
Role of Mn-LIPA in Sex Hormone Regulation and Gonadal Development in the Oriental River Prawn, Macrobrachium nipponense
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Molecular Cloning and Structural Analysis of the LIPA Gene
2.2. Expression of the Mn-LIPA Gene in Different Tissues in Males
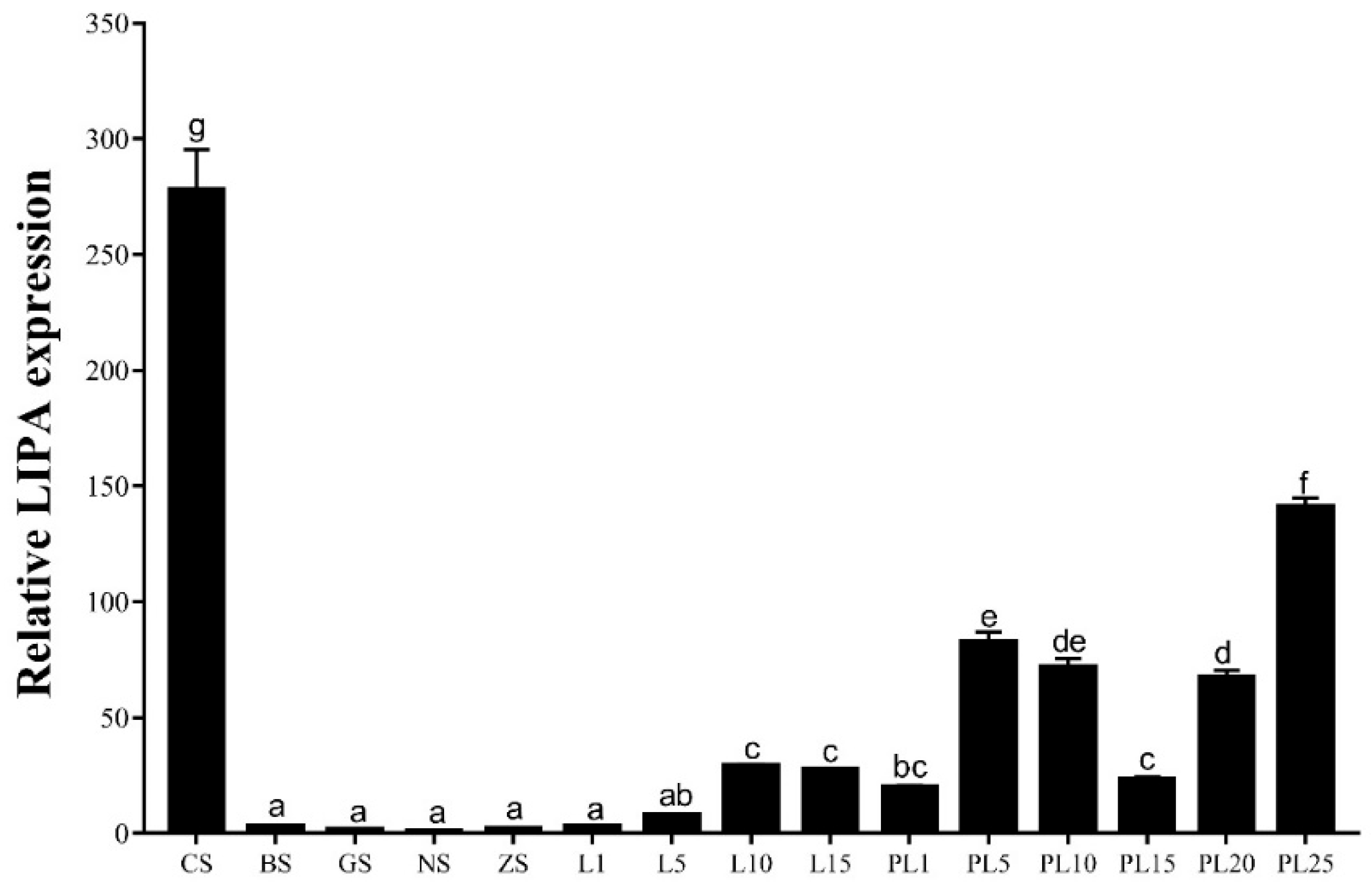
2.3. Expression of the Mn-LIPA Gene during Different Developmental Stages
2.4. Localization of the Mn-LIPA Gene in Testes and Hepatopancreas
2.5. Effects of Mn-DCHR RNAi Knockdown on PL10-Stage M. nipponense
2.5.1. Sex Ratio
2.5.2. Steroid Hormone Content
2.5.3. RNA Interference Efficiency
2.6. Effects of Mn-DCHR RNAi Knockdown on Male M. nipponense
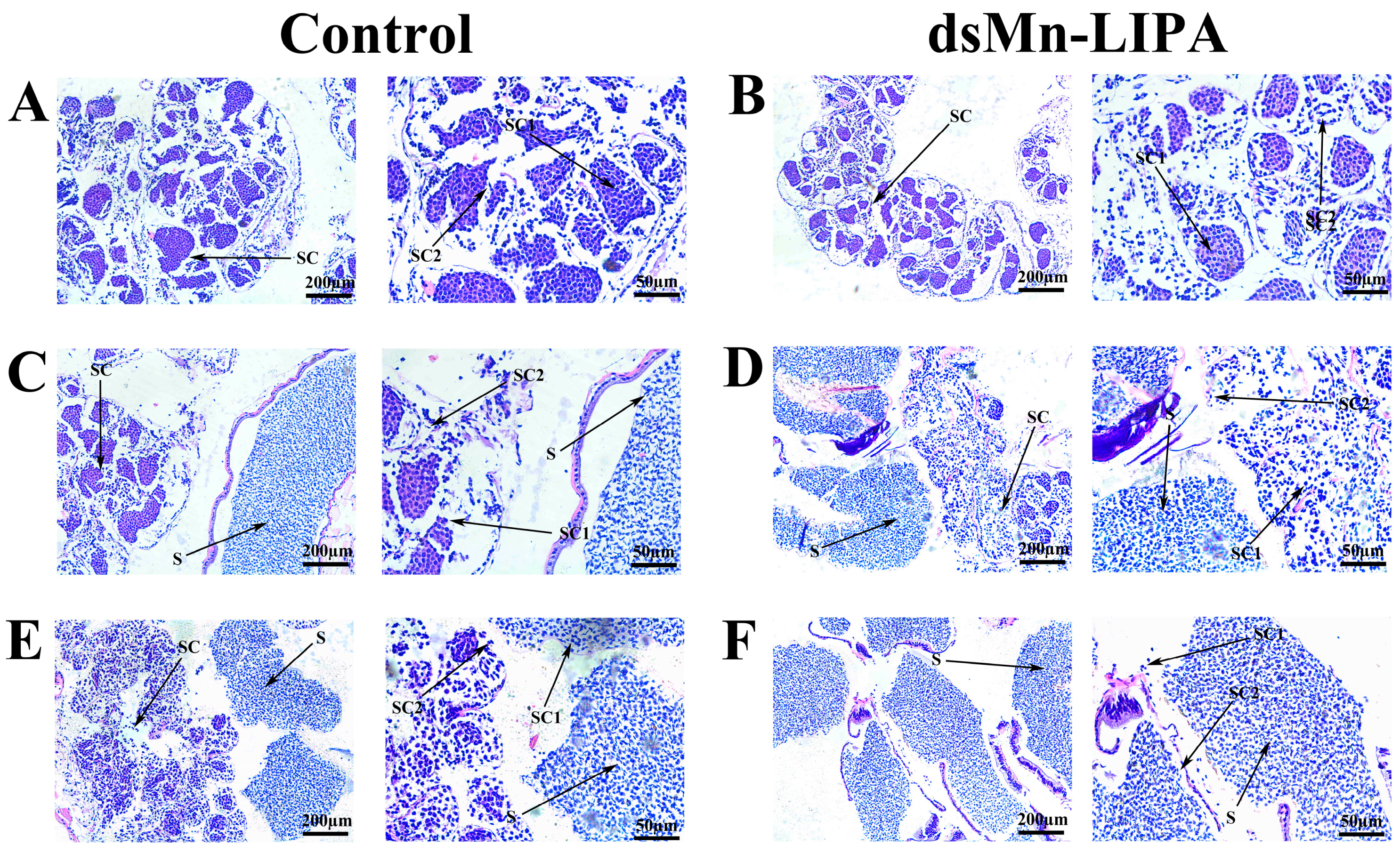
2.6.1. Histological Observations of Gonads
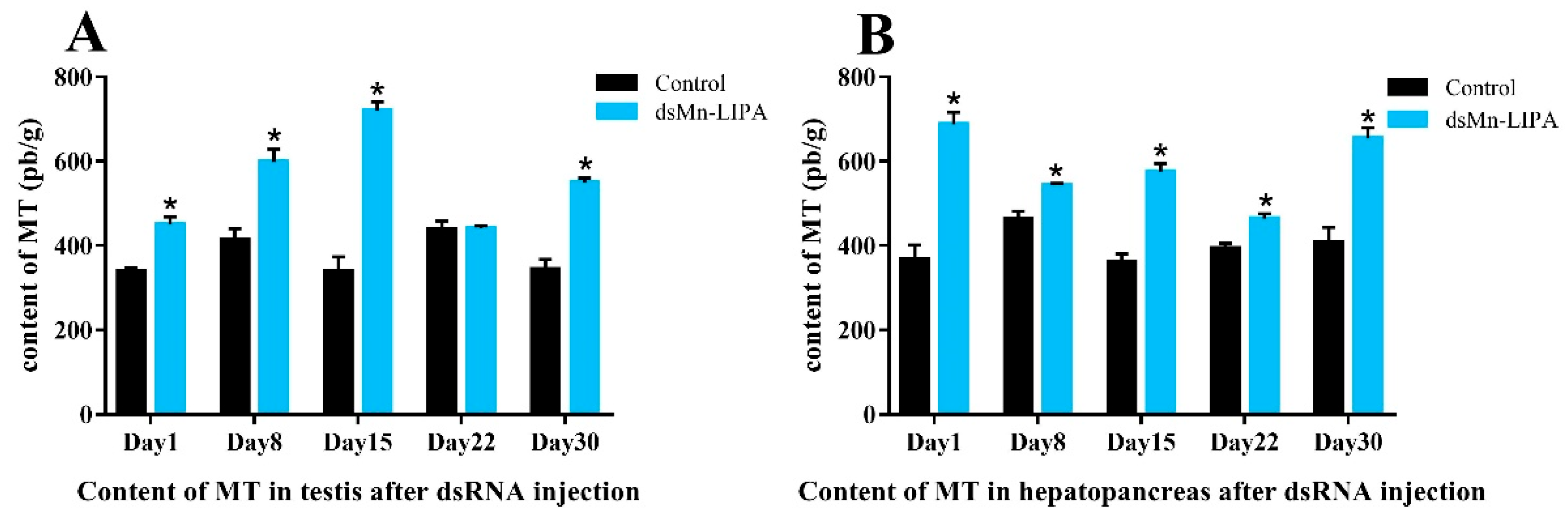
2.6.2. Steroid Hormone Content
2.7. Effects of Steroid Hormones and RNAi on Male Sex-Related Genes in Male M. nipponense
3. Discussion
4. Materials and Methods
4.1. Experimental Prawns
4.2. Nucleotide Sequencing and Bioinformatics Analysis
4.3. Tissue Expression Analysis by Quantitative Real-Time PCR
4.4. In Situ Hybridization (ISH) and Histological Observations
4.5. Sex Ratio Statistics
4.6. RNA Interference of LIPA
4.7. Enzyme Linked Immunosorbent Assay (ELISA)
4.8. Dietary Preparation
4.9. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, Y.; Ng, P. The freshwater palaemonid prawns (Crustacea: Decapoda: Caridea) of Myanmar. Hydrobiologia 2002, 487, 59–83. [Google Scholar] [CrossRef]
- Grave, S.; Ghane, A. The establishment of the Oriental River Prawn, Macrobrachium nipponense (de Haan, 1849) in Anzali Lagoon (Iran). Aquat. Invasions 2006, 1, 204–208. [Google Scholar] [CrossRef]
- Gul, I.; Abbas, M.; Kausar, S.; Luo, J.; Gao, X.; Mu, Y.; Fan, W.; Cui, H. Insight into crustacean cathepsins: Structure-evolutionary relationships and functional roles in physiological processes. Fish Shellfish Immunol. 2023, 139, 108852. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, Z.; Li, M.; Jiang, Q.; Wu, D.; Huang, Y.; Jiao, Y.; Zhang, M.; Zhao, Y. Effects of nanoplastics on antioxidant and immune enzyme activities and related gene expression in juvenile Macrobrachium nipponense. J. Hazard. Mater. 2020, 398, 122990. [Google Scholar] [CrossRef] [PubMed]
