
Maternal–Infant Factors in Relation to Extracellular Vesicle and Particle miRNA in Prenatal Plasma and in Postpartum Human Milk
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Participant Demographics
2.2. EVP miRNA Composition of Maternal Plasma during Pregnancy and Paired Human Milk Postpartum
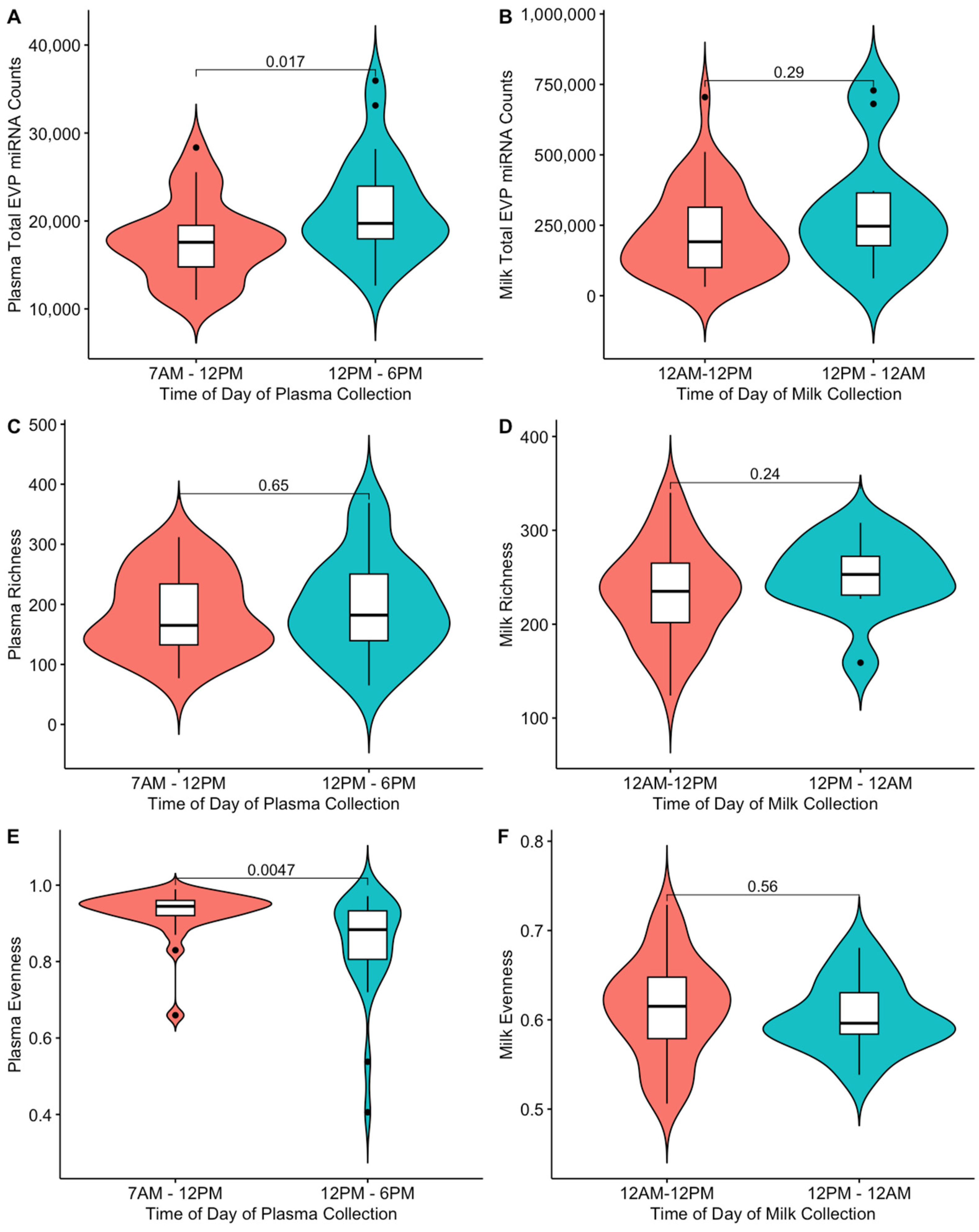
2.3. Time of Day of Blood Collection Is Associated with Maternal Plasma EVP miRNA Composition during Pregnancy
2.4. Delivery via Cesarean Section Is Associated with Lower Total EVP miRNA Counts in Milk and Higher Sample Evenness
2.5. Parity, Pre-Pregnancy Weight Status, and Infant Age at Sample Collection Are Negatively Associated with Total miRNA Counts in Human Milk EVPs
3. Discussion
4. Materials and Methods
4.1. Participant Recruitment
4.2. Maternal Blood Collection and Plasma EVP miRNA Extraction
4.3. Human Milk Collection, Processing, and EVP miRNA Extraction
4.4. Data Processing
4.5. Measures of EVP miRNA Composition
4.6. Statistical Analyses for Plasma EVP miRNA Composition
4.7. Statistical Analyses for Human Milk EVP miRNA Composition
4.8. Post Hoc Analyses Examining Individual miRNAs
4.9. Target Gene Identification and Pathway Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doyle, L.M.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Nair, S.; Salomon, C. Extracellular vesicles as critical mediators of maternal-fetal communication during pregnancy and their potential role in maternal metabolism. Placenta 2020, 98, 60–68. [Google Scholar] [CrossRef]
- Golan-Gerstl, R.; Reif, S. Extracellular vesicles in human milk. Curr. Opin. Clin. Nutr. Metab. Care 2022, 25, 209–215. [Google Scholar] [CrossRef]
- Hu, Y.; Thaler, J.; Nieuwland, R. Extracellular vesicles in human milk. Pharmaceuticals 2021, 14, 1050. [Google Scholar] [CrossRef]
- Sarker, S.; Scholz-Romero, K.; Perez, A.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E.; Salomon, C. Placenta-derived exosomes continuously increase in maternal circulation over the first trimester of pregnancy. J. Transl. Med. 2014, 12, 204. [Google Scholar] [CrossRef]
- Salomon, C.; Torres, M.J.; Kobayashi, M.; Scholz-Romero, K.; Sobrevia, L.; Dobierzewska, A.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E. A gestational profile of placental exosomes in maternal plasma and their effects on endothelial cell migration. PLoS ONE 2014, 9, e98667. [Google Scholar] [CrossRef] [PubMed]
- Foley, H.B.; Howe, C.G.; Eckel, S.P.; Chavez, T.; Gevorkian, L.; Reyes, E.G.; Kapanke, B.; Martinez, D.; Xue, S.; Suglia, S.F.; et al. Extracellular vesicle-enriched miRNA profiles across pregnancy in the MADRES cohort. PLoS ONE 2021, 16, e0251259. [Google Scholar] [CrossRef]
