
Genome-Wide Association Study and Identification of Candidate Genes Associated with Seed Number per Pod in Soybean
 ,
,
Abstract
:1. Introduction
2. Results
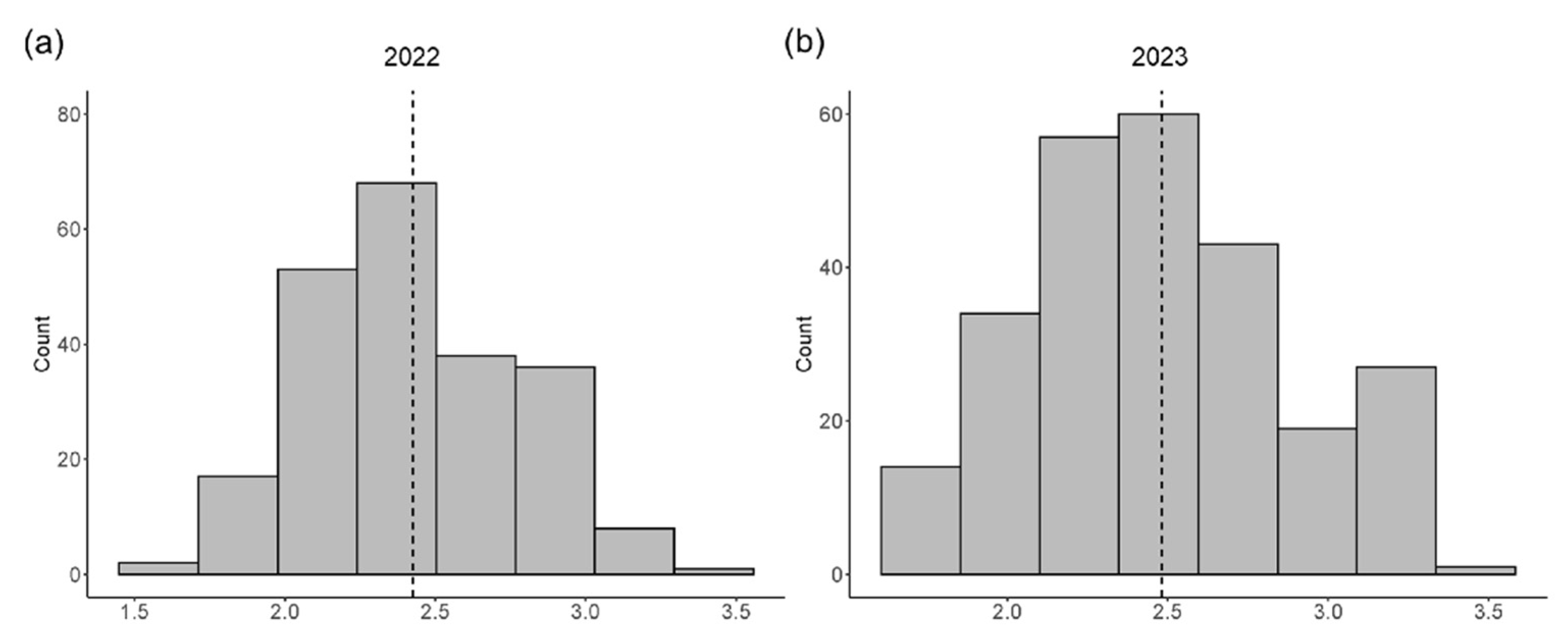
2.1. Phenotypic Variations of the SNPP Trait
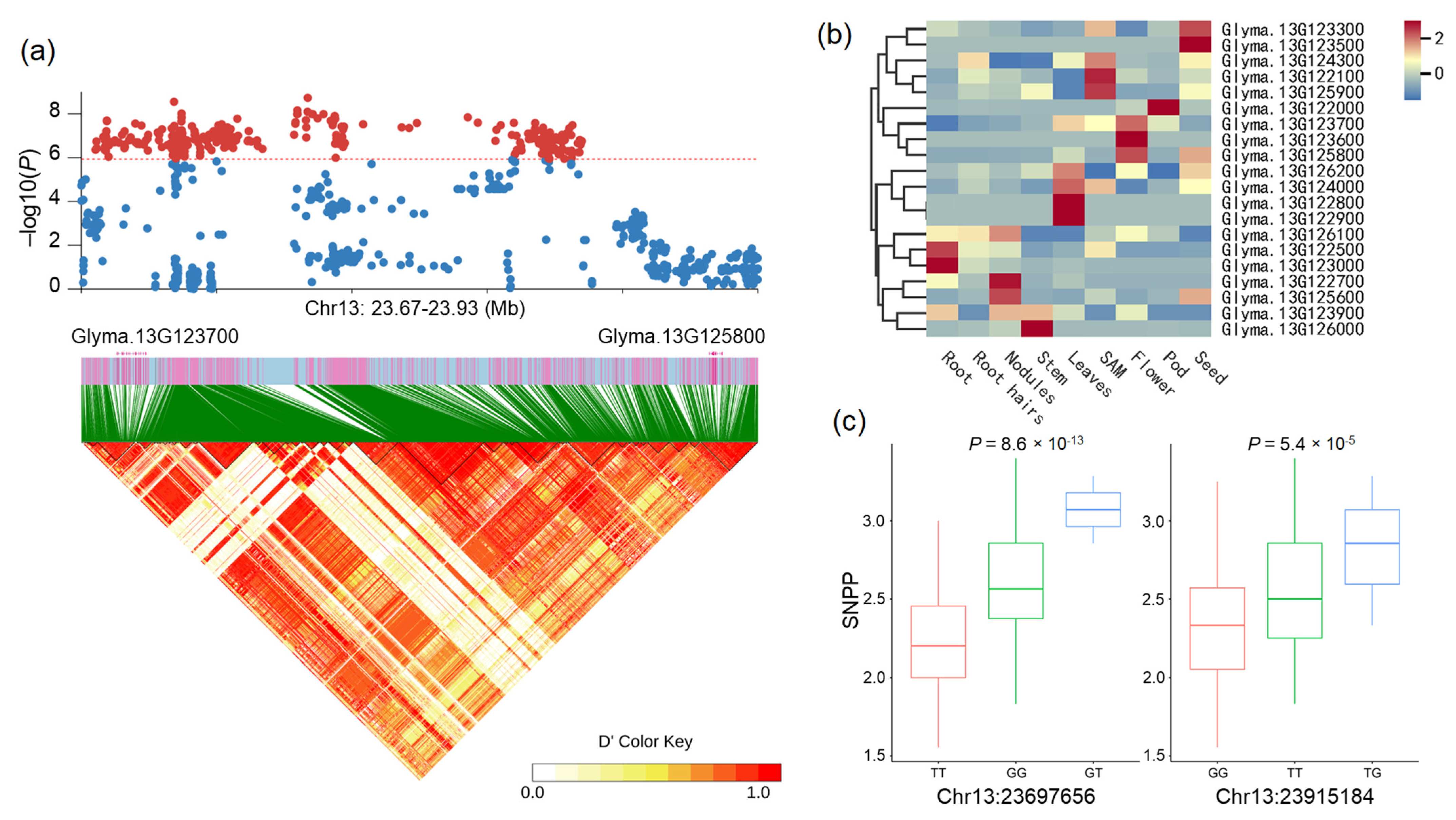
2.2. Genome-Wide Association Analysis
2.3. Identification of Candidate Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Sampling
4.2. Library Construction and SNP Identification
4.3. Statistical Analysis
4.4. GWAS
4.5. Identification of Candidate Genes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sá, A.G.A.; Moreno, Y.M.F.; Carciofi, B.A.M. Plant proteins as high-quality nutritional source for human diet. Trends Food Sci. Technol. 2020, 97, 170–184. [Google Scholar] [CrossRef]
- Carter, T.E., Jr.; Nelson, R.L.; Sneller, C.H.; Cui, Z. Genetic Diversity in Soybean. In Soybeans: Improvement, Production, and Uses; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2004; pp. 303–416. [Google Scholar]
- Pedersen, P.; Lauer, J.G. Response of Soybean Yield Components to Management System and Planting Date. Agron. J. 2004, 96, 1372–1381. [Google Scholar] [CrossRef]
- Takahashi, N. Linkage Relation between the Genes for the Form of Leaves and the Number of Seeds per Pod of Soybeans. Jpn. J. Genet. 1934, 9, 208–225. [Google Scholar] [CrossRef]
- You, M.G.; Liu, Y.B.; Zhou, T.J.; Gai, J.Y. Effects of leaf shape on seed yield and its components in soybeans. Soybean Genet. Newsl. 1995, 22, 66–70. [Google Scholar]
- Jeong, N.; Suh, S.J.; Kim, M.-H.; Lee, S.; Moon, J.-K.; Kim, H.S.; Jeong, S.-C. Ln Is a Key Regulator of Leaflet Shape and Number of Seeds per Pod in Soybean. Plant Cell 2012, 24, 4807–4818. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Li, W.; Li, G.; Wang, Z.; Zhou, Z.; Ma, Y.; Shen, Y.; Li, C.; Wu, Y.; Zhu, B.; et al. Cloning of Ln gene through combined approach of map-based cloning and association study in soybean. J. Genet. Genom. 2013, 40, 93–96. [Google Scholar] [CrossRef]
- Palmer, R.G.; Albertsen, M.C.; Heer, H. Pollen production in soybeans with respect to genotype, environment, and stamen position. Euphytica 1978, 27, 427–433. [Google Scholar] [CrossRef]
