
Suppression of Borna Disease Virus Replication during Its Persistent Infection Using the CRISPR/Cas13b System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
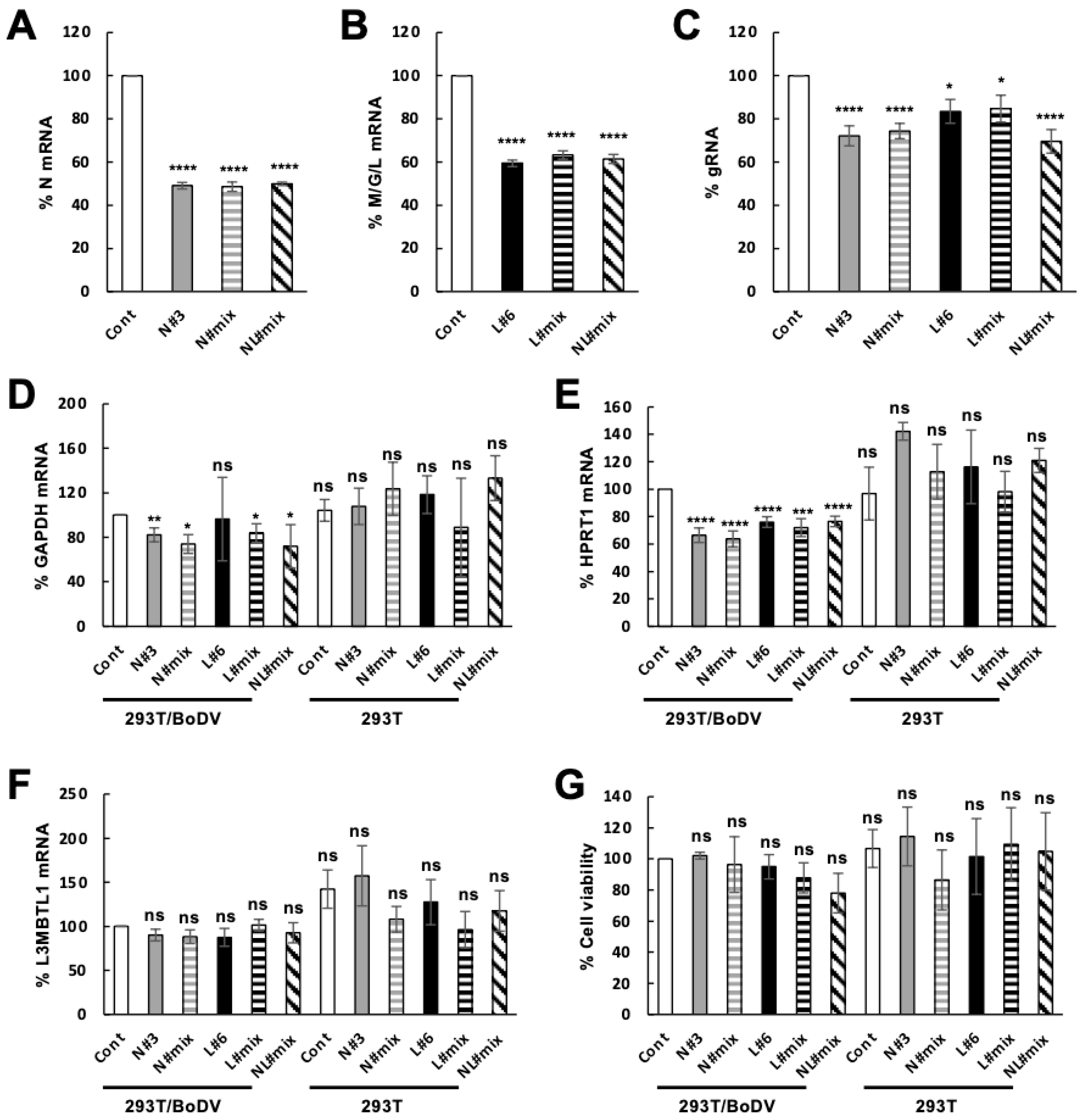
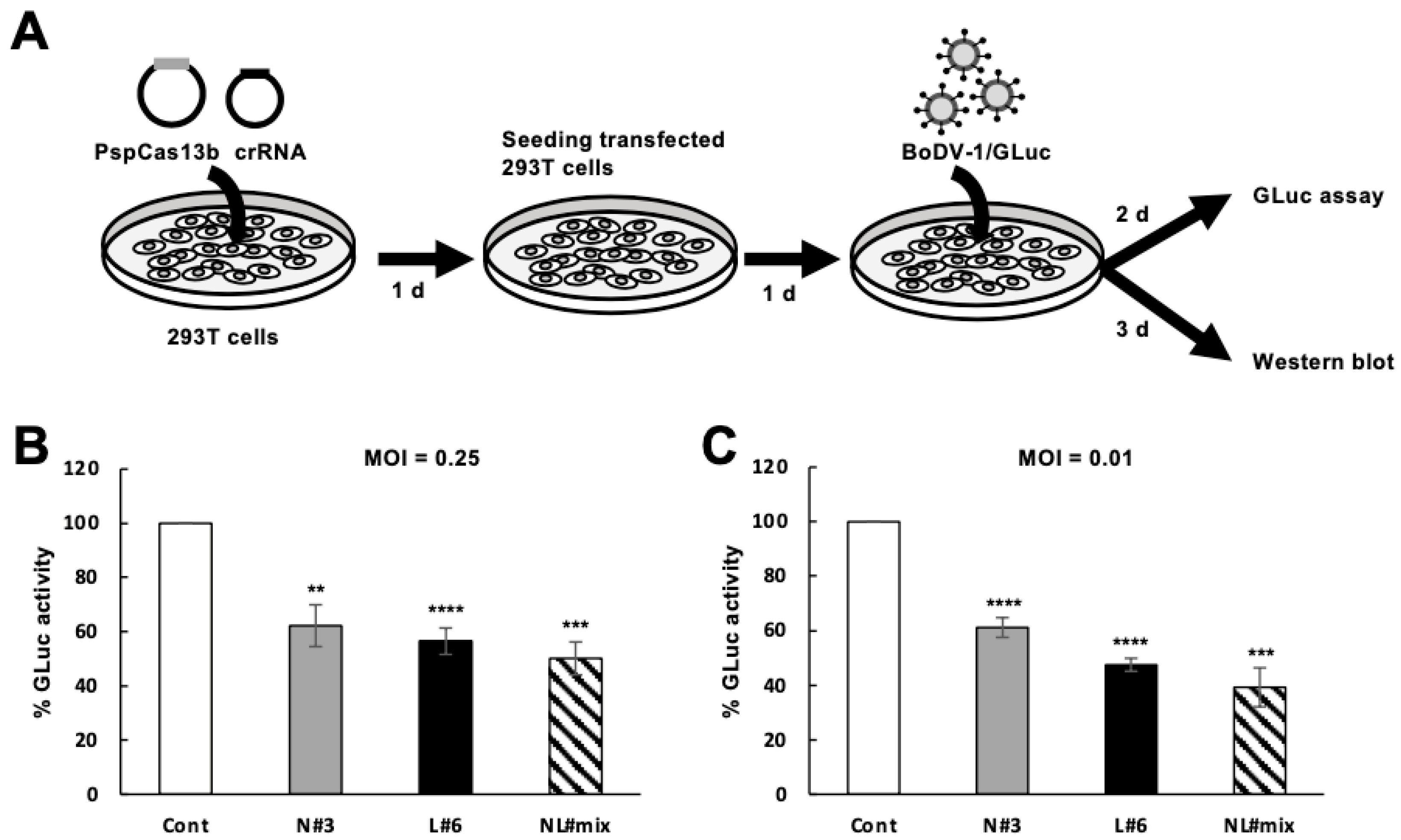
2. Results
2.1. Characterization of the CRISPR/Cas13b System in Viral mRNA Targeting in Persistently Infected Cells
2.2. Characterization of the CRISPR/Cas13b System Using Multiple crRNAs in Viral mRNA Targeting in Persistently Infected Cells
2.3. Downregulation of Viral Load in Persistently Infected Cells by the CRISPR/Cas13b System
2.4. Downregulation of De Novo BoDV-1 Infection by the CRISPR/Cas13b System
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Plasmids
4.3. Virus Preparation
4.4. Transfection
4.5. Virus Infection
4.6. Quantitative Real-Time RT-PCR (RT-qPCR)
4.7. Cell Viability Assay
4.8. Western Blot
4.9. GLuc Assay
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amarasinghe, G.K.; Ayllón, M.A.; Bào, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [PubMed]
- Staeheli, P.; Sauder, C.; Hausmann, J.; Ehrensperger, F.; Schwemmle, M. Epidemiology of Borna disease virus. J. Gen. Virol. 2000, 81, 2123–2135. [Google Scholar] [CrossRef]
- Honda, T. Bornavirus infection in human diseases and its molecular neuropathology. Clin. Exp. Neuroimmunol. 2022, 13, 7–16. [Google Scholar] [CrossRef]
- Schulze, V.; Große, R.; Fürstenau, J.; Forth, L.F.; Ebinger, A.; Richter, M.T.; Tappe, D.; Mertsch, T.; Klose, K.; Schlottau, K.; et al. Borna disease outbreak with high mortality in an alpaca herd in a previously unreported endemic area in Germany. Transbound. Emerg. Dis. 2020, 67, 2093–2107. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.; Tappe, D.; Höper, D.; Herden, C.; Boldt, A.; Mawrin, C.; Niederstraßer, O.; Müller, T.; Jenckel, M.; van der Grinten, E.; et al. A Variegated Squirrel Bornavirus Associated with Fatal Human Encephalitis. N. Engl. J. Med. 2015, 373, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Schlottau, K.; Forth, L.; Angstwurm, K.; Höper, D.; Zecher, D.; Liesche, F.; Hoffmann, B.; Kegel, V.; Seehofer, D.; Platen, S.; et al. Fatal Encephalitic Borna Disease Virus 1 in Solid-Organ Transplant Recipients. N. Engl. J. Med. 2018, 379, 1377–1379. [Google Scholar] [CrossRef] [PubMed]
- Korn, K.; Coras, R.; Bobinger, T.; Herzog, S.M.; Lücking, H.; Stöhr, R.; Huttner, H.B.; Hartmann, A.; Ensser, A. Fatal Encephalitis Associated with Borna Disease Virus 1. N. Engl. J. Med. 2018, 379, 1375–1377. [Google Scholar] [CrossRef] [PubMed]
- Eisermann, P.; Rubbenstroth, D.; Cadar, D.; Thomé-Bolduan, C.; Eggert, P.; Schlaphof, A.; Leypoldt, F.; Stangel, M.; Fortwängler, T.; Hoffmann, F.; et al. Active Case Finding of Current Bornavirus Infections in Human Encephalitis Cases of Unknown Etiology, Germany, 2018–2020. Emerg. Infect. Dis. 2021, 27, 1371–1379. [Google Scholar] [CrossRef]
