
NO Deficiency Compromises Inter- and Intrahemispheric Blood Flow Adaptation to Unilateral Carotid Artery Occlusion
 , , ,
, , ,
Abstract
:1. Introduction
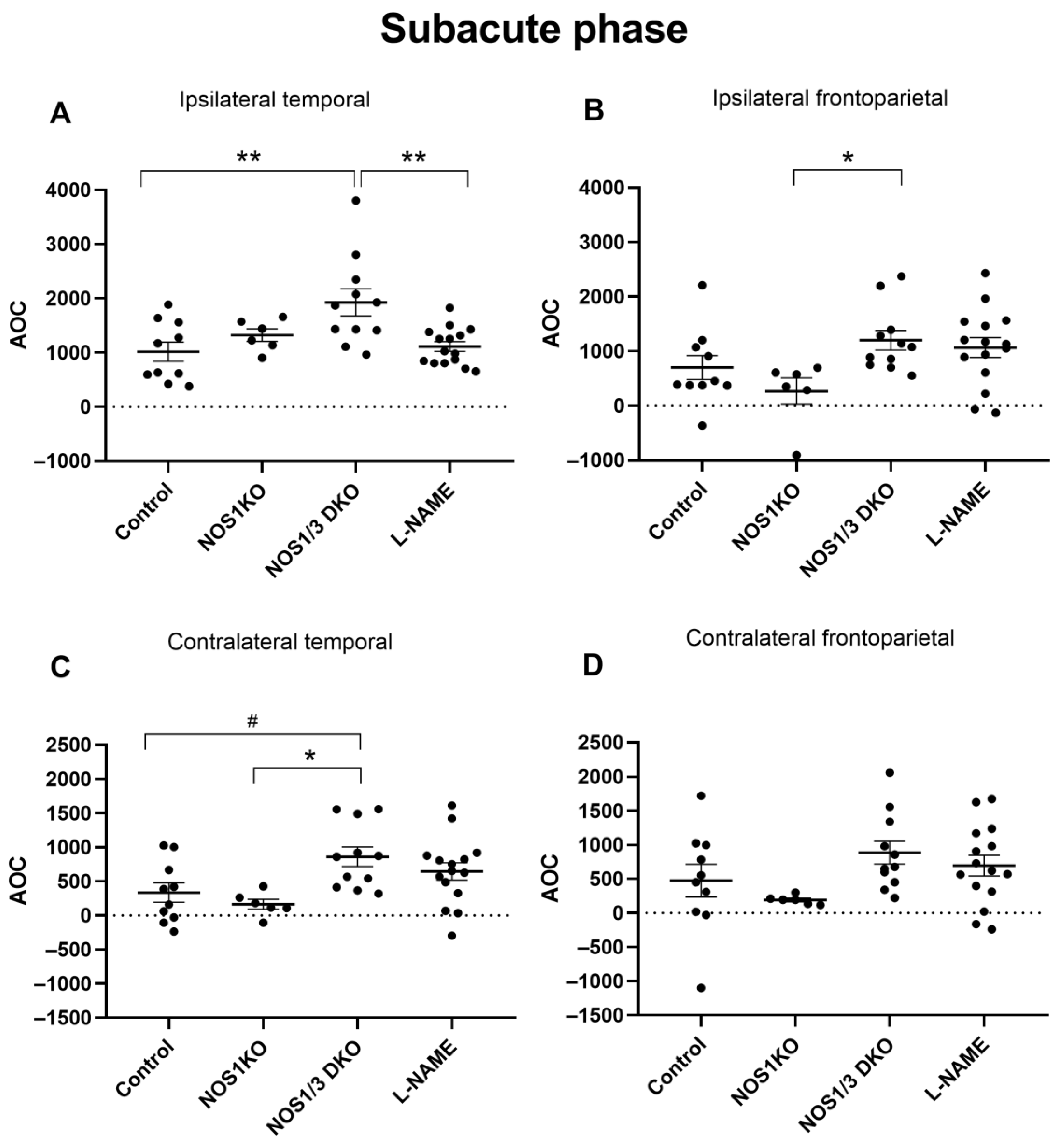
2. Results
2.1. Body Measures
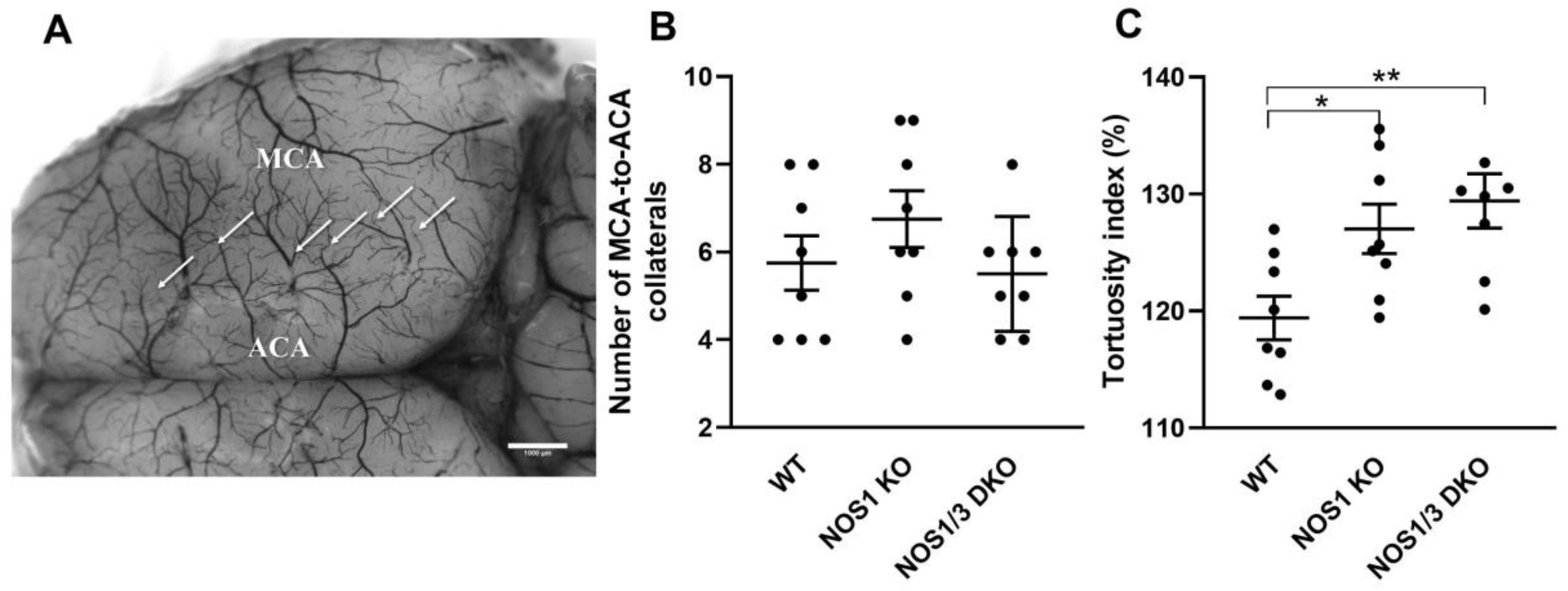
2.2. Morphological Parameters of Leptomeningeal Collaterals
2.3. Acid-Base, Blood gas, Hematocrit, and Electrolyte Parameters
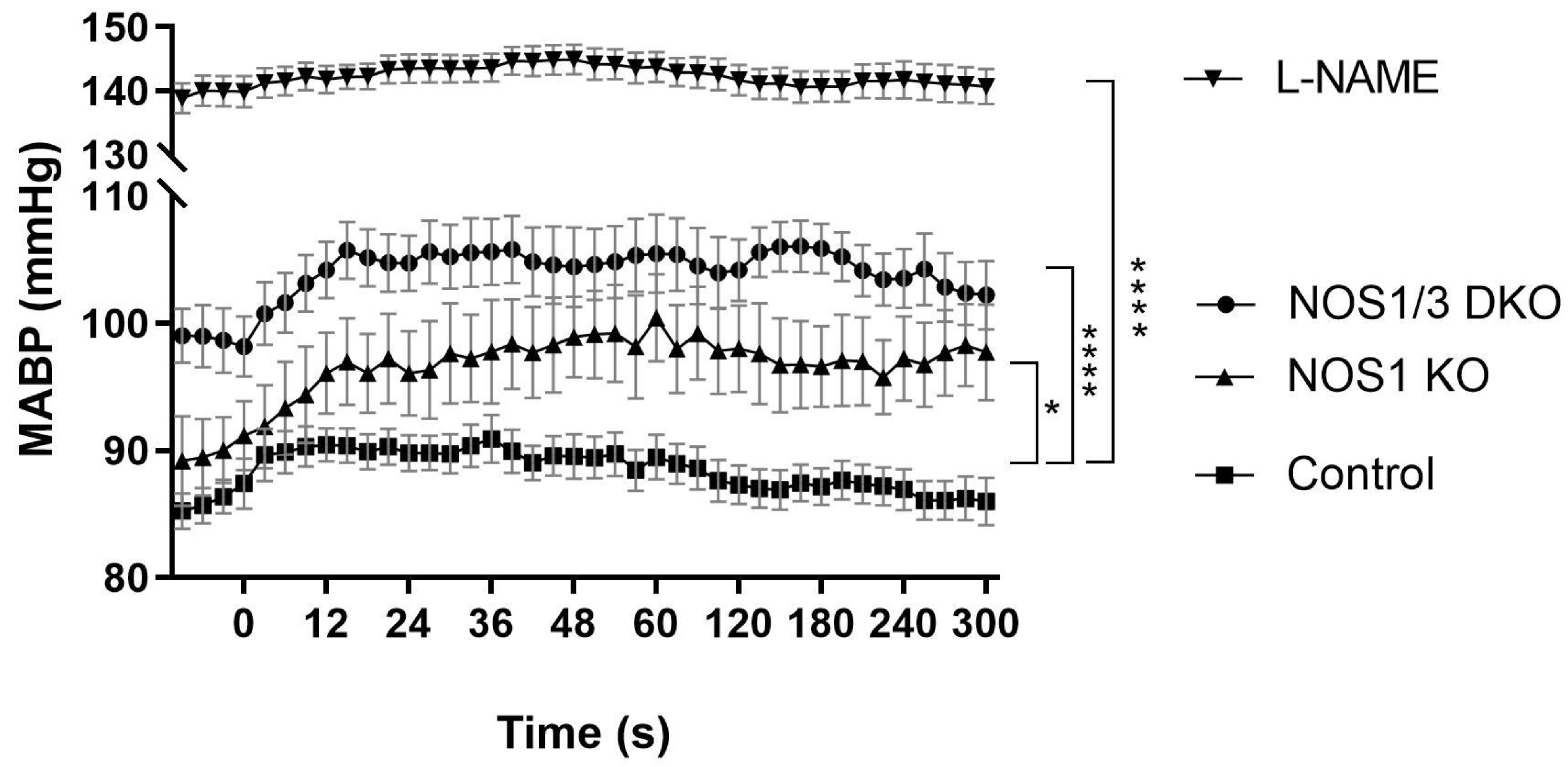
2.4. Blood Pressure
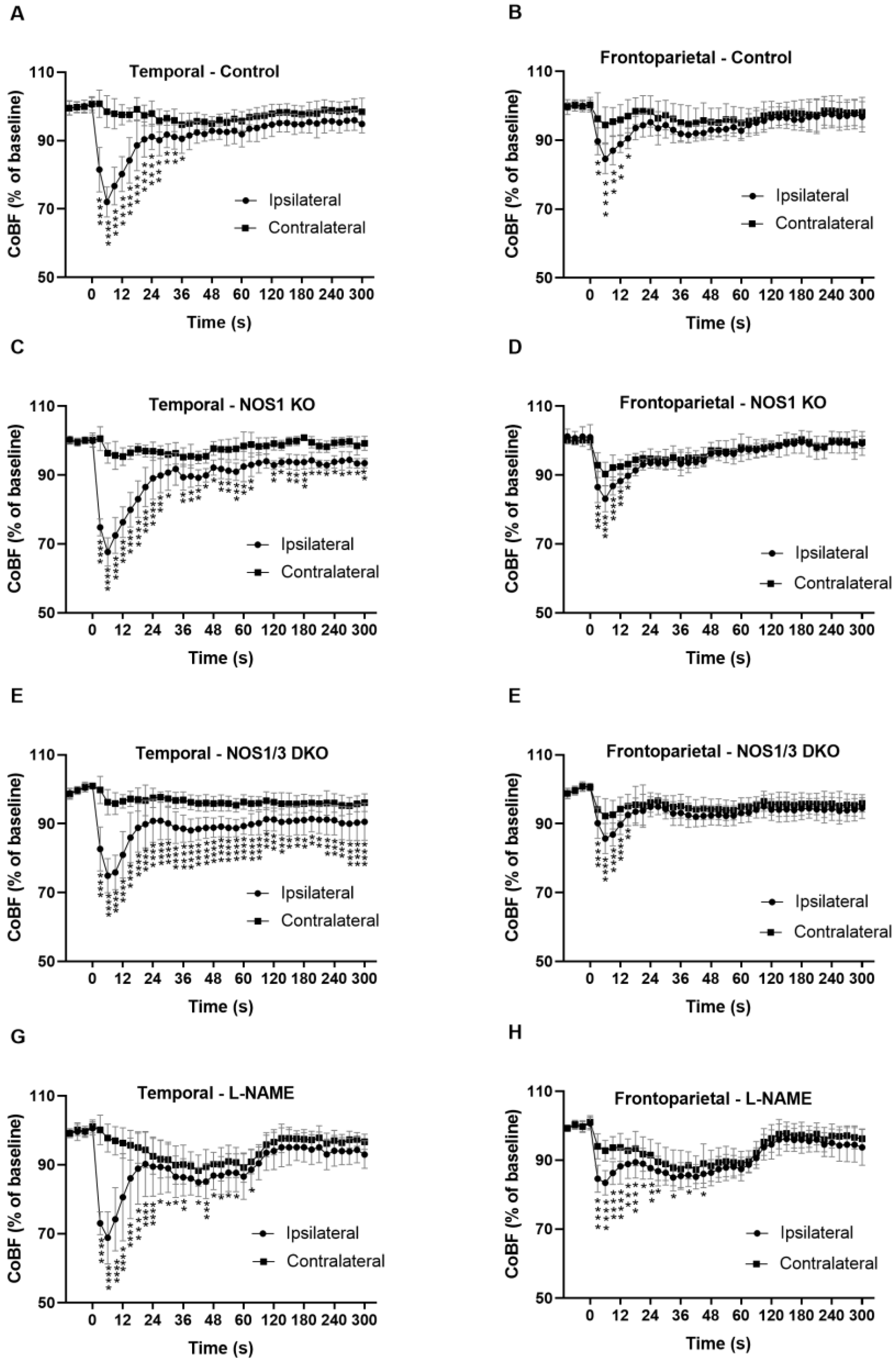
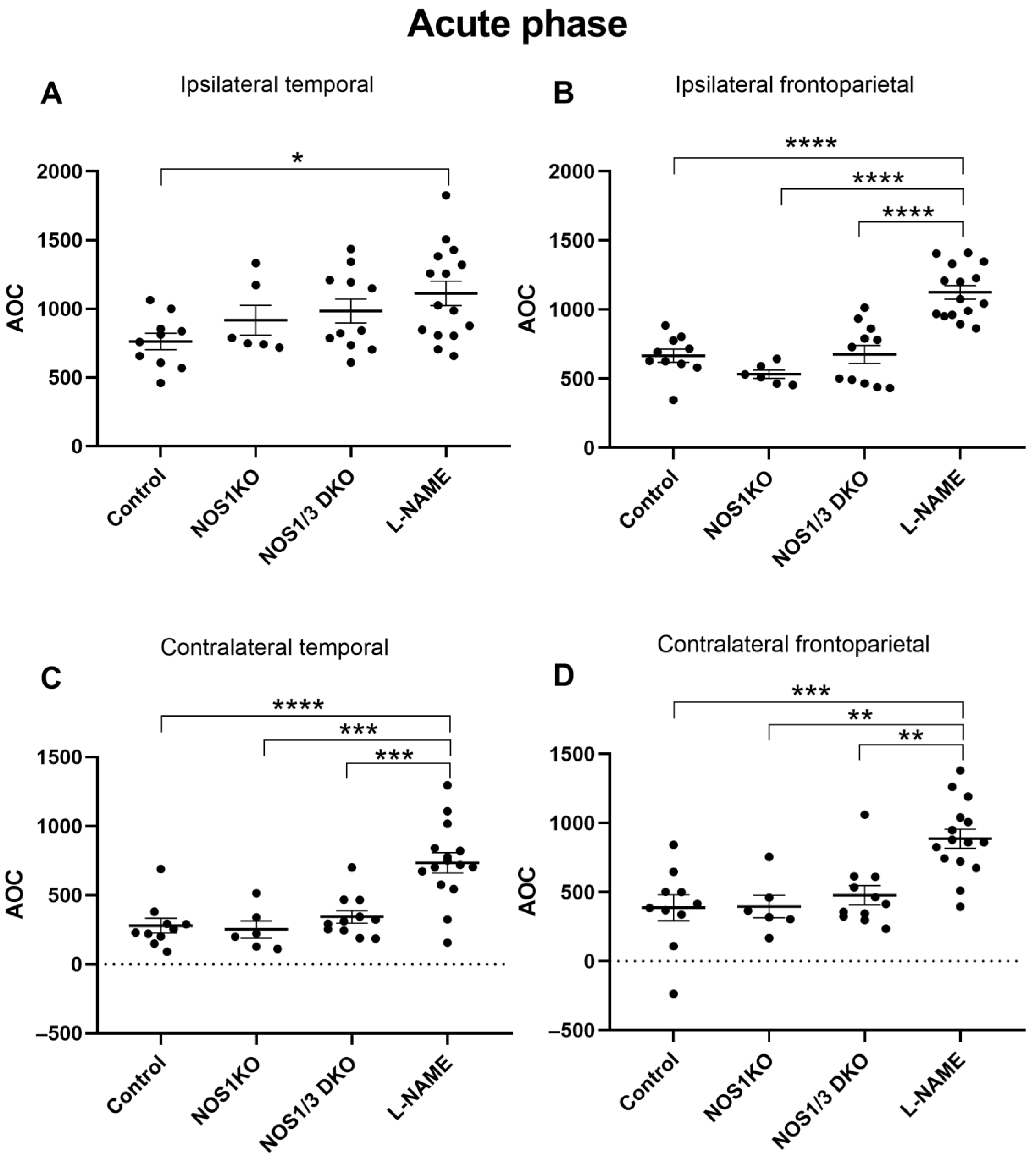
2.5. Changes in Regional CoBF after Unilateral CAO
3. Discussion
Implications of the Results in a Clinical Setting
4. Materials and Methods
4.1. Experimental Animals
4.2. Surgical Procedures, Carotid Artery Occlusion
4.3. Measurement of CAO-Induced Changes in CoBF
4.4. Morphological Analysis of the Cerebrocortical Vasculature
4.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Weerd, M.; Greving, J.P.; Hedblad, B.; Lorenz, M.W.; Mathiesen, E.B.; O’Leary, D.H.; Rosvall, M.; Sitzer, M.; de Borst, G.J.; Buskens, E.; et al. Prediction of asymptomatic carotid artery stenosis in the general population: Identification of high-risk groups. Stroke 2014, 45, 2366–2371. [Google Scholar] [CrossRef] [PubMed]
- Istvan, L.; Czako, C.; Elo, A.; Mihaly, Z.; Sotonyi, P.; Varga, A.; Ungvari, Z.; Csiszar, A.; Yabluchanskiy, A.; Conley, S.; et al. Imaging retinal microvascular manifestations of carotid artery disease in older adults: From diagnosis of ocular complications to understanding microvascular contributions to cognitive impairment. Geroscience 2021, 43, 1703–1723. [Google Scholar] [CrossRef] [PubMed]
- Istvan, L.; Czako, C.; Benyo, F.; Elo, A.; Mihaly, Z.; Sotonyi, P.; Varga, A.; Nagy, Z.Z.; Kovacs, I. The effect of systemic factors on retinal blood flow in patients with carotid stenosis: An optical coherence tomography angiography study. Geroscience 2022, 44, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Lineback, C.M.; Stamm, B.; Sorond, F.; Caprio, F.Z. Carotid disease, cognition, and aging: Time to redefine asymptomatic disease? Geroscience 2023, 45, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Yi, X.; Ruan, J.; Duan, X.; Luo, H.; Lv, Z. Influence of collateral circulation on cerebral blood flow and frontal lobe cognitive function in patients with severe internal carotid artery stenosis. BMC Neurol. 2019, 19, 151. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Duan, R.; Liu, Z.; Zhang, C.; Li, T.; Wei, Y.; Ma, N.; Wang, R. Altered functional connectivity is related to impaired cognition in left unilateral asymptomatic carotid artery stenosis patients. BMC Neurol. 2021, 21, 350. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, K.; Qiang, W.; Fan, X.; Chen, Z. Study of cognitive function in patients with severe asymptomatic carotid artery stenosis by a computerized neuropsychological assessment device. Front Psychol 2023, 14, 1055244. [Google Scholar] [CrossRef]