- Ekwudo, M.; Malek, M.; Anderson, C.; Yampolsky, L. The interplay between prior selection, mild intermittent exposure, and acute severe exposure in phenotypic and transcriptional response to hypoxia. Ecol. Evol. 2022, 12, e9319. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Zhang, W.; Xiong, Y.; Fu, H. Recent progress of male sexual differentiation and development in the oriental river prawn (Macrobrachium nipponense): A review. Rev. Aquac. 2023, 15, 305–317. [Google Scholar] [CrossRef]
- Xiong, Y.; Wang, S.; Gui, J.; Mei, J. Artificially induced sex-reversal leads to transition from genetic to temperature-dependent sex determination in fish species. Sci. China Life Sci. 2020, 63, 157–159. [Google Scholar] [CrossRef]
- Morris, G.; Braund, P.; Moore, J.; Samani, N.; Codd, V.; Webb, T. Coronary artery disease–associated LIPA coding variant rs1051338 reduces lysosomal acid lipase levels and activity in lysosomes. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1050–1057. [Google Scholar] [CrossRef]
- Ameis, D.; Merkel, M.; Eckerskorn, C.; Greten, H. Purification, characterization and molecular cloning of human hepatic lysosomal acid lipase. Eur. J. Biochem. 1994, 219, 905–914. [Google Scholar] [CrossRef]
- Zhang, H. Lysosomal acid lipase and lipid metabolism: New mechanisms, new questions, and new therapies. Curr. Opin. Lipidol. 2018, 29, 218. [Google Scholar] [CrossRef]
- Sequencing, R.; Consortium, A. The Rhesus macaque genome sequence informs biomedical and evolutionary analyses. Science 2007, 316, 222–234. [Google Scholar] [CrossRef]
- Holmes, R.; Cox, L.; VandeBerg, J. Comparative studies of mammalian acid lipases: Evidence for a new gene family in mouse and rat (Lipo). Comp. Biochem. Physiol. Part D Genom. Proteom. 2010, 5, 217–226. [Google Scholar] [CrossRef] [PubMed]
- De, P.; Osoegawa, K.; Zhu, B.; Marra, M.; Schein, J.; Bosdet, I.; Fjell, C. Genome sequence of the Brown Norway rat yields insights into mammalian evolution. Nature 2004, 428, 493–521. [Google Scholar] [CrossRef]
