
Potential Therapeutic Application and Mechanism of Action of Stem Cell-Derived Extracellular Vesicles (EVs) in Systemic Lupus Erythematosus (SLE)

Abstract
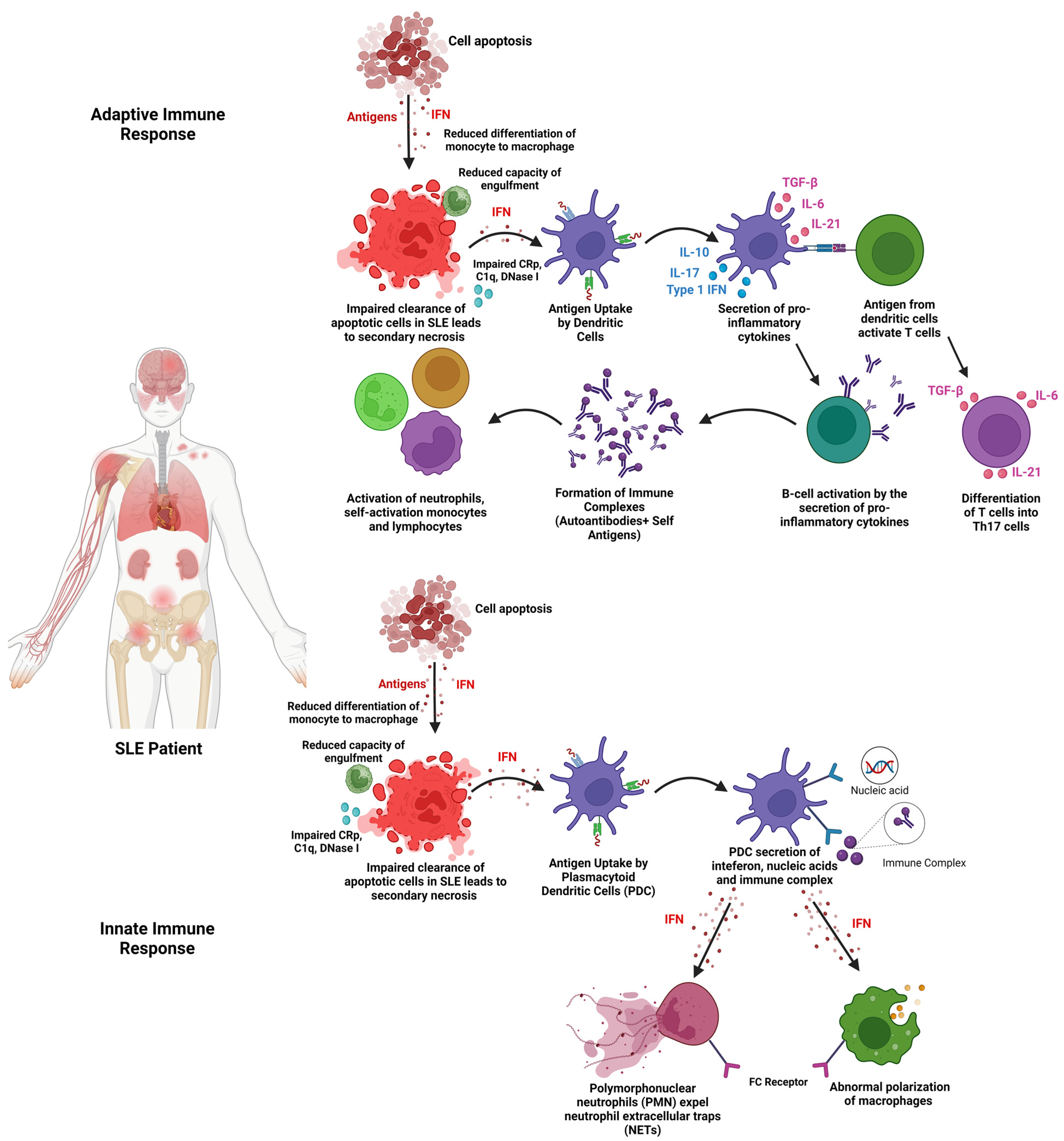
:1. Introduction
2. Extracellular Vesicles (EVs)
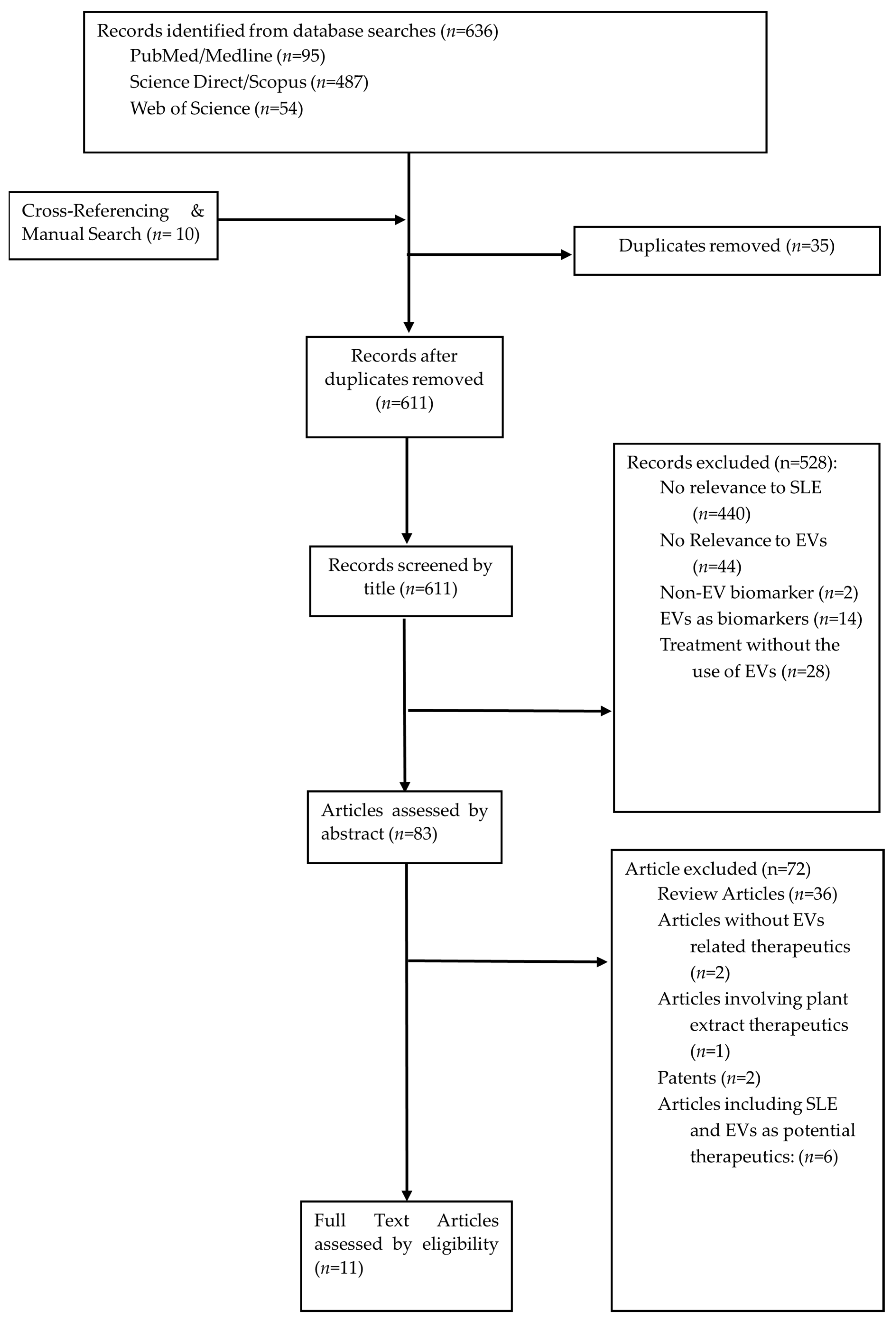
3. Methods
4. Identification of Relevant Studies
5. Study Selection
6. Collecting, Summarising, and Reporting Results
7. Results
8. Discussion
8.1. Isolation of EVs
8.2. Characterisation of EVs
8.3. Range of Dose of EVs Administered
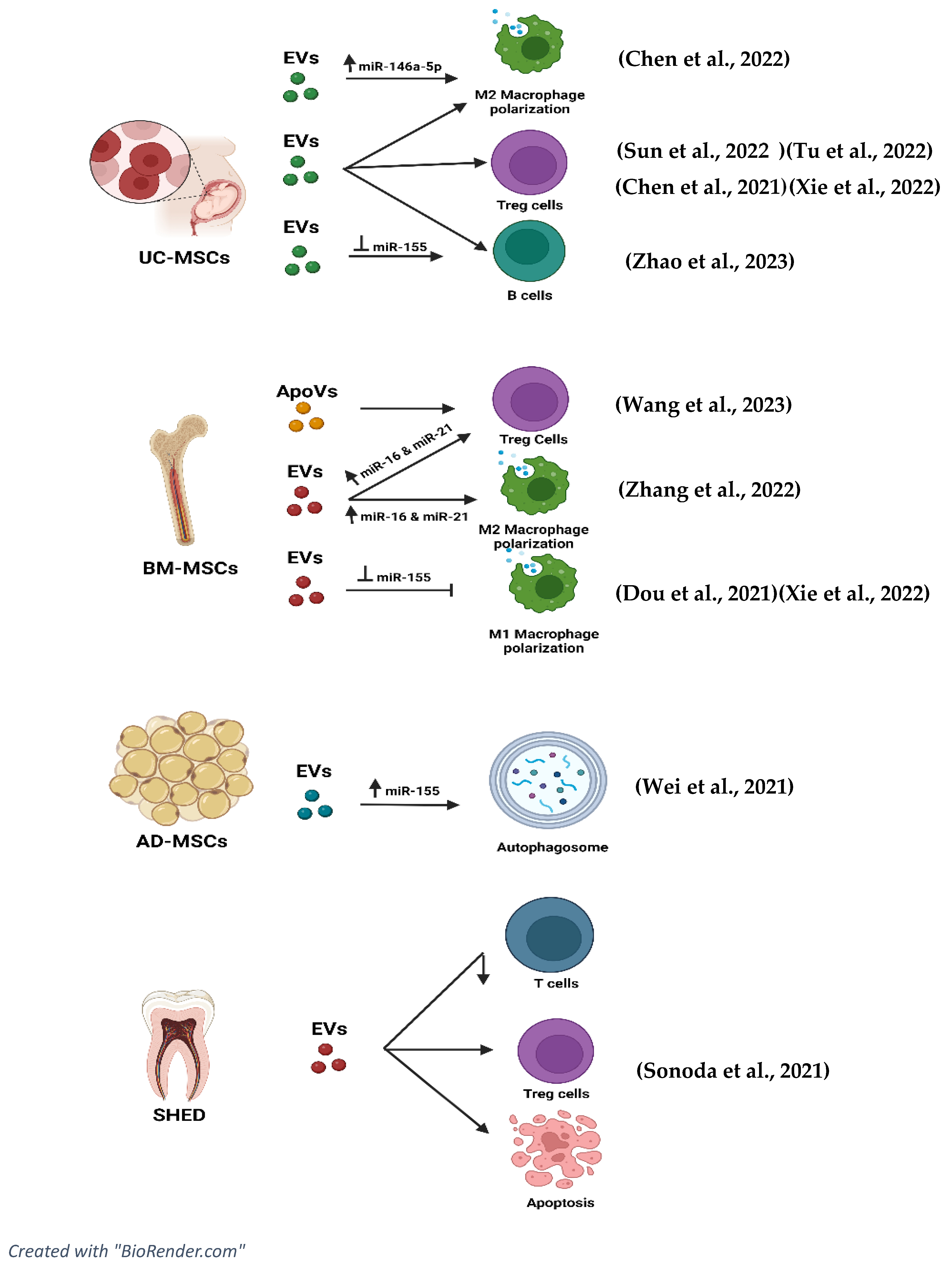
8.4. Mechanism of Action (EVs)
8.4.1. Effects of EVs on Pro-Inflammatory Cytokines
8.4.2. Effects of EVs on Anti-Inflammatory Cytokines
8.4.3. Effects of EVs on T Cell Lineage
8.4.4. Effects of EVs on B Cells
8.4.5. Effects of EVs on Lupus Nephritis (c-Complements)
8.5. Role of miRNAs and tsRNAs in Extracellular Vesicles in Ameliorating the Disease Progression of SLE
8.6. EVs and Signalling Pathways
9. Prospects for Future Research
9.1. Standardisation of EVs
9.2. Enhancing Immunomodulatory Effects of EVs
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, M.; Li, C.; She, Z.; Wu, F.; Mao, J.; Hun, M.; Luo, S.; Wan, W.; Tian, J.; Wen, C. Human umbilical cord mesenchymal stem cells derived extracellular vesicles regulate acquired immune response of lupus mouse in vitro. Sci. Rep. 2022, 12, 13101. [Google Scholar] [CrossRef]
- Justiz Vaillant, A.A.; Goyal, A.; Varacallo, M. Systemic Lupus Erythematosus. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Kuhn, A.; Bonsmann, G.; Anders, H.-J.; Herzer, P.; Tenbrock, K.; Schneider, M. The Diagnosis and Treatment of Systemic Lupus Erythematosus. Dtsch. Arztebl. Int. 2015, 112, 423–432. [Google Scholar] [CrossRef]
- Kamen, D.L. Environmental influences on systemic lupus erythematosus expression. Rheum. Dis. Clin. N. Am. 2014, 40, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Kernder, A.; Richter, J.G.; Fischer-Betz, R.; Winkler-Rohlfing, B.; Brinks, R.; Aringer, M.; Schneider, M.; Chehab, G. Delayed diagnosis adversely affects outcome in systemic lupus erythematosus: Cross sectional analysis of the LuLa cohort. Lupus 2021, 30, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Imran, S.; Thabah, M.M.; Azharudeen, M.; Ramesh, A.; Bobby, Z.; Negi, V.S. Liver Abnormalities in Systemic Lupus Erythematosus: A Prospective Observational Study. Cureus 2021, 13, e15691. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-L.; Zheng, H.-J.; Zhang, L.-Y.; Xu, Q.; Lin, C.-S. Case report: Joint deformity associated with systemic lupus erythematosus. Immun. Inflamm. Dis. 2022, 10, e717. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, J.F.; Cesic, D.; Keser, G.; Bukelica, M.; Karanagnostis, S.; Khamashta, M.A.; Hughes, G.R. Allergic disorders in systemic lupus erythematosus. Lupus 1993, 2, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Wuthisiri, W.; Lai, Y.H.; Capasso, J.; Blidner, M.; Salz, D.; Kruger, E.; Levin, A.V. Autoimmune retinopathy associated with systemic lupus erythematosus: A diagnostic dilemma. Taiwan J. Ophthalmol. 2017, 7, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Abu Bakar, F.; Sazliyana Shaharir, S.; Mohd, R.; Mohamed Said, M.S.; Rajalingham, S.; Wei Yen, K. Burden of Systemic Lupus Erythematosus on Work Productivity and Daily Living Activity: A Cross-Sectional Study Among Malaysian Multi-Ethnic Cohort. Arch. Rheumatol. 2020, 35, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.H.; Gopal, S.; Idris, I.B.; Aizuddin, A.N.; Miskam, H.M. Mental Health Status Among Systemic Lupus Erythematosus (SLE) Patients at Tertiary Hospital in Malaysia. Asian Soc. Work. J. 2020, 5, 30–39. [Google Scholar] [CrossRef]
- Fava, A.; Petri, M. Systemic lupus erythematosus: Diagnosis and clinical management. J. Autoimmun. 2019, 96, 1–13. [Google Scholar] [CrossRef]
- Kaul, A.; Gordon, C.; Crow, M.K.; Touma, Z.; Urowitz, M.B.; van Vollenhoven, R.; Ruiz-Irastorza, G.; Hughes, G. Systemic lupus erythematosus. Nat. Rev. Dis. Primers 2016, 2, 16039. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Lu, M.-P.; Wang, J.-H.; Xu, M.; Yang, S.-R. Immunological pathogenesis and treatment of systemic lupus erythematosus. World J. Pediatr. 2020, 16, 19–30. [Google Scholar] [CrossRef]
