
Clinical Relevance of Vaginal and Endometrial Microbiome Investigation in Women with Repeated Implantation Failure and Recurrent Pregnancy Loss

Abstract
:1. Introduction
2. Role of Microbiota in Modulating Immune Tolerance during Early Pregnancy
2.1. Embryo Implantation
2.2. Immunology of Early Pregnancy
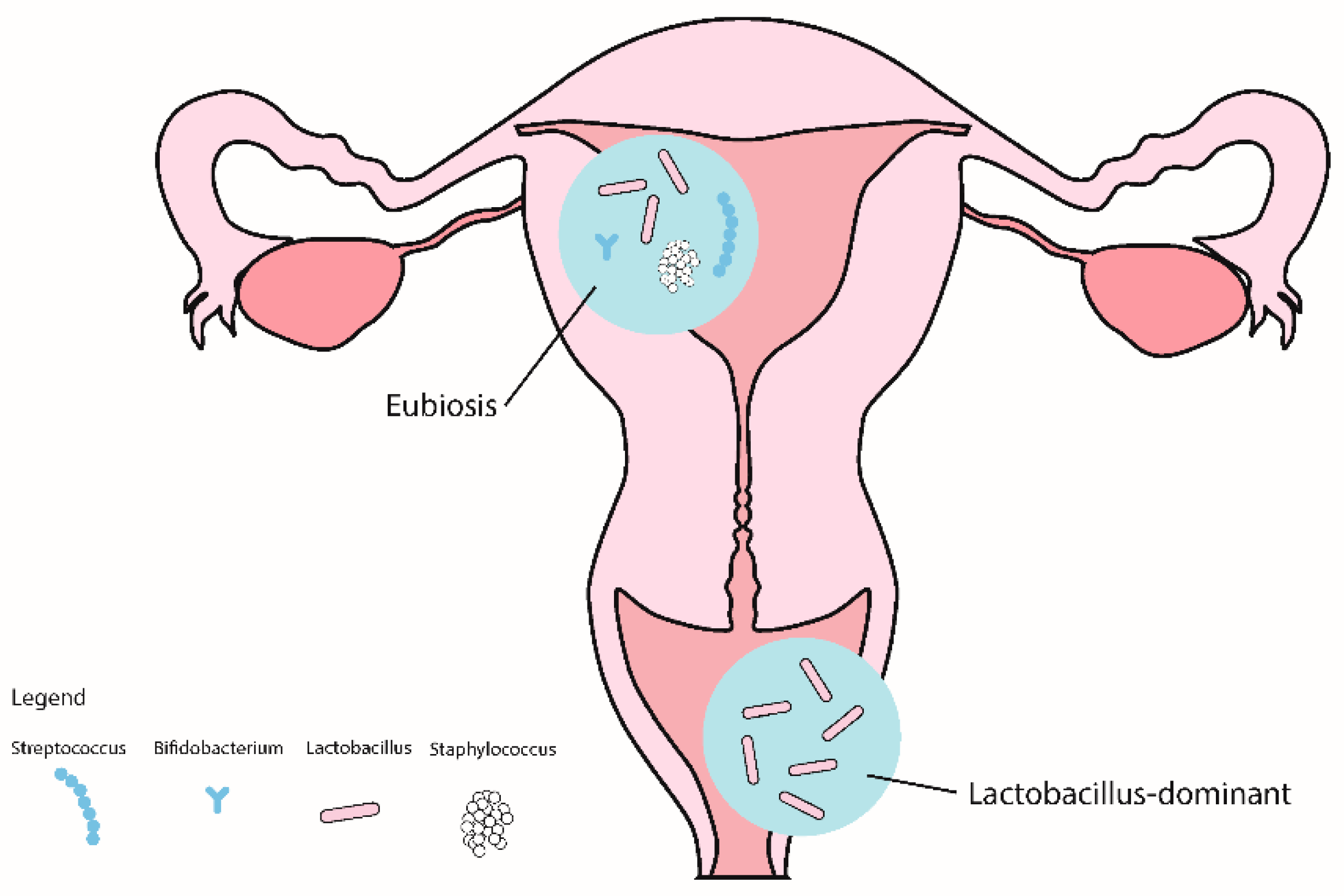
2.3. The Role of the Reproductive Tract Microbiota in Early Pregnancy
3. Repeated Implantation Failure and Recurrent Pregnancy Loss
3.1. Definition, Prevalence, and Prognosis of Repeated Implantation Failure
3.2. Definition, Prevalence, and Prognosis of Recurrent Pregnancy Loss
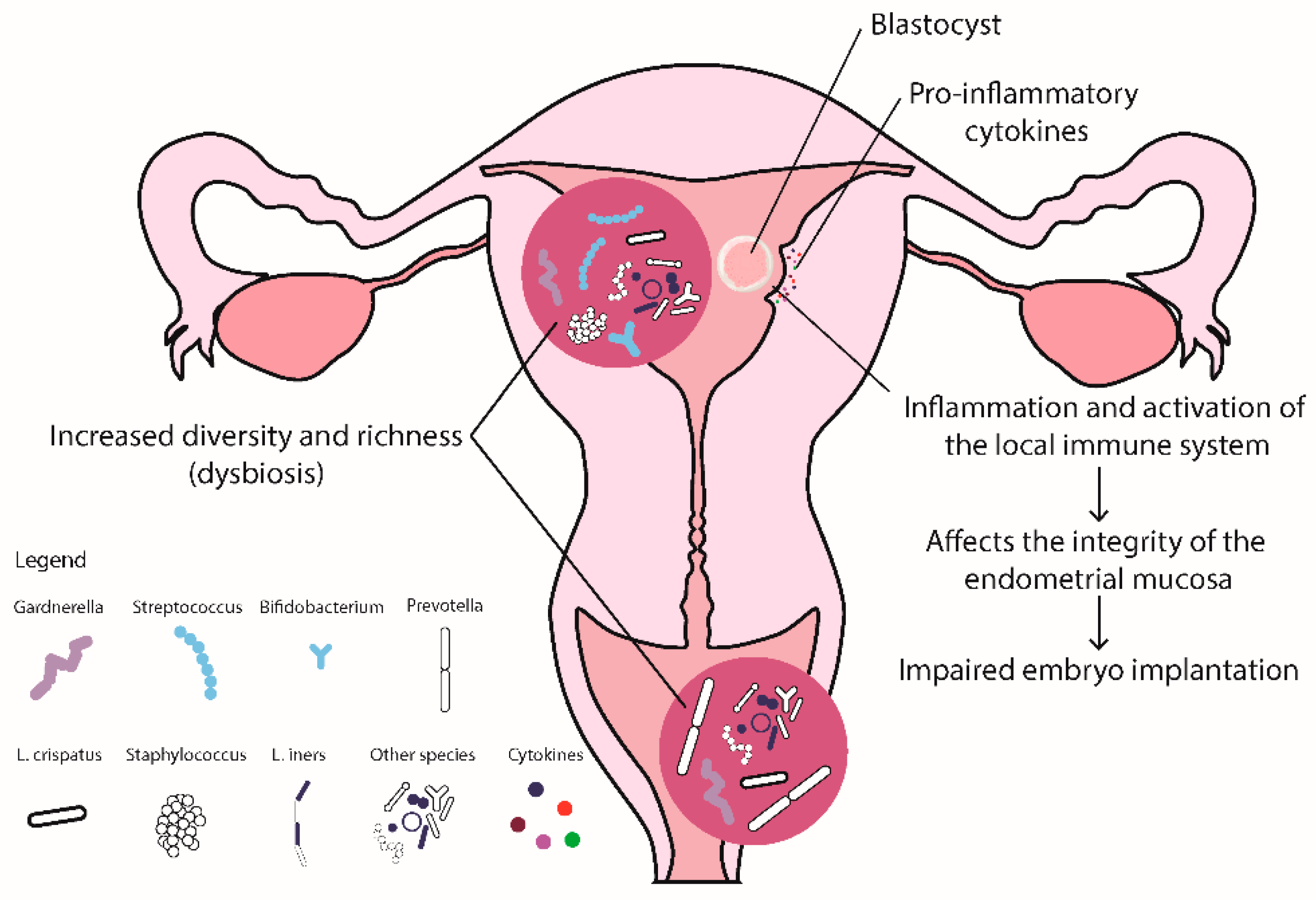
3.3. The Role of Reproductive Tract Microbiota in RIF and RPL
3.4. Repeated Implantation Failure
3.4.1. Vaginal Microbiome
3.4.2. Uterine Microbiome
3.4.3. Molecular Immunological Characteristics of RIF
Natural Killer Cells
T Lymphocytes
3.4.4. Reproductive Microbiome and Immunology in RIF
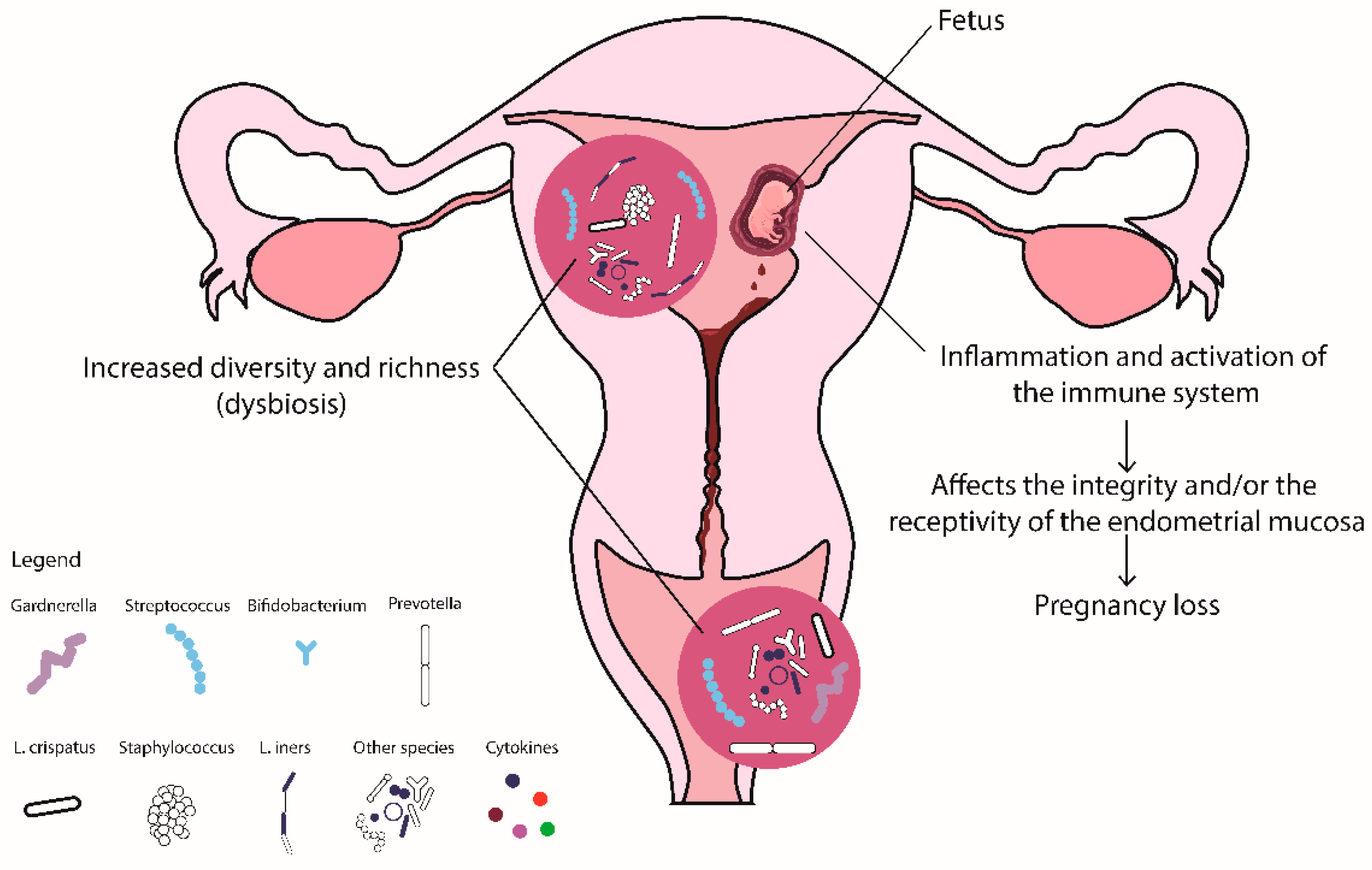
3.5. Recurrent Pregnancy Loss
3.5.1. Vaginal Microbiome
3.5.2. Uterine Microbiome
3.5.3. Molecular Immunological Characteristics of RPL
NK Cells
Cytokines
T lymphocytes
3.5.4. Reproductive Tract Microbiome and Immunology in RPL
4. Summary
5. Challenges and Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersen, A.N.; Goossens, V.; Bhattacharya, S.; Ferraretti, A.P.; Kupka, M.S.; De Mouzon, J.; Nygren, K.G.; European Society of Human Reproduction and Embryology (ESHRE). Assisted reproductive technology and intrauterine inseminations in Europe, 2005: Results generated from European registers by ESHRE: ESHRE. The European IVF Monitoring Programme (EIM), for the European Society of Human Reproduction and Embryology (ESHRE). Hum. Reprod. 2009, 24, 1267–1287. [Google Scholar] [CrossRef]
- Macklon, N.S.; Geraedts, J.P.; Fauser, B.C. Conception to ongoing pregnancy: The ‘black box’ of early pregnancy loss. Hum. Reprod. Update 2002, 8, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Senn, V.; Bassler, D.; Choudhury, R.; Scholkmann, F.; Righini-Grunder, F.; Vuille-Dit-Bile, R.N.; Restin, T. Microbial Colonization From the Fetus to Early Childhood-A Comprehensive Review. Front. Cell. Infect. Microbiol. 2020, 10, 573735. [Google Scholar] [CrossRef] [PubMed]
