
Effect of HY7602 Fermented Deer Antler on Physical Fatigue and Antioxidant Activity in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
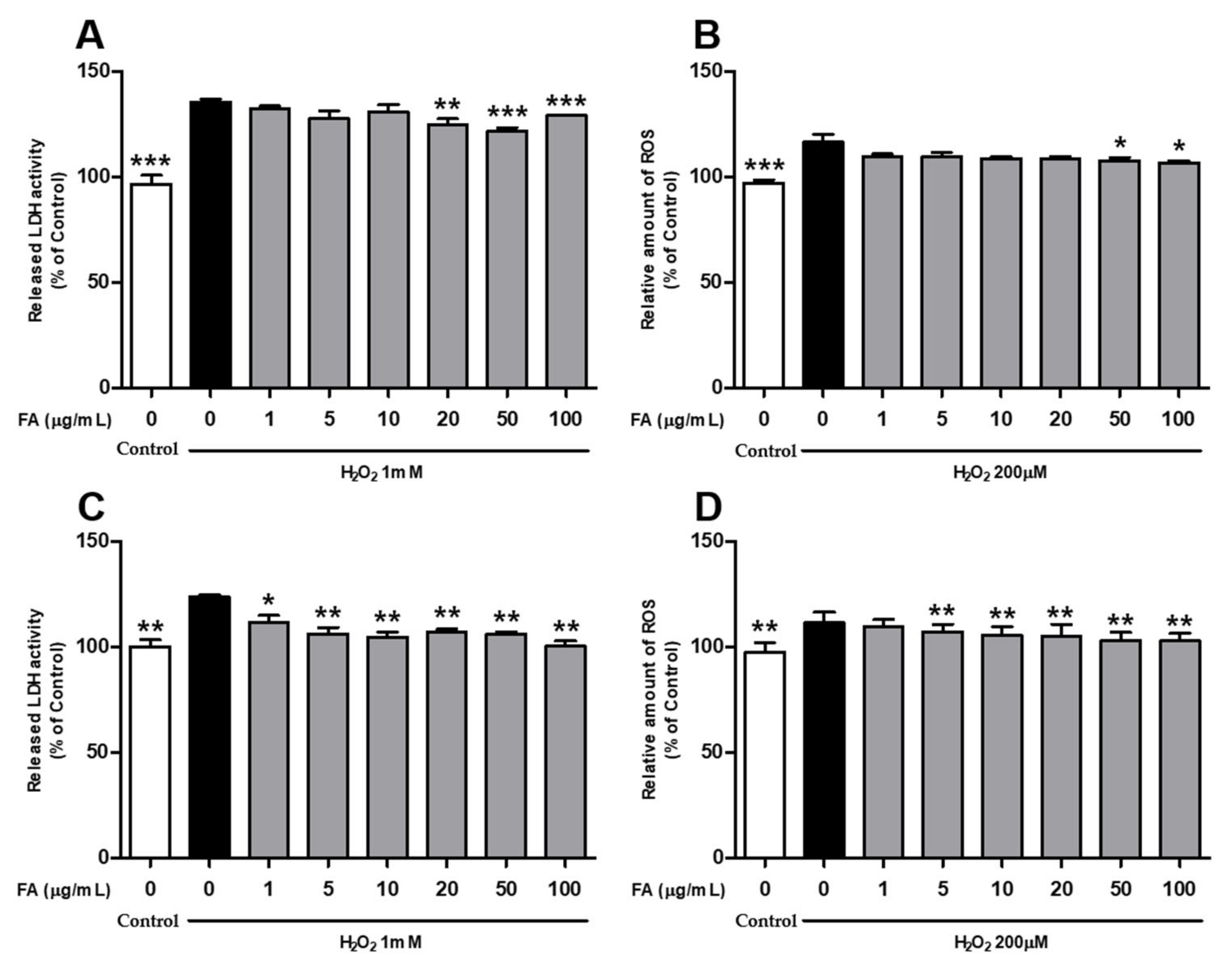
2.1. L. curvatus HY7602-Fermented Antler Inhibits Oxidative Stress Damage in Cells
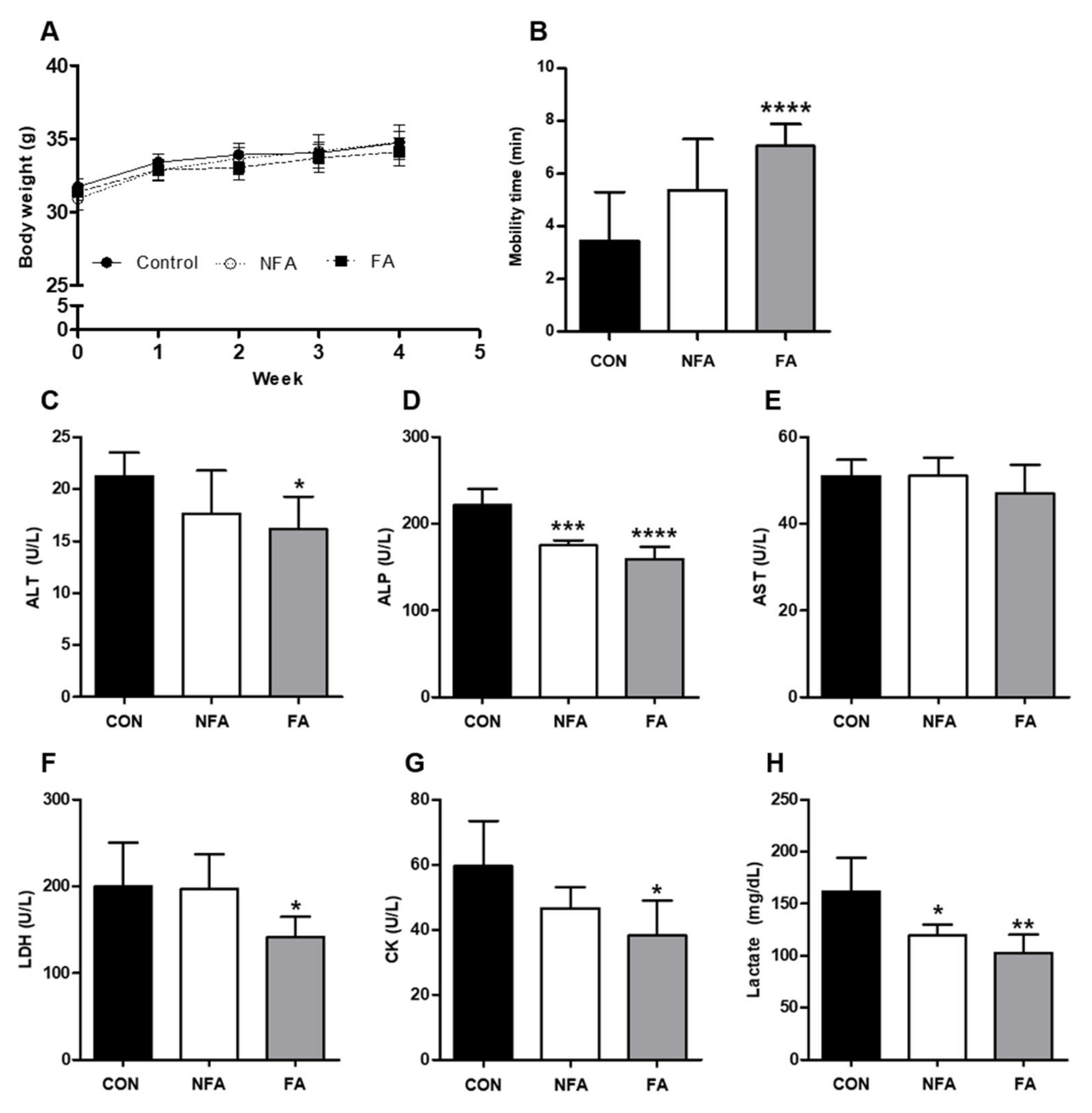
2.2. L. curvatus HY7602 Fermented Antler Ameliorates Physical Fatigue
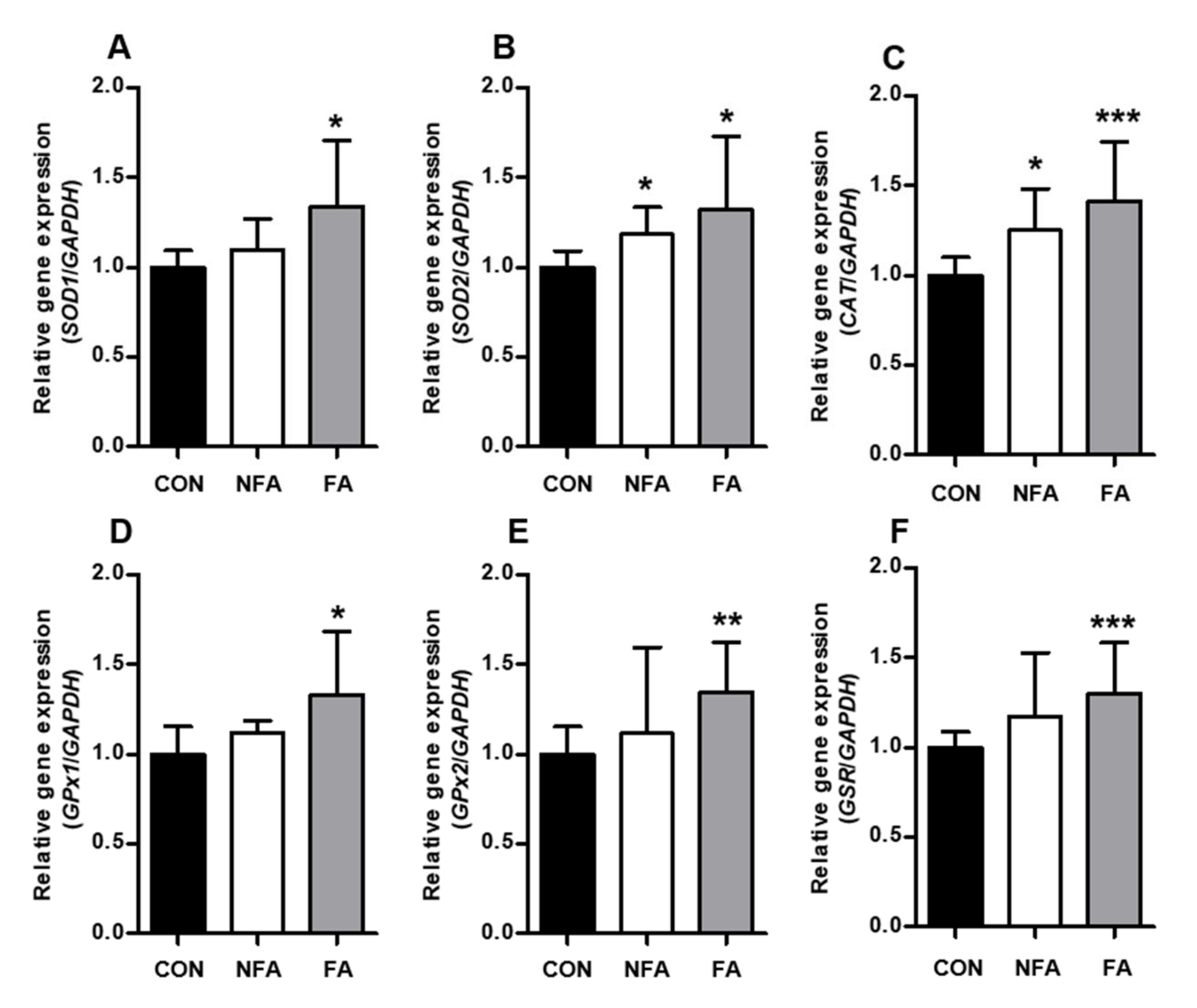
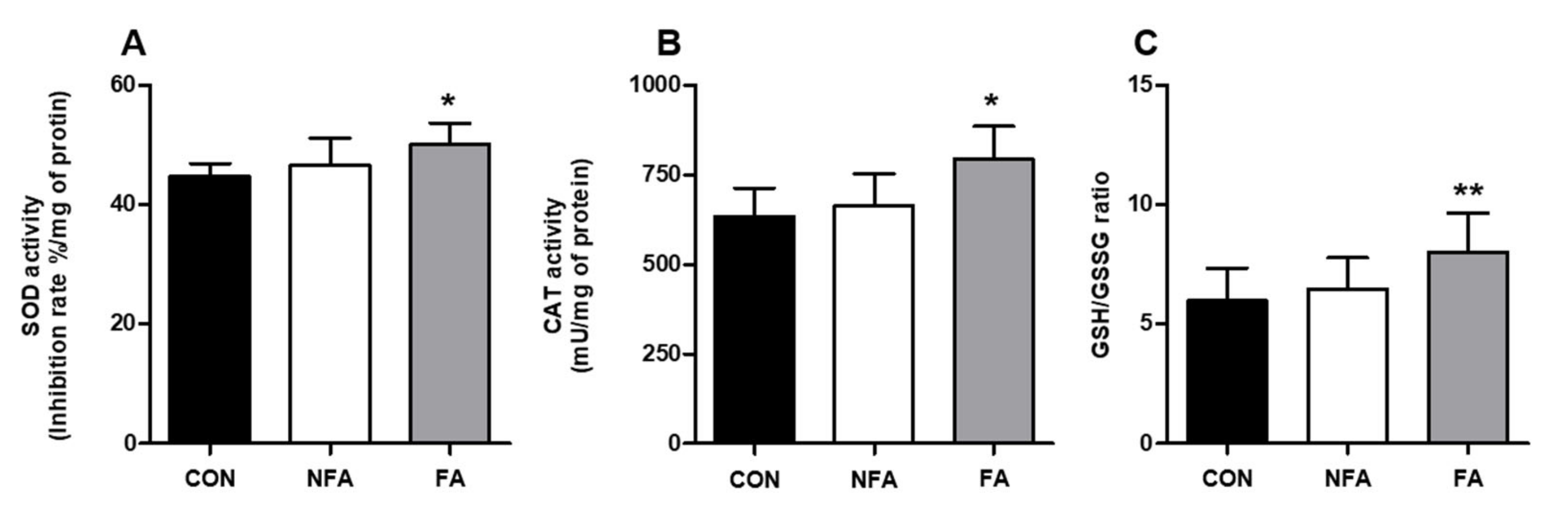
2.3. L. curvatus HY7602 Fermented Antler Upregulates Antioxidant-Related Genes and Antioxidant Enzyme Activity in the Liver
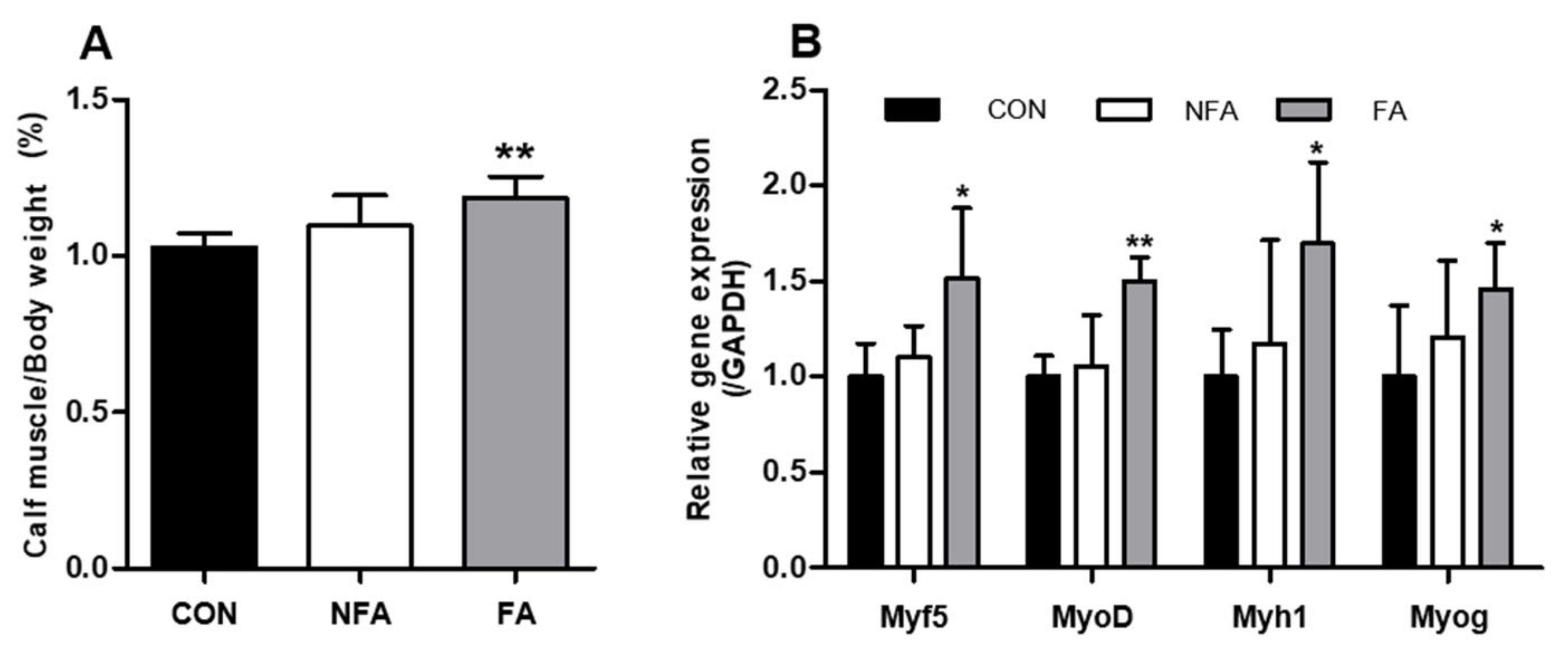
2.4. L. curvatus HY7602 Fermented Antler Promotes Muscle Regeneration
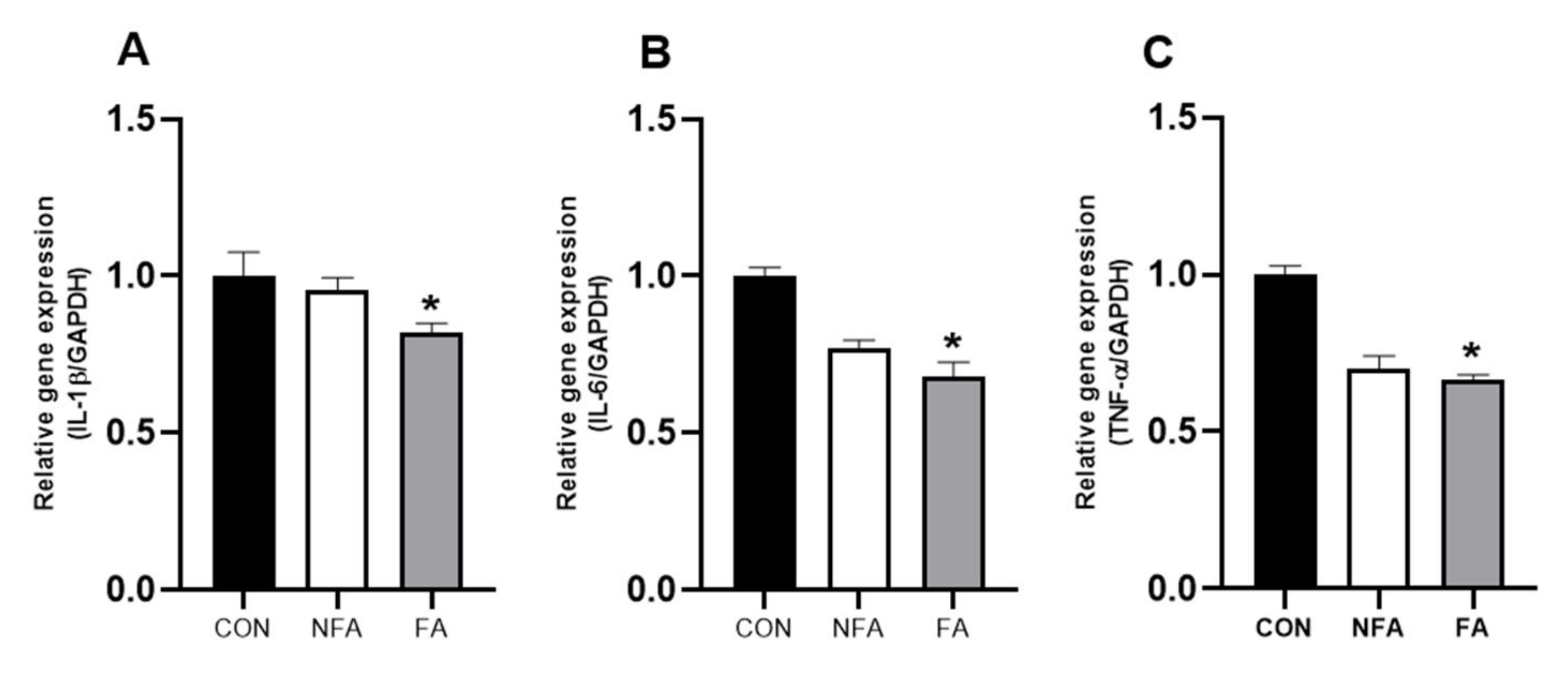
2.5. L. curvatus HY7602 Fermented Antler Downregulates the mRNA Expression of Cytokines in Muscle
3. Discussion
4. Materials and Methods
4.1. Preparation of L. curvatus HY7602-Fermented Deer Antler
4.2. In Vitro Studies
4.2.1. Cell Culture
4.2.2. Measurement of Cell Viability
4.2.3. Determination of ROS Levels
4.3. Animals and Diets
4.4. Forced Swimming Test
4.5. Sample Collection and Serum Biochemistry
4.6. Real-Time Polymerase Chain Reaction (RT-PCR)
4.7. Evaluation of Antioxidant Enzyme Activities
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Behrens, M.; Gube, M.; Chaabene, H.; Prieske, O.; Zenon, A.; Broscheid, K.C.; Schega, L.; Husmann, F.; Weippert, M. Fatigue and human performance: An updated framework. Sports Med. 2023, 53, 7–31. [Google Scholar] [CrossRef]
- Ware, N.C. Society, mind and body in chronic fatigue syndrome: An anthropological view. Ciba Found. Symp. 1993, 173, 62–73, discussion 73–82. [Google Scholar]
- Abd-Elfattah, H.M.F.; Abdelazeim, H.M.; Elshennawy, S.; Elshennawy, S. Physical and cognitive consequences of fatigue: A review. J. Adv. Res. 2015, 6, 351–358. [Google Scholar] [CrossRef]