- McKeon, K.P.; Jiang, S.H. Treatment of systemic lupus erythematosus. Aust. Prescr. 2020, 43, 85–90. [Google Scholar] [CrossRef]
- Sakthiswary, R.; Suresh, E. Methotrexate in systemic lupus erythematosus: A systematic review of its efficacy. Lupus 2014, 23, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, L.; Shi, K.; Zhang, K.; Zheng, C.; Jin, Y. Extracellular Vesicles for Immunomodulation in Tissue Regeneration. Tissue Eng. Part C Methods 2022, 28, 393–404. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, aau6977. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kim, Y.; Ha, S.; Sheller-Miller, S.; Yoo, J.; Choi, C.; Park, C.H. The emerging role of exosomes as novel therapeutics: Biology, technologies, clinical applications, and the next. Am. J. Reprod. Immunol. 2021, 85, e13329. [Google Scholar] [CrossRef]
- Battistelli, M.; Falcieri, E. Apoptotic Bodies: Particular Extracellular Vesicles Involved in Intercellular Communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef]
- Huldani, H.; Jasim, S.A.; Bokov, D.O.; Abdelbasset, W.K.; Shalaby, M.N.; Thangavelu, L.; Margiana, R.; Qasim, M.T. Application of extracellular vesicles derived from mesenchymal stem cells as potential therapeutic tools in autoimmune and rheumatic diseases. Int. Immunopharmacol. 2022, 106, 108634. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Jeyaraj, M.; Qasim, M.; Kim, J.-H. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef]
- Li, M.; Li, S.; Du, C.; Zhang, Y.; Li, Y.; Chu, L.; Han, X.; Galons, H.; Zhang, Y.; Sun, H.; et al. Exosomes from different cells: Characteristics, modifications, and therapeutic applications. Eur. J. Med. Chem. 2020, 207, 112784. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, M.; Liu, S.; Guo, J.; Lu, Y.; Cheng, J.; Liu, J. Macrophage-derived extracellular vesicles: Diverse mediators of pathology and therapeutics in multiple diseases. Cell Death Dis. 2020, 11, 924. [Google Scholar] [CrossRef] [PubMed]
- Castro-Manrreza, M.E.; Montesinos, J.J. Immunoregulation by mesenchymal stem cells: Biological aspects and clinical applications. J. Immunol. Res. 2015, 2015, 394917. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.A.; Smuczek, B.; Valadão, I.C.; Dzik, L.M.; Iglesia, R.P.; Cruz, M.C.; Zelanis, A.; de Siqueira, A.S.; Serrano, S.M.T.; Goldberg, G.S.; et al. AHNAK enables mammary carcinoma cells to produce extracellular vesicles that increase neighboring fibroblast cell motility. Oncotarget 2016, 7, 49998–50016. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, P.M.M.; Alkhilaiwi, F.; Cavalli, I.J.; Malheiros, D.; de Souza Fonseca Ribeiro, E.M.; Cavalli, L.R. Extracellular vesicles from triple-negative breast cancer cells promote proliferation and drug resistance in non-tumorigenic breast cells. Breast Cancer Res. Treat. 2018, 172, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Stronati, E.; Conti, R.; Cacci, E.; Cardarelli, S.; Biagioni, S.; Poiana, G. Extracellular Vesicle-Induced Differentiation of Neural Stem Progenitor Cells. Int. J. Mol. Sci. 2019, 20, 3691. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Mei, J.; Ma, Y.; Jiang, S.; Zhang, J.; Yi, S.; Feng, C.; Liu, Y.; Liu, Y. Tumor hijacks macrophages and microbiota through extracellular vesicles. Exploration 2022, 2, 20210144. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.M.; Kroemer, G.; Zitvogel, L. Extracellular vesicles: Masters of intercellular communication and potential clinical interventions. J. Clin. Investig. 2016, 126, 1139–1143. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Wang, C. A review of the regulatory mechanisms of extracellular vesicles-mediated intercellular communication. Cell Commun. Signal. 2023, 21, 77. [Google Scholar] [CrossRef]
- Yekula, A.; Muralidharan, K.; Kang, K.M.; Wang, L.; Balaj, L.; Carter, B.S. From laboratory to clinic: Translation of extracellular vesicle based cancer biomarkers. Methods 2020, 177, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.L.; Escayg, A. Extracellular vesicles in the treatment of neurological disorders. Neurobiol. Dis. 2021, 157, 105445. [Google Scholar] [CrossRef] [PubMed]