- Ospina-Prieto, S.; Chaiwangyen, W.; Herrmann, J.; Groten, T.; Schleussner, E.; Markert, U.R.; Morales-Prieto, D.M. MicroRNA-141 is upregulated in preeclamptic placentae and regulates trophoblast invasion and intercellular communication. Transl. Res. 2016, 172, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Biró, O.; Alasztics, B.; Molvarec, A.; Joó, J.; Nagy, B.; Rigó, J. Various levels of circulating exosomal total-miRNA and miR-210 hypoxamiR in different forms of pregnancy hypertension. Pregnancy Hypertens 2017, 10, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Schuster, J.; Cheng S bin Padbury, J.; Sharma, S. Placental extracellular vesicles and pre-eclampsia. Am. J. Reprod. Immunol. 2020, 85, e13297. [Google Scholar] [CrossRef]
- Nair, S.; Guanzon, D.; Jayabalan, N.; Lai, A.; Scholz-Romero, K.; Kalita de Croft, P.; Ormazabal, V.; Palma, C.; Diaz, E.; McCarthy, E.A.; et al. Extracellular vesicle-associated miRNAs are an adaptive response to gestational diabetes mellitus. J. Transl. Med. 2021, 19, 360. [Google Scholar] [CrossRef]
- Howe, C.G.; Foley, H.B.; Farzan, S.F.; Chavez, T.A.; Johnson, M.; Meeker, J.D.; Bastain, T.M.; Marsit, C.J.; Breton, C.V. Urinary metals and maternal circulating extracellular vesicle microRNA in the MADRES pregnancy cohort. Epigenetics 2022, 17, 1128–1142. [Google Scholar] [CrossRef]
- Howe, C.G.; Foley, H.B.; Kennedy, E.M.; Eckel, S.P.; Chavez, T.A.; Faham, D.; Grubbs, B.H.; Al-Marayati, L.; Lerner, D.; Suglia, S.; et al. Extracellular vesicle microRNA in early versus late pregnancy with birth outcomes in the MADRES study. Epigenetics 2021, 17, 269–285. [Google Scholar] [CrossRef]
- Bianco-Miotto, T.; Craig, J.M.; Gasser, Y.P.; van Dijk, S.J.; Ozanne, S.E. Epigenetics and DOHaD: From basics to birth and beyond. J. Dev. Orig. Health Dis. 2017, 8, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Du, X.; Li, J.; Lönnerdal, B. Human milk exosomes and their microRNAs survive digestion in vitro and are taken up by human intestinal cells. Mol. Nutr. Food Res. 2017, 61, 1–11. [Google Scholar] [CrossRef]
- Lin, D.; Chen, T.; Xie, M.; Li, M.; Zeng, B.; Sun, R.; Zhu, Y.; Ye, D.; Wu, J.; Sun, J.; et al. Oral Administration of Bovine and Porcine Milk Exosome Alter miRNAs Profiles in Piglet Serum. Sci. Rep. 2020, 10, 6983. [Google Scholar] [CrossRef] [PubMed]
- Manca, S.; Upadhyaya, B.; Mutai, E.; Desaulniers, A.T.; Cederberg, R.A.; White, B.R.; Zempleni, J. Milk exosomes are bioavailable and distinct microRNA cargos have unique tissue distribution patterns. Sci. Rep. 2018, 8, 11321. [Google Scholar] [CrossRef] [PubMed]
- Cione, E.; Zambrini, A.S.V.; Cannataro, R. MicroRNAs and Extracellular Vesicles in Milk: RNA-Based Micronutrients? J. Nutr. 2021, 151, 1378–1379. [Google Scholar] [CrossRef]
- Cui, J.; Zhou, B.; Ross, S.A.; Zempleni, J. Nutrition, microRNAs, and human health. Adv. Nutr. 2017, 8, 105–112. [Google Scholar] [CrossRef]
- Melnik, B.C.; Kakulas, F.; Geddes, D.T.; Hartmann, P.E.; John, S.M.; Carrera-Bastos, P.; Cordain, L.; Schmitz, G. Milk miRNAs: Simple nutrients or systemic functional regulators? Nutr. Metab. 2016, 13, 42. [Google Scholar] [CrossRef]
