
Benzothiadiazinone-1,1-Dioxide Carbonic Anhydrase Inhibitors Suppress the Growth of Drug-Resistant Mycobacterium tuberculosis Strains
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
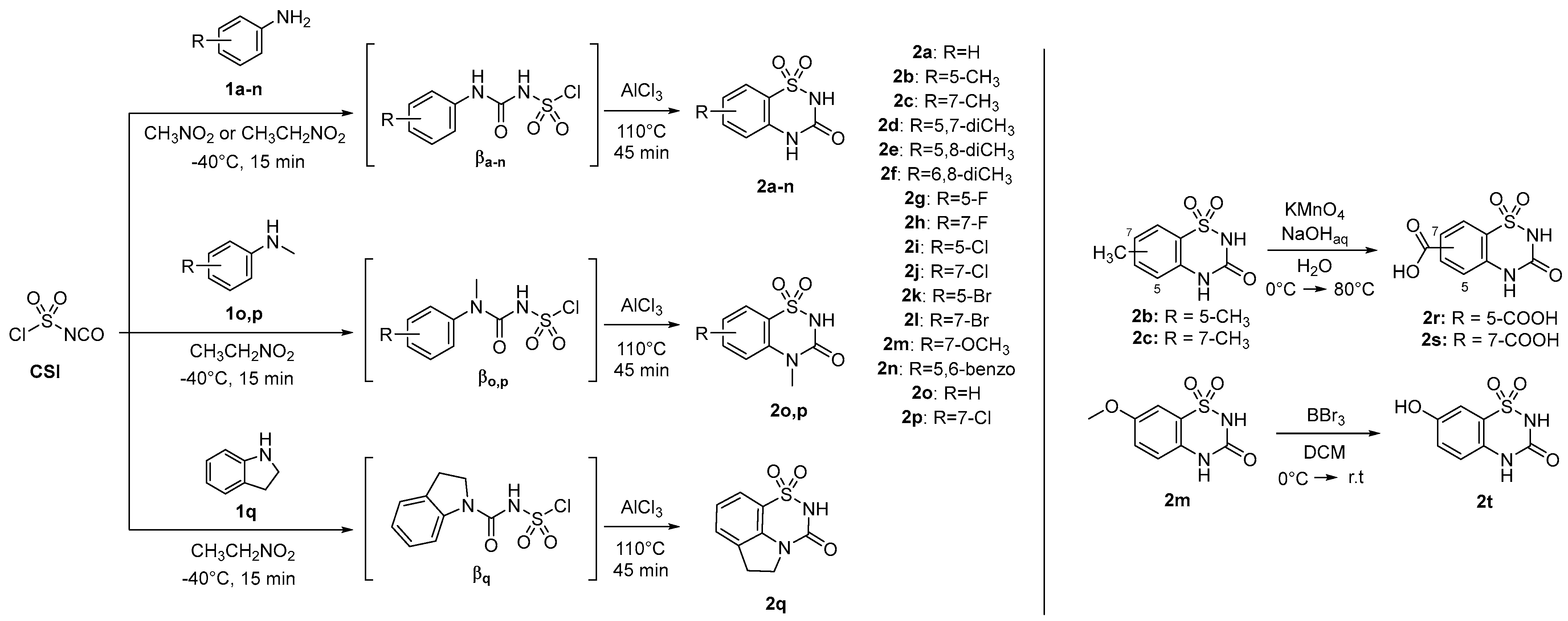
2.1. Design and Chemistry
2.2. Carbonic Anhydrase Inhibition Assay