- Yue, W.; Wang, A.; Zhu, R.; Yan, Z.; Zheng, S.; Wang, J.; Huo, J.; Liu, Y.; Li, X.; Ji, Y. Association between Carotid Artery Stenosis and Cognitive Impairment in Stroke Patients: A Cross-Sectional Study. PLoS ONE 2016, 11, e0146890. [Google Scholar] [CrossRef]
- Katan, M.; Luft, A. Global Burden of Stroke. Semin. Neurol. 2018, 38, 208–211. [Google Scholar] [CrossRef]
- Zirak, P.; Delgado-Mederos, R.; Dinia, L.; Marti-Fabregas, J.; Durduran, T. Microvascular versus macrovascular cerebral vasomotor reactivity in patients with severe internal carotid artery stenosis or occlusion. Acad. Radiol. 2014, 21, 168–174. [Google Scholar] [CrossRef]
- Bokkers, R.P.; van Osch, M.J.; van der Worp, H.B.; de Borst, G.J.; Mali, W.P.; Hendrikse, J. Symptomatic carotid artery stenosis: Impairment of cerebral autoregulation measured at the brain tissue level with arterial spin-labeling MR imaging. Radiology 2010, 256, 201–208. [Google Scholar] [CrossRef]
- Koppl, T.; Schneider, M.; Pohl, U.; Wohlmuth, B. The influence of an unilateral carotid artery stenosis on brain oxygenation. Med. Eng. Phys. 2014, 36, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Semenyutin, V.B.; Asaturyan, G.A.; Nikiforova, A.A.; Aliev, V.A.; Panuntsev, G.K.; Iblyaminov, V.B.; Savello, A.V.; Patzak, A. Predictive Value of Dynamic Cerebral Autoregulation Assessment in Surgical Management of Patients with High-Grade Carotid Artery Stenosis. Front. Physiol. 2017, 8, 872. [Google Scholar] [CrossRef] [PubMed]
- White, R.P.; Markus, H.S. Impaired dynamic cerebral autoregulation in carotid artery stenosis. Stroke 1997, 28, 1340–1344. [Google Scholar] [CrossRef] [PubMed]
- Cassot, F.; Lauwers, F.; Fouard, C.; Prohaska, S.; Lauwers-Cances, V. A novel three-dimensional computer-assisted method for a quantitative study of microvascular networks of the human cerebral cortex. Microcirculation 2006, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Bai, X.; Wang, T.; Dmytriw, A.A.; Patel, A.B.; Jiao, L. Carotid Stenting Versus Endarterectomy for Asymptomatic Carotid Artery Stenosis: A Systematic Review and Meta-Analysis. Stroke 2022, 53, 3047–3054. [Google Scholar] [CrossRef]
- Bonati, L.H.; Jansen, O.; de Borst, G.J.; Brown, M.M. Management of atherosclerotic extracranial carotid artery stenosis. Lancet Neurol. 2022, 21, 273–283. [Google Scholar] [CrossRef]
- Schmid, S.; Tsantilas, P.; Knappich, C.; Kallmayer, M.; Breitkreuz, T.; Zimmermann, A.; Eckstein, H.H.; Kuehnl, A. Age but not sex is associated with higher risk of in-hospital stroke or death after carotid artery stenting in symptomatic and asymptomatic carotid stenosis. J. Vasc. Surg. 2019, 69, 1090–1101.e3. [Google Scholar] [CrossRef]