- Shah, K.B.; Chernausek, S.D.; Garman, L.D.; Pezant, N.P.; Plows, J.F.; Kharoud, H.K.; Demerath, E.W.; Fields, D.A. Human milk exosomal microrna: Associations with maternal overweight/obesity and infant body composition at 1 month of life. Nutrients 2021, 13, 1091. [Google Scholar] [CrossRef]
- Zamanillo, R.; Sánchez, J.; Serra, F.; Palou, A. Breast milk supply of microrna associated with leptin and adiponectin is affected by maternal overweight/obesity and influences infancy BMI. Nutrients 2019, 11, 2589. [Google Scholar] [CrossRef]
- Tao, S.C.; Guo, S.C. Extracellular vesicles: Potential participants in circadian rhythm synchronization. Int. J. Biol. Sci. 2018, 14, 1610. [Google Scholar] [CrossRef]
- Khalyfa, A.; Gaddameedhi, S.; Crooks, E.; Zhang, C.; Li, Y.; Qiao, Z.; Trzepizur, W.; Kay, S.A.; Andrade, J.; Satterfield, B.C.; et al. Circulating exosomal MiRNAs signal circadian misalignment to peripheral metabolic tissues. Int. J. Mol. Sci. 2020, 21, 6396. [Google Scholar] [CrossRef]
- Yeung, C.Y.C.; Dondelinger, F.; Schoof, E.M.; Georg, B.; Lu, Y.; Zheng, Z.; Zhang, J.; Hannibal, J.; Fahrenkrug, J.; Kjaer, M. Circadian regulation of protein cargo in extracellular vesicles. Sci. Adv. 2022, 8, eabc9061. [Google Scholar] [CrossRef]
- Zahoor, M.A.; Yao, X.D.; Henrick, B.M.; Verschoor, C.P.; Abimiku, A.; Osawe, S.; Rosenthal, K.L. Expression profiling of human milk derived exosomal microRNAs and their targets in HIV-1 infected mothers. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Guo, M.M.; Zhang, K.; Zhang, J.H. Human Breast Milk–Derived Exosomal miR-148a-3p Protects Against Necrotizing Enterocolitis by Regulating p53 and Sirtuin 1. Inflammation 2022, 45, 1254–1268. [Google Scholar] [CrossRef] [PubMed]
- Carney, M.C.; Tarasiuk, A.; Diangelo, S.L.; Silveyra, P.; Podany, A.; Birch, L.L.; Paul, I.M.; Kelleher, S.; Hicks, S.D. Metabolism-related microRNAs in maternal breast milk are influenced by premature delivery. Pediatr. Res. 2017, 82, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.; Liao, Y.; Du, X.; Xu, W.; Li, J.; Lönnerdal, B. Exosomal MicroRNAs in Milk from Mothers Delivering Preterm Infants Survive in Vitro Digestion and Are Taken Up by Human Intestinal Cells. Mol. Nutr. Food Res. 2018, 62, e1701050. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; MacIntyre, D.A.; Binkhamis, R.; Cook, J.; Sykes, L.; Bennett, P.R.; Terzidou, V. Maternal plasma miRNAs as potential biomarkers for detecting risk of small-for-gestational-age births. EBioMedicine 2020, 62, 103145. [Google Scholar] [CrossRef]
- Cabiati, M.; Randazzo, E.; Salvadori, C.; Peroni, D.; Federico, G.; del Ry, S. Circulating microRNAs associated with C-type natriuretic peptide in childhood obesity. Peptides 2020, 133, 170387. [Google Scholar] [CrossRef]
- Ouyang, S.; Tang, R.; Liu, Z.; Ma, F.; Li, Y.; Wu, J. Characterization and predicted role of microRNA expression profiles associated with early childhood obesity. Mol. Med. Rep. 2017, 16, 3799–3806. [Google Scholar] [CrossRef]