- Hillier, W. International chicken genome sequencing consortium. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Holmes, R.; VandeBerg, J.; Cox, L. Genomics and proteomics of vertebrate cholesterol ester lipase (LIPA) and cholesterol 25-hydroxylase (CH25H). 3 Biotech 2011, 1, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Lam, P.; Ashbrook, A.; Zygmunt, D.; Yan, C.; Du, H.; Martin, P. Therapeutic efficacy of rscAAVrh74. miniCMV. LIPA gene therapy in a mouse model of lysosomal acid lipase deficiency. Mol. Ther. Methods Clin. Dev. 2022, 26, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Pasta, A.; Borro, P.; Cremonini, A.; Formisano, E.; Tozzi, G.; Cecchi, S.; Fresa, R.; Labanca, S.; Djahandideh, A.; Sukkar, S. Effect of a common missense variant in LIPA gene on fatty liver disease and lipid phenotype: New perspectives from a single-center observational study. Pharmacol. Res. Perspect. 2021, 9, e00820. [Google Scholar] [CrossRef]
- Hou, Z.; Sket, B. A review of Gammaridae (Crustacea: Amphipoda): The family extent, its evolutionary history, and taxonomic redefinition of genera. Zool. J. Linn. Soc. 2016, 176, 323–348. [Google Scholar] [CrossRef]
- Uawisetwathana, U.; Leelatanawit, R.; Klanchui, A.; Prommoon, J.; Klinbunga, S.; Karoonuthaisiri, N. Insights into eyestalk ablation mechanism to induce ovarian maturation in the black tiger shrimp. PLoS ONE 2011, 6, e24427. [Google Scholar] [CrossRef]
- Liang, S.; Zeng, C.; Liang, J.; Luo, X.; Gao, X.; Ke, C.; Li, Y.; Chen, L.; Liu, H.; Guo, Y. Effects of eyestalk ablation on growth, feeding behavior, movement and circadian gene expression in Haliotis discus hannai. Aquac. Rep. 2023, 33, 101807. [Google Scholar] [CrossRef]
- Cai, P.; Yuan, H.; Gao, Z.; Qiao, H.; Zhang, W.; Jiang, S.; Xiong, Y.; Gong, Y.; Wu, Y.; Jin, S.; et al. 17β-Estradiol Induced Sex Reversal and Gonadal Transcriptome Analysis in the Oriental River Prawn (Macrobrachium nipponense): Mechanisms, Pathways, and Potential Harm. Int. J. Mol. Sci. 2023, 24, 8481. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.; Cronan, J. Lipoic acid metabolism in Escherichia coli: Sequencing and functional characterization of the lipA and lipB genes. J. Bacteriol. 1993, 175, 1325–1336. [Google Scholar] [CrossRef]
- Jin, L.; Lloyd, R.V. In situ hybridization: Methods and applications. J. Clin. Lab. Anal. 1997, 11, 2–9. [Google Scholar] [CrossRef]
- Norbeck, L.; Sheridan, M. An in vitro model for evaluating peripheral regulation of growth in fish: Effects of 17β-estradiol and testosterone on the expression of growth hormone receptors, insulin-like growth factors, and insulin-like growth factor type 1 receptors in rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol. 2011, 173, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Abo-Al-Ela, H.G. Hormones and fish monosex farming: A spotlight on immunity. Fish Shellfish Immunol. 2018, 72, 23–30. [Google Scholar] [CrossRef]
