
Transforming Growth Factor α Evokes Aromatase Expression in Gastric Parietal Cells during Rat Postnatal Development

Abstract
:1. Introduction
2. Results
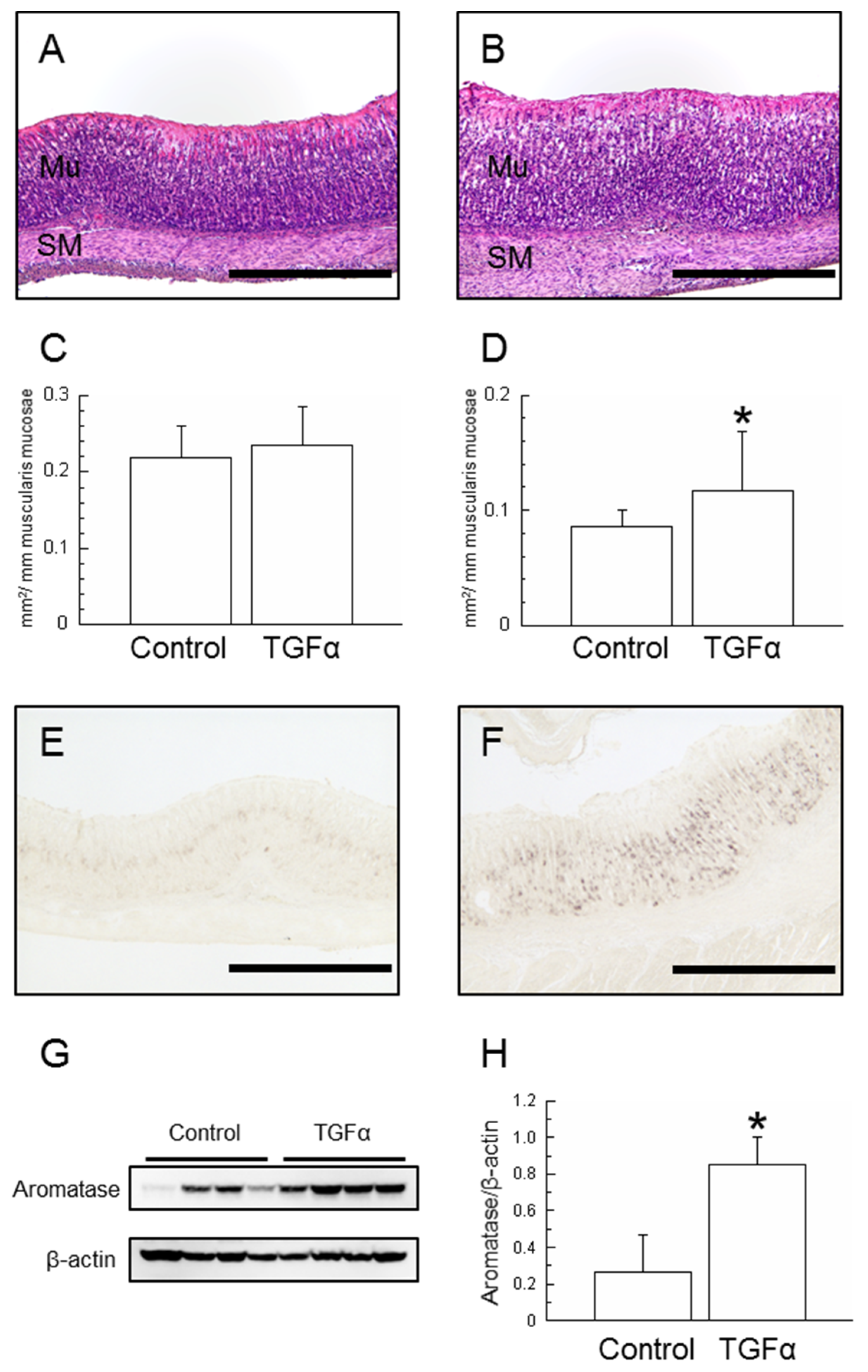
2.1. Effects of TGFα Administration
2.1.1. Morphological Changes in the Stomach
2.1.2. Aromatase Expression in the Stomach
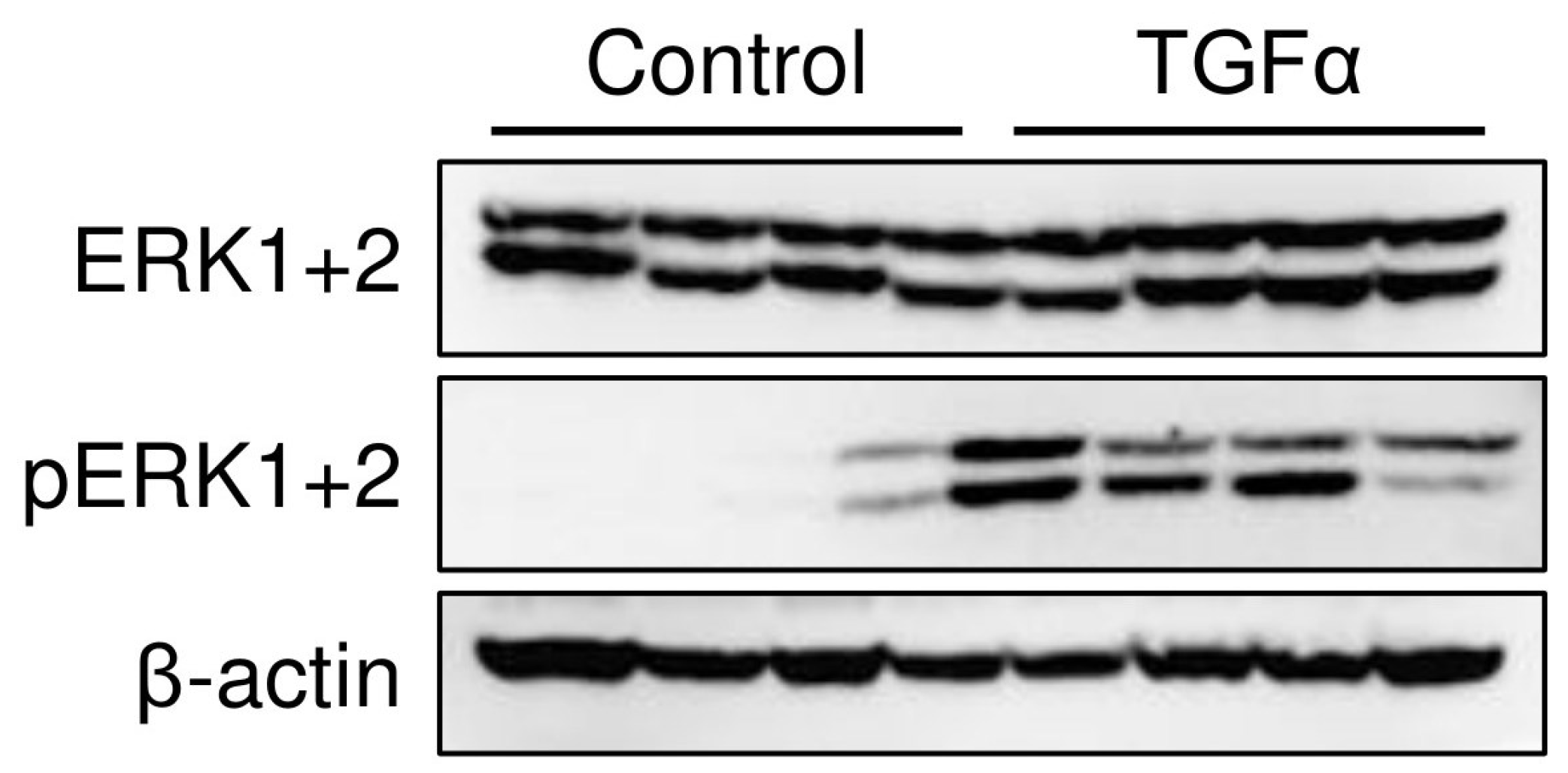
2.1.3. ERK1+2 Expression in the Stomach
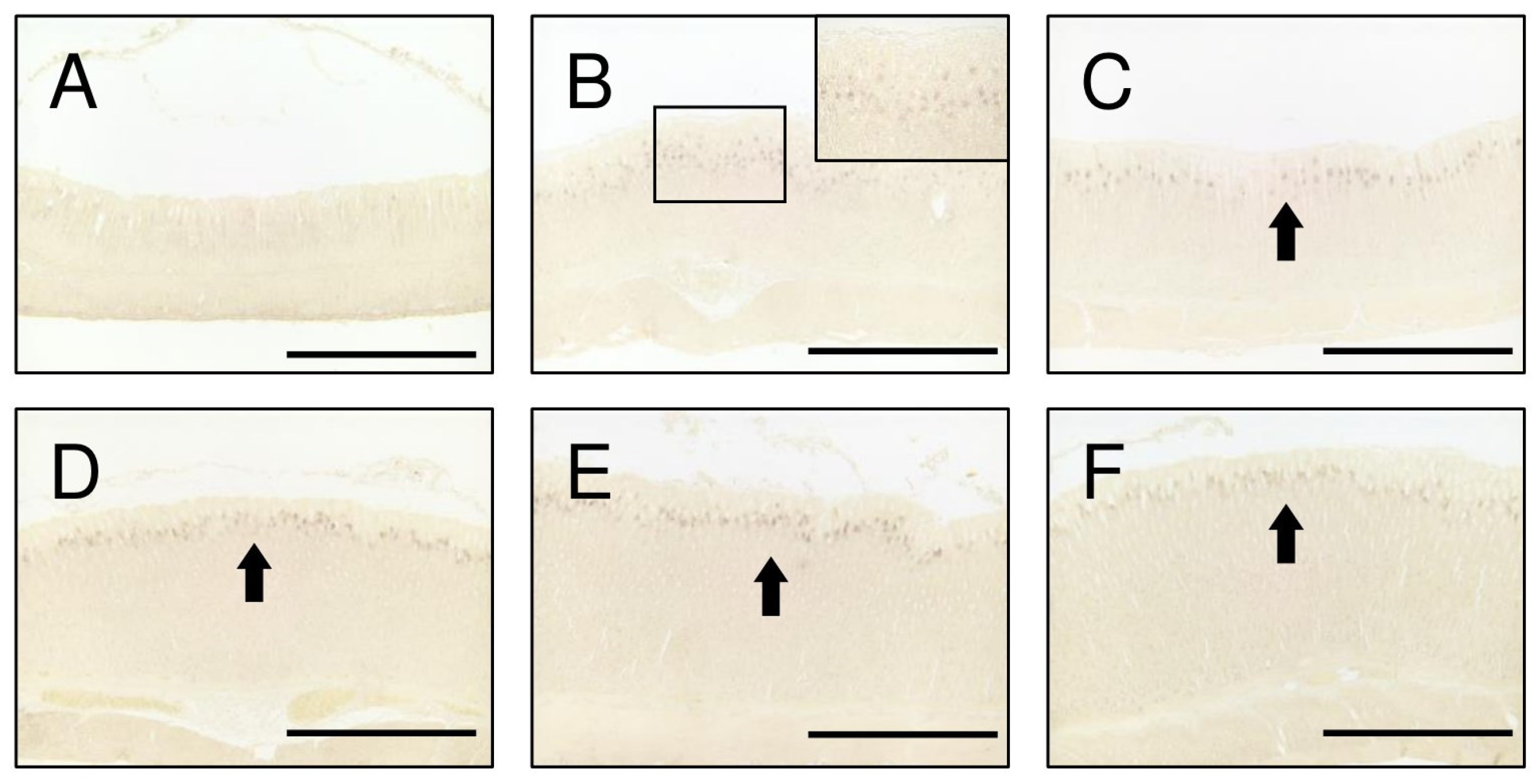
2.2. TGFα Expression in the Stomach during Postnatal Development
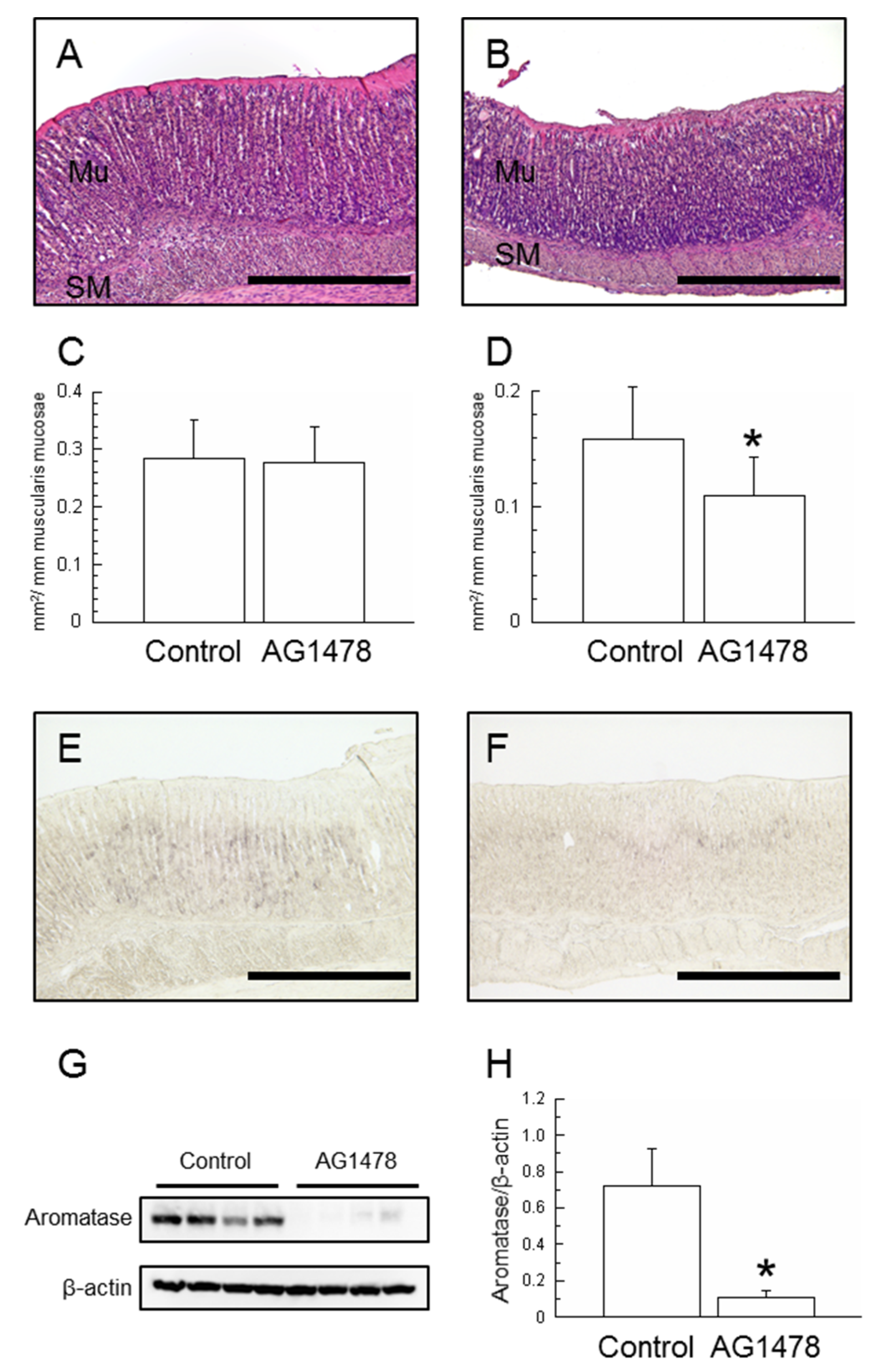
2.3. Effects of AG1478 Administration
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Histological Analysis
4.3. Western Blotting
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hori, T.; Ide, M.; Miyake, T. Ovarian estrogen secretion during the estrous cycle and under the influence of exogenous gonadotropins in rats. Endocrinol. Jpn. 1968, 15, 215–222. [Google Scholar] [CrossRef]
- Yoshinaga, K.; Hawkins, R.A.; Stocker, J.F. Estrogen secretion by the rat ovary in vivo during the estrous cycle and pregnancy. Endocrinology 1969, 85, 103–112. [Google Scholar] [CrossRef]
- Brown-Grant, K.; Exley, D.; Naftolin, F. Peripheral plasma oestradiol and luteinizing hormone concentrations during the oestrous cycle of the rat. J. Endocrinol. 1970, 48, 295–296. [Google Scholar] [CrossRef]