- Alexander, K.L.; Targan, S.R.; Elson, C.O., III. Microbiota activation and regulation of innate and adaptive immunity. Immunol. Rev. 2014, 260, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Postler, T.S.; Ghosh, S. Understanding the Holobiont: How Microbial Metabolites Affect Human Health and Shape the Immune System. Cell Metab. 2017, 26, 110–130. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Ye, Y.; Wang, F.; Chen, F.; Zheng, C. The Role of Microbiota in Infant Health: From Early Life to Adulthood. Front. Immunol. 2021, 12, 708472. [Google Scholar] [CrossRef]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1894070. [Google Scholar] [CrossRef]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; Ter Horst, R.; Jansen, T.; Jacobs, L.; Bonder, M.J.; et al. Linking the Human Gut Microbiome to Inflammatory Cytokine Production Capacity. Cell 2016, 167, 1125–1136.e8. [Google Scholar] [CrossRef]
- Tersigni, C.; D’Ippolito, S.; Di Nicuolo, F.; Marana, R.; Valenza, V.; Masciullo, V.; Scaldaferri, F.; Malatacca, F.; de Waure, C.; Gasbarrini, A.; et al. Recurrent pregnancy loss is associated to leaky gut: A novel pathogenic model of endometrium inflammation? J. Transl. Med. 2018, 16, 102. [Google Scholar] [CrossRef]
- Freudenberg, M.A.; Kalis, C.; Chvatchko, Y.; Merlin, T.; Gumenscheimer, M.; Galanos, C. Role of interferons in LPS hypersensitivity. J. Endotoxin Res. 2003, 9, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Cimadomo, D.; Craciunas, L.; Vermeulen, N.; Vomstein, K.; Toth, B. Definition, diagnostic and therapeutic options in recurrent implantation failure: An international survey of clinicians and embryologists. Hum. Reprod. 2021, 36, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Bernabeu, A.; Lledo, B.; Diaz, M.C.; Lozano, F.M.; Ruiz, V.; Fuentes, A.; Lopez-Pineda, A.; Moliner, B.; Castillo, J.C.; Ortiz, J.A.; et al. Effect of the vaginal microbiome on the pregnancy rate in women receiving assisted reproductive treatment. J. Assist. Reprod. Genet. 