
Svx Peptidases of Phytopathogenic Pectolytic Bacteria: Structural, Catalytic and Phytoimmune Properties
 , , , , and
, , , , and 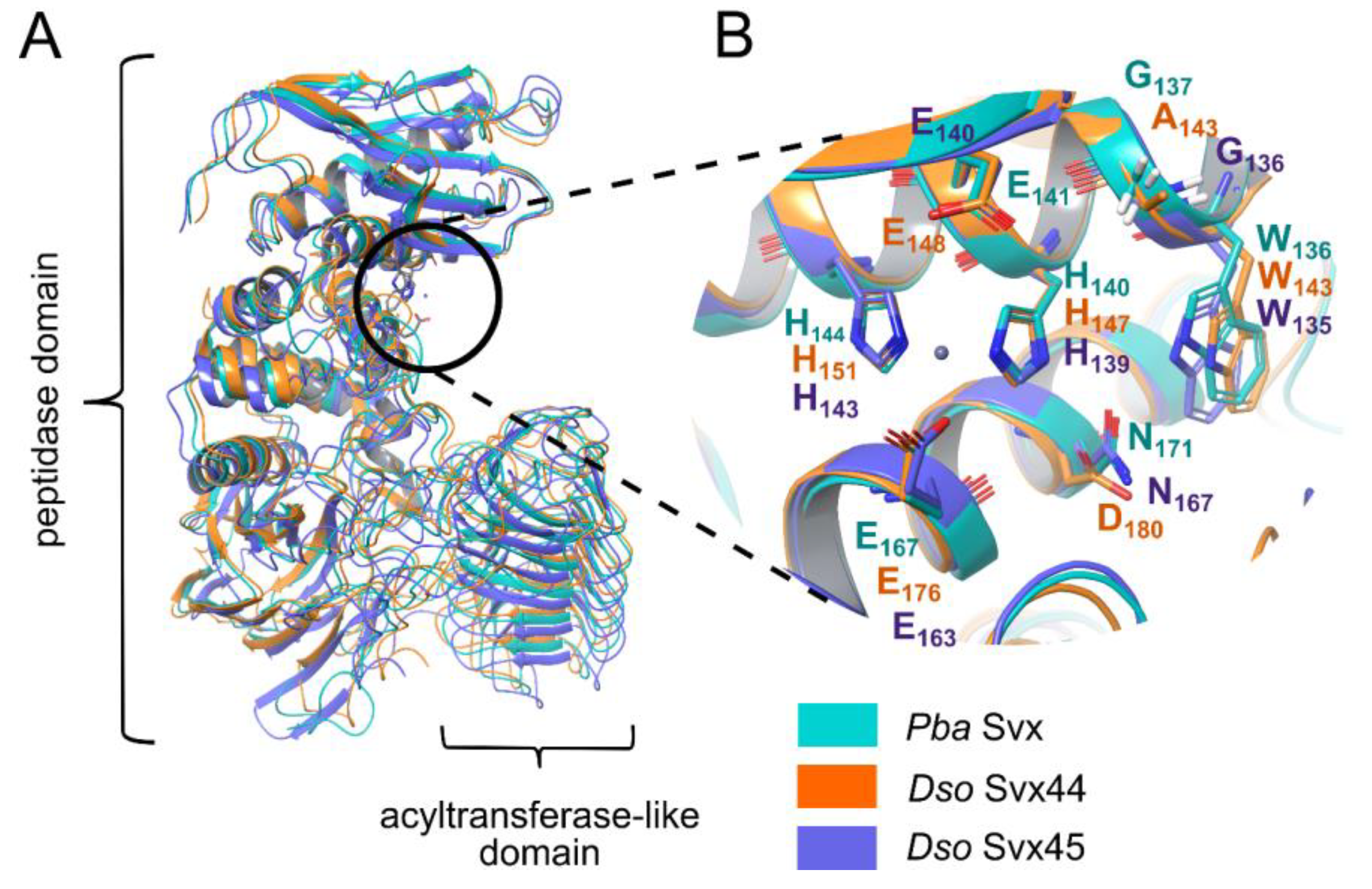{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
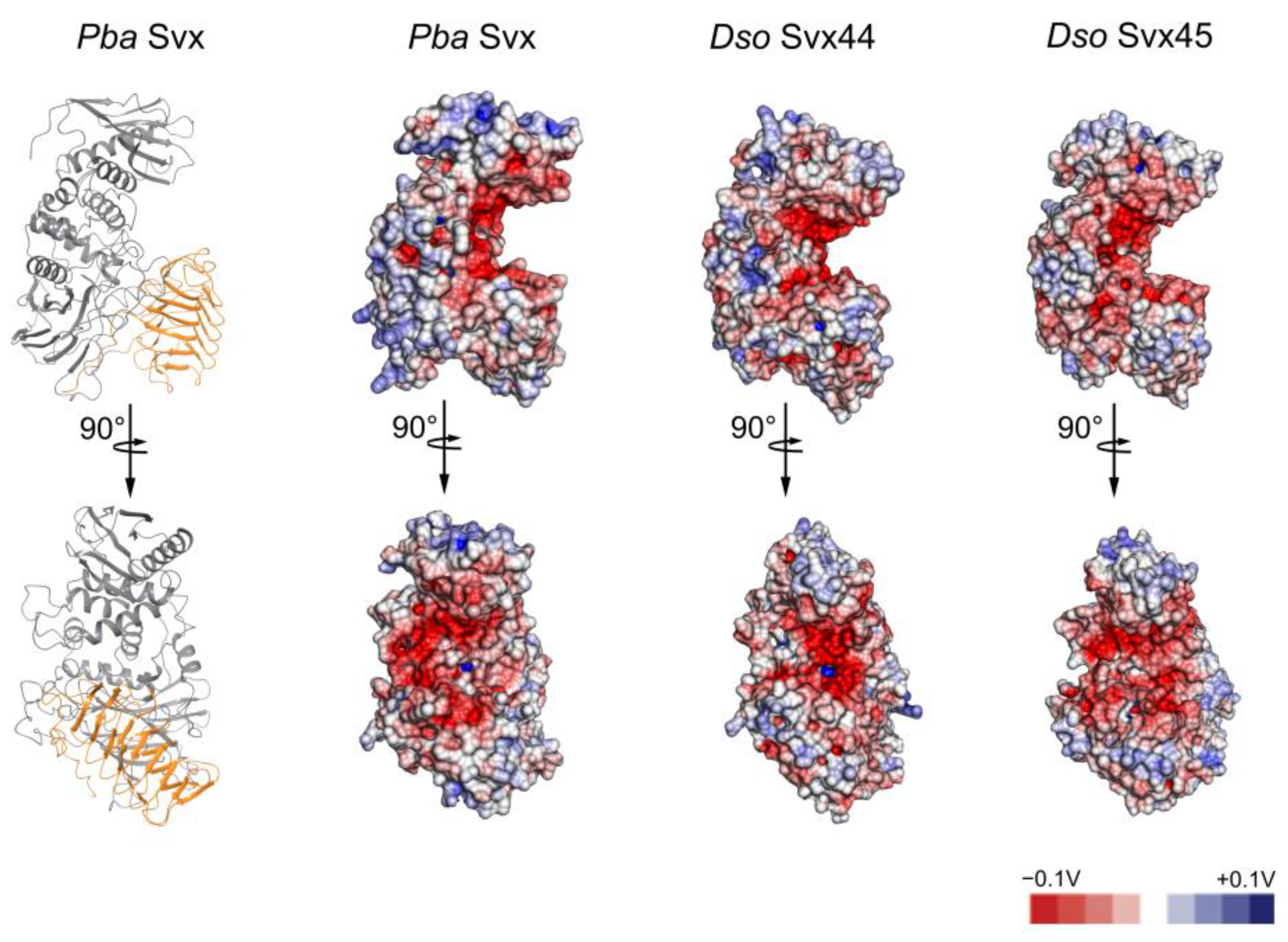
2.1. Comparative Analysis of the Structure of the Svx-like Proteins of Pba and Dso
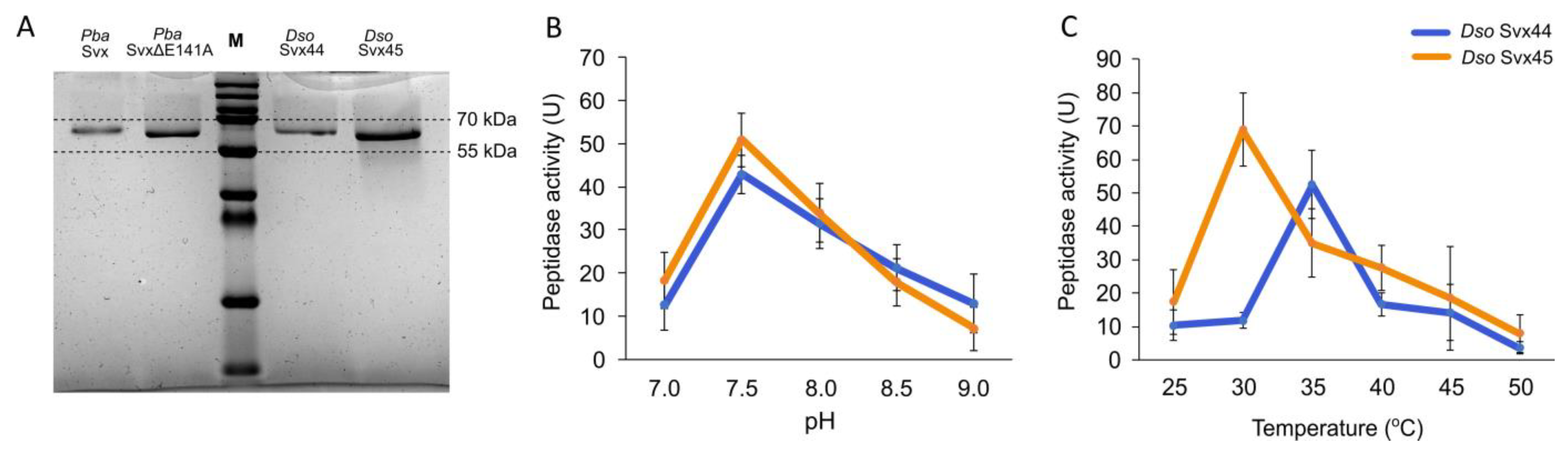
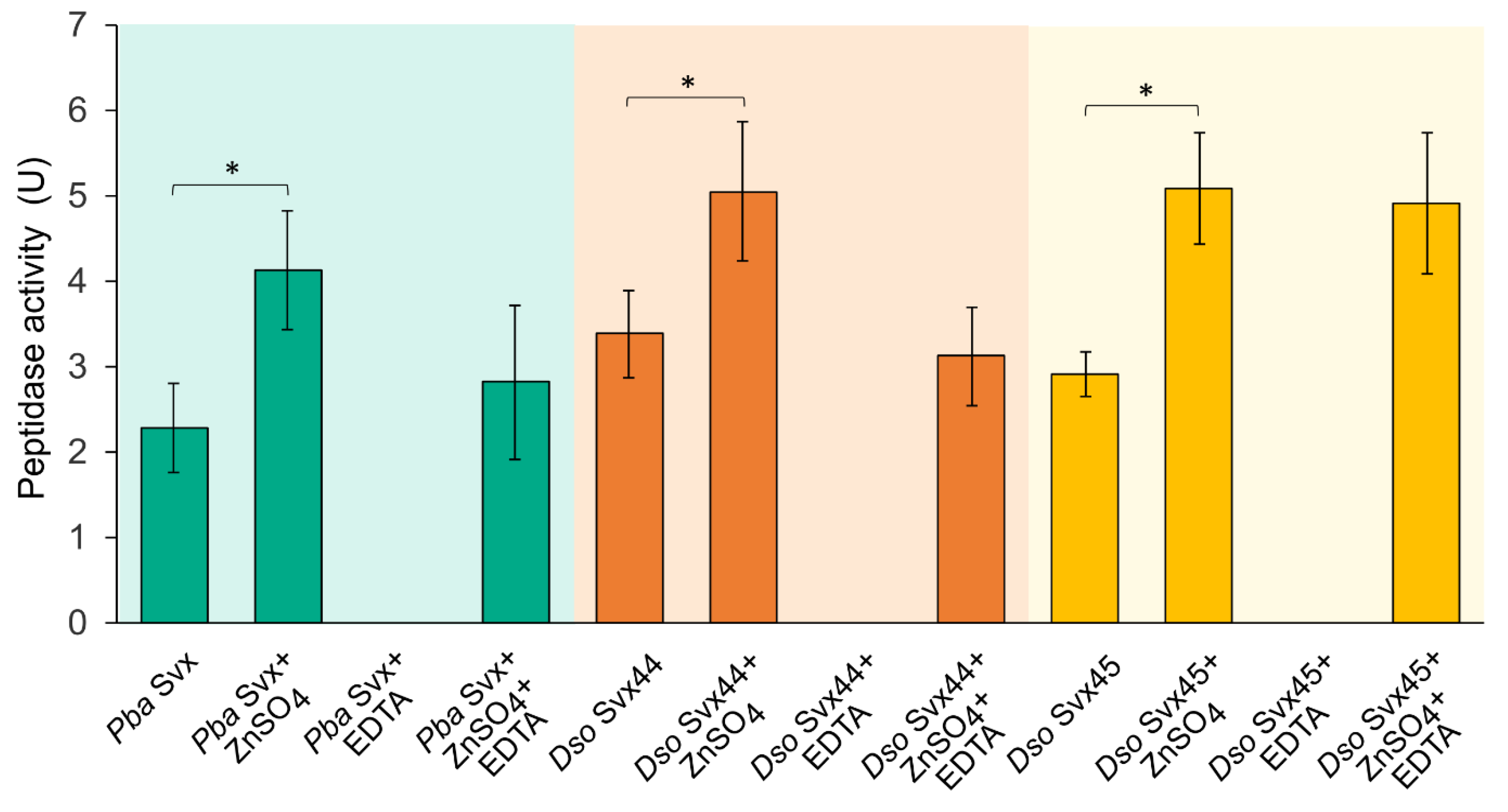
2.2. Catalytic Properties of Dso Svx-like Proteins
2.3. Phytoimmune Properties of Svx Proteins of Pba and Dso
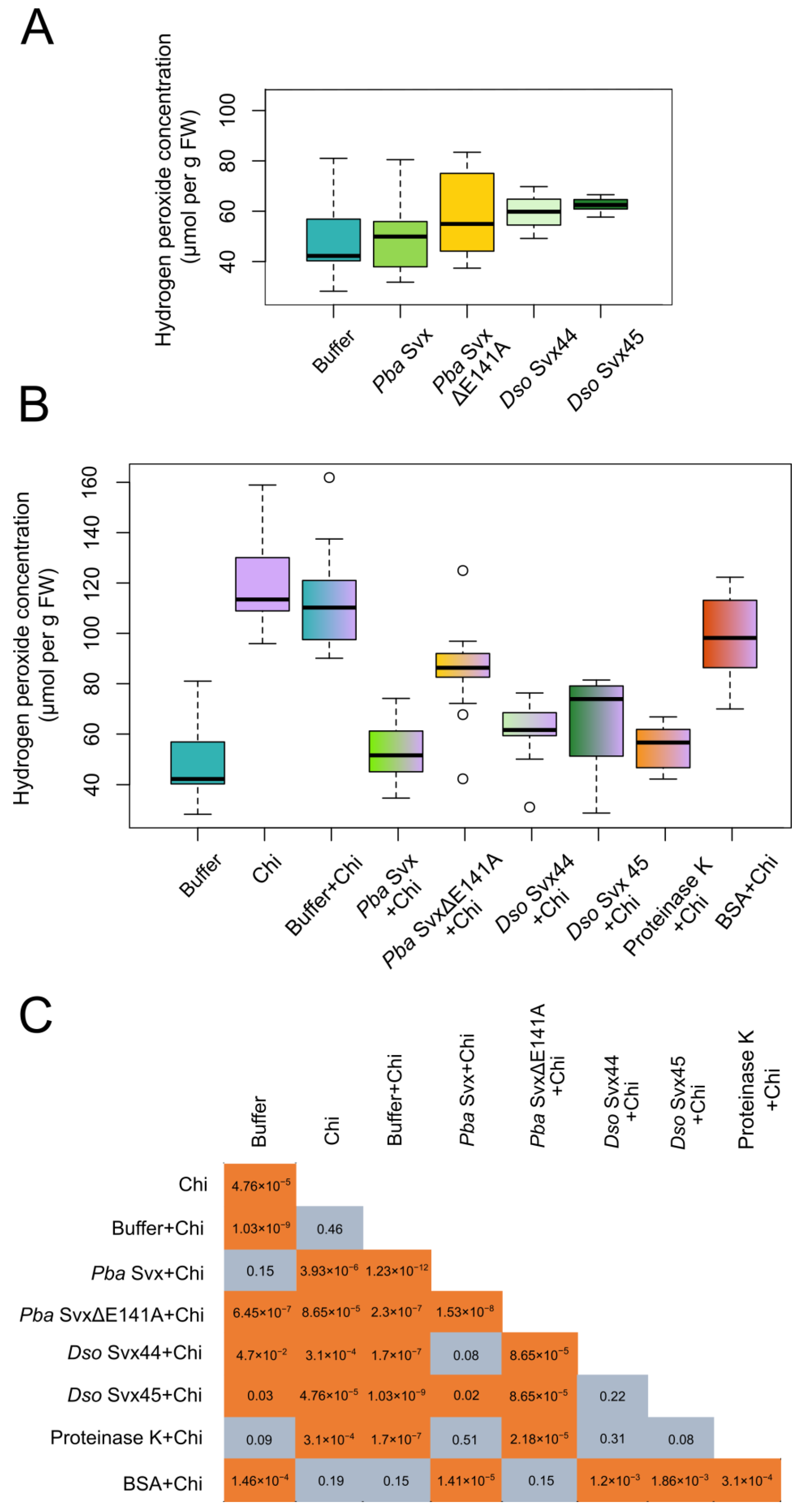
2.3.1. Effect of the Svx Proteins on Hydrogen Peroxide Levels in Plant Tissues
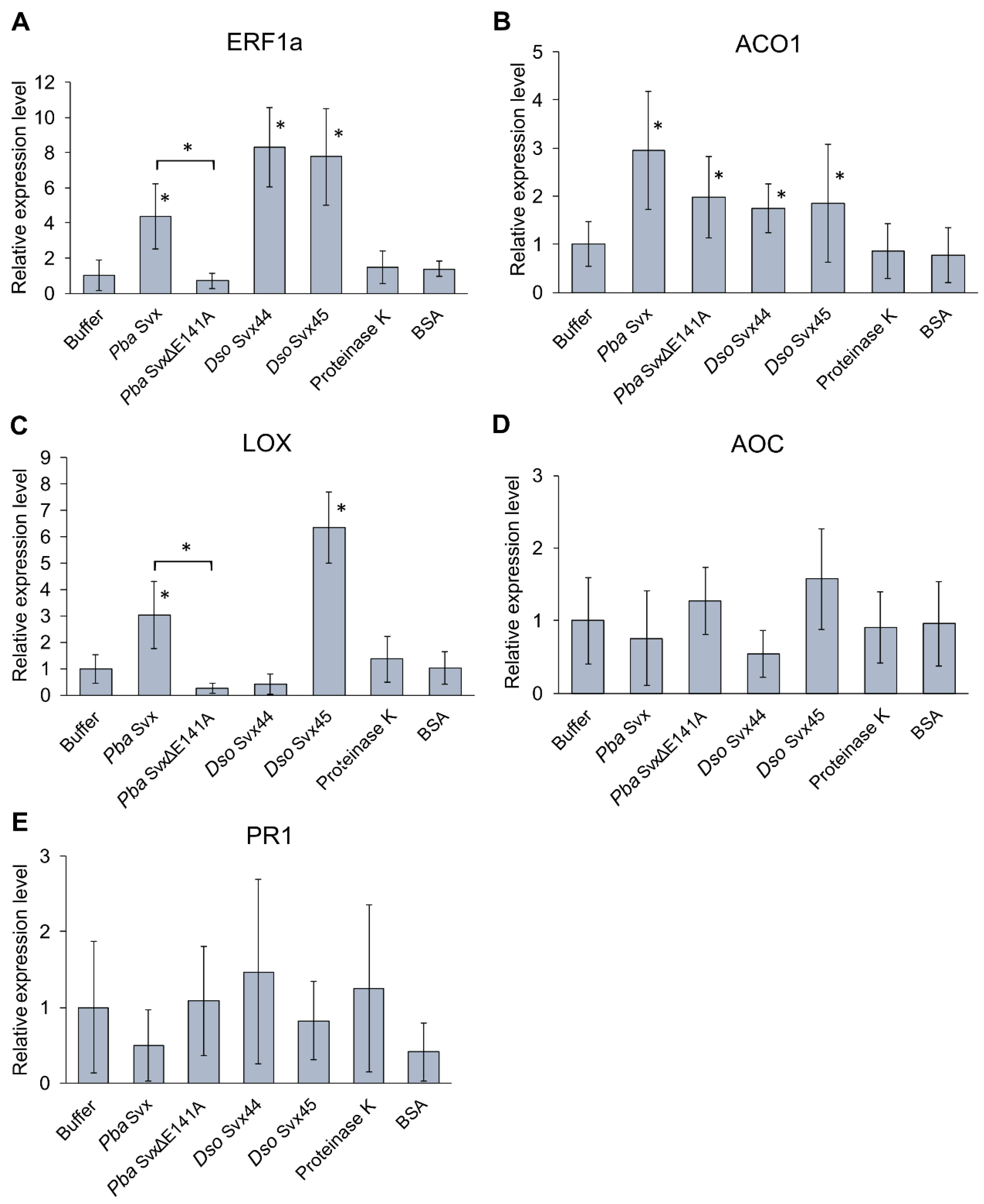
2.3.2. Effect of Svx Proteins on the Expression of Infection-Related Genes
3. Discussion
4. Materials and Methods
4.1. In Silico Analysis of the Protein Structure
4.2. Molecular Dynamics Simulations
4.3. Gene Cloning and Protein Purification
4.4. Peptidase Activity Assays
4.5. Determination of the Hydrogen Peroxide Level
4.6. Gene Expression Analysis Using qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van der Wolf, J.; Acuña, I.; De Boer, S.; Brurberg, M.; Cahill, G.; Charkowski, A.; Coutinho, T.; Davey, T.; Dees, M.; Degefu, Y.; et al. Diseases Caused by Pectobacterium and Dickeya Species Around the World. In Plant Diseases Caused by Dickeya and Pectobacterium Species, 1st ed.; Van Gijsegem, F., van der Wolf, J., Toth, I.K., Eds.; Springer: Cham, Switzerland, 2021; pp. 215–261. [Google Scholar] [CrossRef]
- Charkowski, A.; Blanco, C.; Condemine, G.; Expert, D.; Franza, T.; Hayes, C.; Hugouvieux-Cotte-Pattat, N.; López Solanilla, E.; Low, D.; Moleleki, L.; et al. The role of secretion systems and small molecules in soft-rot Enterobacteriaceae pathogenicity. Annu. Rev. Phytopathol. 2012, 50, 425–449. [Google Scholar] [CrossRef] [PubMed]
- Toth, I.K.; Birch, P.R. Rotting softly and stealthily. Curr. Opin. Plant Biol. 2005, 8, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Van Gijsegem, F.; Hugouvieux-Cotte-Pattat, N.; Kraepiel, Y.; Lojkowska, E.; Moleleki, L.; Gorshkov, V.; Yedidia, I. Molecular Interactions of Pectobacterium and Dickeya with Plants. In Plant Diseases Caused by Dickeya and Pectobacterium Species, 1st ed.; Van Gijsegem, F., van der Wolf, J., Toth, I.K., Eds.; Springer: Cham, Switzerland, 2021; pp. 85–147. [Google Scholar] [CrossRef]