- Li, F.; Zhang, H. Lysosomal acid lipase in lipid metabolism and beyond. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 850–856. [Google Scholar] [CrossRef]
- Mondy, N.; Grossi, V.; Cathalan, E.; Delbecque, J.; Mermillod, F.; Douady, C. Sterols and steroids in a freshwater crustacean (Proasellus meridianus): Hormonal response to nutritional input. Invertebr. Biol. 2014, 133, 99–107. [Google Scholar] [CrossRef]
- Vogt, G. Functional cytology of the hepatopancreas of decapod crustaceans. J. Morphol. 2019, 280, 1405–1444. [Google Scholar] [CrossRef]
- Karplus, I.; Sagi, A.; Khalaila, I.; Barki, A. The Influence of Androgenic Gland Implantation on the Agonistic Behavior of Female Crayfish (Cherax quadricarinatus) in Interactions with Males. Behaviour 2003, 140, 649–663. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, Q.; Chen, H.; Zhu, X.; Cui, Z.; Qiu, G. The morphological and histological observation of embryonic development in the oriental river prawn Macrobrachium nipponense. J. Shanghai Ocean Univ. 2012, 21, 33–40. [Google Scholar]
- Tautz, D.; Pfeifle, C. A non-radioactive in situ hybridization method for the localization of specific RNAs in Drosophila embryos reveals translational control of the segmentation gene hunchback. Chromosoma 1989, 98, 81–85. [Google Scholar] [CrossRef]
- Farhadi, A.; Cui, W.; Zheng, H.; Li, S.; Zhang, Y.; Ikhwanuddin, M.; Ma, H. The regulatory mechanism of sexual development in decapod crustaceans. Front. Mar. Sci. 2021, 8, 679687. [Google Scholar] [CrossRef]
- Das, S.; Durica, D. Ecdysteroid receptor signaling disruption obstructs blastemal cell proliferation during limb regeneration in the fiddler crab, Uca pugilator. Mol. Cell. Endocrinol. 2013, 365, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zhou, Y.; Liu, M.; Tao, T.; Jiang, Q.; Zhu, D. The nuclear receptor E75 from the swimming crab, Portunus trituberculatus: cDNA cloning, transcriptional analysis, and putative roles on expression of ecdysteroid-related genes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2016, 200, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhu, X.; Li, Z.; Xu, W.; Dong, J.; Wei, H.; Li, Y.; Li, X. Genetic diversity and structure of Chinese grass shrimp, Palaemonetes sinensis, inferred from transcriptome-derived microsatellite markers. BMC Genet. 