- Akbar, N.; Azzimato, V.; Choudhury, R.P.; Aouadi, M. Extracellular vesicles in metabolic disease. Diabetologia 2019, 62, 2179–2187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, D.; Gao, Y.; Lin, C.; An, Q.; Feng, Y.; Liu, Y.; Liu, D.; Luo, H.; Wang, D. The Biology and Function of Extracellular Vesicles in Cancer Development. Front. Cell Dev. Biol. 2021, 9, 777441. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Kalimuthu, S.; Oh, J.M.; Gangadaran, P.; Baek, S.H.; Jeong, S.Y.; Lee, S.-W.; Lee, J.; Ahn, B.-C. Enhancement of antitumor potency of extracellular vesicles derived from natural killer cells by IL-15 priming. Biomaterials 2019, 190–191, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Collino, F.; Deregibus, M.C.; Grange, C.; Tetta, C.; Camussi, G. Microvesicles derived from human bone marrow mesenchymal stem cells inhibit tumor growth. Stem Cells Dev. 2013, 22, 758–771. [Google Scholar] [CrossRef]
- Katsuda, T.; Oki, K.; Ochiya, T. Potential application of extracellular vesicles of human adipose tissue-derived mesenchymal stem cells in Alzheimer’s disease therapeutics. In Renewal and Cell-Cell Communication; Turksen, K., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2015; Volume 1212, pp. 171–181. [Google Scholar] [CrossRef]
- d’Angelo, M.; Cimini, A.; Castelli, V. Insights into the Effects of Mesenchymal Stem Cell-Derived Secretome in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 5241. [Google Scholar] [CrossRef]
- Cosenza, S.; Toupet, K.; Maumus, M.; Luz-Crawford, P.; Blanc-Brude, O.; Jorgensen, C.; Noël, D. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics 2018, 8, 1399–1410. [Google Scholar] [CrossRef]
- Nojehdehi, S.; Soudi, S.; Hesampour, A.; Rasouli, S.; Soleimani, M.; Hashemi, S.M. Immunomodulatory effects of mesenchymal stem cell-derived exosomes on experimental type-1 autoimmune diabetes. J. Cell. Biochem. 2018, 119, 9433–9443. [Google Scholar] [CrossRef]
- Chen, X.; Su, C.; Wei, Q.; Sun, H.; Xie, J.; Nong, G. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Alleviate Diffuse Alveolar Hemorrhage Associated with Systemic Lupus Erythematosus in Mice by Promoting M2 Macrophage Polarization via the microRNA-146a-5p/NOTCH1 Axis. Immunol. Investig. 2022, 51, 1975–1993. [Google Scholar] [CrossRef]
- Tu, J.; Zheng, N.; Mao, C.; Liu, S.; Zhang, H.; Sun, L. UC-BSCs Exosomes Regulate Th17/Treg Balance in Patients with Systemic Lupus Erythematosus via miR-19b/KLF13. Cells 2022, 11, 4123. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, W.; Yuan, Z.; Li, M.; Wang, G.; Wang, L.; Liu, Y.; Diao, B. Exosome Derived from Human Umbilical Cord Mesenchymal Cell Exerts Immunomodulatory Effects on B Cells from SLE Patients. J. Immunol. Res. 2023, 2023, 3177584. [Google Scholar] [CrossRef]
- Chen, X.; Wei, Q.; Sun, H.M.; Zhang, X.B.; Yang, C.R.; Tao, Y.; Nong, G.M. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Regulate Macrophage Polarization to Attenuate Systemic Lupus Erythematosus-Associated Diffuse Alveolar Hemorrhage in Mice. Int. J. Stem Cells 2021, 14, 331–340. [Google Scholar] [CrossRef]
- Sun, W.; Yan, S.; Yang, C.; Yang, J.; Wang, H.; Li, C.; Zhang, L.; Zhao, L.; Zhang, J.; Cheng, M.; et al. Mesenchymal Stem Cells-derived Exosomes Ameliorate Lupus by Inducing M2 Macrophage Polarization and Regulatory T Cell Expansion in MRL/lpr Mice. Immunol. Investig. 2022, 51, 1785–1803. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.S.; Zhang, Z.W.; Yan, L.; Mo, Y.J.; Qiu, X.W.; Mi, X.B.; Lai, K. miR-20a Overexpression in Adipose-Derived Mesenchymal Stem Cells Promotes Therapeutic Efficacy in Murine Lupus Nephritis by Regulating Autophagy. Stem Cells Int. 2021, 2021, 3746335. [Google Scholar] [CrossRef] [PubMed]