- Gorshkov, V.; Parfirova, O. Host plant physiological transformation and microbial population heterogeneity as important determinants of the Soft Rot Pectobacteriaceae–plant interactions. Semin. Cell Dev. Biol. 2023, 148, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Lapwood, D.H.; Harris, R.I. The spread of Erwinia carotovora subsp. atroseptica and subsp. carotovora from stem lesions and degenerating seed tubers to progeny tubers in soil. Potato Res. 1982, 25, 41–50. [Google Scholar] [CrossRef]
- Hélias, V.; Andrivon, D.; Jouan, B. Development of symptoms caused by Erwinia carotovora ssp. atroseptica under field conditions and their effects on the yield of individual potato plants. Plant Pathol. 2000, 49, 23–32. [Google Scholar] [CrossRef]
- Pérombelon, M.C.M. Potato diseases caused by soft rot erwinias: An overview of pathogenesis. Plant Pathol. 2002, 51, 1–12. [Google Scholar] [CrossRef]
- Tsror, L.; Lebiush, S.; Erlich, O.; Ben-Daniel, B.; Van DerWolf, J. First report of latent infection of Cyperus rotundus caused by a biovar 3.(Syn. Erwinia chrysanthemi) in Israel. New Dis. Rep. 2010, 22, 2044-0588. [Google Scholar] [CrossRef]
- Tsror, L.; Lebiush, S.; Erlich, O.; Galilov, I.; Chalupowicz, L.; Reuven, M.; Dror, O.; Manulis-Sasson, S. First report of latent infection of Malva nicaeensis caused by Pectobacterium carotovorum subsp. brasiliense in Israel. New Dis. Rep. 2019, 39, 2044-0588. [Google Scholar] [CrossRef]
- Fikowicz-Krosko, J.; Czajkowski, R. Systemic colonization and expression of disease symptoms on bittersweet nightshade (Solanum dulcamara) infected with a GFP-tagged Dickeya solani IPO2222 (IPO2254). Plant Dis. 2018, 102, 619–627. [Google Scholar] [CrossRef]
- Ageichik, A.V.; Evtushenkov, A.N.; Nikolaichik, Y.A. The role of type III secretion system in Erwinia carotovora subsp. atroseptica virulence. Plant Protect. Sci. 2002, 38, 523–527. [Google Scholar] [CrossRef]