2019, 36, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Kyono, K.; Hashimoto, T.; Kikuchi, S.; Nagai, Y.; Sakuraba, Y. A pilot study and case reports on endometrial microbiota and pregnancy outcome: An analysis using 16S rRNA gene sequencing among IVF patients, and trial therapeutic intervention for dysbiotic endometrium. Reprod. Med. Biol. 2019, 18, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, M.; Mekaru, K.; Tanaka, S.E.; Arai, W.; Ashikawa, K.; Sakuraba, Y.; Nakamura, R.; Oishi, S.; Akamine, K.; Aoki, Y. Endometrial and vaginal microbiomes influence assisted reproductive technology outcomes. JBRA Assist. Reprod. 2022, 27, 267. [Google Scholar] [CrossRef] [PubMed]
- Moreno, I.; Codoner, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazan, J.; Alonso, R.; Alama, P.; Remohi, J.; Pellicer, A.; et al. Evidence that the endometrial microbiota has an effect on implantation success or failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef]
- Grewal, K.; Lee, Y.S.; Smith, A.; Brosens, J.J.; Bourne, T.; Al-Memar, M.; Kundu, S.; MacIntyre, D.A.; Bennett, P.R. Chromosomally normal miscarriage is associated with vaginal dysbiosis and local inflammation. BMC Med. 2022, 20, 38. [Google Scholar] [CrossRef]
- Manhanzva, M.T.; Abrahams, A.G.; Gamieldien, H.; Froissart, R.; Jaspan, H.; Jaumdally, S.Z.; Barnabas, S.L.; Dabee, S.; Bekker, L.G.; Gray, G.; et al. Inflammatory and antimicrobial properties differ between vaginal Lactobacillus isolates from South African women with non-optimal versus optimal microbiota. Sci. Rep. 2020, 10, 6196. [Google Scholar] [CrossRef]
- Koedooder, R.; Mackens, S.; Budding, A.; Fares, D.; Blockeel, C.; Laven, J.; Schoenmakers, S. Identification and evaluation of the microbiome in the female and male reproductive tracts. Hum. Reprod. Update 2019, 25, 298–325. [Google Scholar] [CrossRef]
- Bischof, P.; Campana, A. Trophoblast differentiation and invasion: Its significance for human embryo implantation. Early Pregnancy 1997, 3, 81–95. [Google Scholar]
- Norwitz, E.R.; Schust, D.J.; Fisher, S.J. Implantation and the survival of early pregnancy. N. Engl. J. Med. 2001, 345, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.D.; Bagchi, I.; Dey, S.K.; Enders, A.C.; Fazleabas, A.T.; Lessey, B.A.; Yoshinaga, K. Embryo implantation. Dev. Biol. 2000, 223, 217–237. [Google Scholar] [CrossRef] [PubMed]