- Dou, R.; Zhang, X.; Xu, X.; Wang, P.; Yan, B. Mesenchymal stem cell exosomal tsRNA-21109 alleviate systemic lupus erythematosus by inhibiting macrophage M1 polarization. Mol. Immunol. 2021, 139, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, S.; Murata, S.; Kato, H.; Zakaria, F.; Kyumoto-Nakamura, Y.; Uehara, N.; Yamaza, H.; Kukita, T.; Yamaza, T. Targeting of Deciduous Tooth Pulp Stem Cell-Derived Extracellular Vesicles on Telomerase-Mediated Stem Cell Niche and Immune Regulation in Systemic Lupus Erythematosus. J. Immunol. 2021, 206, 3053–3063. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Johnson-Stephenson, T.K.; Wang, W.; Wang, Y.; Li, J.; Li, L.; Zen, K.; Chen, X.; Zhu, D. Mesenchymal stem cell-derived exosome-educated macrophages alleviate systemic lupus erythematosus by promoting efferocytosis and recruitment of IL-17+ regulatory T cell. Stem Cell Res. Ther. 2022, 13, 484. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Hao, M.; Kou, X.; Sui, B.; Sanmillan, M.L.; Zhang, X.; Liu, D.; Tian, J.; Yu, W.; Chen, C.; et al. Apoptotic vesicles ameliorate lupus and arthritis via phosphatidylserine-mediated modulation of T cell receptor signaling. Bioact. Mater. 2023, 25, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Nordin, F.; Shaharir, S.S.; Abdul Wahab, A.; Mustafar, R.; Abdul Gafor, A.H.; Mohamed Said, M.S.; Rajalingham, S.; Shah, S.A. Serum and urine interleukin-17A levels as biomarkers of disease activity in systemic lupus erythematosus. Int. J. Rheum. Dis. 2019, 22, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Mohd, R.; Chin, S.-F.; Shaharir, S.S.; Cham, Q.S. Involvement of Gut Microbiota in SLE and Lupus Nephritis. Biomedicines 2023, 11, 653. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.M.; Edwards, M.R.; Mu, Q.; Yu, Y.; Vieson, M.D.; Reilly, C.M.; Ahmed, S.A.; Bankole, A.A. Gut Microbiota in Human Systemic Lupus Erythematosus and a Mouse Model of Lupus. Appl. Environ. Microbiol. 2018, 84, e02288-17. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Al-Haidari, A.; Sun, J.; Kazi, J.U. T cell receptor (TCR) signaling in health and disease. Signal Transduct. Target. Ther. 2021, 6, 412. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Park, J.; Takamatsu, H.; Konaka, H.; Aoki, W.; Aburaya, S.; Ueda, M.; Nishide, M.; Koyama, S.; Hayama, Y.; et al. Apoptosis-derived membrane vesicles drive the cGAS-STING pathway and enhance type I IFN production in systemic lupus erythematosus. Ann. Rheum. Dis. 2018, 77, 1507–1515. [Google Scholar] [CrossRef]
- Young, N.A.; Valiente, G.R.; Hampton, J.M.; Wu, L.-C.; Burd, C.J.; Willis, W.L.; Bruss, M.; Steigelman, H.; Gotsatsenko, M.; Amici, S.A.; et al. Estrogen-regulated STAT1 activation promotes TLR8 expression to facilitate signaling via microRNA-21 in systemic lupus erythematosus. Clin. Immunol. 2017, 176, 12–22. [Google Scholar] [CrossRef]
- Nelson, B.C.; Maragh, S.; Ghiran, I.C.; Jones, J.C.; DeRose, P.C.; Elsheikh, E.; Vreeland, W.N.; Wang, L. Measurement and standardization challenges for extracellular vesicle therapeutic delivery vectors. Nanomedicine 2020, 15, 2149–2170. [Google Scholar] [CrossRef] [PubMed]
- Willis, G.R.; Kourembanas, S.; Mitsialis, S.A. Toward Exosome-Based Therapeutics: Isolation, Heterogeneity, and Fit-for-Purpose Potency. Front. Cardiovasc. Med. 2017, 4, 63. [Google Scholar] [CrossRef]
- Collino, F.; Lopes, J.A.; Tapparo, M.; Tortelote, G.G.; Kasai-Brunswick, T.H.; Lopes, G.M.C.; Almeida, D.B.; Skovronova, R.; Wendt, C.H.C.; de Miranda, K.R.; et al. Extracellular Vesicles Derived from Induced Pluripotent Stem Cells Promote Renoprotection in Acute Kidney Injury Model. Cells 2020, 9, 453. [Google Scholar] [CrossRef]
- Azevedo, R.I.; Minskaia, E.; Fernandes-Platzgummer, A.; Vieira, A.I.S.; da Silva, C.L.; Cabral, J.M.S.; Lacerda, J.F. Mesenchymal stromal cells induce regulatory T cells via epigenetic conversion of human conventional CD4 T cells in vitro. Stem Cells 2020, 38, 1007–1019. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | EVs Type/miRNA Studied/Control | Disease Model | Modifications | EV Characterisation (Method) and Tracking | Pathway of Action | |
|---|---|---|---|---|---|---|