- Grosse, L.; Lieftüchter, V.; Vollmuth, Y.; Hoffmann, F.; Olivieri, M.; Reiter, K.; Tacke, M.; Heinen, F.; Borggraefe, I.; Osterman, A.; et al. First detected geographical cluster of BoDV-1 encephalitis from same small village in two children: Therapeutic considerations and epidemiological implications. Infection 2023, 51, 1383–1398. [Google Scholar] [CrossRef]
- Hornig, M.; Solbrig, M.; Horscroft, N.; Weissenböck, H.; Lipkin, W.I. Borna disease virus infection of adult and neonatal rats: Models for neuropsychiatric disease. Curr. Top. Microbiol. Immunol. 2001, 253, 157–177. [Google Scholar] [CrossRef]
- Pletnikov, M.V.; Rubin, S.A.; Vasudevan, K.; Moran, T.H.; Carbone, K.M. Developmental brain injury associated with abnormal play behavior in neonatally Borna disease virus-infected Lewis rats: A model of autism. Behav. Brain Res. 1999, 100, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Pletnikov, M.V.; Moran, T.H.; Carbone, K.M. Borna disease virus infection of the neonatal rat: Developmental brain injury model of autism spectrum disorders. Front. Biosci. 2002, 7, d593–d607. [Google Scholar] [CrossRef] [PubMed]
- Pletnikov, M.V.; Jones, M.L.; Rubin, S.A.; Moran, T.H.; Carbone, K.M. Rat model of autism spectrum disorders. Genetic background effects on Borna disease virus-induced developmental brain damage. Ann. N. Y. Acad. Sci. 2001, 939, 318–319. [Google Scholar] [CrossRef]
- Lancaster, K.; Dietz, D.M.; Moran, T.H.; Pletnikov, M.V. Abnormal social behaviors in young and adult rats neonatally infected with Borna disease virus. Behav. Brain Res. 2007, 176, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Pletnikov, M.V.; Rubin, S.A.; Carbone, K.M.; Moran, T.H.; Schwartz, G.J. Neonatal Borna disease virus infection (BDV)-induced damage to the cerebellum is associated with sensorimotor deficits in developing Lewis rats. Brain Res. Dev. Brain Res. 2001, 126, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hans, A.; Bajramovic, J.J.; Syan, S.; Perret, E.; Dunia, I.; Brahic, M.; Gonzalez-Dunia, D. Persistent, noncytolytic infection of neurons by Borna disease virus interferes with ERK 1/2 signaling and abrogates BDNF-induced synaptogenesis. FASEB J. 2004, 18, 863–865. [Google Scholar] [CrossRef] [PubMed]