- Weise, J.; Kuschke, S.; Bahr, M. Gender-specific risk of perioperative complications in carotid endarterectomy patients with contralateral carotid artery stenosis or occlusion. J. Neurol. 2004, 251, 838–844. [Google Scholar] [CrossRef]
- Cohen, J.E.; Gomori, J.M.; Itshayek, E.; Pikis, S.; Keigler, G.; Eichel, R.; Leker, R.R. Ischemic complications after tailored carotid artery stenting in different subpopulations with high-grade stenosis: Feared but rare. J. Clin. Neurosci. 2015, 22, 189–194. [Google Scholar] [CrossRef]
- Chuang, Y.M.; Huang, K.L.; Chang, Y.J.; Chang, C.H.; Chang, T.Y.; Wu, T.C.; Lin, C.P.; Wong, H.F.; Liu, S.J.; Lee, T.H. Associations between Circle of Willis morphology and white matter lesion load in subjects with carotid artery stenosis. Eur. Neurol. 2011, 66, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.S.; Pavol, M.A.; Cheung, Y.K.; Strom, I.; Slane, K.; Asllani, I.; Lazar, R.M. Dissociation among hemodynamic measures in asymptomatic high grade carotid artery stenosis. J. Neurol. Sci. 2016, 367, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Ando, T.; Sekine, T.; Murai, Y.; Orita, E.; Takagi, R.; Amano, Y.; Iwata, K.; Nakaza, M.; Ogawa, M.; Obara, M.; et al. Multiparametric flow analysis using four-dimensional flow magnetic resonance imaging can detect cerebral hemodynamic impairment in patients with internal carotid artery stenosis. Neuroradiology 2020, 62, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Dankbaar, J.W.; Kerckhoffs, K.G.P.; Horsch, A.D.; van der Schaaf, I.C.; Kappelle, L.J.; Velthuis, B.K.; DUST Investigators. Internal Carotid Artery Stenosis and Collateral Recruitment in Stroke Patients. Clin. Neuroradiol. 2018, 28, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Kunieda, T.; Miyake, K.; Sakamoto, H.; Iwasaki, Y.; Iida, S.; Morise, S.; Fujita, K.; Nakamura, M.; Kaneko, S.; Kusaka, H. Leptomeningeal Collaterals Strongly Correlate with Reduced Cerebrovascular Reactivity Measured by Acetazolamide-challenged Single-photon Emission Computed Tomography Using a Stereotactic Extraction Estimation Analysis in Patients with Unilateral Internal Carotid Artery Stenosis. Intern. Med. 2017, 56, 2857–2863. [Google Scholar] [CrossRef] [PubMed]
- Waaijer, A.; van Leeuwen, M.S.; van der Worp, H.B.; Verhagen, H.J.; Mali, W.P.; Velthuis, B.K. Anatomic variations in the circle of Willis in patients with symptomatic carotid artery stenosis assessed with multidetector row CT angiography. Cerebrovasc. Dis. 2007, 23, 267–274. [Google Scholar] [CrossRef]
- Wang, J.; Ji, J.; Qiu, J.; Wang, Y. Incompleteness of circle of Willis and silent brain infarction in patients with internal carotid artery stenosis. J Clin Neurosci 2022, 98, 73–77. [Google Scholar] [CrossRef]