- Holeva, M.C.; Bell, K.S.; Hyman, L.J.; Avrova, A.O.; Whisson, S.C.; Birch, P.R.; Toth, I.K. Use of a pooled transposon mutation grid to demonstrate roles in disease development for Erwinia carotovora subsp. atroseptica putative type III secreted effector (DspE/a) and helper (HrpN) proteins. Mol. Plant Microbe Interact. 2004, 17, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Rojas, C.M.; Ham, J.H.; Deng, W.L.; Doyle, J.J.; Collmer, A. HecA, a member of a class of adhesins produced by diverse pathogenic bacteria, contributes to the attachment, aggregation, epidermal cell killing, and virulence phenotypes of Erwinia chrysanthemi EC16 on Nicotiana clevelandii seedlings. Proc. Natl. Acad. Sci. USA 2002, 99, 13142–13147. [Google Scholar] [CrossRef]
- Mattinen, L.; Nissinen, R.; Riipi, T.; Kalkkinen, N.; Pirhonen, M. Host-extract induced changes in the secretome of the plant pathogenic bacterium Pectobacterium atrosepticum. Proteomics 2007, 7, 3527–3537. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Coulthurst, S.J.; Pritchard, L.; Hedley, P.E.; Ravensdale, M.; Humphris, S.; Burr, T.; Takle, G.; Brurberg, M.B.; Birch, P.R.J.; et al. Quorum sensing coordinates brute force and stealth modes of infection in the plant pathogen Pectobacterium atrosepticum. PLoS Pathog. 2008, 4, e1000093. [Google Scholar] [CrossRef] [PubMed]
- Gorshkov, V.; Gubaev, R.; Petrova, O.; Daminova, A.; Gogoleva, N.; Ageeva, M.; Gogolev, Y. Transcriptome profiling helps to identify potential and true molecular switches of stealth to brute force behavior in Pectobacterium atrosepticum during systemic colonization of tobacco plants. Eur. J. Plant Pathol. 2018, 152, 957–976. [Google Scholar] [CrossRef]
- Gorshkov, V.Y.; Toporkova, Y.Y.; Tsers, I.D.; Smirnova, E.O.; Ogorodnikova, A.V.; Gogoleva, N.E.; Gogolev, Y.V. Differential modulation of the lipoxygenase cascade during typical and latent Pectobacterium atrosepticum infections. Ann. Bot. 2022, 129, 271–286. [Google Scholar] [CrossRef]
- Tsers, I.; Parfirova, O.; Moruzhenkova, V.; Petrova, O.; Gogoleva, N.; Vorob’ev, V.; Gorshkov, V. A Switch from Latent to Typical Infection during Pectobacterium atrosepticum—Tobacco Interactions: Predicted and True Molecular Players. Int. J. Mol. Sci. 2023, 24, 13283. [Google Scholar] [CrossRef]