- Kamitani, W.; Ono, E.; Yoshino, S.; Kobayashi, T.; Taharaguchi, S.; Lee, B.-J.; Yamashita, M.; Kobayashi, T.; Okamoto, M.; Taniyama, H.; et al. Glial expression of Borna disease virus phosphoprotein induces behavioral and neurological abnormalities in transgenic mice. Proc. Natl. Acad. Sci. USA 2003, 100, 8969–8974. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Fujino, K.; Okuzaki, D.; Ohtaki, N.; Matsumoto, Y.; Horie, M.; Daito, T.; Itoh, M.; Tomonaga, K. Upregulation of insulin-like growth factor binding protein 3 in astrocytes of transgenic mice that express Borna disease virus phosphoprotein. J. Virol. 2011, 85, 4567–4571. [Google Scholar] [CrossRef]
- Teng, D.; Ueda, K.; Honda, T. Impact of Borna Disease Virus Infection on the Transcriptome of Differentiated Neuronal Cells and Its Modulation by Antiviral Treatment. Viruses 2023, 15, 942. [Google Scholar] [CrossRef]
- Prat, C.M.A.; Schmid, S.; Farrugia, F.; Cenac, N.; Le Masson, G.; Schwemmle, M.; Gonzalez-Dunia, D. Mutation of the protein kinase C site in borna disease virus phosphoprotein abrogates viral interference with neuronal signaling and restores normal synaptic activity. PLoS Pathog. 2009, 5, e1000425. [Google Scholar] [CrossRef]
- Zocher, M.; Czub, S.; Schulte-Mönting, J.; de La Torre, J.C.; Sauder, C. Alterations in neurotrophin and neurotrophin receptor gene expression patterns in the rat central nervous system following perinatal Borna disease virus infection. J. Neurovirol. 2000, 6, 462–477. [Google Scholar] [CrossRef] [PubMed]
- Nobach, D.; Müller, J.; Tappe, D.; Herden, C. Update on immunopathology of bornavirus infections in humans and animals. Adv. Virus Res. 2020, 107, 159–222. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, T.; Yamamoto, Y.; Sakai, M.; Tomonaga, K.; Honda, T. Antiviral activity of favipiravir (T-705) against mammalian and avian bornaviruses. Antivir. Res. 2017, 143, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Inagaki, H.; Araki, K.; Kariwa, H.; Arikawa, J.; Takashima, I. Inhibition of Borna disease virus replication by ribavirin in persistently infected cells. Arch. Virol. 1998, 143, 2039–2044. [Google Scholar] [CrossRef] [PubMed]
- Musser, J.M.B.; Heatley, J.J.; Koinis, A.V.; Suchodolski, P.F.; Guo, J.; Escandon, P.; Tizard, I.R. Ribavirin Inhibits Parrot Bornavirus 4 Replication in Cell Culture. PLoS ONE 2015, 10, e0134080. [Google Scholar] [CrossRef] [PubMed]
- Reuter, A.; Horie, M.; Höper, D.; Ohnemus, A.; Narr, A.; Rinder, M.; Beer, M.; Staeheli, P.; Rubbenstroth, D. Synergistic antiviral activity of Ribavirin and interferon-α against parrot bornaviruses in avian cells. J. Gen. Virol. 2016, 97, 2096–2103. [Google Scholar] [CrossRef] [PubMed]
- Teng, D.; Obika, S.; Ueda, K.; Honda, T. A Small Interfering RNA Cocktail Targeting the Nucleoprotein and Large Protein Genes Suppresses Borna Disease Virus Infection. Front. Microbiol. 2019, 10, 2781. [Google Scholar] [CrossRef] [PubMed]