2019, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B. Molecular and evolutionary dynamics of animal sex-chromosome turnover. Nat. Ecol. Evol. 2019, 3, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Toyota, K.; Miyakawa, H.; Hiruta, C.; Sato, T.; Katayama, H.; Ohira, T.; Iguchi, T. Sex determination and differentiation in decapod and cladoceran crustaceans: An overview of endocrine regulation. Genes 2021, 12, 305. [Google Scholar] [CrossRef]
- Zheng, S.; Tao, W.; Yang, H.; Kocher, T.D.; Wang, Z.; Peng, Z.; Jin, L.; Pu, D.; Zhang, Y.; Wang, D. Identification of sex chromosome and sex-determining gene of southern catfish (Silurus meridionalis) based on XX, XY and YY genome sequencing. Proc. R. Soc. B 2022, 289, 20212645. [Google Scholar] [CrossRef]
- Tan, K.; Jiang, H.; Jiang, D.; Wang, W. Sex reversal and the androgenic gland (AG) in Macrobrachium rosenbergii: A review. Aquac. Fish. 2020, 5, 283–288. [Google Scholar] [CrossRef]
- Amir, S.; Eviatar, S.; Isam, K. Sexual differentiation in decapod crustaceans: Role of the androgenic gland. Int. J. Invertebr. Reprod. 1997, 31, 55–61. [Google Scholar] [CrossRef]
- Ventura, T.; Sagi, A. The insulin-like androgenic gland hormone in crustaceans: From a single gene silencing to a wide array of sexual manipulation-based biotechnologies. Biotechnol. Adv. 2012, 30, 1543–1550. [Google Scholar] [CrossRef] [PubMed]
- Maya, B.; Debby, I.; Haim, B. Regulation of sperm motility by PIP2(4,5) and actin polymerization. Dev. Biol. 2013, 381, 62–72. [Google Scholar] [CrossRef]
- Breitbart, H.; Etkovitz, N. Role and regulation of EGFR in actin remodeling in sperm capacitation and the acrosome reaction. Asian J. Androl. 2011, 13, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Hamada, D.; Kamikubo, H.; Hirata, K.; Kataoka, M.; Yamamoto, M.; Ikawa, M.; Okabe, M.; Hagihara, Y. Molecular dissection of IZUMO1, a sperm protein essential for sperm-egg fusion. Development 2013, 140, 3221–3229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zarkower, D. DMRT proteins and coordination of mammalian spermatogenesis. Stem Cell Res. 2017, 24, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Abayed, F.; Manor, R.; Aflalo, E.; Sagi, A. Screening for Dmrt genes from embryo to mature Macrobrachium rosenbergii prawns. Gen. Comp. Endocrinol. 