- Enard, C.; Diolez, A.; Expert, D. Systemic virulence of Erwinia chrysanthemi 3937 requires a functional iron assimilation system. J. Bacteriol. 1988, 170, 2419–2426. [Google Scholar] [CrossRef]
- Franza, T.; Mahé, B.; Expert, D. Erwinia chrysanthemi requires a second iron transport route dependent of the siderophore achromobactin for extracellular growth and plant infection. Mol. Microbiol. 2005, 55, 261–275. [Google Scholar] [CrossRef]
- Dellagi, A.; Segond, D.; Rigault, M.; Fagard, M.; Simon, C.; Saindrenan, P.; Expert, D. Microbial siderophores exert a subtle role in Arabidopsis during infection by manipulating the immune response and the iron status. Plant Physiol. 2009, 150, 1687–1696. [Google Scholar] [CrossRef]
- Gorshkov, V.; Parfirova, O.; Petrova, O.; Gogoleva, N.; Kovtunov, E.; Vorob’ev, V.; Gogolev, Y. The knockout of enterobactin-related gene in Pectobacterium atrosepticum results in reduced stress resistance and virulence towards the primed plants. Int. J. Mol. Sci. 2021, 22, 9594. [Google Scholar] [CrossRef] [PubMed]
- Islamov, B.; Petrova, O.; Mikshina, P.; Kadyirov, A.; Vorob’ev, V.; Gogolev, Y.; Gorshkov, V. The Role of Pectobacterium atrosepticum Exopolysaccharides in Plant-Pathogen Interactions. Int. J. Mol Sci. 2021, 22, 12781. [Google Scholar] [CrossRef] [PubMed]
- Corbett, M.; Virtue, S.; Bell, K.; Birch, P.; Burr, T.; Hyman, L.; Lilley, K.; Poock, S.; Toth, I.; Salmond, G. Identification of a new quorum-sensing-controlled virulence factor in Erwinia carotovora subsp. atroseptica secreted via the type II targeting pathway. Mol. Plant Microbe Interact. 2005, 18, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Tendiuk, N.; Konnova, T.; Petrova, O.; Osipova, E.; Mukhametzyanov, T.; Makshakova, O.; Gorshkov, V. Structure-Functional Characteristics of the Svx Protein-The Virulence Factor of the Phytopathogenic Bacterium Pectobacterium atrosepticum. Int. J. Mol. Sci. 2022, 23, 6914. [Google Scholar] [CrossRef] [PubMed]
- Cassab, G.I. Plant cell wall proteins. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 281–309. [Google Scholar] [CrossRef] [PubMed]
- Herger, A.; Dünser, K.; Kleine-Vehn, J.; Ringli, C. Leucine-rich repeat extensin proteins and their role in cell wall sensing. Curr. Biol. 2019, 29, R851–R858. [Google Scholar] [CrossRef]