- Tischner, T.; Allphin, L.; Chase, K.; Orf, J.H.; Lark, K.G. Genetics of Seed Abortion and Reproductive Traits in Soybean. Crop Sci. 2003, 43, 464–473. [Google Scholar] [CrossRef]
- Palmer, R.G.; Kilen, T.C. Qualitative genetics and cytogenetics. Agron. A Ser. Monogr. Am. Soc. Agron. 1987, 16, 135–209. [Google Scholar]
- Cregan, P.B.; Jarvik, T.; Bush, A.L.; Shoemaker, R.C.; Lark, K.G.; Kahler, A.L.; Kaya, N.; VanToai, T.T.; Lohnes, D.G.; Chung, J.; et al. An Integrated Genetic Linkage Map of the Soybean Genome. Crop Sci. 1999, 39, 1464–1490. [Google Scholar] [CrossRef]
- Wang, Q.; Zhu, Y.; Zhou, M.; Zhang, W.; Zhang, H.; Chen, X.; Chen, H.; Cui, X. Genome-wide association analysis and candidate genes predication of leaf characteristics traits in soybean (Glycine max L.). Acta Agron. Sin. 2024, 50, 623–632. [Google Scholar]
- Weiss, M.G. Genetic Linkage in Soybeans. Linkage Group IV. Crop Sci. 1970, 10, 368–370. [Google Scholar] [CrossRef]
- Mandl, F.A.; Buss, G.R. Comparison of Narrow and Broad Leaflet Isolines of Soybean. Crop Sci. 1981, 21, 25–27. [Google Scholar] [CrossRef]
- Dinkins, R.D.; Keim, K.R.; Farno, L.; Edwards, L.H. Expression of the Narrow Leaflet Gene for Yield and Agronomic Traits in Soybean. J. Hered. 2002, 93, 346–351. [Google Scholar] [CrossRef]
- Jeong, N.; Moon, J.-K.; Kim, H.S.; Kim, C.-G.; Jeong, S.-C. Fine genetic mapping of the genomic region controlling leaflet shape and number of seeds per pod in the soybean. Theor. Appl. Genet. 2011, 122, 865–874. [Google Scholar] [CrossRef]
- Dong, S.-S.; He, W.-M.; Ji, J.-J.; Zhang, C.; Guo, Y.; Yang, T.-L. LDBlockShow: A fast and convenient tool for visualizing linkage disequilibrium and haplotype blocks based on variant call format files. Brief. Bioinform. 2020, 22, bbaa227. [Google Scholar] [CrossRef]
- Mallory, A.; Vaucheret, H. Form, Function, and Regulation of ARGONAUTE Proteins. Plant Cell 2010, 22, 3879–3889. [Google Scholar] [CrossRef]