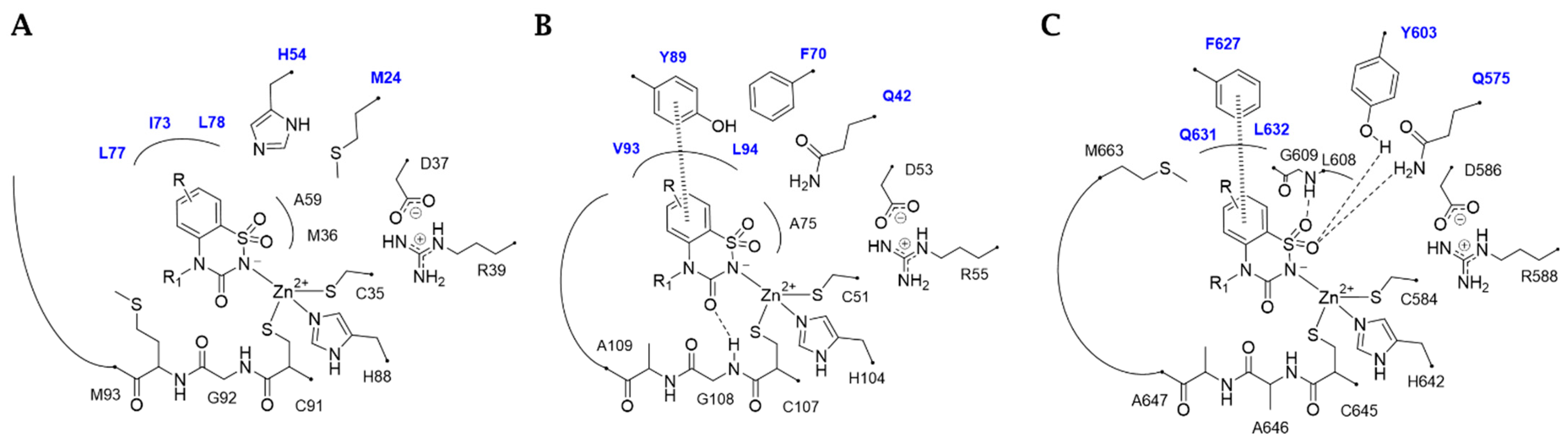
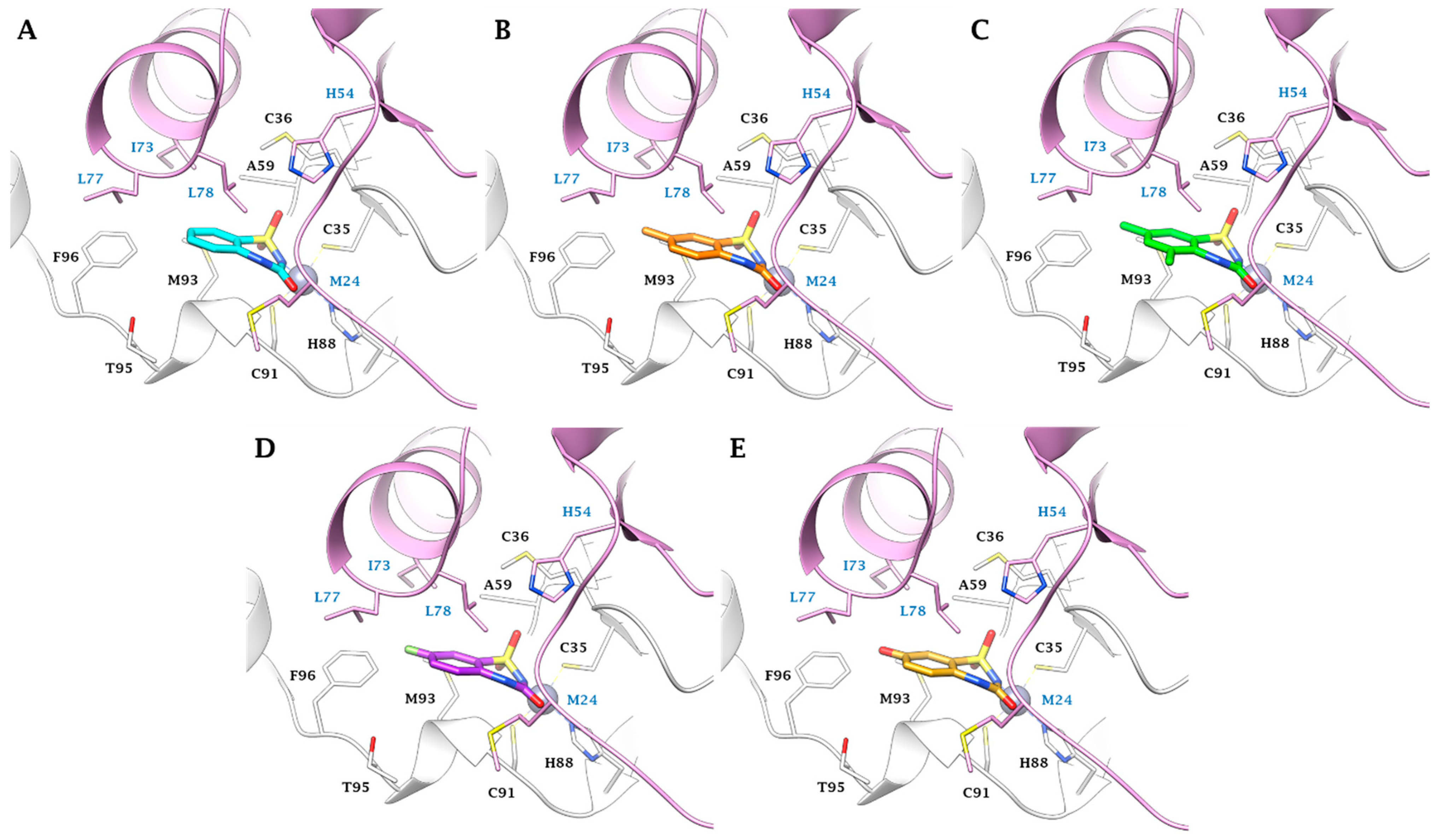
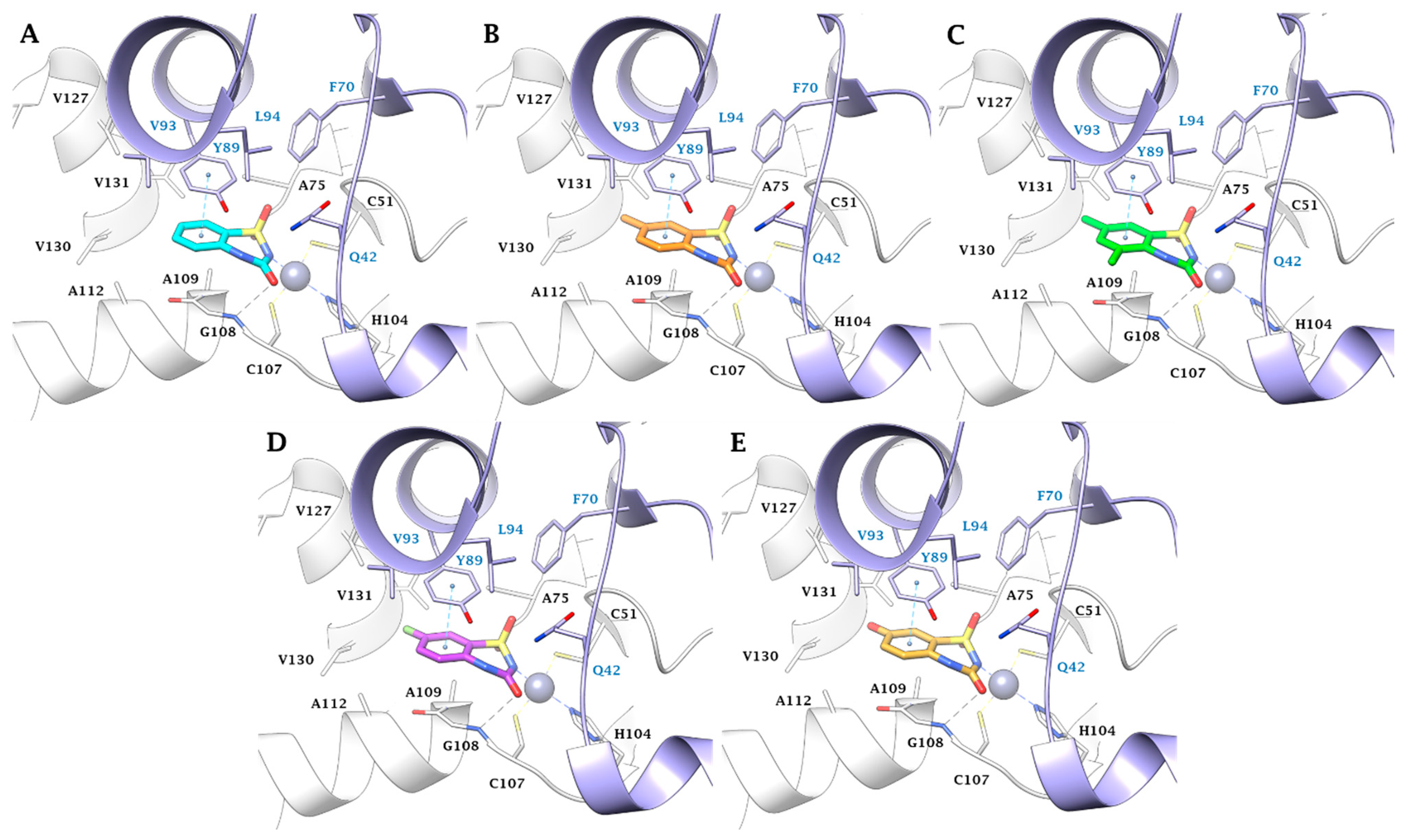
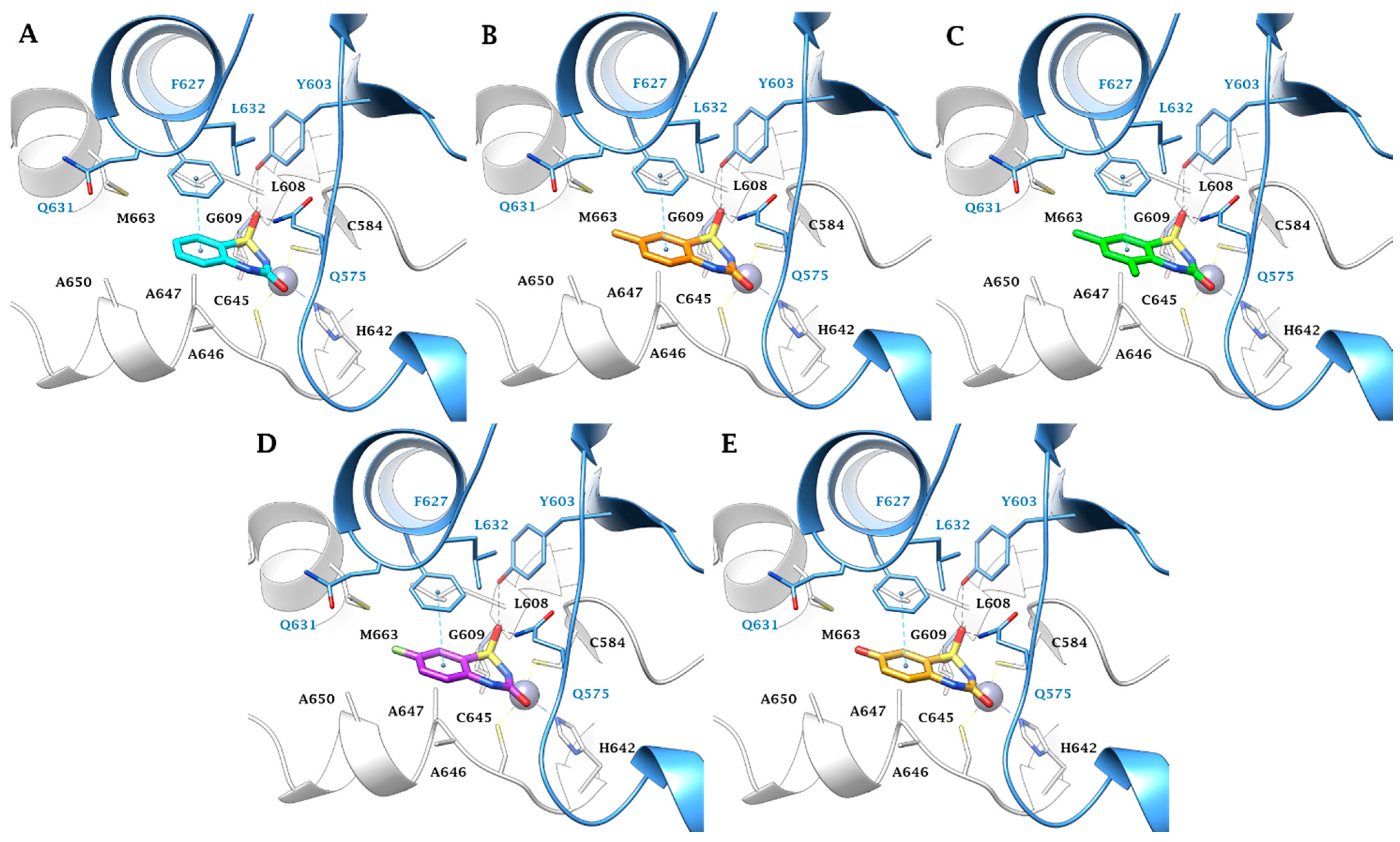
2.3. In Silico Study
2.4. Tuberculostatic Activity Assay
3. Material and Methods
3.1. Chemistry
3.2. Carbonic Anhydrase CO2 Hydration Catalytic/Inhibition Assay and Ki Determination
3.3. In Silico Studies
3.4. Tuberculostatic Activity Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2023; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2023. Available online: https://www.who.int/publications/i/item/9789240083851 (accessed on 12 December 2023).