- Shaikh, A.A. Estrone and estradiol levels in the ovarian venous blood from rats during the estrous cycle and pregnancy. Biol. Reprod. 1971, 5, 297–307. [Google Scholar] [CrossRef]
- Dupon, C.; Kim, M.H. Peripheral plasma levels of testosterone, androstenedione, and oestradiol during the rat oestrous cycle. J. Endocrinol. 1973, 59, 653–654. [Google Scholar] [CrossRef]
- Butcher, R.L.; Collins, W.E.; Fugo, N.W. Plasma concentration of LH, FSH, prolactin, progesterone and estradiol-17beta throughout the 4-day estrous cycle of the rat. Endocrinology 1974, 94, 1704–1708. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, R.A.; Freedman, B.; Marshall, A.; Killen, E. Oestradiol-17 beta and prolactin levels in rat peripheral plasma. Br. J. Cancer 1975, 32, 179–185. [Google Scholar] [CrossRef]
- Nelson, L.R.; Bulun, S.E. Estrogen production and action. J. Am. Acad. Dermatol. 2001, 45 (Suppl. S3), S116–S124. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, T.; Shirasawa, N.; Matsuzawa, M.; Yamada, K.; Shelangouski, M.; Ito, T.; Tsuruo, Y. Gastric parietal cells: Potent endocrine role in secreting estrogen as a possible regulator of gastro-hepatic axis. Endocrinology 2002, 143, 3162–3170. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, T.; Shirasawa, N.; Ito, T.; Tsuruo, Y. Estrogen-producing steroidogenic pathways in parietal cells of the rat gastric mucosa. Life Sci. 2004, 74, 2327–2337. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Yamamoto, Y.; Yamagishi, N.; Kanai, Y. Stomach secretes estrogen in response to the blood triglyceride levels. Commun. Biol. 2021, 4, 1364. [Google Scholar] [CrossRef]
- Osaki, L.H.; Curi, M.A.F.; Alvares, E.P.; Gama, P. Early weaning accelerates the differentiation of mucous neck cells in rat gastric mucosa: Possible role of TGFalpha/EGFR. Differentiation 2010, 79, 48–56. [Google Scholar] [CrossRef]
- Hsuan, J.J. Transforming Growth Factor (TGF) Alpha. In Encyclopedia of Endocrine Diseases; Academic Press: Cambridge, MA, USA, 2004; pp. 605–611. [Google Scholar]
- Schubert, M.L. Gastric secretion. Curr. Opin. Gastroenterol. 2005, 21, 636–643. [Google Scholar] [CrossRef]
- Wang, L.; Lucey, M.R.; Fras, A.M.; Wilson, E.J.; Del Valle, J. Epidermal growth factor and transforming growth factor-alpha directly inhibit parietal cell function through a similar mechanism. J. Pharmacol. Exp. Ther. 1993, 265, 308–313. [Google Scholar] [PubMed]
- Coffey, R.J.; Romano, M.; Goldenring, J. Roles for transforming growth factor-alpha in the stomach. J. Clin. Gastroenterol. 1995, 21 (Suppl. S1), S36–S39. [Google Scholar] [PubMed]