- Dekel, N.; Gnainsky, Y.; Granot, I.; Racicot, K.; Mor, G. The role of inflammation for a successful implantation. Am. J. Reprod. Immunol. 2014, 72, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Franasiak, J.M.; Alecsandru, D.; Forman, E.J.; Gemmell, L.C.; Goldberg, J.M.; Llarena, N.; Margolis, C.; Laven, J.; Schoenmakers, S.; Seli, E. A review of the pathophysiology of recurrent implantation failure. Fertil. Steril. 2021, 116, 1436–1448. [Google Scholar] [CrossRef] [PubMed]
- Teklenburg, G.; Salker, M.; Heijnen, C.; Macklon, N.S.; Brosens, J.J. The molecular basis of recurrent pregnancy loss: Impaired natural embryo selection. Mol. Hum. Reprod. 2010, 16, 886–895. [Google Scholar] [CrossRef]
- Brosens, J.J.; Bennett, P.R.; Abrahams, V.M.; Ramhorst, R.; Coomarasamy, A.; Quenby, S.; Lucas, E.S.; McCoy, R.C. Maternal selection of human embryos in early gestation: Insights from recurrent miscarriage. Semin. Cell Dev. Biol. 2022, 131, 14–24. [Google Scholar] [CrossRef]
- Teklenburg, G.; Salker, M.; Molokhia, M.; Lavery, S.; Trew, G.; Aojanepong, T.; Mardon, H.J.; Lokugamage, A.U.; Rai, R.; Landles, C.; et al. Natural selection of human embryos: Decidualizing endometrial stromal cells serve as sensors of embryo quality upon implantation. PLoS ONE 2010, 5, e10258. [Google Scholar] [CrossRef]
- Mor, G.; Cardenas, I. The immune system in pregnancy: A unique complexity. Am. J. Reprod. Immunol. 2010, 63, 425–433. [Google Scholar] [CrossRef]
- Gaynor, L.M.; Colucci, F. Uterine Natural Killer Cells: Functional Distinctions and Influence on Pregnancy in Humans and Mice. Front. Immunol. 2017, 8, 467. [Google Scholar] [CrossRef]
- Al-Memar, M.; Bobdiwala, S.; Fourie, H.; Mannino, R.; Lee, Y.S.; Smith, A.; Marchesi, J.R.; Timmerman, D.; Bourne, T.; Bennett, P.R.; et al. The association between vaginal bacterial composition and miscarriage: A nested case-control study. BJOG 2020, 127, 264–274. [Google Scholar] [CrossRef]