- Alsayed, S.S.R.; Gunosewoyo, H. Tuberculosis: Pathogenesis, Current Treatment Regimens and New Drug Targets. Int. J. Mol. Sci. 2023, 24, 5202. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Chan, J. Tuberculosis: Latency and Reactivation. Infect. Immun. 2001, 69, 4195–4201. [Google Scholar] [CrossRef]
- Pai, M.; Behr, M.A.; Dowdy, D.; Dheda, K.; Divangahi, M.; Boehme, C.C.; Ginsberg, A.; Swaminathan, S.; Spigelman, M.; Getahun, H.; et al. Tuberculosis. Nat. Rev. Dis. Primers 2016, 27, 16076. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Grigsby, S.J.; Philips, J.A. Immune evasion and provocation by Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2022, 20, 750–766. [Google Scholar] [CrossRef] [PubMed]
- Philips, J.A.; Ernst, J.D. Tuberculosis Pathogenesis and Immunity. Annu. Rev. Pathol. Mech. Dis. 2012, 7, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.; Acharya, A.; Gautam, S.; Ghimire, S.P.; Mishra, G.; Parajuli, N.; Sapkota, B. Advances in diagnosis of Tuberculosis: An update into molecular diagnosis of Mycobacterium tuberculosis. Mol. Biol. Rep. 2020, 47, 4065–4075. [Google Scholar] [CrossRef]
- McMahon, P.; Pisapia, D.J.; Schweitzer, A.D.; Heier, L.; Souweidane, M.M.; Roytman, M. Central nervous system tuberculoma mimicking a brain tumor: A case report. Radiol. Case Rep. 2023, 19, 414–417. [Google Scholar] [CrossRef]
- Daher Ede, F.; da Silva, G.B., Jr.; Barros, E.J. Renal tuberculosis in the modern era. Am. J. Trop. Med. Hyg. 2013, 88, 54–64. [Google Scholar] [CrossRef]
- Shaikh, G.A.; Yaqoob, S.; Batool, F.; Waseem, R.; Shah, H.H.; Abbasi, A.A.; Lashari, N.; Dave, T. A rare case of tuberculous myocarditis: A diagnostic challenge in a tuberculosis-endemic country. Clin. Case Rep. 2023, 11, e8224. [Google Scholar] [CrossRef]
- Chen, B.; Bao, Y.; Chen, J.; Zhang, Y.; Wen, Q.; Wang, K.; Cheng, X.; Lv, J. Isolated soft tissue tuberculosis: A case report and literature review. Front. Med. 2023, 10, 1205446. [Google Scholar] [CrossRef] [PubMed]
- Baidya, A.; Singha, A.; Bhattacharjee, R.; Dalal, B.S. Tuberculosis of the thyroid gland: Two case reports. Oxf. Med. Case Rep. 2015, 2015, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Ganchua, S.K.C.; White, A.G.; Klein, E.C.; Flynn, J.L. Lymph nodes-The neglected battlefield in tuberculosis. PLoS Pathog. 2020, 16, e1008632. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, O.H.; Sunil, J.; Prakash, S.; Vijaykumar, C.; Kumbhar, U.S. Isolated pancreatic tuberculosis masquerading pancreatic malignancy. BMJ Case Rep. 2023, 16, e254250. [Google Scholar] [CrossRef] [PubMed]
- Salih, A.M.; Qaradakhy, A.J.; Hassan, S.H.; Abdullah, A.M.; Dhahir, H.M.; Karim, S.O.; Sofi, H.A.; Abdalla, B.A.; Ali, M.H.; Kakamad, F.H. Tuberculous granulomatous inflammation of parathyroid adenoma manifested as primary hyperparathyroidism: A case report and a review of the literature. Med. Int. 2023, 3, 49. [Google Scholar] [CrossRef]
- Gadde, A.B.; Jha, P.K.; Bansal, S.B.; Rana, A.; Jain, M.; Bansal, D.; Yadav, D.K.; Mahapatra, A.K.; Sethi, S.K.; Kher, V. Renal Transplantation in Patients with Tuberculosis: A Single-center Experience from an Endemic Region. Transplant. Direct 2023, 9, e1541. [Google Scholar] [CrossRef]
- Malihy, Z.; Benaissa, E.; Ben Lahlou, Y.; Maleb, A.; Elouennass, M. Osteoarticular tuberculosis of the ankle, a rare localization: A case report. Access Microbiol. 2023, 5, 000654.v3. [Google Scholar] [CrossRef]
- Tsui, J.K.; Poon, S.H.L.; Fung, N.S.K. Ocular manifestations and diagnosis of tuberculosis involving the uvea: A case series. Trop. Dis. Travel Med. Vaccines 2023, 9, 20. [Google Scholar] [CrossRef]
- Singh, A.; Singh, B.K.; Singh, V.K.; Nagarajan, S. Presumptive ocular tuberculosis: A case series of rare cases. Oman J. Ophthalmol. 2023, 16, 529–532. [Google Scholar]
- Pottakkat, B.; Kumar, A.; Rastogi, A.; Krishnani, N.; Kapoor, V.K.; Saxena, R. Tuberculosis of the spleen as a cause of Fever of unknown origin and splenomegaly. Gut Liver 2010, 4, 94–97. [Google Scholar] [CrossRef]