- Zarrinkoob, L.; Wahlin, A.; Ambarki, K.; Birgander, R.; Eklund, A.; Malm, J. Blood Flow Lateralization and Collateral Compensatory Mechanisms in Patients With Carotid Artery Stenosis. Stroke 2019, 50, 1081–1088. [Google Scholar] [CrossRef]
- Bokkers, R.P.; Wessels, F.J.; van der Worp, H.B.; Zwanenburg, J.J.; Mali, W.P.; Hendrikse, J. Vasodilatory capacity of the cerebral vasculature in patients with carotid artery stenosis. AJNR Am. J. Neuroradiol. 2011, 32, 1030–1033. [Google Scholar] [CrossRef]
- Gottler, J.; Kaczmarz, S.; Kallmayer, M.; Wustrow, I.; Eckstein, H.H.; Zimmer, C.; Sorg, C.; Preibisch, C.; Hyder, F. Flow-metabolism uncoupling in patients with asymptomatic unilateral carotid artery stenosis assessed by multi-modal magnetic resonance imaging. J. Cereb. Blood Flow Metab. 2019, 39, 2132–2143. [Google Scholar] [CrossRef]
- Igarashi, T.; Sakatani, K.; Fujiwara, N.; Murata, Y.; Suma, T.; Shibuya, T.; Hirayama, T.; Katayama, Y. Monitoring of hemodynamic change in patients with carotid artery stenosis during the tilt test using wearable near-infrared spectroscopy. Adv. Exp. Med. Biol. 2013, 789, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Milanlioglu, A.; Yaman, A.; Kolukisa, M.; Asil, T. Evaluation of cerebral hemodynamic status in patients with unilateral symptomatic carotid artery stenosis during motor tasks, through use of transcranial Doppler sonography. Arq. Neuropsiquiatr. 2022, 80, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T.; Nyul-Toth, A.; Balasubramanian, P.; Tarantini, S.; Ahire, C.; Yabluchanskiy, A.; Csipo, T.; Farkas, E.; Wren, J.D.; Garman, L.; et al. Nicotinamide mononucleotide (NMN) supplementation promotes neurovascular rejuvenation in aged mice: Transcriptional footprint of SIRT1 activation, mitochondrial protection, anti-inflammatory, and anti-apoptotic effects. Geroscience 2020, 42, 527–546. [Google Scholar] [CrossRef] [PubMed]
- Levit, A.; Hachinski, V.; Whitehead, S.N. Neurovascular unit dysregulation, white matter disease, and executive dysfunction: The shared triad of vascular cognitive impairment and Alzheimer disease. Geroscience 2020, 42, 445–465. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Roman, R.J. Reversal of cerebral hypoperfusion: A novel therapeutic target for the treatment of AD/ADRD? Geroscience 2021, 43, 1065–1067. [Google Scholar] [CrossRef] [PubMed]
- Tarantini, S.; Balasubramanian, P.; Delfavero, J.; Csipo, T.; Yabluchanskiy, A.; Kiss, T.; Nyul-Toth, A.; Mukli, P.; Toth, P.; Ahire, C.; et al. Treatment with the BCL-2/BCL-xL inhibitor senolytic drug ABT263/Navitoclax improves functional hyperemia in aged mice. Geroscience 2021, 43, 2427–2440. [Google Scholar] [CrossRef] [PubMed]