- Gruet, M.; Temesi, J.; Rupp, T.; Levy, P.; Millet, G.Y.; Verges, S. Stimulation of the motor cortex and corticospinal tract to assess human muscle fatigue. Neuroscience 2013, 231, 384–399. [Google Scholar] [CrossRef]
- Van Cutsem, J.; Marcora, S.; De Pauw, K.; Bailey, S.; Meeusen, R.; Roelands, B. The effects of mental fatigue on physical performance: A systematic review. Sports Med. 2017, 47, 1569–1588. [Google Scholar] [CrossRef]
- Xu, R.; Zhang, C.; He, F.; Zhao, X.; Qi, H.; Zhou, P.; Zhang, L.; Ming, D. How physical activities affect mental fatigue based on EEG energy, connectivity, and complexity. Front. Neurol. 2018, 9, 915. [Google Scholar] [CrossRef] [PubMed]
- Jose, V.Ñ.; Mari-Carmen, G.C.; Ana, L.; Rafael, M.; Juan, B.M.; Federico, V.P.; Juan, S. Free Radicals in Exhaustive Physical Exercise: Mechanism of Production, and Protection by Antioxidants. IUBMB Life 2000, 50, 271–277. [Google Scholar]
- Reid, M.B. Free radicals and muscle fatigue: Of ROS, canaries, and the IOC. Free Radic. Biol. Med. 2008, 44, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.S.; Oh, H.; Rhee, S.G.; Yoo, Y.D. Regulation of reactive oxygen species generation in cell signaling. Mol. Cells 2011, 32, 491–509. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Young, I.S.; Woodside, J.V. Antioxidants in health and disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef]
- Ziad, M.; Zaher, M.A.J.; Saleh, A.A. Chapter 6. Nonenzymatic exogenous and endogenous antioxidants. In Free Radical Medicine and Biology; IntechOpen Limited: London, UK, 2019. [Google Scholar]