- Rehman, A.U.; Khattak, M.; Mushtaq, U.; Latif, M.; Ahmad, I.; Rasool, M.F.; Shakeel, S.; Hayat, K.; Hussain, R.; Alhazmi, G.A.; et al. The impact of diabetes mellitus on the emergence of multi-drug resistant tuberculosis and treatment failure in TB-diabetes comorbid patients: A systematic review and meta-analysis. Front. Public Health 2023, 11, 1244450. [Google Scholar] [CrossRef]
- Mbewana Ntshanka, N.G.; Msagati, T.A.M. Trends and Progress on Antibiotic-Resistant Mycobacterium tuberculosis and Genes in relation to Human Immunodeficiency Virus. Can. J. Infect. Dis. Med. Microbiol. 2023, 2023, 6659212. [Google Scholar] [CrossRef]
- Lall, D. Tuberculosis and undernutrition in India. Lancet Glob. Health 2023, 11, e1859. [Google Scholar] [CrossRef]
- Wagnew, F.; Alene, K.A.; Kelly, M.; Gray, D. Geospatial Overlap of Undernutrition and Tuberculosis in Ethiopia. Int. J. Environ. Res. Public Health 2023, 20, 7000. [Google Scholar] [CrossRef]
- Corleis, B.; Tzouanas, C.N.; Wadsworth, M.H., 2nd; Cho, J.L.; Linder, A.H.; Schiff, A.E.; Zessin, B.; Stei, F.; Dorhoi, A.; Dickey, A.K.; et al. Tobacco smoke exposure recruits inflammatory airspace monocytes that establish permissive lung niches for Mycobacterium tuberculosis. Sci. Transl. Med. 2023, 15, eadg3451. [Google Scholar] [CrossRef] [PubMed]
- Boisson-Walsh, A.; Thompson, P.; Fried, B.; Shea, C.M.; Ngimbi, P.; Lumande, F.; Tabala, M.; Kashamuka, M.M.; Babakazo, P.; Domino, M.E.; et al. Childhood immunization uptake determinants in Kinshasa, Democratic Republic of the Congo: Ordered regressions to assess timely infant vaccines administered at birth and 6-weeks. Glob. Health Res. Policy 2023, 8, 50. [Google Scholar] [CrossRef]
- Cotton, M.F.; Rabie, H. Planning to introduce novel tuberculosis vaccines in high burden settings: How could this be done? Lancet Glob. Health 2023, 11, e484–e485. [Google Scholar] [CrossRef]
- Tait, D.R.; Hatherill, M.; Van Der Meeren, O.; Ginsberg, A.M.; Van Brakel, E.; Salaun, B.; Scriba, T.J.; Akite, E.J.; Ayles, H.M.; Bollaerts, A.; et al. Final Analysis of a Trial of M72/AS01E Vaccine to Prevent Tuberculosis. N. Engl. J. Med. 2019, 381, 2429–2439. [Google Scholar] [CrossRef]
- Blossey, A.M.; Brückner, S.; May, M.; Parzmair, G.P.; Sharma, H.; Shaligram, U.; Grode, L.; Kaufmann, S.H.E.; Netea, M.G.; Schindler, C. VPM1002 as Prophylaxis Against Severe Respiratory Tract Infections Including Coronavirus Disease 2019 in the Elderly: A Phase 3 Randomized, Double-Blind, Placebo-Controlled, Multicenter Clinical Study. Clin. Infect. Dis. 2023, 76, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Tkachuk, A.P.; Bykonia, E.N.; Popova, L.I.; Kleymenov, D.A.; Semashko, M.A.; Chulanov, V.P.; Fitilev, S.B.; Maksimov, S.L.; Smolyarchuk, E.A.; Manuylov, V.A.; et al. Safety and Immunogenicity of the GamTBvac, the Recombinant Subunit Tuberculosis Vaccine Candidate: A Phase II, Multi-Center, Double-Blind, Randomized, Placebo-Controlled Study. Vaccines 2020, 8, 652. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Marco-García, M.; Gamell, A.; Cusó, T.; Monsonís, M.; Latre, C.; Fortuny, C.; Noguera-Julian, A. Toxicity of the Increased Recommended Doses of First-line Anti-tuberculosis Oral Drugs in Children in a Reference Center in Spain. Arch. Bronconeumol. 2023, 59, 612–615. [Google Scholar] [CrossRef] [PubMed]
- Ndambuki, J.; Nzomo, J.; Muregi, L.; Mutuku, C.; Makokha, F.; Nthusi, J.; Ambale, C.; Lynen, L.; Decroo, T. Comparison of first-line tuberculosis treatment outcomes between previously treated and new patients: A retrospective study in Machakos subcounty, Kenya. Int. Health 2021, 13, 272–280. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Consolidated Guidelines on Tuberculosis. Module 4: Treatment—Drug-Resistant Tuberculosis Treatment, 2022 Update; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2022. Available online: https://www.who.int/publications/i/item/9789240063129 (accessed on 12 December 2023).