2019, 282, 113205. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Manor, R.; Aflalo, E.; Weil, S.; Raviv, S.; Glazer, L.; Sagi, A. Temporal silencing of an androgenic gland-specific insulin-like gene affecting phenotypical gender differences and spermatogenesis. Endocrinology 2009, 150, 1278–1286. [Google Scholar] [CrossRef]
- Xu, H.; Chen, Y.; Wang, Y.; Luo, J.; Li, J.; Shen, S.; Yang, J.; Ma, W. Full Functional Sex Reversal Achieved Through Silencing of MroDmrt11E Gene in Macrobrachium rosenbergii: Production of All-Male Monosex Freshwater Prawn. Front. Endocrinol. 2022, 12, 1867. [Google Scholar] [CrossRef]
- Wei, W.; Huang, J.; Yang, Q.; Zhou, F.; Jiang, S.; Li, Y.; Jiang, S.; Yang, L. Molecular characterization and functional analysis of DMRT11E in black tiger shrimp (Penaeus monodon). Aquac. Rep. 2022, 22, 100982. [Google Scholar] [CrossRef]
- Cai, P.; Yuan, H.; Gao, Z.; Daka, P.; Qiao, H.; Zhang, W.; Jiang, S.; Xiong, Y.; Gong, Y.; Wu, Y.; et al. Sex Reversal Induced by Dietary Supplementation with 17α-Methyltestosterone during the Critical Period of Sex Differentiation in Oriental River Prawn (Macrobrachium nipponense). Animals 2023, 13, 1369. [Google Scholar] [CrossRef]
- Zhu, Q.; Han, C.; Liu, S.; Ouyang, H.; Liu, D.; Zhang, Z.; Huang, J.; Han, L.; Li, S.; Li, G.; et al. Development and gene expression analysis of gonad during 17α-methyltestosterone-induced sex reversal in mandarin fish (Siniperca chuatsi). Aquac. Rep. 2022, 23, 101049. [Google Scholar] [CrossRef]
- Liang, F.; Sun, C.; Li, S.; Hou, T.; Li, C. Therapeutic effect and immune mechanism of chitosan-gentamicin conjugate on Pacific white shrimp (Litopenaeus vannamei) infected with Vibrio parahaemolyticus. Carbohydr. Polym. 2021, 269, 118334. [Google Scholar] [CrossRef] [PubMed]
- Greisy, Z.; Gamal, A. Monosex production of tilapia, Oreochromis niloticus using different doses of 17α-methyltestosterone with respect to the degree of sex stability after one year of treatment. Egypt. J. Aquat. Res. 2012, 38, 59–66. [Google Scholar] [CrossRef]