- Rodríguez-Leal, D.; Castillo-Cobián, A.; Rodríguez-Arévalo, I.; Vielle-Calzada, J.-P. A Primary Sequence Analysis of the ARGONAUTE Protein Family in Plants. Front. Plant Sci. 2016, 7, 1347. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, W.; Guo, M.; Liu, S.; Liu, L.; Yu, Y.; Mo, B.; Chen, X.; Gao, L. Origin, evolution and diversification of plant ARGONAUTE proteins. Plant J. 2022, 109, 1086–1097. [Google Scholar] [CrossRef] [PubMed]
- Oliver, C.; Santos, J.L.; Pradillo, M. Accurate Chromosome Segregation at First Meiotic Division Requires AGO4, a Protein Involved in RNA-Dependent DNA Methylation in Arabidopsis thaliana. Genetics 2016, 204, 543–553. [Google Scholar] [CrossRef]
- Martín-Merchán, A.; Moro, B.; Bouet, A.; Bologna, N.G. Domain organization, expression, subcellular localization, and biological roles of ARGONAUTE proteins in Arabidopsis. J. Exp. Bot. 2023, 74, 2374–2388. [Google Scholar] [CrossRef]
- Latrasse, D.; Benhamed, M.; Henry, Y.; Domenichini, S.; Kim, W.; Zhou, D.-X.; Delarue, M. The MYST histone acetyltransferases are essential for gametophyte development in Arabidopsis. BMC Plant Biol. 2008, 8, 121. [Google Scholar] [CrossRef]
- Schubert, V.; Weißleder, A.; Ali, H.; Fuchs, J.; Lermontova, I.; Meister, A.; Schubert, I. Cohesin gene defects may impair sister chromatid alignment and genome stability in Arabidopsis thaliana. Chromosoma 2009, 118, 591–605. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Y.; Li, C.; Zang, J.; Gao, E.; Kroon, J.T.; Qu, X.; Hussey, P.J.; Wang, P. Exo84c interacts with VAP27 to regulate exocytotic compartment degradation and stigma senescence. Nat. Commun. 2023, 14, 4888. [Google Scholar] [CrossRef]
- Tejos, R.; Rodriguez-Furlán, C.; Adamowski, M.; Sauer, M.; Norambuena, L.; Friml, J. PATELLINS are regulators of auxin-mediated PIN1 relocation and plant development in Arabidopsis thaliana. J. Cell Sci. 2018, 131, jcs204198. [Google Scholar] [CrossRef]
- Cucinotta, M.; Di Marzo, M.; Guazzotti, A.; de Folter, S.; Kater, M.M.; Colombo, L. Gynoecium size and ovule number are interconnected traits that impact seed yield. J. Exp. Bot. 2020, 71, 2479–2489. [Google Scholar] [CrossRef] [PubMed]