- Toth, L.; Czigler, A.; Hegedus, E.; Komaromy, H.; Amrein, K.; Czeiter, E.; Yabluchanskiy, A.; Koller, A.; Orsi, G.; Perlaki, G.; et al. Age-related decline in circulating IGF-1 associates with impaired neurovascular coupling responses in older adults. Geroscience 2022, 44, 2771–2783. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Roman, R.J.; Fan, F. Hippocampus is more susceptible to hypoxic injury: Has the Rosetta Stone of regional variation in neurovascular coupling been deciphered? Geroscience 2022, 44, 127–130. [Google Scholar] [CrossRef]
- Prado, R.; Watson, B.D.; Wester, P. Effects of nitric oxide synthase inhibition on cerebral blood flow following bilateral carotid artery occlusion and recirculation in the rat. J. Cereb. Blood Flow Metab. 1993, 13, 720–723. [Google Scholar] [CrossRef]
- Benyó, Z.; Lacza, Z.; Hortobágyi, T.; Görlach, C.; Wahl, M. Functional importance of neuronal nitric oxide synthase in the endothelium of rat basilar arteries. Brain Res. 2000, 877, 79–84. [Google Scholar] [CrossRef]
- Benyó, Z.; Szabó, C.; Stuiver, B.T.; Bohus, B.; Sándor, P. Hypothalamic blood flow remains unaltered following chronic nitric oxide synthase blockade in rats. Neurosci. Lett. 1995, 198, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Hortobágyi, L.; Kis, B.; Hrabák, A.; Horváth, B.; Huszty, G.; Schweer, H.; Benyó, B.; Sándor, P.; Busija, D.W.; Benyó, Z. Adaptation of the hypothalamic blood flow to chronic nitric oxide deficiency is independent of vasodilator prostanoids. Brain Res. 2007, 1131, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Król, M.; Kepinska, M. Human Nitric Oxide Synthase-Its Functions, Polymorphisms, and Inhibitors in the Context of Inflammation, Diabetes and Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 22, 56. [Google Scholar] [CrossRef] [PubMed]
- Pál, É.; Ungvári, Z.; Benyó, Z.; Várbíró, S. Role of Vitamin D Deficiency in the Pathogenesis of Cardiovascular and Cerebrovascular Diseases. Nutrients 2023, 15, 334. [Google Scholar] [CrossRef] [PubMed]
- Nagy, D.; Hricisák, L.; Walford, G.P.; Lékai, Á.; Karácsony, G.; Várbíró, S.; Ungvári, Z.; Benyó, Z.; Pál, É. Disruption of Vitamin D Signaling Impairs Adaptation of Cerebrocortical Microcirculation to Carotid Artery Occlusion in Hyperandrogenic Female Mice. Nutrients 2023, 15, 3869. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.L.; Dawson, T.M.; Bredt, D.S.; Snyder, S.H.; Fishman, M.C. Targeted disruption of the neuronal nitric oxide synthase gene. Cell 1993, 75, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Iversen, N.K.; Malte, H.; Baatrup, E.; Wang, T. The normal acid-base status of mice. Respir. Physiol. Neurobiol. 2012, 180, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Shesely, E.G.; Maeda, N.; Kim, H.S.; Desai, K.M.; Krege, J.H.; Laubach, V.E.; Sherman, P.A.; Sessa, W.C.; Smithies, O. Elevated blood pressures in mice lacking endothelial nitric oxide synthase. Proc. Natl. Acad. Sci. USA 1996, 93, 13176–13181. [Google Scholar] [CrossRef]