- Dünser, K.; Gupta, S.; Herger, A.; Feraru, M.I.; Ringli, C.; Kleine-Vehn, J. Extracellular matrix sensing by FERONIA and Leucine-Rich Repeat Extensins controls vacuolar expansion during cellular elongation in Arabidopsis thaliana. EMBO J. 2019, 38, e100353. [Google Scholar] [CrossRef]
- Parker, J.E.; Barber, C.E.; Fan, M.J.; Daniels, M.J. Interaction of Xanthomonas campestris with Arabidopsis thaliana: Characterization of a gene from X. c. pv. raphani that confers avirulence to most A. thaliana accessions. Mol. Plant Microbe Interact. 1993, 6, 216–224. [Google Scholar] [CrossRef]
- Kazemi-Pour, N.; Condemine, G.; Hugouvieux-Cotte-Pattat, N. The secretome of the plant pathogenic bacterium Erwinia chrysanthemi. Proteomics 2004, 4, 3177–3186. [Google Scholar] [CrossRef]
- Xing, R.; Liu, Y.; Li, K.; Yu, H.; Liu, S.; Yang, Y.; Chen, X.; Li, P. Monomer composition of chitooligosaccharides obtained by different degradation methods and their effects on immunomodulatory activities. Carbohydr. Polym. 2017, 157, 1288–1297. [Google Scholar] [CrossRef]
- Cannon, M.C.; Terneus, K.; Hall, Q.; Tan, L.; Wang, Y.; Wegenhart, B.L.; Chen, L.; Lamport, D.T.; Chen, Y.; Kieliszewski, M.J. Self-assembly of the plant cell wall requires an extensin scaffold. Proc. Natl. Acad. Sci. USA 2008, 105, 2226–2231. [Google Scholar] [CrossRef] [PubMed]
- Nguema-Ona, E.; Vicré-Gibouin, M.; Gotté, M.; Plancot, B.; Lerouge, P.; Bardor, M.; Driouich, A. Cell wall O-glycoproteins and N-glycoproteins: Aspects of biosynthesis and function. Front. Plant Sci. 2014, 5, 499. [Google Scholar] [CrossRef] [PubMed]
- Kieliszewski, M.J.; Kamyab, A.; Leykam, J.F.; Lamport, D.T.A. A Histidine-Rich Extensin from Zea mays Is an Arabinogalactan Protein. Plant Physiol. 1992, 99, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.H.; Weizbauer, R.A.; Singamaneni, S.; Xu, F.; Genin, G.M.; Pickard, B.G. Structures formed by a cell membrane-associated arabinogalactan-protein on graphite or mica alone and with Yariv phenylglycosides. Ann. Bot. 2014, 114, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Verdugo-Perales, M.; Velez-de la Rocha, R.; León-Félix, J.; Osuna-Enciso, T.; Heredia, J.B.; Osuna-Castro, J.A.; Islas-Osuna, M.A.; Sañudo-Barajas, J.A. Gene expression of an arabinogalactan lysine-rich protein CaAGP18 during vegetative and reproductive development of bell pepper (Capsicum annuum L.). 