.; Guoyun, Z.; Jungang, D.; Dong, Z. Cytological Observation of Anther Development of Photoperiod/Thermo-sensitive Male Sterile Line Huiyou50S in Brassica napus. Acta Agron. Sin. 2012, 38, 541–548. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, D.; Wang, Z.; Guo, Y.; Sun, X.; Li, W.; Zhi, W.; Hu, S. Cytological observation of anther structure and genetic investigation of a thermo-sensitive genic male sterile line 373S in Brassica napus L. BMC Plant Biol. 2020, 20, 8. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, Q.; Wang, N.; Tao, B.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Fu, T.; Li, Q.; et al. Tapetal Expression of BnaC.MAGL8.a Causes Male Sterility in Arabidopsis. Front. Plant Sci. 2019, 10, 763. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Shukla, P.; Kirti, P.B. Targeted expression of a cysteine protease (AdCP) in tapetum induces male sterility in Indian mustard, Brassica juncea. Funct. Integr. Genom. 2019, 19, 703–714. [Google Scholar] [CrossRef]
- Zunaira, F.; Muhammad Nouman, R.; Muhammad Shoaib, F.; Yifan, L.; Huadong, W.; Mayra, A.; Jinxing, T.; Chaozhi, M.; Cheng, D.; Jing, W.; et al. Induction of Male Sterility by Targeted Mutation of a Restorer-of-Fertility Gene with CRISPR/Cas9-Mediated Genome Editing in Brassica napus L. Plants 2022, 11, 3501. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Zeng, X.; Wang, S.; Li, K.; Yuan, R.; Gao, H.; Luo, J.; Liu, F.; Wu, Y.; Li, Y.; et al. Aberrant Meiotic Prophase I Leads to Genic Male Sterility in the Novel TE5A Mutant of Brassica napus. Sci. Rep. 2016, 6, 33955. [Google Scholar] [CrossRef]
- Yu, C.; Xu, X.; Ge, J.; Guo, Y.; Dong, J.; Dong, Z. Premature breakdown of tapetum associated with reverse thermo-sensitive genic male-sterile line Huiyou50S in rapeseed (Brassica napus). Acta Physiol. Plant. 2016, 38, 54. [Google Scholar] [CrossRef]
- Song, J.M.; Liu, D.X.; Xie, W.Z.; Yang, Z.; Guo, L.; Liu, K.; Yang, Q.Y.; Chen, L.L. BnPIR: Brassica napus pan-genome information resource for 1689 accessions. Plant Biotechnol. J. 2021, 19, 412–414. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Elisabeth, G.; Alexandre, G.; Christine, H.; Ivan, I.; Ron, D.A.; Amos Marc, B. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Bolser, D.M.; Staines, D.M.; Perry, E.; Kersey, P.J. Ensembl Plants: Integrating Tools for Visualizing, Mining, and Analyzing Plant Genomic Data. Methods Mol. Biol. 2017, 1533, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. EvolView, an online tool for visualizing, annotating and managing phylogenetic trees. Nucleic Acids Res. 2012, 40, W569–W572. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, S.; Wei, L.; Huang, Y.; Liu, D.; Jia, Y.; Luo, C.; Lin, Y.; Liang, C.; Hu, Y.; et al. BnIR: A multi-omics database with various tools for Brassica napus research and breeding. Mol. Plant 2023, 16, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Location | Size (aa) | MW (Da) | PI | Instability Index | Aliphatic Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| BnaA01T0045500ZS | BnaSPPL1-1 | ChrA01:2433664:2434985 | 372 | 41018.71 | 8.04 | 42.55 | 119.01 | 0.677 | Plasma membrane |
| BnaA03T0536700ZS | BnaSPPL1-2 | ChrA03:30318596:30319895 | 369 | 40534.99 | 8.07 | 42.34 | 115.45 | 0.661 | Plasma membrane |
| BnaA04T0115800ZS | BnaSPPL1-3 | ChrA04:13250245:13250983 | 162 | 17983.62 | 8.44 | 33.35 | 120.8 | 0.722 | Plasma membrane |
| BnaA06T0148900ZS | BnaSPPL1-4 | ChrA06:9010737:9011720 | 181 | 20081.97 | 8.9 | 35.97 | 112.43 | 0.556 | Vacuole |