- Takada, H.; Sasagawa, Y.; Yoshimura, M.; Tanaka, K.; Iwayama, Y.; Hayashi, T.; Isomura-Matoba, A.; Nikaido, I.; Kurisaki, A. Single-cell transcriptomics uncovers EGFR signaling-mediated gastric progenitor cell differentiation in stomach homeostasis. Nat. Commun. 2023, 14, 3750. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H. Estrogen synthesis in gastric parietal cells and secretion into portal vein. Anat. Sci. Int. 2020, 95, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Yoshida, S.; Sun, Y.J.; Shirasawa, N.; Naito, A. Postnatal development of gastric aromatase and portal venous estradiol-17β levels in male rats. J. Endocrinol. 2013, 218, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.J.; Goldenring, J.R.; Modlin, I.M.; Coffey, R.J. Inhibition of parietal cell H+ secretion by transforming growth factor alpha: A possible autocrine regulatory mechanism. Surgery 1990, 108, 220–227. [Google Scholar] [PubMed]
- Yuan, Q.X.; McRoberts, J.A.; Lakshmanan, J.; Yagi, H.; Hyman, P.E. Newborn rabbit gastric smooth muscle cell culture: EGF and TGF-alpha are potent mitogens. J. Pediatr. Gastroenterol. Nutr. 1993, 17, 153–160. [Google Scholar]
- Levitzki, A.; Gazit, A. Tyrosine kinase inhibition: An approach to drug development. Science 1995, 267, 1782–1788. [Google Scholar] [CrossRef]