- Bouayed, J.; Bohn, T. Exogenous antioxidants—Double-edged swords in cellular redox state: Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid. Med. Cell. Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.F.; Reid, M.B. Muscle-derived ROS and thiol regulation in muscle fatigue. J. Appl. Physiol. 2008, 104, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.J.; O’Farrell, S. Free radicals and muscle damage. Br. Med. Bull. 1993, 49, 630–641. [Google Scholar] [CrossRef] [PubMed]
- Lian, D.; Chen, M.M.; Wu, H.; Deng, S.; Hu, X. The role of oxidative stress in skeletal muscle myogenesis and muscle disease. Antioxidants 2022, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Docherty, S.; Harley, R.; McAuley, J.J.; Crowe, L.A.N.; Pedret, C.; Kirwan, P.D.; Siebert, S.; Millar, N.L. The effect of exercise on cytokines: Implications for musculoskeletal health: A narrative review. BMC Sports Sci. Med. Rehabil. 2022, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.L. Cytokine hypothesis of overtraining: A physiological adaptation to excessive stress? Med. Sci. Sports Exer. 2000, 32, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Camus, G.; Deby-Dupont, G.; Duchateau, J.; Deby, C.; Pincemail, J.; Lamy, M. Are similar inflammatory factors involved in strenuous exercise and sepsis? Intensive Care Med. 1994, 20, 602–610. [Google Scholar] [CrossRef]
- Karshikoff, B.; Sundelin, T.; Lasselin, J. Role of inflammation in human fatigue: Relevance of multidimensional assessments and potential neuronal mechanisms. Front. Immunol. 2017, 8, 229498. [Google Scholar] [CrossRef]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of oxidative stress in the pathogenesis of non-alcoholic fatty liver disease: Implications for prevention and therapy. Antioxidants 2021, 10, 174. [Google Scholar] [CrossRef]