- Herrero-Aguayo, V.; Jiménez-Vacas, J.M.; Sáez-Martínez, P.; Gómez-Gómez, E.; López-Cánovas, J.L.; Garrido-Sánchez, L.; Herrera-Martínez, A.D.; García-Bermejo, L.; MacÍas-González, M.; López-Miranda, J.; et al. Influence of Obesity in the miRNome: MiR-4454, a Key Regulator of Insulin Response Via Splicing Modulation in Prostate. J. Clin. Endocrinol. Metab. 2021, 106, e469–e484. [Google Scholar] [CrossRef] [PubMed]
- Ramos, B.R.A.; Tronco, J.A.; Carvalho, M.; Felix, T.F.; Reis, P.P.; Silveira, J.C.; Silva, M.G. Circulating Extracellular Vesicles microRNAs Are Altered in Women Undergoing Preterm Birth. Int. J. Mol. Sci. 2023, 24, 5527. [Google Scholar] [CrossRef]
- Gray, C.; McCowan, L.M.; Patel, R.; Taylor, R.S.; Vickers, M.H. Maternal plasma miRNAs as biomarkers during mid-pregnancy to predict later spontaneous preterm birth: A pilot study. Sci. Rep. 2017, 7, 815. [Google Scholar] [CrossRef]
- Gutierrez, M.J.; Gomez, J.L.; Perez, G.F.; Pancham, K.; Val, S.; Pillai, D.K.; Giri, M.; Ferrante, S.; Freishtat, R.; Rose, M.C.; et al. Airway secretory micrornaome changes during rhinovirus infection in early childhood. PLoS ONE 2016, 11, e0162244. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Gu, W.W.; Gu, Y.; Yan, N.N.; Mao, Y.Y.; Zhen, X.X.; Wang, J.M.; Yang, J.; Shi, H.J.; Zhang, X.; et al. Association of the peripheral blood levels of circulating microRNAs with both recurrent miscarriage and the outcomes of embryo transfer in an in vitro fertilization process. J. Transl. Med. 2018, 16, 186. [Google Scholar] [CrossRef] [PubMed]
- Zierden, H.C.; Marx-Rattner, R.; Rock, K.D.; Montgomery, K.R.; Anastasiadis, P.; Folts, L.; Bale, T.L. Extracellular vesicles are dynamic regulators of maternal glucose homeostasis during pregnancy. Sci. Rep. 2023, 13, 1–14. [Google Scholar]
- Saenz-de-Juano, M.D.; Silvestrelli, G.; Ulbrich, S.E. Circadian Rhythm Does Not Affect the miRNA Cargo of Bovine Raw Milk Extracellular Vesicles. Int. J. Mol. Sci. 2023, 24, 10210. [Google Scholar] [CrossRef]
- Khodayar-Pardo, P.; Mira-Pascual, L.; Collado, M.C.; Martínez-Costa, C. Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J. Perinatol. 2014, 34, 599–605. [Google Scholar] [CrossRef]
- Dewey, K.G. Maternal and fetal stress are associated with impaired lactogenesis in humans. J. Nutr. 2001, 131, 3012S–3015S. [Google Scholar] [CrossRef]
- Dewey, K.G.; Nommsen-Rivers, L.A.; Heinig, M.J.; Cohen, R.J. Risk factors for suboptimal infant breastfeeding behavior, delayed onset of lactation, and excess neonatal weight loss. Pediatrics 2003, 112, 607–619. [Google Scholar] [CrossRef]
- Evans, K.C.; Evans, R.G.; Royal, R.; Esterman, A.J.; James, S.L. Effect of caesarean section on breast milk transfer to the normal term newborn over the first week of life. Arch. Dis. Child.-Fetal Neonatal Ed. 2003, 88, 380F–382. [Google Scholar] [CrossRef]
- Samuel, T.M.; Thielecke, F.; Lavalle, L.; Chen, C.; Fogel, P.; Giuffrida, F.; Dubascoux, S.; Martínez-Costa, C.; Haaland, K.; Marchini, G.; et al. Mode of Neonatal Delivery Influences the Nutrient Composition of Human Milk: Results from a Multicenter European Cohort of Lactating Women. Front. Nutr. 2022, 9, 834394. [Google Scholar] [CrossRef]