| Chen et al. [43] | miRNA overexpression Exosomes hUC-MSCs NOTCH1 overexpression or inhibition Control: PBS | C57BL/6J mice were induced with 0.5 mL pristane. | miR-146a-5p antagonist and agonist transfected to show inhibition/ overexpression | TEM Western blot assay to detect CD9 and CD36 NTA | miRNA expression: Antagomir miR-146a-5p shows lung injuries. Higher expression of NOTCH1, IL-1β, and iNOS. miR-146a-5p lowers levels of IL-10 and TGF-β. Exosomes inhibit NOTCH 1 expression by promoting the polarisation of M2 macrophage via miR-146a-5p. | Control Antagomir NC (exo). No significant difference in miR-146a-5p, NOTCH1, IL-1β, iNOS, CD206, Arg-1, and IL-10 expression levels. |
| No Modifications | EVs: Bleeding and inflammation in the lung tissues were alleviated post-treatment. Decreased levels of NOTCH1, IL-1β, and iNOS levels while increased levels of CD206, Arginase-1, and IL-10. | Control PBS Healthy Group (No evident changes). | ||||
| Overexpression modified with pcDNA3.1—NOTCH1 or pcDNA2.1 Inhibition—short hairpin (sh) RNA lentiviral vectors | NOTCH 1 Overexpression: Aggravated bleeding and inflammation. NOTCH1, IL-1β, and iNOS levels increased while CD206, Arginase-1, and IL-10 decreased. IL-6 and TNF-α concentrations increased while IL-10 and TGF-β decreased. NOTCH 1 Inhibition: Degree of injuries was reduced, and M2 macrophage polarisation was accelerated by sh-NOTCH1. Reducing NOTCH1, IL-1β, and iNOS. Enhanced CD206, Arginase-1, and IL-10 anti-inflammatory markers. Reduced expression of IL-6, TNF-α, and increased IL-10, or TGF-β. | Control (Agomir NC, Con) No difference in expression of miR-146a-5p, NOTCH1, IL-1 β, iNOS, CD206, Arg-1, and IL-10. | ||||
| Dou et al. [49] | tsRNA expression and Inhibition Human BMMSCs Exosomes Control: PBS | In vitro TPH-1 cell line + PMA to induce macrophage differentiation. (Mononuclear macrophage line) | tsRNA-21109 inhibition and expression | Hollow spherical microvesicle morphology Expression of CD36 Size: (100–200 nm) TEM | tsRNA expression: Reduced expression of tsRNA-21109 can be seen in patients with SLE. tsRNA-21109 can inhibit the M1 macrophage polarisation through Rap1, Ras, Hippo, Wnt, MAPK, and TGF-beta signalling pathways. Inhibition of tsRNA-21109: This leads to increased TNF-α and IL-1β, CD80, NOS2, and MCP-1 markers. Decreased in M2 markers CD206, MRC-2, and Arg-1. | Control MSCs-exo showed lower levels of CD80/ARG-1 compared to tsRNA-21109 inhibition. |
| Exosomes derived from MSCs | BMMSC EVs: Decrease in M1/M2 polarisation where expression of CD80, NOS2, and MCP-1 were decreased while CD206, MRC-2, and ARG1 increased. Reduction in TNF-α and IL-1β. Changed the TRF expression of M1 macrophages. | Control: Untreated macrophage culture showed expression of CD80, NOS2, and MCP-1 while reduction in CD206, MRC-2, and ARG-1. | ||||
| Sonoda et al. [50] | RNA and miRNA expression (SHED-EVs) SHED-EVs on BMMSC EVs Control: PBS | In vitro and In vivo Immunocompromised NOD-SCID mice BM-MSCs derived from NOD-SCID mice. | Small interfering RNA for Tert (siRNATert) and RAB27A (siRNARAB27A) | Surface Ag expression was analysed using an ExoAB Ab kit and R-PE-conjugated anti-rabbit IgG AB Particle size measured with NTA (69–478 nm) | RNA and siRNA on SHED EVs and effect on BMMSCs: MIR346 binds to a region in the TERTmRNA to upregulate TERT expression. Improves the functions of hematopoietic niche formation and immune regulation in recipient BMMSCs through the epigenetically regulated TERT telomerase activity pathway. | Control siRNACont (silencing RNA) Expression of Sca-1+, c-kit+, and CD45+ cells. |
| No Modifications | SHED EVs: Reduced the peripheral autoantibody levels, renal functions, and levels of CD4+ IL-17+ IFN-gamma and increased CD4+ CD25+ Foxp3+ in the PBMCs of the mice. | Control PBS solution Nontransplanted control (lpr-BMMSCs). Control wild type (B6-BMMSCs). Did not recover the TERT expression and telomerase activity. Reduced levels of Sca-1+, c-kit+, and CD45+ cells. | ||||
| No Modifications | SHED EVs on BMMSC EVs: Reduced CD4+ IL-17+ IFN-gamma cells and increased levels of CD4+ CD25+ Foxp3+ (Treg cells) and Annexin-V+ 7AAD+ cells. Reduced disease progression of SLE in mice. | |||||