- Hallensleben, W.; Staeheli, P. Inhibition of Borna disease virus multiplication by interferon: Cell line differences in susceptibility. Arch. Virol. 1999, 144, 1209–1216. [Google Scholar] [CrossRef]
- Teng, D.; Ueda, K.; Honda, T. HAND2 suppresses favipiravir efficacy in treatment of Borna disease virus infection. Antiviral Res. 2024, 222, 105812. [Google Scholar] [CrossRef]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef]
- Koonin, E.V.; Makarova, K.S.; Zhang, F. Diversity, classification and evolution of CRISPR-Cas systems. Curr. Opin. Microbiol. 2017, 37, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Deveau, H.; Garneau, J.E.; Moineau, S. CRISPR/Cas System and Its Role in Phage-Bacteria Interactions. Annu. Rev. Microbiol. 2010, 64, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Karginov, F.V.; Hannon, G.J. The CRISPR system: Small RNA-guided defense in bacteria and archaea. Mol. Cell 2010, 37, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR-Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA editing with CRISPR-Cas13. Science 2017, 358, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Konermann, S.; Lotfy, P.; Brideau, N.J.; Oki, J.; Shokhirev, M.N.; Hsu, P.D. Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell 2018, 173, 665–676.e14. [Google Scholar] [CrossRef]
- Kavuri, N.R.; Ramasamy, M.; Qi, Y.; Mandadi, K. Applications of CRISPR/Cas13-Based RNA Editing in Plants. Cells 2022, 11, 2665. [Google Scholar] [CrossRef]
- Yan, W.X.; Chong, S.; Zhang, H.; Makarova, K.S.; Koonin, E.V.; Cheng, D.R.; Scott, D.A. Cas13d Is a Compact RNA-Targeting Type VI CRISPR Effector Positively Modulated by a WYL-Domain-Containing Accessory Protein. Mol. Cell 2018, 70, 327–339.e5. [Google Scholar] [CrossRef]
- Freije, C.A.; Myhrvold, C.; Boehm, C.K.; Lin, A.E.; Welch, N.L.; Carter, A.; Metsky, H.C.; Luo, C.Y.; Abudayyeh, O.O.; Gootenberg, J.S.; et al. Programmable Inhibition and Detection of RNA Viruses Using Cas13. Mol. Cell 2019, 76, 826–837.e11. [Google Scholar] [CrossRef]
- Abbott, T.R.; Dhamdhere, G.; Liu, Y.; Lin, X.; Goudy, L.; Zeng, L.; Chemparathy, A.; Chmura, S.; Heaton, N.S.; Debs, R.; et al. Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influenza. Cell 2020, 181, 865–876.e12. [Google Scholar] [CrossRef]
- Xu, C.; Zhou, Y.; Xiao, Q.; He, B.; Geng, G.; Wang, Z.; Cao, B.; Dong, X.; Bai, W.; Wang, Y.; et al. Programmable RNA editing with compact CRISPR–Cas13 systems from uncultivated microbes. Nat. Methods 2021, 18, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, J.; Wang, Q.; Wang, Y.; Kang, C. Rapid design and development of CRISPR-Cas13a targeting SARS-CoV-2 spike protein. Theranostics 2021, 11, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Singsuksawat, E.; Onnome, S.; Posiri, P.; Suphatrakul, A.; Srisuk, N.; Nantachokchawapan, R.; Praneechit, H.; Sae-kow, C.; Chidpratum, P.; Sangiamsuntorn, K.; et al. Potent programmable antiviral against dengue virus in primary human cells by Cas13b RNP with short spacer and delivery by VLP. Mol. Ther. Methods Clin. Dev. 2021, 21, 729–740. [Google Scholar] [CrossRef]