- Huang, J.; Liu, S.; Ouyang, H.; Han, C.; Li, M.; Han, L.; Li, S.; Li, G.; Lin, H.; Zhang, Y. Production of XX neo-male mandarin fish, Siniperca chuatsi, after sexual differentiation by oral administration of 17α-methyltestosterone. Water Biol. Secur. 2023, 2, 100095. [Google Scholar] [CrossRef]
- Voorhees, J.; Mamer, E.; Schill, D.; Adams, M.; Martinez, C.; Barnes, M. 17β-Estradiol can induce sex reversal in Brown Trout. Fishes 2023, 8, 103. [Google Scholar] [CrossRef]
- Black, D.; Minkin, M.; Graham, S.; Bernick, B.; Mirkin, S. Effects of combined 17β-estradiol and progesterone on weight and blood pressure in postmenopausal women of the REPLENISH trial. Menopause 2021, 28, 32. [Google Scholar] [CrossRef]
- Du, Y.; Ma, K.; Qiu, G. Discovery of the genes in putative GnRH signaling pathway with focus on characterization of GnRH-like receptor transcripts in the brain and ovary of the oriental river prawn Macrobrachium nipponense. Aquaculture 2015, 442, 1–11. [Google Scholar] [CrossRef]
- Liu, Z.; Xiao, J.; Xia, Y.; Wu, Q.; Zhao, C.; Li, D. Selection and validation of reference genes for RT-qPCR-based analyses of Anastatus japonicus Ashmead (Hymenoptera: Helicopteridae). Front. Physiol. 2022, 13, 1046204. [Google Scholar] [CrossRef]
- Andreani, J.; Le, M.; Duflot, I.; Jardot, P.; Rolland, C.; Boxberger, M.; Wurtz, N.; Rolain, J.; Colson, P.; La, B. In vitro testing of combined hydroxychloroquine and azithromycin on SARS-CoV-2 shows synergistic effect. Microb. Pathog. 2020, 145, 104228. [Google Scholar] [CrossRef]
- Sato, K.; Ohuchi, H. The Opsin 3/Teleost multiple tissue opsin system: mRNA localization in the retina and brain of medaka (Oryzias latipes). J. Comp. Neurol. 2021, 529, 2484–2516. [Google Scholar] [CrossRef]
- Faeed, M.; Kasra, R.; PourKazemi, M.; Darboee, M.; Haghighi, S. Study on effect feedings with probiotics in increasing resistance to Aeromonas hydrophila and Changes in gut bacterial communities Sander lucioperca. Biol. J. Microorg. 2018, 7, 1–12. [Google Scholar] [CrossRef]
- Hodar, A.; Vasava, R.; Mahavadiya, D.; Joshi, N. Fish meal and fish oil replacement for aqua feed formulation by using alternative sources: A review. J. Exp. Zool. India 2020, 23, 13–21. [Google Scholar] [CrossRef]