- Smyth, D.R.; Bowman, J.L.; Meyerowitz, E.M. Early flower development in Arabidopsis. Plant Cell 1990, 2, 755–767. [Google Scholar]
- Nemhauser, J.L.; Feldman, L.J.; Zambryski, P.C. Auxin and ETTIN in Arabidopsis gynoecium morphogenesis. Development 2000, 127, 3877–3888. [Google Scholar] [CrossRef]
- Gomez, M.D.; Barro-Trastoy, D.; Escoms, E.; Saura-Sánchez, M.; Sánchez, I.; Briones-Moreno, A.; Vera-Sirera, F.; Carrera, E.; Ripoll, J.-J.; Yanofsky, M.F.; et al. Gibberellins negatively modulate ovule number in plants. Development 2018, 145, dev163865. [Google Scholar] [CrossRef] [PubMed]
- Stamm, P.; Kumar, P.P. Auxin and gibberellin responsive Arabidopsis SMALL AUXIN UP RNA36 regulates hypocotyl elongation in the light. Plant Cell Rep. 2013, 32, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Jiang, W.-B.; Hu, Y.-W.; Wu, P.; Zhu, J.-Y.; Liang, W.-Q.; Wang, Z.-Y.; Lin, W.-H. BR signal influences Arabidopsis ovule and seed number through regulating related genes expression by BZR1. Mol. Plant 2013, 6, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Kim, T.-W.; Oses-Prieto, J.A.; Sun, Y.; Deng, Z.; Zhu, S.; Wang, R.; Burlingame, A.L.; Wang, Z.-Y. BSKs mediate signal transduction from the receptor kinase BRI1 in Arabidopsis. Science 2008, 321, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin Regulates the Activity of Reproductive Meristems, Flower Organ Size, Ovule Formation, and Thus Seed Yield in Arabidopsis thaliana. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Schuler, M.A.; Paquette, S.M.; Werck-Reichhart, D.; Bak, S. Comparative Genomics of Rice and Arabidopsis. Analysis of 727 Cytochrome P450 Genes and Pseudogenes from a Monocot and a Dicot. Plant Physiol. 2004, 135, 756–772. [Google Scholar] [CrossRef]
- Mizutani, M.; Ohta, D. Diversification of P450 genes during land plant evolution. Annu. Rev. Plant Biol. 2010, 61, 291–315. [Google Scholar] [CrossRef]
- Helliwell, C.A.; Chandler, P.M.; Poole, A.; Dennis, E.S.; Peacock, W.J. The CYP88A cytochrome P450, ent-kaurenoic acid oxidase, catalyzes three steps of the gibberellin biosynthesis pathway. Proc. Natl. Acad. Sci. USA 2001, 98, 2065–2070. [Google Scholar] [CrossRef]