- Li, H.; Wang, S.; Dong, X.; Li, Q.; Li, M.; Li, J.; Guo, Y.; Jin, X.; Zhou, Y.; Song, H.; et al. CRISPR-Cas13a Cleavage of Dengue Virus NS3 Gene Efficiently Inhibits Viral Replication. Mol. Ther. Nucleic Acids 2020, 19, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.U.; Salman, H.M.; Khalid, M.F.; Khan, M.H.F.; Anwar, S.; Afzal, S.; Idrees, M.; Chaudhary, S.U. CRISPR-Cas13a mediated targeting of hepatitis C virus internal-ribosomal entry site (IRES) as an effective antiviral strategy. Biomed. Pharmacother. 2021, 136, 111239. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhao, F.; Sun, H.; Wang, Z.; Huang, Y.; Zhu, W.; Xu, F.; Mei, S.; Liu, X.; Zhang, D.; et al. CRISPR-Cas13a Inhibits HIV-1 Infection. Mol. Ther. Nucleic Acids 2020, 21, 147–155. [Google Scholar] [CrossRef]
- Nguyen, H.; Wilson, H.; Jayakumar, S.; Kulkarni, V.; Kulkarni, S. Efficient Inhibition of HIV Using CRISPR/Cas13d Nuclease System. Viruses 2021, 13, 1850. [Google Scholar] [CrossRef]
- Tomonaga, K.; Kobayashi, T.; Ikuta, K. Molecular and cellular biology of Borna disease virus infection. Microbes Infect. 2002, 4, 491–500. [Google Scholar] [CrossRef]
- Honda, T.; Tomonaga, K. Nucleocytoplasmic shuttling of viral proteins in borna disease virus infection. Viruses 2013, 5, 1978–1990. [Google Scholar] [CrossRef]
- Rudolph, M.G.; Kraus, I.; Dickmanns, A.; Eickmann, M.; Garten, W.; Ficner, R. Crystal structure of the borna disease virus nucleoprotein. Structure 2003, 11, 1219–1226. [Google Scholar] [CrossRef]
- Poenisch, M.; Wille, S.; Ackermann, A.; Staeheli, P.; Schneider, U. The X protein of borna disease virus serves essential functions in the viral multiplication cycle. J. Virol. 2007, 81, 7297–7299. [Google Scholar] [CrossRef] [PubMed]
- Neumann, P.; Lieber, D.; Meyer, S.; Dautel, P.; Kerth, A.; Kraus, I.; Garten, W.; Stubbs, M.T. Crystal structure of the Borna disease virus matrix protein (BDV-M) reveals ssRNA binding properties. Proc. Natl. Acad. Sci. USA 2009, 106, 3710–3715. [Google Scholar] [CrossRef] [PubMed]
- Bajramovic, J.J.; Münter, S.; Syan, S.; Nehrbass, U.; Brahic, M.; Gonzalez-Dunia, D. Borna disease virus glycoprotein is required for viral dissemination in neurons. J. Virol. 2003, 77, 12222–12231. [Google Scholar] [CrossRef] [PubMed]
- Mayer, D.; Baginsky, S.; Schwemmle, M. Isolation of viral ribonucleoprotein complexes from infected cells by tandem affinity purification. Proteomics 2005, 5, 4483–4487. [Google Scholar] [CrossRef] [PubMed]