| Tu et al. [44] | miR-19b expression UC-MSCs Exosomes Control: Healthy control group/ Negative control | In vitro model CD4+ cells of PBMC cells | miR-19b derived UC-MSCs that are transduced into CD4+ T cells via exosomes. | Rotund or oval-shaped exosomes NTA Size of exosomes: 132.5 (±) 37.4 nm Expressed CD63 and TSG101 | miR-19b expression: Lowered the levels of TNF, IL-6, and IL-17 and increased levels of IL-10 and TGF-beta. miR-19b inhibited the endogenous expression of KLF13 in T cells. Exosomes derived from UC-MSCs promote the expression of T cells in expressing miR-19b to inhibit the level of KLF13 and improve Th17/Treg cells regulation. | Control miR-19b NC miR-19b higher in PBMC of normal controls. Lower KLF13, TNF-α, IL-6, IL-17, and increased IL-10 and TGF- β. |
| Wei et al. [48] | miR-20a overexpression Adipose tissue-derived stem cells Exosomes Control: PBS | B6.MRL/lpr mice C57BL/6 mice | ADSCs transfected with miR-20a to produce miR-20a-ADSCs-derived exosomes. | CD63 marker was used to identify exosomes. None | miR-20a overexpression: Anti-dsDNA antibody, urine protein, and serum creatinine levels are the lowest in miR-20a-ADSC. Inhibited the Akt/mTOR pathway (LN pathogenesis). Higher autophagy markers of Beclin 1, LC3-II/LC3-I, and p62. Increased podocin and nephrin and higher autophagosomes. Reduced podocyte damage through autophagy. Fewer histopathologic abnormalities reduced C3 and IgG deposits and reduced nephritis scores. | Control Empty lentiviral vector (NC) Normal levels of Anti-dsDNA level and serum creatine. Lower C3 deposits in glomeruli, higher levels of Beclin 1 and LC3-II/LC3-1 and p62. Higher podocin and nephrin expression. Higher autophagosomes. |
| Zhang et al. [51] | miR-16 and miR-21 overexpression and inhibition BMMSCs Exosomes Control: PBS | In Vitro C57BL/6 mice were induced with pristane oil to obtain macrophage from the kidney. In vivo macrophages overexpression of miR-16 and miR-21 | BM-MSCs transduced with lenti-anti-miR-16 and lenti-anti-miR-21 (Exo/Anti-miRNAs) | Size of exosomes: 40 nm Exosome markers were detected using the immunoblot analysis (Alix and TSG-101) ExoGlow-Protein EV labelling kit (green, fluorescent dye) | miR-16 and miR-21 overexpression and inhibition: Reduced proinflammatory cytokines and increased anti-inflammatory cytokines. Reduction in iNOS, PDCD4, and PTEN levels. High expression of CCL20 (ligand on t cells). Higher expression of CD206, Arg-1, B7H4, and CD138 but lower expression of CD86 and iNOS. Macrophages ingested more apoptotic cells. Macrophage polarisation is contributed by the exosomes carrying miR-16 and miR-21 via targeting PDCD4 and PTEN. Increased efferocytosis of apoptotic cells. More production of Treg cells. | Control Macrophages transduced with Lent/ZIPmiR-Cont (ZIPmiR-Cont) Increased levels of miR-16 and miR-21 were elevated, and levels of iNOS, PDCD4, and PTEN decreased. Arg-1, CD206, B7H4 and CD138 markers increased. |
| No modifications | BMMSC EVs: Immune cell infiltration in the renal interstitium and mesangial expansion were significantly reduced. Lessened immune deposits (IgG, IgM, C3, and C1q). Reduced T cell infiltration. High levels of CD206 showing macrophage M2 polarisation. Reduced CD86 and increased CD206. Downregulated iNOS and upregulated Arg-1. Pro-inflammatory cytokines IFN-γ, IL-1β, IL-6, IL-12, and GM-CSF were reduced and anti-inflammatory cytokines IL-10, TGF-β, and M-CSF were increased. Reduced levels of ROS show a shift in macrophage polarisation. T cells differentiate into Treg cells. | |||||
| Zhao et al. [45] | miR-155 Inhibition Exosomes derived from (hucMSCs) Control: Healthy patients | In vitro PBMCs of SLE patients | No Modifications | Vesicles with a sphere shape in a size range of 130 nm NTA to track the exosomes. TEM to determine the morphology. Surface markers CD63 and CD81 | miR-155 inhibition: Inhibition of miR155- in B cells increases the SHIP-1 levels, reducing the activation of B cells. SHIP-1 activity promotes the apoptosis of B cells besides inhibiting its proliferation and overactivation. Inhibition of the ERK/SHIP-1 pathway can reduce inflammation. Inhibition of miR155 reduced IL-6 and TNF-α. | Control Lower expression of miR-155. Decreased levels of SHIP-1. Antagomir NC recorded lower levels of B cell apoptosis. Expressed IL-6 and TNF-α. p-ERK activation in B cells was not inhibited |