- Huang, P.L.; Huang, Z.; Mashimo, H.; Bloch, K.D.; Moskowitz, M.A.; Bevan, J.A.; Fishman, M.C. Hypertension in mice lacking the gene for endothelial nitric oxide synthase. Nature 1995, 377, 239–242. [Google Scholar] [CrossRef]
- Mattson, D.L.; Meister, C.J. Renal cortical and medullary blood flow responses to L-NAME and ANG II in wild-type, nNOS null mutant, and eNOS null mutant mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R991–R997. [Google Scholar] [CrossRef]
- Faraci, F.M. Regulation of the cerebral circulation by endothelium. Pharmacol. Ther. 1992, 56, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Faraci, F.M. Role of endothelium-derived relaxing factor in cerebral circulation: Large arteries vs. microcirculation. Am J Physiol 1991, 261, H1038–H1042. [Google Scholar] [CrossRef] [PubMed]
- Prado, R.; Watson, B.D.; Kuluz, J.; Dietrich, W.D. Endothelium-derived nitric oxide synthase inhibition. Effects on cerebral blood flow, pial artery diameter, and vascular morphology in rats. Stroke 1992, 23, 1118–1123, discussion 1124. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C. Regulation of the cerebral microcirculation during neural activity: Is nitric oxide the missing link? Trends Neurosci. 1993, 16, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Polycarpou, A.; Hricisák, L.; Iring, A.; Safar, D.; Ruisanchez, É.; Horváth, B.; Sándor, P.; Benyó, Z. Adaptation of the cerebrocortical circulation to carotid artery occlusion involves blood flow redistribution between cortical regions and is independent of eNOS. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H972–H980. [Google Scholar] [CrossRef] [PubMed]
- Saqqur, M.; Khan, K.; Derksen, C.; Alexandrov, A.; Shuaib, A. Transcranial Doppler and Transcranial Color Duplex in Defining Collateral Cerebral Blood Flow. J. Neuroimaging 2018, 28, 455–476. [Google Scholar] [CrossRef]
- Matsunaga, T.; Weihrauch, D.W.; Moniz, M.C.; Tessmer, J.; Warltier, D.C.; Chilian, W.M. Angiostatin inhibits coronary angiogenesis during impaired production of nitric oxide. Circulation 2002, 105, 2185–2191. [Google Scholar] [CrossRef] [PubMed]
- Murohara, T.; Asahara, T.; Silver, M.; Bauters, C.; Masuda, H.; Kalka, C.; Kearney, M.; Chen, D.; Symes, J.F.; Fishman, M.C.; et al. Nitric oxide synthase modulates angiogenesis in response to tissue ischemia. J. Clin. Investig. 1998, 101, 2567–2578. [Google Scholar] [CrossRef]
- Loesch, A.; Burnstock, G. Perivascular nerve fibres and endothelial cells of the rat basilar artery: Immuno-gold labelling of antigenic sites for type I and type III nitric oxide synthase. J. Neurocytol. 1998, 27, 197–204. [Google Scholar] [CrossRef]