- Cubitt, B.; Oldstone, C.; Valcarcel, J.; de la Torre, J.C. RNA splicing contributes to the generation of mature mRNAs of Borna disease virus, a non-segmented negative strand RNA virus. Virus Res. 1994, 34, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.A.; Schneemann, A.; Lipkin, W.I. RNA splicing in Borna disease virus, a nonsegmented, negative-strand RNA virus. J. Virol. 1994, 68, 5007–5012. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [PubMed]
- Bot, J.F.; van der Oost, J.; Geijsen, N. The double life of CRISPR–Cas13. Curr. Opin. Biotechnol. 2022, 78, 102789. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, X.; Zhou, J.; Yang, C.; Wang, G.; Tan, Y.; Wu, Y.; Zhang, S.; Yi, K.; Kang, C. The CRISPR-Cas13a Gene-Editing System Induces Collateral Cleavage of RNA in Glioma Cells. Adv. Sci. 2019, 6, 1901299. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Q.; Liu, Q.; Zheng, Y.; Zheng, C.; Yi, K.; Zhao, Y.; Gu, Y.; Wang, Y.; Wang, C.; et al. Dual-Locking Nanoparticles Disrupt the PD-1/PD-L1 Pathway for Efficient Cancer Immunotherapy. Adv. Mater. 2019, 31, 1905751. [Google Scholar] [CrossRef]
- Özcan, A.; Krajeski, R.; Ioannidi, E.; Lee, B.; Gardner, A.; Makarova, K.S.; Koonin, E.V.; Abudayyeh, O.O.; Gootenberg, J.S. Programmable RNA targeting with the single-protein CRISPR effector Cas7-11. Nature 2021, 597, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Buchman, A.; Brogan, D.J.; Sun, R.; Yang, T.; Hsu, P.; Akbari, O.S. Programmable RNA Targeting Using CasRx in Flies. Cris. J. 2020, 3, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Liang, D.; Wilusz, J.E. CRISPR/Cas13 effectors have differing extents of off-target effects that limit their utility in eukaryotic cells. Nucleic Acids Res. 2022, 50, e65. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Huang, J.; Xiao, Q.; He, B.; Dong, X.; Liu, Y.; Yang, X.; Han, D.; Wang, Z.; Wang, X.; et al. High-fidelity Cas13 variants for targeted RNA degradation with minimal collateral effects. Nat. Biotechnol. 2022, 41, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Guo, X.; Li, Z.; Cao, L.; Liu, S.; Guo, Y.; Wang, G.; Luo, Y.; Zhang, Z.; et al. The collateral activity of RfxCas13d can induce lethality in a RfxCas13d knock-in mouse model. Genome Biol. 2023, 24, 20. [Google Scholar] [CrossRef] [PubMed]