3 Biotech 2018, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.C.; Geraats, B.P.J.; Linthorst, H.J.M. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci. 2006, 11, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Gorshkov, V.; Tsers, I. Plant susceptible responses: The underestimated side of plant-pathogen interactions. Biol. Rev. Camb. Philos. Soc. 2022, 97, 45–66. [Google Scholar] [CrossRef]
- Palva, T.K.; Hurtig, M.; Saindrenan, P.; Palva, E.T. Salicylic acid induced resistance to Erwinia carotovora subsp. carotovora in tobacco. Mol. Plant Microbe Interact. 1994, 7, 356–363. [Google Scholar] [CrossRef]
- Joshi, J.R.; Burdman, S.; Lipsky, A.; Yariv, S.; Yedidia, I. Plant phenolic acids affect the virulence of Pectobacterium aroidearum and P. carotovorum ssp. brasiliense via quorum sensing regulation. Mol. Plant Pathol. 2016, 17, 487–500. [Google Scholar] [CrossRef]
- Czajkowski, R.; van der Wolf, J.; Krolicka, A. Salicylic acid can reduce infection symptoms caused by Dickeya solani in tissue culture grown potato (Solanum tuberosum L.) plants. Eur. J. Plant Pathol. 2015, 141, 545–558. [Google Scholar] [CrossRef]
- Burra, D.D.; Mühlenbock, P.; Andreasson, E. Salicylic and jasmonic acid pathways are necessary for defence against Dickeya solani as revealed by a novel method for blackleg disease screening of in vitro grown potato. Plant Biol. 2005, 17, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Figaj, D.; Ambroziak, P.; Przepiora, T.; Skorko-Glonek, J. The role of proteases in the virulence of plant pathogenic bacteria. Int. J. Mol. Sci. 2019, 20, 672. [Google Scholar] [CrossRef] [PubMed]
- Ghigo, J.M.; Wandersman, C. A fourth metalloprotease gene in Erwinia chrysanthemi. Res. Microbiol. 1992, 143, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Nyffenegger, C.; Højrup, P.; Vidal-Melgosa, S.; Yan, K.P.; Fangel, J.U.; Meyer, A.S.; Kirpekar, F.; Willats, W.G.; Mikkelsen, J.D. Characterization of an extensin-modifying metalloprotease: N-terminal processing and substrate cleavage pattern of Pectobacterium carotovorum Prt1. Appl. Microbiol. Biotechnol. 2014, 98, 10077–10089. [Google Scholar] [CrossRef] [PubMed]