- Osaki, L.H.; Figureueiredo, P.M.; Alvares, E.P.; Gama, P. EGFR is involved in control of gastric cell proliferation through activation of MAPK and Src signalling pathways in early-weaned rats. Cell Prolif. 2011, 44, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Redman, R.S.; Sweney, L.R. Changes in diet and patterns of feeding activity of developing rats. J. Nutr. 1976, 106, 615–626. [Google Scholar] [CrossRef]
- Henning, S.J. Postnatal development: Coordination of feeding, digestion, and metabolism. Am. J. Physiol. 1981, 241, G199–G214. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Shirasawa, N.; Naito, A. Age-related alterations of gastric mucosa and estrogen synthesis in rat parietal cells. Histochem. Cell Biol. 2022, 157, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R.; Liu, L.; Fligiel, S.E.; Jaszewski, R.; Majumdar, A.P. Aging alters gastric mucosal responses to epidermal growth factor and transforming growth factor-alpha. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G805–G810. [Google Scholar] [CrossRef]
- Shinohara, H.; Williams, C.S.; McWilliam, D.L.; Koldovský, O.; Philipps, A.F.; Dvorák, B. Transforming growth factor-alpha delays gastric emptying and small intestinal transit in suckling rats. Scand. J. Gastroenterol. 2001, 36, 356–360. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | TGFα | |