- Helliwell, C.A.; Sheldon, C.C.; Olive, M.R.; Walker, A.R.; Zeevaart, J.A.; Peacock, W.J.; Dennis, E.S. Cloning of the Arabidopsis ent-kaurene oxidase gene GA3. Proc. Natl. Acad. Sci. USA 1998, 95, 9019–9024. [Google Scholar] [CrossRef]
- Shi, Z.; Rao, Y.; Xu, J.; Hu, S.; Fang, Y.; Yu, H.; Pan, J.; Liu, R.; Ren, D.; Wang, X.; et al. Characterization and cloning of SMALL GRAIN 4, a novel DWARF11 allele that affects brassinosteroid biosynthesis in rice. Sci. Bull. 2015, 60, 905–915. [Google Scholar] [CrossRef]
- Zhou, Y.; Tao, Y.; Zhu, J.; Miao, J.; Liu, J.; Liu, Y.; Yi, C.; Yang, Z.; Gong, Z.; Liang, G. GNS4, a novel allele of DWARF11, regulates grain number and grain size in a high-yield rice variety. Rice 2017, 10, 34. [Google Scholar] [CrossRef]
- Zhou, Y.; Xie, Y.; Cai, J.; Liu, C.; Zhu, H.; Jiang, R.; Zhong, Y.; Zhang, G.; Tan, B.; Liu, G.; et al. Substitution mapping of QTLs controlling seed dormancy using single segment substitution lines derived from multiple cultivated rice donors in seven cropping seasons. Theor. Appl. Genet. 2017, 130, 1191–1205. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Zhang, H.; Sun, G.; Zhang, W.; Qiu, L. Evolution and association analysis of GmCYP78A10 gene with seed size/weight and pod number in soybean. Mol. Biol. Rep. 2015, 42, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Dai, A.; Wei, H.; Yang, S.; Wang, B.; Jiang, N.; Feng, X. Arabidopsis KLU homologue GmCYP78A72 regulates seed size in soybean. Plant Mol. Biol. 2016, 90, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, W.; Zhang, H.; Liu, X.; Cui, X.; Li, S.; Song, L.; Zhu, Y.; Chen, X.; Chen, H. Comparative selective signature analysis and high-resolution GWAS reveal a new candidate gene controlling seed weight in soybean. Theor. Appl. Genet. 2021, 134, 1329–1341. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Subgroup, G.P.D.P. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.M.; Sul, J.H.; Service, S.K.; Zaitlen, N.A.; Kong, S.-y.; Freimer, N.B.; Sabatti, C.; Eskin, E. Variance component model to account for sample structure in genome-wide association studies. Nat. Genet. 2010, 42, 348–354. [Google Scholar] [CrossRef]