- Ökmen, B.; Kemmerich, B.; Hilbig, D.; Wemhöner, R.; Aschenbroich, J.; Perrar, A.; Huesgen, P.F.; Schipper, K.; Doehlemann, G. Dual function of a secreted fungalysin metalloprotease in Ustilago maydis. New Phytol. 2018, 220, 249–261. [Google Scholar] [CrossRef]
- Naumann, T.A.; Price, N.P. Truncation of class IV chitinases from Arabidopsis by secreted fungal proteases. Mol. Plant Pathol. 2012, 13, 1135–1139. [Google Scholar] [CrossRef]
- Sanz-Martín, J.M.; Pacheco-Arjona, J.R.; Bello-Rico, V.; Vargas, W.A.; Monod, M.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. A highly conserved metalloprotease effector enhances virulence in the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant Pathol. 2016, 17, 1048–1062. [Google Scholar] [CrossRef]
- Dow, J.M.; Clarke, B.R.; Milligan, D.E.; Tang, J.L.; Daniels, M.J. Extracellular proteases from Xanthomonas campestris pv. campestris, the black rot pathogen. Appl. Environ. Microbiol. 1990, 56, 2994–2998. [Google Scholar] [CrossRef]
- Grinter, R.; Hay, I.D.; Song, J.; Wang, J.; Teng, D.; Dhanesakaran, V.; Wilksch, J.J.; Davies, M.R.; Littler, D.; Beckham, S.A.; et al. FusC, a member of the M16 protease family acquired by bacteria for iron piracy against plants. PLoS Biol. 2018, 16, e2006026. [Google Scholar] [CrossRef]
- Jia, Y.; McAdams, S.A.; Bryan, G.T.; Hershey, H.P.; Valent, B. Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. EMBO J. 2000, 19, 4004–4014. [Google Scholar] [CrossRef]
- Han, J.; Wang, X.; Wang, F.; Zhao, Z.; Li, G.; Zhu, X.; Su, J.; Chen, L. The Fungal Effector Avr-Pita Suppresses Innate Immunity by Increasing COX Activity in Rice Mitochondria. Rice 2021, 14, 12. [Google Scholar] [CrossRef]
- Wang, X.; He, M.; Liu, H.; Ding, H.; Liu, K.; Li, Y.; Cheng, P.; Li, Q.; Wang, B. Functional Characterization of the M36 Metalloprotease FgFly1 in Fusarium graminearum. J. Fungi 2022, 8, 726. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- PSI-BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch&PROGRAM=blastp&BLAST_PROGRAMS=psiBlast (accessed on 5 December 2023).