- Tiberi, S.; Utjesanovic, N.; Galvin, J.; Centis, R.; D’Ambrosio, L.; van den Boom, M.; Zumla, A.; Migliori, G.B. Drug resistant TB—Latest developments in epidemiology, diagnostics and management. Int. J. Infect. Dis. 2022, 124, S20–S25. [Google Scholar] [CrossRef] [PubMed]
- Seung, K.J.; Keshavjee, S.; Rich, M.L. Multidrug-Resistant Tuberculosis and Extensively Drug-Resistant Tuberculosis. Cold Spring Harb. Perspect. Med. 2015, 5, a017863. [Google Scholar] [CrossRef] [PubMed]
- Akkahadsee, P.; Sawangjit, R.; Phumart, P.; Chaiyakunapruk, N.; Sakloetsakun, D. Systematic review and network meta-analysis of efficacy and safety of interventions for preventing anti-tuberculosis drug induced liver injury. Sci. Rep. 2023, 13, 19880. [Google Scholar] [CrossRef] [PubMed]
- Molla, Y.; Wubetu, M.; Dessie, B. Anti-Tuberculosis Drug Induced Hepatotoxicity and Associated Factors among Tuberculosis Patients at Selected Hospitals, Ethiopia. Hepatic Med. 2021, 13, 1–8. [Google Scholar] [CrossRef]
- Sekaggya-Wiltshire, C.; von Braun, A.; Scherrer, A.U.; Manabe, Y.C.; Buzibye, A.; Muller, D.; Ledergerber, B.; Gutteck, U.; Corti, N.; Kambugu, A.; et al. Anti-TB drug concentrations and drug-associated toxicities among TB/HIV-coinfected patients. J. Antimicrob. Chemother. 2017, 72, 1172–1177. [Google Scholar] [CrossRef]
- World Health Organization. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report: 2022; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2022. Available online: https://www.who.int/publications/i/item/9789240062702 (accessed on 12 December 2023).
- Mishra, C.B.; Tiwari, M.; Supuran, C.T. Progress in the development of human carbonic anhydrase inhibitors and their pharmacological applications: Where are we today? Med. Res. Rev. 2020, 40, 2485–2565. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. Antibacterial carbonic anhydrase inhibitors: An update on the recent literature. Expert Opin. Ther. Pat. 2020, 30, 963–982. [Google Scholar] [CrossRef]
- Kaur, J.; Cao, X.; Abutaleb, N.S.; Elkashif, A.; Graboski, A.L.; Krabill, A.D.; AbdelKhalek, A.H.; An, W.; Bhardwaj, A.; Seleem, M.N.; et al. Optimization of Acetazolamide-Based Scaffold as Potent Inhibitors of Vancomycin-Resistant Enterococcus. J. Med. Chem. 2020, 63, 9540–9562. [Google Scholar] [CrossRef]
- Abutaleb, N.S.; Elhassanny, A.E.M.; Flaherty, D.P.; Seleem, M.N. In vitro and in vivo activities of the carbonic anhydrase inhibitor, dorzolamide, against vancomycin-resistant enterococci. PeerJ 2021, 9, e11059. [Google Scholar] [CrossRef]
- Abutaleb, N.S.; Elhassanny, A.E.M.; Seleem, M.N. In vivo efficacy of acetazolamide in a mouse model of Neisseria gonorrhoeae infection. Microb. Pathog. 2022, 164, 105454. [Google Scholar] [CrossRef] [PubMed]
- Abuaita, B.H.; Withey, J.H. Bicarbonate Induces Vibrio cholerae virulence gene expression by enhancing ToxT activity. Infect. Immun. 2009, 77, 4111–4120. [Google Scholar] [CrossRef]
- Aspatwar, A.; Kairys, V.; Rala, S.; Parikka, M.; Bozdag, M.; Carta, F.; Supuran, C.T.; Parkkila, S. Mycobacterium tuberculosis β-Carbonic Anhydrases: Novel Targets for Developing Antituberculosis Drugs. Int. J. Mol. Sci. 2019, 20, 5153. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Powderly, W.G. The post-antibiotic era is here. Science 2021, 373, 471. [Google Scholar] [CrossRef] [PubMed]
- Cima, G. WHO warns of ‘post-antibiotic era’. J. Am. Vet. Med. Assoc. 2014, 244, 1356–1357. [Google Scholar]
- Zucca, M.; Savoia, D. The post-antibiotic era: Promising developments in the therapy of infectious diseases. Int. J. Biomed. Sci. 2010, 6, 77–86. [Google Scholar]
- Nocentini, A.; Cadoni, R.; Del Prete, S.; Capasso, C.; Dumy, P.; Gratteri, P.; Supuran, C.T.; Winum, J.Y. Benzoxaboroles as Efficient Inhibitors of the β-Carbonic Anhydrases from Pathogenic Fungi: Activity and Modeling Study. ACS Med. Chem. Lett. 2017, 8, 1194–1198. [Google Scholar] [CrossRef]