- Hermansson, H.; Kumar, H.; Collado, M.C.; Salminen, S.; Isolauri, E.; Rautava, S. Breast milk microbiota is shaped by mode of delivery and intrapartum antibiotic exposure. Front. Nutr. 2019, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Yerlikaya, F.H.; Eryavuz Onmaz, D.; Altunhan, H.; Ilhan, M. Can altered colostrum miRNA expression profile after cesarean delivery be a risk factor for autoimmune diseases? Am. J. Reprod. Immunol. 2021, 86, e13472. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, M.; Lavi-Avnon, Y.; Zagoory-Sharon, O.; Weller, A.; Moran, T.H. Preobesity in the infant OLETF rat: The role of suckling. Dev. Psychobiol. 2007, 49, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, S.; Haba, D.; Schroeder, M.; Smith, G.P.; Weller, A. Independent ingestion and microstructure of feeding patterns in infant rats lacking CCK-1 receptors. Am. J. Physiol. Integr. Comp. Physiol. 2006, 290, R208–R218. [Google Scholar] [CrossRef] [PubMed]
- Marchini, G.; Linden, A. Cholecystokinin, a satiety signal in newborn infants? J. Dev. Physiol. 1992, 17, 215–219. [Google Scholar] [PubMed]
- Ozaki, T.; Mohammad, S.; Morioka, E.; Takiguchi, S.; Ikeda, M. Infant satiety depends on transient expression of cholecystokinin-1 receptors on ependymal cells lining the third ventricle in mice. J. Physiol. 2013, 591, 1295–1312. [Google Scholar] [CrossRef] [PubMed]
- Eccles, S.A. The epidermal growth factor receptor/Erb-B/HER family in normal and malignant breast biology. Int. J. Dev. Biol. 2011, 55, 685–696. [Google Scholar] [CrossRef] [PubMed]
- Darcy, K.M.; Wohlhueter, A.L.; Zangani, D.; Vaughan, M.M.; Russell, J.A.; Masso-Welch, P.A.; Varela, L.M.; Shoemaker, S.F.; Horn, E.; Lee, P.P.H.; et al. Selective changes in EGF receptor expression and function during the proliferation, differentiation and apoptosis of mammary epithelial cells. Eur. J. Cell Biol. 1999, 78, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Pond, A.C.; Bin, X.; Batts, T.; Roarty, K.; Hilsenbeck, S.; Rosen, J.M. Fibroblast growth factor receptor signaling is essential for normal mammary gland development and stem cell function. Stem Cells 2013, 31, 178–189. [Google Scholar] [CrossRef]
- Huang, J.; Dai, B.; Qu, H.; Zhong, Y.; Ma, Y.; Luo, J.; Hadsell, D.; Shi, H. Epidermal growth factor stimulates fatty acid synthesis mainly via plc-γ1/akt signaling pathway in dairy goat mammary epithelial cells. Animals 2020, 10, 930. [Google Scholar] [CrossRef]
- Kupsco, A.; Prada, D.; Valvi, D.; Hu, L.; Petersen, M.S.; Coull, B.; Grandjean, P.; Weihe, P.; Baccarelli, A.A. Human milk extracellular vesicle miRNA expression and associations with maternal characteristics in a population-based cohort from the Faroe Islands. Sci. Rep. 2021, 11, 5840. [Google Scholar] [CrossRef]
- Gomez-Gallego, C.; Garcia-Mantrana, I.; Salminen, S.; Collado, M.C. The human milk microbiome and factors influencing its composition and activity. Semin. Fetal Neonatal Med. 2016, 21, 400–405. [Google Scholar] [CrossRef]