- Mor, G.; Aldo, P.; Alvero, A.B. The unique immunological and microbial aspects of pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Koedooder, R.; Singer, M.; Schoenmakers, S.; Savelkoul, P.H.M.; Morre, S.A.; de Jonge, J.D.; Poort, L.; Cuypers, W.S.S.; Budding, A.E.; Laven, J.S.E.; et al. The ReceptIVFity cohort study protocol to validate the urogenital microbiome as predictor for IVF or IVF/ICSI outcome. Reprod. Health 2018, 15, 202. [Google Scholar] [CrossRef] [PubMed]
- Toson, B.; Simon, C.; Moreno, I. The endometrial microbiome and its impact on human conception. Int. J. Mol. Sci. 2022, 23, 485. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Diaz, D.J.; Jesaveluk, B.; Hayward, J.A.; Tyssen, D.; Alisoltani, A.; Potgieter, M.; Bell, L.; Ross, E.; Iranzadeh, A.; Allali, I.; et al. Lactic acid from vaginal microbiota enhances cervicovaginal epithelial barrier integrity by promoting tight junction protein expression. Microbiome 2022, 10, 141. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Zhang, X.; Liang, Y.; Lin, S.; Qian, W.; Fan, S. Alterations in Vaginal Microbiota and Associated Metabolome in Women with Recurrent Implantation Failure. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S.; Alvi, S.; Bongiovanni, A.M.; Linhares, I.M.; Ledger, W.J. Lactic acid stimulates interleukin-23 production by peripheral blood mononuclear cells exposed to bacterial lipopolysaccharide. FEMS Immunol. Med. Microbiol. 2011, 61, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Baryla, M.; Kaczynski, P.; Goryszewska, E.; Riley, S.C.; Waclawik, A. Prostaglandin F(2alpha) stimulates adhesion, migration, invasion and proliferation of the human trophoblast cell line HTR-8/SVneo. Placenta 2019, 77, 19–29. [Google Scholar] [CrossRef]
- Kaczynski, P.; Goryszewska, E.; Baryla, M.; Waclawik, A. Prostaglandin F2alpha stimulates angiogenesis at the embryo-maternal interface during early pregnancy in the pig. Theriogenology 2020, 142, 169–176. [Google Scholar] [CrossRef]
- Lebedeva, O.P.; Popov, V.N.; Syromyatnikov, M.Y.; Starkova, N.N.; Maslov, A.Y.; Kozarenko, O.N.; Gryaznova, M.V. Female reproductive tract microbiome and early miscarriages. Apmis 2023, 131, 61–76. [Google Scholar] [CrossRef]
- Polanski, L.T.; Baumgarten, M.N.; Quenby, S.; Brosens, J.; Campbell, B.K.; Raine-Fenning, N.J. What exactly do we mean by ‘recurrent implantation failure’? A systematic review and opinion. Reprod. Biomed. Online 2014, 28, 409–423. [Google Scholar] [CrossRef]