- Bonardi, A.; Nocentini, A.; Cadoni, R.; Del Prete, S.; Dumy, P.; Capasso, C.; Gratteri, P.; Supuran, C.T.; Winum, J.Y. Benzoxaboroles: New Potent Inhibitors of the Carbonic Anhydrases of the Pathogenic Bacterium Vibrio cholerae. ACS Med. Chem. Lett. 2020, 11, 2277–2284. [Google Scholar] [CrossRef]
- Güzel, O.; Maresca, A.; Scozzafava, A.; Salman, A.; Balaban, A.T.; Supuran, C.T. Discovery of low nanomolar and subnanomolar inhibitors of the mycobacterial beta-carbonic anhydrases Rv1284 and Rv3273. J. Med. Chem. 2009, 52, 4063–4067. [Google Scholar] [CrossRef]
- Aspatwar, A.; Hammarén, M.; Koskinen, S.; Luukinen, B.; Barker, H.; Carta, F.; Supuran, C.T.; Parikka, M.; Parkkila, S. beta-CA-specific inhibitor dithiocarbamate Fc14-584B: A novel antimycobacterial agent with potential to treat drug-resistant tuberculosis. J. Enzym. Inhib. Med. Chem. 2017, 32, 832–840. [Google Scholar] [CrossRef]
- Bua, S.; Lomelino, C.; Murray, A.B.; Osman, S.M.; ALOthman, Z.A.; Bozdag, M.; Abdel-Aziz, H.A.; Eldehna, W.M.; McKenna, R.; Nocentini, A.; et al. “A Sweet Combination”: Developing Saccharin and Acesulfame K Structures for Selectively Targeting the Tumor-Associated Carbonic Anhydrases IX and XII. J. Med. Chem. 2020, 63, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar] [CrossRef] [PubMed]
- Abdoli, M.; Bonardi, A.; Paoletti, N.; Aspatwar, A.; Parkkila, S.; Gratteri, P.; Supuran, C.T.; Žalubovskis, R. Inhibition Studies on Human and Mycobacterial Carbonic Anhydrases with N-((4-Sulfamoylphenyl)carbamothioyl) Amides. Molecules 2023, 28, 4020. [Google Scholar] [CrossRef] [PubMed]
- Ommi, O.; Paoletti, N.; Bonardi, A.; Gratteri, P.; Bhalerao, H.A.; Sau, S.; Nanduri, S.; Mohammed, A.; Kalia, N.P.; Sonti, R.; et al. Exploration of 3-aryl pyrazole-tethered sulfamoyl carboxamides as carbonic anhydrase inhibitors. Arch. Pharm. 2023, 356, e2300309. [Google Scholar] [CrossRef] [PubMed]
- McGurn, L.D.; Moazami-Goudarzi, M.; White, S.A.; Suwal, T.; Brar, B.; Tang, J.Q.; Espie, G.S.; Kimber, M.S. The structure, kinetics and interactions of the β-carboxysomal β-carbonic anhydrase, CcaA. Biochem. J. 2016, 473, 4559–4572. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger Suite Release 2022-4, Schrödinger, LLC, New York, NY, 2022: (a) Prime, v.5.5; (b) Maestro v.13.2; (c) Epik, v.6.0; (d) Impact, v.9.5; (e) Macromodel v.13.6. (f) Glide, v.9.5.
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, D.K.; Jude, K.M.; Banerjee, A.L.; Haldar, M.; Manokaran, S.; Kooren, J.; Mallik, S.; Christianson, D.W. Structural analysis of charge discrimination in the binding of inhibitors to human carbonic anhydrases I and II. J. Am. Chem. Soc. 2007, 129, 5528–5537. [Google Scholar] [CrossRef]
- Behnke, C.A.; Le Trong, I.; Godden, J.W.; Merritt, E.A.; Teller, D.C.; Bajorath, J.; Stenkamp, R.E. Atomic resolution studies of carbonic anhydrase II. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 616–627. [Google Scholar] [CrossRef]
- Suarez Covarrubias, A.; Larsson, A.M.; Högbom, M.; Lindberg, J.; Bergfors, T.; Björkelid, C.; Mowbray, S.L.; Unge, T.; Jones, T.A. Structure and function of carbonic anhydrases from Mycobacterium tuberculosis. J. Biol. Chem. 2005, 280, 18782–18789. [Google Scholar] [CrossRef]
- Covarrubias, A.S.; Bergfors, T.; Jones, T.A.; Högbom, M. Structural mechanics of the pH-dependent activity of beta-carbonic anhydrase from Mycobacterium tuberculosis. J. Biol. Chem. 2006, 281, 4993–4999. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Lu, C.; Wu, C.; Ghoreishi, D.; Chen, W.; Wang, L.; Damm, W.; Ross, G.A.; Dahlgren, M.K.; Russell, E.; Von Bargen, C.D.; et al. OPLS4: Improving Force Field Accuracy on Challenging Regimes of Chemical Space. J. Chem. Theory Comput. 2021, 17, 4291–4300. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, G.A.; Friesner, R.A.; Tirado-Rives, J.; Jorgensen, W.L. Evaluation and Reparametrization of the OPLS-AA Force Field for Proteins via Comparison with Accurate Quantum Chemical Calculations on Peptides. J. Phys. Chem. B 2001, 105, 6474–6487. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Homorodean, D.; Moisoiu, A.; Borroni, E. Ghid Național Pentru Rețeaua Laboratoarelor TB; Ministerul Sănătății: Bucharest, Romania, 2017. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||||