- Kelley, C.P.; Haerle, M.C.; Wang, E.T. Negative autoregulation mitigates collateral RNase activity of repeat-targeting CRISPR-Cas13d in mammalian cells. Cell Rep. 2022, 40, 111226. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Techakriengkrai, N.; Nedumpun, T.; Suradhat, S. Abrogation of PRRSV infectivity by CRISPR-Cas13b-mediated viral RNA cleavage in mammalian cells. Sci. Rep. 2020, 10, 9617. [Google Scholar] [CrossRef]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Smargon, A.A.; Cox, D.B.T.; Pyzocha, N.K.; Zheng, K.; Slaymaker, I.M.; Gootenberg, J.S.; Abudayyeh, O.A.; Essletzbichler, P.; Shmakov, S.; Makarova, K.S.; et al. Cas13b Is a Type VI-B CRISPR-Associated RNA-Guided RNase Differentially Regulated by Accessory Proteins Csx27 and Csx28. Mol. Cell 2017, 65, 618–630.e7. [Google Scholar] [CrossRef]
- Schneider, P.A.; Briese, T.; Zimmermann, W.; Ludwig, H.; Lipkin, W.I. Sequence conservation in field and experimental isolates of Borna disease virus. J. Virol. 1994, 68, 63–68. [Google Scholar] [CrossRef]
- Proal, A.D.; VanElzakker, M.B. Long COVID or Post-acute Sequelae of COVID-19 (PASC): An Overview of Biological Factors That May Contribute to Persistent Symptoms. Front. Microbiol. 2021, 12, 698169. [Google Scholar] [CrossRef] [PubMed]
- Stein, S.R.; Ramelli, S.C.; Grazioli, A.; Chung, J.Y.; Singh, M.; Yinda, C.K.; Winkler, C.W.; Sun, J.; Dickey, J.M.; Ylaya, K.; et al. SARS-CoV-2 infection and persistence in the human body and brain at autopsy. Nature 2022, 612, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Bussani, R.; Zentilin, L.; Correa, R.; Colliva, A.; Silvestri, F.; Zacchigna, S.; Collesi, C.; Giacca, M. Persistent SARS-CoV-2 infection in patients seemingly recovered from COVID-19. J. Pathol. 2023, 259, 254–263. [Google Scholar] [CrossRef] [PubMed]
- McKinlay, C.J.; Vargas, J.R.; Blake, T.R.; Hardy, J.W.; Kanada, M.; Contag, C.H.; Wender, P.A.; Waymouth, R.M. Charge-altering releasable transporters (CARTs) for the delivery and release of mRNA in living animals. Proc. Natl. Acad. Sci. USA 2017, 114, E448–E456. [Google Scholar] [CrossRef] [PubMed]
- Hendel, A.; Bak, R.O.; Clark, J.T.; Kennedy, A.B.; Ryan, D.E.; Roy, S.; Steinfeld, I.; Lunstad, B.D.; Kaiser, R.J.; Wilkens, A.B.; et al. Chemically modified guide RNAs enhance CRISPR-Cas genome editing in human primary cells. Nat. Biotechnol. 2015, 33, 985–989. [Google Scholar] [CrossRef]
- Sago, C.D.; Lokugamage, M.P.; Paunovska, K.; Vanover, D.A.; Monaco, C.M.; Shah, N.N.; Castro, M.G.; Anderson, S.E.; Rudoltz, T.G.; Lando, G.N.; et al. High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proc. Natl. Acad. Sci. USA 2018, 115, E9944–E9952. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.F.; Chen, G.J.; Luo, Y.L.; Zhang, Y.; Zhao, G.; Lu, Z.D.; Czarna, A.; Gu, Z.; Wang, J. Rational designs of in vivo CRISPR-Cas delivery systems. Adv. Drug Deliv. Rev. 2021, 168, 3–29. [Google Scholar] [CrossRef]