- Tarantini, S.; Nyul-Toth, A.; Yabluchanskiy, A.; Csipo, T.; Mukli, P.; Balasubramanian, P.; Ungvari, A.; Toth, P.; Benyo, Z.; Sonntag, W.E.; et al. Endothelial deficiency of insulin-like growth factor-1 receptor (IGF1R) impairs neurovascular coupling responses in mice, mimicking aspects of the brain aging phenotype. Geroscience 2021, 43, 2387–2394. [Google Scholar] [CrossRef]
- Young, A.P.; Zhu, J.; Bagher, A.M.; Denovan-Wright, E.M.; Howlett, S.E.; Kelly, M.E.M. Endothelin B receptor dysfunction mediates elevated myogenic tone in cerebral arteries from aged male Fischer 344 rats. Geroscience 2021, 43, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Bagi, Z.; Kroenke, C.D.; Fopiano, K.A.; Tian, Y.; Filosa, J.A.; Sherman, L.S.; Larson, E.B.; Keene, C.D.; Degener O’Brien, K.; Adeniyi, P.A.; et al. Association of cerebral microvascular dysfunction and white matter injury in Alzheimer’s disease. Geroscience 2022, 44, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ayo-Martin, O.; Garcia-Garcia, J.; Hernandez-Fernandez, F.; Gomez-Hontanilla, M.; Gomez-Fernandez, I.; Andres-Fernandez, C.; Lamas, C.; Alfaro-Martinez, J.J.; Botella, F.; Segura, T. Cerebral hemodynamics in obesity: Relationship with sex, age, and adipokines in a cohort-based study. Geroscience 2021, 43, 1465–1479. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.W.; Montgomery, P.S.; Wang, M.; Shen, B.; Casanegra, A.I.; Silva-Palacios, F.; Ungvari, Z.; Yabluchanskiy, A.; Csiszar, A.; Waldstein, S.R. Cognitive decrement in older adults with symptomatic peripheral artery disease. Geroscience 2021, 43, 2455–2465. [Google Scholar] [CrossRef] [PubMed]
- Banga, P.V.; Varga, A.; Csobay-Novák, C.; Kolossváry, M.; Szántó, E.; Oderich, G.S.; Entz, L.; Sótonyi, P. Incomplete circle of Willis is associated with a higher incidence of neurologic events during carotid eversion endarterectomy without shunting. J. Vasc. Surg. 2018, 68, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Magyar-Stang, R.; Pál, H.; Csányi, B.; Gaál, A.; Mihály, Z.; Czinege, Z.; Csipo, T.; Ungvari, Z.; Sótonyi, P.; Varga, A.; et al. Assessment of cerebral autoregulatory function and inter-hemispheric blood flow in older adults with internal carotid artery stenosis using transcranial Doppler sonography-based measurement of transient hyperemic response after carotid artery compression. Geroscience 2023, 45, 3333–3357. [Google Scholar] [CrossRef]
- Csobay-Novák, C.; Bárány, T.; Zima, E.; Nemes, B.; Sótonyi, P.; Merkely, B.; Hüttl, K. Role of stent selection in the incidence of persisting hemodynamic depression after carotid artery stenting. J. Endovasc. Ther. 2015, 22, 122–129. [Google Scholar] [CrossRef]