| Ki (nM) a | |||||||
| Cmpd | R | R1 | MtCA1 | MtCA2 | MtCA3 | hCA I b | hCA II b |
| 2a | H | H | 1680 ± 93 | 68.6 ± 3.1 | 28.7 ± 2.1 | 866 | 150 |
| 2b | 5-CH3 | H | 1460 ± 101 | 58.5 ± 1.7 | 21.3 ± 1.7 | 969 | 144 |
| 2c | 7-CH3 | H | 1050 ± 65 | 41.3 ± 2.0 | 18.5 ± 1.6 | 1070 | 872 |
| 2d | 5,7-diCH3 | H | 934 ± 78 | 38.1 ± 1.1 | 15.1 ± 0.9 | 1270 | 764 |
| 2e | 5,8-diCH3 | H | >10,000 | 2370 ± 167 | 1540 ± 91 | 2540 | 2190 |
| 2f | 6,8-diCH3 | H | >10,000 | 4480 ± 277 | 2250 ± 168 | 1920 | 2940 |
| 2g | 5-F | H | 1290 ± 96 | 62.9 ± 2.5 | 16.2 ± 1.1 | 547 | 140 |
| 2h | 7-F | H | 1830 ± 137 | 42.5 ± 1.6 | 24.3 ± 1.4 | 1290 | 534 |
| 2i | 5-Cl | H | 1500 ± 89 | 214 ± 13 | 44.3 ± 2.9 | 930 | 686 |
| 2j | 7-Cl | H | 2460 ± 174 | 67.1 ± 4.3 | 39.8 ± 2.2 | 1860 | 1140 |
| 2k | 5-Br | H | 993 ± 33 | 458 ± 25 | 82.4 ± 4.9 | 2320 | 527 |
| 2l | 7-Br | H | 2960 ± 213 | 200 ± 14 | 90.1 ± 6.7 | 4400 | 2360 |
| 2m | 7-OCH3 | H | 2760 ± 166 | 152 ± 11 | 59.2 ± 4.2 | 3810 | 1920 |
| 2n | 5,6-benzo | H | 1040 ± 41 | 995 ± 35 | 120 ± 8 | 2070 | 484 |
| 2o | H | CH3 | 556 ± 37 | 39.6 ± 2.1 | 32.5 ± 2.4 | 3410 | 853 |
| 2p | 7-Cl | CH3 | 1180 ± 75 | 135 ± 9 | 46.0 ± 3.3 | >10,000 | 2340 |
| 2q | 5-CH2-CH2- | 883 ± 24 | 599 ± 31 | 76.2 ± 5.6 | 6860 | 1060 | |
| 2r | 5-COOH | H | 3280 ± 204 | 105 ± 6 | 65.4 ± 3.9 | 5780 | 2450 |
| 2s | 7-COOH | H | 4870 ± 315 | 223 ± 18 | 101 ± 5 | >10,000 | 6780 |
| 2t | 7-OH | H | 2390 ± 126 | 58.4 ± 4.1 | 45.6 ± 3.5 | 1890 | 3020 |
| AAZ | - | 480 | 9.8 | 104 | 250 | 12.5 | |
 | |||||
| Selectivity Index (SI) | |||||
| Cmpd | R | R1 | hCA II/MtCA1 | hCA II/MtCA2 | hCA II/MtCA3 |
| 2a | H | H | 0.1 | 2.2 | 5.2 |
| 2b | 5-CH3 | H | 0.1 | 2.5 | 6.8 |
| 2c | 7-CH3 | H | 0.8 | 21.1 | 47.1 |
| 2d | 5,7-diCH3 | H | 0.8 | 20.0 | 50.6 |
| 2e | 5,8-diCH3 | H | <0.2 | 0.9 | 1.4 |
| 2f | 6,8-diCH3 | H | <0.3 | 0.7 | 1.3 |
| 2g | 5-F | H | 0.1 | 2.2 | 8.6 |
| 2h | 7-F | H | 0.3 | 12.6 | 22.0 |
| 2i | 5-Cl | H | 0.5 | 3.2 | 15.5 |
| 2j | 7-Cl | H | 0.5 | 17.0 | 28.7 |
| 2k | 5-Br | H | 0.5 | 1.1 | 6.4 |
| 2l | 7-Br | H | 0.8 | 11.8 | 26.2 |
| 2m | 7-OCH3 | H | 0.7 | 12.6 | 32.5 |
| 2n | 5,6-benzo | H | 0.5 | 0.5 | 4.0 |
| 2o | H | CH3 | 1.5 | 21.5 | 26.2 |
| 2p | 7-Cl | CH3 | 2.0 | 17.3 | 50.8 |
| 2q | 5-CH2-CH2- | 1.2 | 1.8 | 13.9 | |
| 2r | 5-COOH | H | 0.7 | 23.2 | 37.4 |
| 2s | 7-COOH | H | 1.4 | 30.4 | 67.4 |
| 2t | 7-OH | H | 1.3 | 51.8 | 66.3 |
| AAZ | - | <0.1 | 1.3 | 0.1 | |
| Strain Susceptible to Rifampicin and Isoniazid | Strain Resistant to Isoniazid | Strain Resistant to Both Rifampicin and Isoniazid | ||||
|---|---|---|---|---|---|---|
| Cmpd | 1 µg/mL | 2 µg/mL | 1 µg/mL | 2 µg/mL | 1 µg/mL | 2 µg/mL |
| 2a | 30 | 0 | 0 | 0 | 0 | 0 |
| 2c | 0 | 0 | 0 | 0 | 0 | 0 |
| 2d | 0 | 0 | 0 | 0 | 0 | 0 |
| 2h | 0 | 0 | 0 | 0 | 0 | 0 |
| 2t | 30–100 | 0 | 30–100 | 0 | 0 | 0 |
| MTZ | 30–100 | 0 | 0 | 0 | 0 | 0 |
| AAZ | 0 | 0 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bua, S.; Bonardi, A.; Mük, G.R.; Nocentini, A.; Gratteri, P.; Supuran, C.T. Benzothiadiazinone-1,1-Dioxide Carbonic Anhydrase Inhibitors Suppress the Growth of Drug-Resistant Mycobacterium tuberculosis Strains. Int. J. Mol. Sci. 2024, 25, 2584. https://doi.org/10.3390/ijms25052584
Bua S, Bonardi A, Mük GR, Nocentini A, Gratteri P, Supuran CT. Benzothiadiazinone-1,1-Dioxide Carbonic Anhydrase Inhibitors Suppress the Growth of Drug-Resistant Mycobacterium tuberculosis Strains. International Journal of Molecular Sciences. 2024; 25(5):2584. https://doi.org/10.3390/ijms25052584
Chicago/Turabian StyleBua, Silvia, Alessandro Bonardi, Georgiana Ramona Mük, Alessio Nocentini, Paola Gratteri, and Claudiu T. Supuran. 2024. "Benzothiadiazinone-1,1-Dioxide Carbonic Anhydrase Inhibitors Suppress the Growth of Drug-Resistant Mycobacterium tuberculosis Strains" International Journal of Molecular Sciences 25, no. 5: 2584. https://doi.org/10.3390/ijms25052584