- Gerber, L.H.; Weinstein, A.A.; Mehta, R.; Younossi, Z.M. Importance of fatigue and its measurement in chronic liver disease. World J. Gastroenterol. 2019, 25, 3669–3683. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, Z.J.; Lao, L.; Wong, C.W.; Feng, Y. The role of oxidative stress and antioxidants in liver diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Pravin, S.K.; Durgacharan, A.B.; Dinesh, M.S. Deer antlers- Traditional use and future perspectives. IJTK 2010, 9, 245–251. [Google Scholar]
- Gordon, S.; Val, B.; Craig, P.; Alan, W.; David, G.; Stephen, H.; Roger, L. The effects of deer antler velvet extract or powder supplementation on aerobic power, erythropoiesis, and muscular strength and endurance characteristics. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 251–265. [Google Scholar]
- Shao, M.J.; Wang, S.R.; Zhao, M.J.; Lv, X.L.; Xu, H.; Li, L.; Gu, H.; Zhang, J.L.; Li, G.; Cui, X.N.; et al. The Effects of Velvet Antler of Deer on Cardiac Functions of Rats with Heart Failure following Myocardial Infarction. eCAM 2012, 2012, 825056. [Google Scholar] [PubMed]
- Louis, C.; Tomás, L.C.; Gemma, S.H.; Martina, P.S.; Francisco, J.P.B.; Carlos, G.A.; Andrés, G.; Carlos, C.; Jose, M.L.; Chunyi, L.; et al. Anti-tumour activity of deer growing antlers and its potential applications in the treatment of malignant gliomas. Sci. Rep. 2021, 11, 42. [Google Scholar]
- Jung, S.; Kim, S.; Jeung, W.; Ra, J.; Heo, K.; Shim, J.; Lee, J. Fermented antler improves endurance during exercise performance by increasing mitochondrial biogenesis and muscle strength in mice. Appl. Sci. 2021, 11, 5386. [Google Scholar] [CrossRef]
- Kim, Y.-T.; Jeon, H.; Kim, S.; Heo, K.; Shim, J.; Lee, J.; Yang, D.; Kang, S.C. Fermented antler recovers stamina, muscle strength and muscle mass in middle-aged mice. Appl. Sci. 2022, 12, 106. [Google Scholar] [CrossRef]