| No Modifications | EVs from hucMSCs: Reduced B cell hyperactivation. Reduced cytokine levels of IL-16, IL-10, INF-γ, IL-17, 1L-4, and TNF-α. High levels of IL-6- due to paracrine effect from MSCs. | Control Does not inhibit B cell proliferation. Does not overactivate B cells. IL-16, IL-10 and TNF-α were expressed. IL-6 is lower in the control group. Lower expression of miR-155. | ||||
| Chen et al. [46] | hucMSCs-Exo Control: PBS | C57BL/6J mice (SPF) induced by pristine to develop DAH | No Modifications | Round or elliptical shape with an intact capsule Exosome markers CD63, TSG101, and Alix Diameter range: 40–120 nm | EVs derived from hUC-MSC: Enhances the polarisation of M2 macrophage to alleviate DAH. Improved phagocytosis of macrophages in DAH. Facilitated the transformation of macrophages from M1 to M2 phenotype. iNOS, IL-6, TNF-α, and IL-1β deceased in M1 macrophage phenotype. Arg1, IL-10, TGF-β, and chi3l3 levels were upregulated in the M2 macrophage phenotype. | Control No changes in the F4/80 + CD11b + CD86 + CD206− cells. No changes in IL-6, iNOS, TNF-α, IL-10, and TGF- β levels. No changes in phagocytosis |
| Sun et al. [47] | hucMSCs-Exo Control: PBS | In vitro and In vivo pTHP-1 macrophages C57BL/6lpr-/-(B6.lpr) | No Modifications | The saucer-like shape of exosomes The diameter of exosomes ranged from 80 nm to 150 nm Markers determined from Western blot: CD9, TSG101, and GAPDH | EVs derived from hucMSCs (In vitro) Macrophage proliferation, inflammation, and M1 polarisation were inhibited post-treatment. Reduced expression of IL-1β and TNF-α and increased CD163+ M cells and CD206 + M cells but reduced HLA-DR+ and CD68+ cells. (In vivo) Reduced deposition of C3 immune complex. Increased CD14+ CD163+ markers of M2 cells and decreased CD14 + CD11c+ markers of M1 cells. The collagen fibre deposition in the glomeruli was significantly reduced. Increased serum IL-10 and decreased serum TNF-α and IFN-α. Increased CD4+ CD25 + FoxP3+ Treg cells. Alleviated nephritis and lung injury in MRL/lpr mice. Promoted the polarisation of M2 and Treg in MRL/lpr mice and the survival of mice. Reduction in pulmonary septal thickening and fibrosis. | Control No expression of IL-1 β and TNF-α. No significant difference in the deposition of C3 in glomeruli. No reduction in pulmonary septal thickening and fibrosis. Expression of CD14+ CD11c+ M1 cell infiltration. Lower CD14+ CD163+ M2 cells compared to EV treatment. No changes in IL-10, TNF-α and IFN-α. |
| Xie et al. [1] | hucMSCs-Exo Control: PBS | In vitro Splenic mononuclear cells of MRL/lpr mice | No modifications | Size of exosomes: 139.1 nm diameter TEM and NTA | EVs derived from hucMSCs: CD4+ T cells undergone inhibition in splenic mononuclear cells of MRL/lpr mice. Promoted Th17 cell differentiation. Increased the cytokine concentration of IL-17. The cytokine concentration of TGF -was increased. | Control Higher CD4+ T cells Lower Th17 cells Reduced concentration of IFNγ Lower levels of IL-4. |
| Wang et al. [52] | (BMMSCs) derived from C57b16. Apoptotic vesicles (ApoVs) derived from apoptotic BMMSCs. | In Vitro and In Vivo MRL/Ipr mice Apoptosis induced with staurosporine (STS) | No modifications | Spherical shape apoVs size = (100–350 nm) Apoptosis marker PtdSer/PS to validate the apoVs | apoVs derived from BMMSCs (in vivo): Decreased T cells in lymphoid tissue. Increased CD4+ T effector cells. Decreased CD4+ T naïve cells. Reduction in IFNγ+ and CD4+ T cells. Increased Foxp3+ CD4 + T cells effector. Reduction in lymphoproliferation. Reduced levels of anti-dsDNA and IgG. (In vitro) Reduced expansion of Th1 (IFNγ + CD4+), Th17 (IL-17A + CD4+) and Th2 (IL-4 + CD4+). Reduced levels of cytokines, including IFNγ, IL-17A, and IL-10. Foxp3+ and CD4+ T cells remained the same. Suppressed CD4+ T cells and IL-2 levels. ApoEVs diminished the phosphorylation of the TCR signalling pathway. | Control Non-Apoptotic control Higher Th17 cells. Severe arthritis. |