- Tan, B.K.; Vandekerckhove, P.; Kennedy, R.; Keay, S.D. Investigation and current management of recurrent IVF treatment failure in the UK. BJOG 2005, 112, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, C.; Ledger, W.; Wang, Q.; Liu, F.; Demirol, A.; Gurgan, T.; Cutting, R.; Ong, K.; Sallam, H.; Li, T.C. Recurrent implantation failure: Definition and management. Reprod. Biomed. Online 2014, 28, 14–38. [Google Scholar] [CrossRef] [PubMed]
- Busnelli, A.; Reschini, M.; Cardellicchio, L.; Vegetti, W.; Somigliana, E.; Vercellini, P. How common is real repeated implantation failure? An indirect estimate of the prevalence. Reprod. Biomed. Online 2020, 40, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Pirtea, P.; De Ziegler, D.; Tao, X.; Sun, L.; Zhan, Y.; Ayoubi, J.M.; Seli, E.; Franasiak, J.M.; Scott, R.T., Jr. Rate of true recurrent implantation failure is low: Results of three successive frozen euploid single embryo transfers. Fertil. Steril. 2021, 115, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Koot, Y.E.M.; Hviid Saxtorph, M.; Goddijn, M.; de Bever, S.; Eijkemans, M.J.C.; Wely, M.V.; van der Veen, F.; Fauser, B.; Macklon, N.S. What is the prognosis for a live birth after unexplained recurrent implantation failure following IVF/ICSI? Hum. Reprod. 2019, 34, 2044–2052. [Google Scholar] [CrossRef] [PubMed]
- Vomstein, K.; Voss, P.; Molnar, K.; Ainsworth, A.; Daniel, V.; Strowitzki, T.; Toth, B.; Kuon, R.J. Two of a kind? Immunological and clinical risk factors differ between recurrent implantation failure and recurrent miscarriage. J. Reprod. Immunol. 2020, 141, 103166. [Google Scholar] [CrossRef] [PubMed]
- Orvieto, R.; Meltcer, S.; Nahum, R.; Rabinson, J.; Anteby, E.Y.; Ashkenazi, J. The influence of body mass index on in vitro fertilization outcome. Int. J. Gynaecol. Obstet. 2009, 104, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Waylen, A.L.; Metwally, M.; Jones, G.L.; Wilkinson, A.J.; Ledger, W.L. Effects of cigarette smoking upon clinical outcomes of assisted reproduction: A meta-analysis. Hum. Reprod Update 2009, 15, 31–44. [Google Scholar] [CrossRef]