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- SignalP—6.0—Services—DTU Health Tech. Available online: https://services.healthtech.dtu.dk/service.php?SignalP-6.0 (accessed on 5 December 2023).
- Maestro, Schrödinger. Available online: https://www.schrodinger.com/products/maestro (accessed on 5 December 2023).
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- AlphaFold2 TIB Serve. Available online: https://alphafold2.biodesign.ac.cn/ (accessed on 5 December 2023).
- Nakamura, H.; Nishida, S. Numerical calculations of electrostatic potentials of protein-solvent systems by the self consistent boundary method. J. Phys. Soc. Jpn. 1987, 56, 1609–1622. [Google Scholar] [CrossRef]
- eF-Surf. Available online: https://pdbj.org/eF-surf/top.do (accessed on 5 December 2023).
- Case, D.A.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Ghoreishi, D.; Gilson, M.K.; et al. AMBER 2018; University of California: San Francisco, CA, USA, 2018. [Google Scholar]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Laemmli, U. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Akpinar, O.; Penner, M.H. Peptidase activity assays using protein substrates. Curr. Prot. Food Anal. Chem. 2002, 3, C2.2.1–C2.2.10. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tendiuk, N.; Diakonova, A.; Petrova, O.; Mukhametzyanov, T.; Makshakova, O.; Gorshkov, V. Svx Peptidases of Phytopathogenic Pectolytic Bacteria: Structural, Catalytic and Phytoimmune Properties. Int. J. Mol. Sci. 2024, 25, 756. https://doi.org/10.3390/ijms25020756
Tendiuk N, Diakonova A, Petrova O, Mukhametzyanov T, Makshakova O, Gorshkov V. Svx Peptidases of Phytopathogenic Pectolytic Bacteria: Structural, Catalytic and Phytoimmune Properties. International Journal of Molecular Sciences. 2024; 25(2):756. https://doi.org/10.3390/ijms25020756
Chicago/Turabian StyleTendiuk, Natalia, Anastasiya Diakonova, Olga Petrova, Timur Mukhametzyanov, Olga Makshakova, and Vladimir Gorshkov. 2024. "Svx Peptidases of Phytopathogenic Pectolytic Bacteria: Structural, Catalytic and Phytoimmune Properties" International Journal of Molecular Sciences 25, no. 2: 756. https://doi.org/10.3390/ijms25020756