- Li, M.-X.; Yeung, J.M.Y.; Cherny, S.S.; Sham, P.C. Evaluating the effective numbers of independent tests and significant p-value thresholds in commercial genotyping arrays and public imputation reference datasets. Hum. Genet. 2012, 131, 747–756. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Signal | Peak Loci | Chromosome | Position | p | Environment |
|---|---|---|---|---|---|
| 3_1.1 | Chr03:1129563 | 3 | 1129563 | 3.37 × 10−7 | NJ23, BLUP |
| 3_3.3 | Chr03:3343094 | 3 | 3343094 | 4.74 × 10−6 | NJ23, BLUP |
| 3_5.1 | Chr03:5123032 | 3 | 5123032 | 1.10 × 10−6 | NJ23, BLUP |
| 4_3.4 | Chr04:3411371 | 4 | 3411371 | 1.31 × 10−6 | NJ22, BLUP |
| 4_39.2 | Chr04:39179195 | 4 | 39179195 | 1.82 × 10−6 | NJ23, BLUP |
| 4_40.7 | Chr04:40744574 | 4 | 40744574 | 9.34 × 10−8 | NJ23, BLUP |
| 5_17.4 | Chr05:17445053 | 5 | 17445053 | 2.18 × 10−8 | NJ22, BLUP |
| 6_50.5 | Chr06:50457715 | 6 | 50457715 | 7.03 × 10−10 | NJ22, NJ23, BLUP |
| 7_6.2 | Chr07:6182076 | 7 | 6182076 | 9.50 × 10−6 | NJ23, BLUP |
| 10_3.3 | Chr10:3346012 | 10 | 3346012 | 8.03 × 10−10 | NJ22, NJ23, BLUP |
| 13_23.6 | Chr13:23620712 | 13 | 23620712 | 4.79 × 10−10 | NJ22, NJ23, BLUP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Zhang, W.; Xu, W.; Zhang, H.; Liu, X.; Chen, X.; Chen, H. Genome-Wide Association Study and Identification of Candidate Genes Associated with Seed Number per Pod in Soybean. Int. J. Mol. Sci. 2024, 25, 2536. https://doi.org/10.3390/ijms25052536
Wang Q, Zhang W, Xu W, Zhang H, Liu X, Chen X, Chen H. Genome-Wide Association Study and Identification of Candidate Genes Associated with Seed Number per Pod in Soybean. International Journal of Molecular Sciences. 2024; 25(5):2536. https://doi.org/10.3390/ijms25052536
Chicago/Turabian StyleWang, Qiong, Wei Zhang, Wenjing Xu, Hongmei Zhang, Xiaoqing Liu, Xin Chen, and Huatao Chen. 2024. "Genome-Wide Association Study and Identification of Candidate Genes Associated with Seed Number per Pod in Soybean" International Journal of Molecular Sciences 25, no. 5: 2536. https://doi.org/10.3390/ijms25052536