- Demmelmair, H.; Jiménez, E.; Collado, M.C.; Salminen, S.; McGuire, M.K. Maternal and perinatal factors associated with the human milk microbiome. Curr. Dev. Nutr. 2020, 4, nzaa027. [Google Scholar] [CrossRef]
- Lundgren, S.N.; Madan, J.C.; Karagas, M.R.; Morrison, H.G.; Hoen, A.G.; Christensen, B.C. Microbial Communities in Human Milk Relate to Measures of Maternal Weight. Front. Microbiol. 2019, 10, 2886. [Google Scholar] [CrossRef] [PubMed]
- Han, E.; Abrams, B.; Sridhar, S.; Xu, F.; Hedderson, M. Validity of Self-Reported Pre-Pregnancy Weight and Body Mass Index Classification in an Integrated Health Care Delivery System. Paediatr. Périnat. Epidemiol. 2016, 30, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Gilbert-Diamond, D.; Cottingham, K.L.; Gruber, J.F.; Punshon, T.; Sayarath, V.; Gandolfi, A.J.; Baker, E.R.; Jackson, B.P.; Folt, C.L.; Karagas, M.R. Rice consumption contributes to arsenic exposure in US women. Proc. Natl. Acad. Sci. USA 2011, 108, 20656–20660. [Google Scholar] [CrossRef]
- Howe, C.G.; Armstrong, D.A.; Muse, M.E.; Gilbert-Diamond, D.; Gui, J.; Hoen, A.G.; Palys, T.J.; Barnaby, R.L.; Stanton, B.A.; Jackson, B.P.; et al. Periconceptional and Prenatal Exposure to Metals and Extracellular Vesicle and Particle miRNAs in Human Milk: A Pilot Study. Expo. Heal. 2022, 15, 731–743. [Google Scholar] [CrossRef]
- Frørup, C.; Mirza, A.H.; Yarani, R.; Nielsen, L.B.; Mathiesen, E.R.; Damm, P.; Svare, J.; Engelbrekt, C.; Størling, J.; Johannesen, J.; et al. Plasma Exosome-Enriched Extracellular Vesicles from Lactating Mothers with Type 1 Diabetes Contain Aberrant Levels of miRNAs During the Postpartum Period. Front. Immunol. 2021, 12, 744509. [Google Scholar] [CrossRef]
- Witwer, K.W.; Goberdhan, D.C.I.; O’Driscoll, L.; Théry, C.; Welsh, J.A.; Blenkiron, C.; Buzás, E.I.; Di Vizio, D.; Erdbrügger, U.; Falcón-Pérez, J.M.; et al. Updating MISEV: Evolving the minimal requirements for studies of extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12182. [Google Scholar] [CrossRef]
- Salas, L.A.; Lundgren, S.N.; Browne, E.P.; Punska, E.C.; Anderton, D.L.; Karagas, M.R.; Arcaro, K.F.; Christensen, B.C. Prediagnostic breast milk DNA methylation alterations in women who develop breast cancer. Hum. Mol. Genet. 2020, 29, 662–673. [Google Scholar] [CrossRef]
- Waggott, D.; Chu, K.; Yin, S.; Wouters, B.G.; Liu, F.F.; Boutros, P.C. NanoStringNorm: An extensible R package for the pre-processing of nanostring mRNA and miRNA data. Bioinformatics 2012, 28, 1546–1548. [Google Scholar] [CrossRef]
- Hill, T.C.J.; Walsh, K.A.; Harris, J.A.; Moffett, B.F. Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Rousseeuw, P.; Croux, C.; Todorov, V.; Ruckstuhl, A.; Salibian-Barrera, M.; Verbeke, T.; Koller, M.; Maechler, M.M. Robustbase: Basic Robust Statistics; R Package. 2015. Available online: http://CRAN.R-project.org/package=robustbase (accessed on 21 November 2023).