- Saleh, S.; Widodo, M.; Kilawati, Y. The Effect of 17α-Methyltestosterone Hormone Application on Masculinization and Growth Rate of Vaname Shrimp Larvae (Litopenaeus Vannamei, Boone 1931). Russ. J. Agric. Socio-Econ. Sci. 2018, 79, 352–356. [Google Scholar] [CrossRef]
- Rosales, M.; Castillo, S.; Pohlenz, C.; Gatlin, D. Evaluation of dried yeast and threonine fermentation biomass as partial fish meal replacements in the diet of red drum Sciaenops ocellatus. Anim. Feed Sci. Technol. 2017, 232, 190–197. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′→3′) | Usage |
|---|---|---|
| LIPA-F1 | ACTCATTTCTTTGTGGGACCCTT | ORF |
| LIPA-R1 | GAAATTCCTTGGGCAAAGTGGAA | ORF |
| LIPA-F2 | CGTAGCAACAGTTACAAGCATCA | ORF |
| LIPA-R2 | AAGGGTCCCACAAAGAAATGAGT | ORF |
| LIPA-F3 | ACTCAAACACTCTTCCACTTTGC | ORF |
| LIPA-R3 | ATAGACCAGAGGCTTTGCATCAG | ORF |
| Mn-LIPA-F | ACCGCCAGAATATGATGTTGCTA | qPCR |
| Mn-LIPA-R | AAAGGAAGTCCAGGTGGCTAAAT | qPCR |
| Mn-IAG-F | CGCCTCCGTCTGCCTGAGATAC | qPCR |
| Mn-IAG-R | CCTCCTCCTCCACCTTCAATGC | qPCR |
| Mn-SG-F | ACCCTAGCCCCAGTACGTGTT | qPCR |
| Mn-SG-R | AGAGGTGGTGAAGCTGTCTCTCA | qPCR |
| Mn-Dmrt11E-F | ACGACCTTAGTAGGATGGACAGT | qPCR |
| Mn-Dmrt11E-R | GAGTGGAGGCAATAGAATGGGTA | qPCR |
| Mn-EIF-F | CATGGATGTACCTGTGGTGAAAC | qPCR |
| Mn-EIF-R | CTGTCAGCAGAAGGTCCTCATTA | qPCR |
| dsMn-LIPA-F | TAATACGACTCACTATAGGGACTGACCCATAACCCTGCTG | dsRNA |
| dsMn-LIPA-R | TAATACGACTCACTATAGGGTCAGTTGCCCAAAGGAAGTC | dsRNA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, P.; Zhang, W.; Jiang, S.; Xiong, Y.; Qiao, H.; Yuan, H.; Gao, Z.; Zhou, Y.; Jin, S.; Fu, H. Role of Mn-LIPA in Sex Hormone Regulation and Gonadal Development in the Oriental River Prawn, Macrobrachium nipponense. Int. J. Mol. Sci. 2024, 25, 1399. https://doi.org/10.3390/ijms25031399
Cai P, Zhang W, Jiang S, Xiong Y, Qiao H, Yuan H, Gao Z, Zhou Y, Jin S, Fu H. Role of Mn-LIPA in Sex Hormone Regulation and Gonadal Development in the Oriental River Prawn, Macrobrachium nipponense. International Journal of Molecular Sciences. 2024; 25(3):1399. https://doi.org/10.3390/ijms25031399
Chicago/Turabian StyleCai, Pengfei, Wenyi Zhang, Sufei Jiang, Yiwei Xiong, Hui Qiao, Huwei Yuan, Zijian Gao, Yongkang Zhou, Shubo Jin, and Hongtuo Fu. 2024. "Role of Mn-LIPA in Sex Hormone Regulation and Gonadal Development in the Oriental River Prawn, Macrobrachium nipponense" International Journal of Molecular Sciences 25, no. 3: 1399. https://doi.org/10.3390/ijms25031399