- Jeon, H.; Kim, Y.; Jang, W.Y.; Kim, J.; Heo, K.; Shim, J.; Lee, J.; Yang, D.; Kang, S.C. Effects of Lactobacillus curvatus HY7602-fermented antlers in dexamethasone-induced muscle atrophy. Fermentation 2022, 8, 454. [Google Scholar] [CrossRef]
- Jeon, H.; Lee, K.; Kim, J.; Shim, J.; Lee, J. Effect of Lactobacillus curvatus HY7602-fermented antler on sarcopenia in mice. Fermentation 2023, 9, 429. [Google Scholar] [CrossRef]
- Li, J.; Yang, Q.; Han, L.; Pan, C.; Lei, C.; Chen, H.; Lan, X. C2C12 Mouse Myoblasts Damage Induced by Oxidative Stress Is Alleviated by the Antioxidant Capacity of the Active Substance Phloretin. Front. Cell Dev. Biol. 2020, 8, 541260. [Google Scholar] [CrossRef]
- Mersch-Sundermann, V.; Knasmüller, S.; Wu, X.J.; Darroudi, F.; Kassie, F. Use of a human-derived liver cell line for the detection of cytoprotective, antigenotoxic and cogenotoxic agents. Toxicology 2004, 198, 329–340. [Google Scholar] [CrossRef]
- Ghoneum, M.; Abdulmalek, S.; Pan, D. Reversal of age-associated oxidative stress in mice by PFT, a novel kefir product. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420950149. [Google Scholar] [CrossRef]
- Hamilton, R.T.; Walsh, M.E.; Van, R.H. Mouse Models of Oxidative Stress Indicate a Role for Modulating Healthy Aging. J. Clin. Exp. Pathol. 2012, 4, 5. [Google Scholar] [CrossRef]
- Rosenthal, T.C.; Majeroni, B.A.; Pretorius, R.; Malik, K. Fatigue: An overview. Am. Fam. Phys. 2008, 78, 1173–1179. [Google Scholar]
- Cornuz, J.; Guessous, I.; Favrat, B. Fatigue: A practical approach to diagnosis in primary care. CMAJ 2006, 174, 765–767. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chu, Z.; Luo, Y.; Yang, F.; Cao, F.; Luo, F.; Lin, Q. Dietary polysaccharides exert anti-fatigue functions via the gut-muscle axis: Advances and prospectives. Foods 2023, 12, 3083. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lu, X.; Chen, P.; Shen, Y.; Zheng, B.; Guo, Z. Anti-fatigue effect of glycoprotein from hairtail (Trichiurus lepturus) by-products in a behavioral mouse model. Food Chem. X 2023, 18, 100645. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, X.; Liu, Y.; Zhang, Z. Effects of exercise-induced ROS on the pathophysiological functions of skeletal muscle. Oxid. Med. Cell. Longev. 2021, 2021, 3846122. [Google Scholar] [CrossRef]
- Nemes, R.; Koltai, E.; Taylor, A.W.; Suzuki, K.; Gyori, F.; Radak, Z. Reactive oxygen and nitrogen species regulate key metabolic, anabolic, and catabolic pathways in skeletal muscle. Antioxidants 2018, 7, 85. [Google Scholar] [CrossRef]
- Sharma, P.; Nandave, M.; Nandave, D.; Yadav, S.; Vargas-De-La-Cruz, C.; Singh, S.; Tandon, R.; Ramniwas, S.; Behl, T. Reactive oxygen species (ROS)-mediated oxidative stress in chronic liver diseases and its mitigation by medicinal plants. Am. J. Transl. Res. 2023, 15, 6321–6341. [Google Scholar]
- Swain, M.G.; Jones, D.E.J. Fatigue in chronic liver disease: New insights and therapeutic approaches. Liver Int. 2019, 39, 6–19. [Google Scholar] [CrossRef]
- Sjogren, M.H.; Mph, E. MonitoringTransaminase levels and vigorous exercise. Gastroenterol. Hepatol. 2007, 3, 913–914. [Google Scholar]
- Khatri, P.; Neupane, A.; Sapkota, S.R.; Bashyal, B.; Sharma, D.; Chhetri, A.; Chirag, K.C.; Banjade, A.; Sapkota, P.; Bhandari, S. Strenuous exercise-induced tremendously elevated transaminases levels in a healthy adult: A diagnostic Dilemma. Case Rep. Hepatol. 2021, 2021, 6653266. [Google Scholar] [CrossRef]
- Callegari, G.A.; Novaes, J.S.; Neto, G.R.; Dias, I.; Garrido, N.D.; Dani, C. Creatine kinase and lactate dehydrogenase responses after different resistance and aerobic exercise protocols. J. Hum. Kinet. 2017, 58, 65–72. [Google Scholar] [CrossRef]