- Hauschild, A.C.; Pastrello, C.; Ekaputeri, G.K.A.; Bethune-Waddell, D.; Abovsky, M.; Ahmed, Z.; Kotlyar, M.; Lu, R.; Jurisica, I. MirDIP 5.2: Tissue context annotation and novel microRNA curation. Nucleic Acids Res. 2023, 51, D217–D225. [Google Scholar] [CrossRef]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Demographic | Mean (SD) or n (%) |
|---|---|
| Maternal Age (yrs) | 32.6 (4.0) |
| Maternal Education | |
| High School Graduate or Less | 4 (7.4) |
| Some College | 3 (5.6) |
| College Graduate or More | 47 (87.0) |
| Maternal Relationship Status | |
| Married | 45 (83.3) |
| Separated or Never Married | 9 (16.7) |
| Maternal Pre-Pregnancy BMI (kg/m2) | 23.9 (4.9) |
| Maternal Pre-Pregnancy Weight Status | |
| BMI < 18.5 kg/m2 | 3 (5.6) |
| BMI 18.5–25 kg/m2 | 39 (72.2) |
| BMI > 25 kg/m2 | 12 (22.2) |
| Gestational Weight Gain (lbs) | 34.8 (18.9) |
| Infant Sex | |
| Female | 30 (55.6) |
| Male | 24 (44.4) |
| Gestational Age at Delivery (wks) | 39.2 (1.5) |
| Delivery | |
| Preterm (<37 weeks) | 6 (11.1) |
| Term (≥37 weeks) | 48 (88.9) |
| Delivery Mode | |
| Vaginal | 40 (74.1) |
| Cesarean Section | 14 (25.9) |
| Gestational Age at Plasma Collection (wks) | 28.8 (2.8) |
| Time of Day of Plasma Collection | |
| Morning (7:00 AM–12:00 PM) | 27 (50.0) |
| Afternoon (12:00 PM–6:00 PM) | 27 (50.0) |
| Infant Age at Milk Collection (wks) | 6.1 (0.8) |
| Time of Day of Milk Collection a | |
| Morning (12:00 AM–12:00 PM) | 40 (76.9) |
| Afternoon (12:00 PM–12:00 AM) | 12 (23.1) |
| Total Counts a | Richness | Evenness | ||||
|---|---|---|---|---|---|---|
| Effect Size (95% CI) | p Value | Effect Size (95% CI) | p Value | Effect Size (95% CI) | p Value | |
| Maternal Age (yrs) | 0.003 (−0.022, 0.028) | 0.824 | −3.463 (−8.636, 1.709) | 0.195 | −0.002 (−0.009, 0.005) | 0.517 |
| Parity b | ||||||
| Nulliparous | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| Parous | 0.157 (−0.047, 0.361) | 0.137 | 18.096 (−23.588, 59.780) | 0.399 | 0.057 (0, 0.114) | 0.056 |
| Pre-Pregnancy BMI (kg/m2) | −0.001 (−0.022, 0.019) | 0.887 | −0.294 (−4.432, 3.844) | 0.890 | 0.001 (−0.005, 0.007) | 0.701 |
| Pre-Pregnancy Weight Statusc | ||||||
| BMI 18.5–25 kg/m2 | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| BMI > 25 kg/m2 | 0.006 (−0.218, 0.229) | 0.961 | 15.197 (−34.074, 64.468) | 0.548 | 0.036 (−0.031, 0.103) | 0.301 |
| Infant Sex | ||||||
| Female | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| Male | −0.096 (−0.294, 0.103) | 0.349 | −6.903 (−47.798, 33.992) | 0.742 | 0.002 (−0.054, 0.058) | 0.945 |
| GA at Blood Collection (days) | 0.001 (−0.004, 0.006) | 0.720 | 0.405 (−0.614, 1.423) | 0.440 | 0 (−0.001, 0.002) | 0.866 |
| Time of Day of Blood Collection | ||||||
| 7 AM–12 PM | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| 12 PM–6 PM | 0.234 (0.037, 0.430) | 0.024 | 8.693 (−31.48, 48.867) | 0.673 | −0.093 (−0.148, −0.038) | 0.002 |
| Total Counts a | Richness | Evenness | ||||
|---|---|---|---|---|---|---|