- Pál, É.; Hricisák, L.; Lékai, Á.; Nagy, D.; Fülöp, Á.; Erben, R.G.; Várbíró, S.; Sándor, P.; Benyó, Z. Ablation of Vitamin D Signaling Compromises Cerebrovascular Adaptation to Carotid Artery Occlusion in Mice. Cells 2020, 9, 1457. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wild Type | NOS1 KO | NOS1/3 DKO | |
|---|---|---|---|
| body weight (g) | 31.04 ± 0.57 | 26.13 ± 1.17 **** | 26.04 ± 0.20 **** |
| heart weight (g) | 0.153 ± 0.004 | 0.133 ± 0.008 * | 0.121 ± 0.004 **** |
| heart weight/body weight (‰) | 5.05 ± 0.14 | 5.29 ± 0.25 | 4.66 ± 0.18 |
| tibial length (mm) | 18.0 (17.9–18.0) | 18.0 (17.0–18.1) | 17.5 (17.5–18.0) |
| body weight/tibial length (g/mm) | 1.698 ± 0.026 | 1.416 ± 0.058 **** | 1.483 ± 0.024 **** |
| heart weight/tibial length (mg/mm) | 8.549 ± 0.205 | 7.451 ± 0.362 * | 6.892 ± 0.255 *** |
| brain weight (g) | 0.456 ± 0.004 | 0.435 ± 0.007 * | 0.430 ± 0.003 *** |
| brain weight/body weight (%) | 1.503 ± 0.024 | 1.747 ± 0.082 ** | 1.653 ± 0.022 * |
| brain weight/tibial length (mg/mm) | 25.48 ± 0.27 | 24.54 ± 0.59 | 24.49 ± 0.38 |
| Control | NOS1KO | NOS1/3 DKO | L-NAME | |
|---|---|---|---|---|
| pH | 7.28 ± 0.04 | 7.26 ± 0.10 | 7.29 ± 0.06 | 7.24 ± 0.08 |
| SBE (mmol/L) | −8.97 ± 0.88 | −10.03 ± 2.00 | −10.43 ± 1.89 | −10.17 ± 0.94 |
| HCO3− (mmol/L) | 17.96 ± 0.52 | 15.93 ± 1.46 | 18.40 ± 0.42 | 17.33 ± 0.83 |
| pCO2 (mmHg) | 39.8 ± 1.7 | 38.2 ± 2.7 | 40.3 ± 1.8 | 40.5 ± 1.9 |
| pO2 (mmHg) | 107.7 ± 2.29 | 119.7 ± 7.0 | 102.6 ± 2.0 | 112.2 ± 3.3 |
| satO2 (%) | 97.21 ± 0.27 | 97.80 ± 0.52 | 96.82 ± 0.41 | 96.95 ± 0.52 |
| Hct (%) | 41.4 ± 0.4 | 39.0 ± 0.8 | 41.5 ± 0.4 | 43.4 ± 0.9 |
| Na+ (mmol/L) | 155.6 ± 1.2 | 153.0 ± 1.2 | 156.5 ± 1.3 | 154.4 ± 0.9 |
| K+ (mmol/L) | 4.62 (3.88–4.98) | 4.82 (4.70–4.93) | 4.87 (4.37–4.48) | 5.07 (4.30–6.20) |
| Ca2+ (mmol/L) | 1.30 ± 0.04 | 1.30 ± 0.05 | 1.31 ± 0.03 | 1.24 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hricisák, L.; Pál, É.; Nagy, D.; Delank, M.; Polycarpou, A.; Fülöp, Á.; Sándor, P.; Sótonyi, P.; Ungvári, Z.; Benyó, Z. NO Deficiency Compromises Inter- and Intrahemispheric Blood Flow Adaptation to Unilateral Carotid Artery Occlusion. Int. J. Mol. Sci. 2024, 25, 697. https://doi.org/10.3390/ijms25020697
Hricisák L, Pál É, Nagy D, Delank M, Polycarpou A, Fülöp Á, Sándor P, Sótonyi P, Ungvári Z, Benyó Z. NO Deficiency Compromises Inter- and Intrahemispheric Blood Flow Adaptation to Unilateral Carotid Artery Occlusion. International Journal of Molecular Sciences. 2024; 25(2):697. https://doi.org/10.3390/ijms25020697
Chicago/Turabian StyleHricisák, László, Éva Pál, Dorina Nagy, Max Delank, Andreas Polycarpou, Ágnes Fülöp, Péter Sándor, Péter Sótonyi, Zoltán Ungvári, and Zoltán Benyó. 2024. "NO Deficiency Compromises Inter- and Intrahemispheric Blood Flow Adaptation to Unilateral Carotid Artery Occlusion" International Journal of Molecular Sciences 25, no. 2: 697. https://doi.org/10.3390/ijms25020697