- Wan, J.J.; Qin, Z.; Wang, P.Y.; Sun, Y.; Liu, X. Muscle fatigue: General understanding and treatment. Exp. Mol. Med. 2017, 49, e384. [Google Scholar] [CrossRef] [PubMed]
- Ament, W.; Verkerke, G.J. Exercise and fatigue. Sports Med. 2009, 39, 389–422. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.E.; Huang, W.C.; Liao, C.C.; Chang, Y.K.; Kan, N.W.; Huang, C.C. Resveratrol Protects against Physical Fatigue and Improves Exercise Performance in Mice. Molecules 2013, 18, 4689–4702. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B. Teaching the basics of redox biology to medical and graduate students: Oxidants, antioxidants and disease mechanisms. Redox Biol. 2013, 1, 244–257. [Google Scholar] [CrossRef]
- Agnieszka, M.C.; Anna, S.S.; Gerard, D. Antioxidant defense in children with chronic viral hepatitis B and C. Med. Sci. Monit. 2000, 6, 713–718. [Google Scholar]
- Michele, T.; Silvia, B.; Francesco, D.R.; Saro, O.; Gian, B.S.; Sergio, T.; Franco, Z. Glutathione and detoxification. Cancer Treat. Rev. 1990, 17, 203–208. [Google Scholar]
- Elaine, M.C.; Matthew, B.G. Inflammation, free radicals, and antioxidants. Nutrition 1996, 12, 274–277. [Google Scholar]
- Ismail, N.A.; Okasha, S.H.; Dhawan, A.; Abdel-Rahman, A.O.; Shaker, O.G.; Sadik, N.A. Antioxidant enzyme activities in hepatic tissue from children with chronic cholestatic liver disease. Saudi J. Gastroenterol. 2010, 16, 90–94. [Google Scholar] [CrossRef]
- Baumert, P.; Temple, S.; Stanley, J.M.; Cocks, M.; Strauss, J.A.; Shepherd, S.O.; Drust, B.; Lake, M.J.; Stewart, C.E.; Erskine, R.M. Neuromuscular fatigue and recovery after strenuous exercise depends on skeletal muscle size and stem cell characteristics. Sci. Rep. 2021, 11, 7733. [Google Scholar] [CrossRef]