| BnaC01T0051800ZS | BnaSPPL1-5 | ChrC01:2869609:2870914 | 372 | 40985.54 | 8.38 | 44.11 | 118.47 | 0.69 | Plasma membrane |
| BnaC03T0754800ZS | BnaSPPL1-6 | ChrC03:73142539:73142914 | 86 | 9306.36 | 8.71 | 26.01 | 136.05 | 1.052 | Extracell |
| BnaC07T0513500ZS | BnaSPPL1-7 | ChrC07:58840667:58841964 | 369 | 40612.19 | 8 | 41.16 | 113.85 | 0.656 | Plasma membrane |
| BnaA09T0148500ZS | BnaSPPL2-1 | ChrA09:9059052:9062997 | 539 | 59646.09 | 5.83 | 30.28 | 117.37 | 0.465 | Plasma membrane |
| BnaC09T0161800ZS | BnaSPPL2-2 | ChrC09:12535426:12539195 | 523 | 58342.52 | 6.47 | 32.66 | 117.25 | 0.422 | Plasma membrane |
| BnaA04T0273700ZS | BnaSPPL3-1 | ChrA04:24217292:24217905 | 106 | 11527.72 | 5.15 | 37.04 | 113.96 | 0.632 | Plasma membrane |
| BnaA04T0273800ZS | BnaSPPL3-2 | ChrA04:24217912:24219967 | 316 | 35405.88 | 8.77 | 43.34 | 106.71 | 0.447 | Plasma membrane |
| BnaA05T0036100ZS | BnaSPPL3-3 | ChrA05:1990653:1993716 | 541 | 59360.25 | 5.02 | 45.91 | 109.74 | 0.45 | Plasma membrane |
| BnaC04T0039100ZS | BnaSPPL3-4 | ChrC04:3621314:3624386 | 541 | 59330.22 | 4.96 | 46.27 | 109.04 | 0.455 | Plasma membrane |
| BnaC04T0591400ZS | BnaSPPL3-5 | ChrC04:69507056:69509666 | 447 | 49674.56 | 9.02 | 48.29 | 106.82 | 0.301 | Plasma membrane |
| BnaA09T0696100ZS | BnaSPPL4-1 | ChrA09:64489204:64492337 | 539 | 59719.51 | 7.82 | 39.9 | 115.12 | 0.472 | Plasma membrane |
| BnaA09T0717600ZS | BnaSPPL4-2 | ChrA09:65650441:65653480 | 563 | 62510.74 | 8.04 | 41.33 | 113.68 | 0.448 | Plasma membrane |
| BnaA10T0003600ZS | BnaSPPL4-3 | ChrA10:210105:213788 | 540 | 59798.44 | 6.41 | 35.29 | 114.37 | 0.505 | Plasma membrane |
| BnaC05T0005500ZS | BnaSPPL4-4 | ChrC05:446704:450260 | 540 | 59710.27 | 6.21 | 35.34 | 114.74 | 0.507 | Plasma membrane |
| BnaA08T0309700ZS | BnaSPPL5-1 | ChrA08:27801075:27803833 | 542 | 59207.14 | 5.43 | 36.34 | 104.34 | 0.399 | Plasma membrane |
| BnaC08T0020500ZS | BnaSPPL5-2 | ChrC08:1651410:1654438 | 540 | 58824.06 | 5.81 | 38.24 | 105.43 | 0.463 | Plasma membrane |
| BnaA02T0326000ZS | BnaSPP1 | ChrA02:29422039:29424283 | 343 | 37885.76 | 8.65 | 34.41 | 109.42 | 0.613 | Plasma membrane |
| BnaA06T0399700ZS | BnaSPP2 | ChrA06:45757268:45759541 | 335 | 37165.11 | 8.82 | 32 | 112.03 | 0.655 | Plasma membrane |
| BnaA09T0223100ZS | BnaSPP3 | ChrA09:15794170:15796398 | 336 | 37266.19 | 8.81 | 34.5 | 112.53 | 0.654 | Plasma membrane |
| BnaC02T0439400ZS | BnaSPP4 | ChrC02:54167767:54170281 | 343 | 37915.78 | 8.65 | 36.31 | 109.13 | 0.6 | Plasma membrane |
| BnaC07T0282000ZS | BnaSPP5 | ChrC07:42592380:42594688 | 335 | 37139.07 | 8.82 | 32.31 | 112.33 | 0.665 | Plasma membrane |
| BnaC09T0259800ZS | BnaSPP6 | ChrC09:24544910:24547180 | 337 | 37553.64 | 9.15 | 36.28 | 113.06 | 0.659 | Plasma membrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Zhu, W.; Tian, M.; Liu, R.; Ruan, Y.; Liu, C. Genome-Wide Identification of the SPP/SPPL Gene Family and BnaSPPL4 Regulating Male Fertility in Rapeseed (Brassica napus L.). Int. J. Mol. Sci. 2024, 25, 3936. https://doi.org/10.3390/ijms25073936
Li G, Zhu W, Tian M, Liu R, Ruan Y, Liu C. Genome-Wide Identification of the SPP/SPPL Gene Family and BnaSPPL4 Regulating Male Fertility in Rapeseed (Brassica napus L.). International Journal of Molecular Sciences. 2024; 25(7):3936. https://doi.org/10.3390/ijms25073936
Chicago/Turabian StyleLi, Guangze, Wenjun Zhu, Minyu Tian, Rong Liu, Ying Ruan, and Chunlin Liu. 2024. "Genome-Wide Identification of the SPP/SPPL Gene Family and BnaSPPL4 Regulating Male Fertility in Rapeseed (Brassica napus L.)" International Journal of Molecular Sciences 25, no. 7: 3936. https://doi.org/10.3390/ijms25073936