| Effect Size (95% CI) | p Value | Effect Size (95% CI) | p Value | Effect Size (95% CI) | p Value | |
| Maternal Age (yrs) | −0.034 (−0.099, 0.031) | 0.307 | −1.855 (−5.237, 1.527) | 0.288 | 0 (−0.003, 0.004) | 0.806 |
| Parity b | ||||||
| Primiparous | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| Multiparous | −0.526 (−1.032, −0.019) | 0.047 | −17.262 (−43.69, 9.166) | 0.207 | 0.014 (−0.014, 0.041) | 0.331 |
| Pre-Pregnancy BMI (kg/m2) | −0.032 (−0.086, 0.023) | 0.257 | 0.711 (−2.155, 3.576) | 0.629 | 0.001 (−0.002, 0.004) | 0.704 |
| Pre-Pregnancy Weight Status c | ||||||
| BMI 18.5–25 kg/m2 | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| BMI > 25 kg/m2 | −0.645 (−1.232, −0.058) | 0.037 | −13.548 (−45.98, 18.884) | 0.417 | 0.022 (−0.012, 0.055) | 0.209 |
| Infant Sex | ||||||
| Female | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| Male | 0.389 (−0.124, 0.901) | 0.144 | 22.036 (−4.706, 48.777) | 0.113 | −0.018 (−0.045, 0.01) | 0.219 |
| Gestational Weight Gain (lbs) | 0.009 (−0.005, 0.023) | 0.196 | 0.48 (−0.233, 1.193) | 0.193 | −0.001 (−0.001, 0) | 0.120 |
| Gestational Age at Birth (wks) | 0.034 (−0.151, 0.218) | 0.723 | −0.169 (−9.811, 9.474) | 0.973 | −0.001 (−0.011, 0.009) | 0.870 |
| Delivery Mode | ||||||
| Vaginal | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| Cesarean Section | −0.703 (−1.28, −0.125) | 0.021 | −14.626 (−44.762, 15.51) | 0.346 | 0.040 (0.009, 0.071) | 0.015 |
| Infant Age at Milk Collection (days) | −0.049 (−0.095, −0.004) | 0.040 | −0.655 (−3.039, 1.73) | 0.593 | 0 (−0.003, 0.002) | 0.774 |
| Milk Collection Time b | ||||||
| 12:00 AM–12:00 PM | Ref. | Ref. | Ref. | Ref. | Ref. | Ref. |
| 12:00 PM–12:00 AM | 0.21 (−0.412, 0.833) | 0.511 | 13.052 (−19.668, 45.771) | 0.438 | −0.012 (−0.047, 0.022) | 0.481 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muse, M.E.; Armstrong, D.A.; Hoen, A.G.; Gilbert-Diamond, D.; Gui, J.; Palys, T.J.; Kolling, F.W.; Christensen, B.C.; Karagas, M.R.; Howe, C.G. Maternal–Infant Factors in Relation to Extracellular Vesicle and Particle miRNA in Prenatal Plasma and in Postpartum Human Milk. Int. J. Mol. Sci. 2024, 25, 1538. https://doi.org/10.3390/ijms25031538
Muse ME, Armstrong DA, Hoen AG, Gilbert-Diamond D, Gui J, Palys TJ, Kolling FW, Christensen BC, Karagas MR, Howe CG. Maternal–Infant Factors in Relation to Extracellular Vesicle and Particle miRNA in Prenatal Plasma and in Postpartum Human Milk. International Journal of Molecular Sciences. 2024; 25(3):1538. https://doi.org/10.3390/ijms25031538
Chicago/Turabian StyleMuse, Meghan E., David A. Armstrong, Anne G. Hoen, Diane Gilbert-Diamond, Jiang Gui, Thomas J. Palys, Frederick W. Kolling, Brock C. Christensen, Margaret R. Karagas, and Caitlin G. Howe. 2024. "Maternal–Infant Factors in Relation to Extracellular Vesicle and Particle miRNA in Prenatal Plasma and in Postpartum Human Milk" International Journal of Molecular Sciences 25, no. 3: 1538. https://doi.org/10.3390/ijms25031538