| Study | Disease Model | Source of EVs | Range of Dose | Experimental Groups | Outcome |
|---|---|---|---|---|---|
| [43] | DAH mouse model induced with pristane. | hUC-MSCs | 100 μL of 0.2 mg/mL EVs via intravenous injection through the tail vein every 2 days for 14 days. | n = 90 female rats 5 groups
| Promote polarisation of macrophages into M2 phenotype. Increases anti-inflammatory cytokines. Alleviation of bleeding and inflammation in the lungs. |
| [49] | In-Vitro TPH-1 cell line+PMA to induce macrophage polarisation and IFN γ+LPS to induce M1 phenotype. | BM-MSCs | None | None | Decrease in M1 polarisation and increase the polarisation into M2 phenotype. Reduction pro-inflammatory cytokines and increased levels of anti-inflammatory cytokines. |
| [50] | In Vivo Immunocompromised NOD-SCID mice In-Vitro BM-MSCs derived from NOD-SCID mice. | SHED | 100 μg of SHED EVs in 100 mL of PBS solution once in 4 weeks. | 3 groups
| Increased anti-inflammatory cytokines. Reduced disease progression of SLE. Recovered immune condition and renal functions. |
| [44] | In-Vitro Model CD4+ cells of PBMC cells. | UC-MSCs | None | None | Down-regulation of pro-inflammatory cytokines and up-regulation of anti-inflammatory cytokines. Regulation of Treg and T helper cells. |
| [48] | B6.MRL/Ipr Mice and C57BL/6 mice. | ADSC | 2 × 105 cells per 10 g animal weight in 150 μL for 14 days | 4 groups
| Reduction of disease severity and delayed lupus nephritis progression. Activation of autophagy and reduction of podocyte damage |
| [51] | In-Vitro Macrophage obtained from kidney of pristane induced mice. | BM-MSCs | None | None | Increased differentiation of T cells into Treg cells. Increased expression of anti-inflammatory markers. Reduction of lupus nephritis. |
| [45] | In-Vitro PBMC of SLE patients. | UC-MSCs | None | None | Increased immunomodulatory effects of B cells. Promotion of B cell apoptosis, prevention of B cell proliferation and activation. Regulation of the autoimmune reaction in PBMC of SLE patients. |
| [46] | DAH mouse model induced with pristane | hUC-MSCs | 100 μL of 0.2 mg/mL EVs via intravenous injection through the tail vein every 2 days for 14 days | n = 40 4 groups
| Promoted polarisation of macrophages into M2 phenotype. Increased anti-inflammatory cytokines. Improved phagocytosis of macrophages. |
| [47] | In-vitro pTHP-1 macrophages In-vivo C57BL/6lpr-/- (B6.lpr) | hUC-MSCs | Single dose of EVs of 200 μg protein concentration. | n = 18 3 groups
| Inhibited the proliferation of macrophages. Inhibited inflammation and M1 polarisation. Alleviated nephritis and lung injuries in mice. Promote polarisation of M2 and Treg in mice. Increased survival of mice. |
| [1] | In-Vitro Splenic Mononuclear Cells | hUC-MSCs | None | None | Promotion of the differentiation of Th17 cells. Regulation of immune cells and cytokine levels. |
| [52] | In Vitro & In Vivo MRL/Ipr mice | ApoVs BM-MSCs | Weekly administration for 4 weeks | 3 groups
| Promotion of immunomodulatory effects on T cells. Amelioration of inflammation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajeev Kumar, S.; Sakthiswary, R.; Lokanathan, Y. Potential Therapeutic Application and Mechanism of Action of Stem Cell-Derived Extracellular Vesicles (EVs) in Systemic Lupus Erythematosus (SLE). Int. J. Mol. Sci. 2024, 25, 2444. https://doi.org/10.3390/ijms25042444
Rajeev Kumar S, Sakthiswary R, Lokanathan Y. Potential Therapeutic Application and Mechanism of Action of Stem Cell-Derived Extracellular Vesicles (EVs) in Systemic Lupus Erythematosus (SLE). International Journal of Molecular Sciences. 2024; 25(4):2444. https://doi.org/10.3390/ijms25042444
Chicago/Turabian StyleRajeev Kumar, Sushmitha, Rajalingham Sakthiswary, and Yogeswaran Lokanathan. 2024. "Potential Therapeutic Application and Mechanism of Action of Stem Cell-Derived Extracellular Vesicles (EVs) in Systemic Lupus Erythematosus (SLE)" International Journal of Molecular Sciences 25, no. 4: 2444. https://doi.org/10.3390/ijms25042444