- Norheim, K.B.; Jonsson, G.; Omdal, R. Biological mechanisms of chronic fatigue. Rheumatology 2011, 50, 1009–1018. [Google Scholar] [CrossRef]
- Cohen, T.V.; Many, G.M.; Fleming, B.D.; Gnocchi, V.F.; Ghimbovschi, S.; Mosser, D.M.; Hoffman, E.P.; Partridge, T.A. Upregulated IL-1β in dysferlin-deficient muscle attenuates regeneration by blunting the response to pro-inflammatory macrophages. Skelet. Muscle 2015, 5, 24. [Google Scholar] [CrossRef]
- VanderVeen, B.N.; Fix, D.K.; Montalvo, R.N.; Counts, B.R.; Smuder, A.J.; Murphy, E.A.; Koh, H.J.; Carson, J.A. The regulation of skeletal muscle fatigability and mitochondrial function by chronically elevated interleukin-6. Exp. Physiol. 2019, 104, 385–397. [Google Scholar] [CrossRef]
- Eruslanov, E.; Kusmartsev, S. Identification of ROS Using Oxidized DCFDA and flow-cytometry. In Advanced Protocols in Oxidative Stress II; Armstrong, D., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 57–72. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, H.; Lee, K.; Kim, Y.-T.; Kim, J.-Y.; Shim, J.-J.; Lee, J.-H. Effect of HY7602 Fermented Deer Antler on Physical Fatigue and Antioxidant Activity in Mice. Int. J. Mol. Sci. 2024, 25, 3318. https://doi.org/10.3390/ijms25063318
Jeon H, Lee K, Kim Y-T, Kim J-Y, Shim J-J, Lee J-H. Effect of HY7602 Fermented Deer Antler on Physical Fatigue and Antioxidant Activity in Mice. International Journal of Molecular Sciences. 2024; 25(6):3318. https://doi.org/10.3390/ijms25063318
Chicago/Turabian StyleJeon, Hyejin, Kippeum Lee, Yong-Tae Kim, Joo-Yun Kim, Jae-Jung Shim, and Jae-Hwan Lee. 2024. "Effect of HY7602 Fermented Deer Antler on Physical Fatigue and Antioxidant Activity in Mice" International Journal of Molecular Sciences 25, no. 6: 3318. https://doi.org/10.3390/ijms25063318