|---|---|---|
| Body weight (g) | 34.0 ± 1.7 | 42.4 ± 1.3 * |
| Stomach weight (mg) | 251.1 ± 14.4 | 350.7 ± 36.2 * |
| Stomach weight ratio (mg/100 g BW) | 745.8 ± 29.0 | 836.9 ± 82.4 * |
| Control | AG1478 | |
|---|---|---|
| Body weight (g) | 55.8 ± 5.1 | 51.7 ± 1.0 |
| Stomach weight (mg) | 342.3 ± 31.1 | 307.7 ± 6.0 * |
| Stomach weight ratio (mg/100 g BW) | 613.6 ± 14.2 | 594.9 ± 5.8 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, H.; Naito, A.; Kawagishi, K. Transforming Growth Factor α Evokes Aromatase Expression in Gastric Parietal Cells during Rat Postnatal Development. Int. J. Mol. Sci. 2024, 25, 2119. https://doi.org/10.3390/ijms25042119
Kobayashi H, Naito A, Kawagishi K. Transforming Growth Factor α Evokes Aromatase Expression in Gastric Parietal Cells during Rat Postnatal Development. International Journal of Molecular Sciences. 2024; 25(4):2119. https://doi.org/10.3390/ijms25042119
Chicago/Turabian StyleKobayashi, Hiroto, Akira Naito, and Kyutaro Kawagishi. 2024. "Transforming Growth Factor α Evokes Aromatase Expression in Gastric Parietal Cells during Rat Postnatal Development" International Journal of Molecular Sciences 25, no. 4: 2119. https://doi.org/10.3390/ijms25042119