- Nakamura, Y.; Takahashi, H.; Shoya, Y.; Nakaya, T.; Watanabe, M.; Tomonaga, K.; Iwahashi, K.; Ameno, K.; Momiyama, N.; Taniyama, H.; et al. Isolation of Borna Disease Virus from Human Brain Tissue. J. Virol. 2000, 74, 4601–4611. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Tomonaga, K.; Honda, T. Development of an RNA Virus-Based Episomal Vector Capable of Switching Transgene Expression. Front. Microbiol. 2019, 10, 2485. [Google Scholar] [CrossRef]
- Pleschka, S.; Staeheli, P.; Kolodziejek, J.; Richt, J.A.; Nowotny, N.; Schwemmle, M. Conservation of coding potential and terminal sequences in four different isolates of Borna disease virus. J. Gen. Virol. 2001, 82, 2681–2690. [Google Scholar] [CrossRef]
- Nakayama, R.; Ueno, Y.; Ueda, K.; Honda, T. Latent infection with Kaposi’s sarcoma-associated herpesvirus enhances retrotransposition of long interspersed element-1. Oncogene 2019, 38, 4340–4351. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Nakayama, R.; Obika, S.; Ohsaki, E.; Ueda, K.; Honda, T. Inhibition of LINE-1 Retrotransposition by Capsaicin. Int. J. Mol. Sci. 2018, 19, 3243. [Google Scholar] [CrossRef]
- Hayashi, Y.; Horie, M.; Daito, T.; Honda, T.; Ikuta, K.; Tomonaga, K. Heat shock cognate protein 70 controls Borna disease virus replication via interaction with the viral non-structural protein X. Microbes Infect. 2009, 11, 394–402. [Google Scholar] [CrossRef]
- Kojima, S.; Honda, T.; Matsumoto, Y.; Tomonaga, K. Heat stress is a potent stimulus for enhancing rescue efficiency of recombinant Borna disease virus. Microbiol. Immunol. 2014, 58, 636–642. [Google Scholar] [CrossRef]
- Honda, T.; Nishikawa, Y.; Nishimura, K.; Teng, D.; Takemoto, K.; Ueda, K. Effects of activation of the LINE-1 antisense promoter on the growth of cultured cells. Sci. Rep. 2020, 10, 22136. [Google Scholar] [CrossRef]
- Quantification of Gel Bands by an Image J Macro, Band/Peak Quantification Tool. Available online: https://www.protocols.io/view/quantification-of-gel-bands-by-an-image-j-macro-ba-bp2l6n4bkgqe/v1 (accessed on 23 January 2024).




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, S.; Ogawa, H.; Katoh, H.; Honda, T. Suppression of Borna Disease Virus Replication during Its Persistent Infection Using the CRISPR/Cas13b System. Int. J. Mol. Sci. 2024, 25, 3523. https://doi.org/10.3390/ijms25063523
Sasaki S, Ogawa H, Katoh H, Honda T. Suppression of Borna Disease Virus Replication during Its Persistent Infection Using the CRISPR/Cas13b System. International Journal of Molecular Sciences. 2024; 25(6):3523. https://doi.org/10.3390/ijms25063523
Chicago/Turabian StyleSasaki, Shigenori, Hirohito Ogawa, Hirokazu Katoh, and Tomoyuki Honda. 2024. "Suppression of Borna Disease Virus Replication during Its Persistent Infection Using the CRISPR/Cas13b System" International Journal of Molecular Sciences 25, no. 6: 3523. https://doi.org/10.3390/ijms25063523