- Zeadna, A.; Son, W.Y.; Moon, J.H.; Dahan, M.H. A comparison of biochemical pregnancy rates between women who underwent IVF and fertile controls who conceived spontaneouslydagger. Hum. Reprod. 2015, 30, 783–788. [Google Scholar] [CrossRef]
- Ford, H.B.; Schust, D.J. Recurrent pregnancy loss: Etiology, diagnosis, and therapy. Rev. Obstet. Gynecol. 2009, 2, 76–83. [Google Scholar]
- Egerup, P.; Kolte, A.M.; Larsen, E.C.; Krog, M.; Nielsen, H.S.; Christiansen, O.B. Recurrent pregnancy loss: What is the impact of consecutive versus non-consecutive losses? Hum. Reprod 2016, 31, 2428–2434. [Google Scholar] [CrossRef] [PubMed]
- Krog, M.C.; Peker, N.; Hugerth, L.W.; Vomstein, K.; Bliddal, S.; Reinhardt Nielsen, J.; Slot, A.; Vauvert Hviid, K.; Engstrand, L.; Schuppe-Koistinen, I. O-262 The vaginal and faecal microbiome in women with recurrent pregnancy loss (RPL) before pregnancy according to the reproductive outcome after referral. Hum. Reprod. 2023, 38, dead093-316. [Google Scholar] [CrossRef]
- Kaiser, J.; Branch, D.W. Recurrent Pregnancy Loss: Generally Accepted Causes and Their Management. Clin. Obstet. Gynecol. 2016, 59, 464–473. [Google Scholar] [CrossRef] [PubMed]
- RPL, E.G.G.o.; Bender Atik, R.; Christiansen, O.B.; Elson, J.; Kolte, A.M.; Lewis, S.; Middeldorp, S.; Nelen, W.; Peramo, B.; Quenby, S.; et al. ESHRE guideline: Recurrent pregnancy loss. Hum. Reprod. Open 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- RPL, E.G.G.o.; Bender Atik, R.; Christiansen, O.B.; Elson, J.; Kolte, A.M.; Lewis, S.; Middeldorp, S.; McHeik, S.; Peramo, B.; Quenby, S.; et al. ESHRE guideline: Recurrent pregnancy loss: An update in 2022. Hum. Reprod. Open 2023, 2023, hoad002. [Google Scholar] [CrossRef]
- Al-Nasiry, S.; Ambrosino, E.; Schlaepfer, M.; Morre, S.A.; Wieten, L.; Voncken, J.W.; Spinelli, M.; Mueller, M.; Kramer, B.W. The Interplay Between Reproductive Tract Microbiota and Immunological System in Human Reproduction. Front. Immunol. 2020, 11, 378. [Google Scholar] [CrossRef]
- Bardos, J.; Fiorentino, D.; Longman, R.E.; Paidas, M. Immunological Role of the Maternal Uterine Microbiome in Pregnancy: Pregnancies Pathologies and Alterated Microbiota. Front. Immunol. 2019, 10, 2823. [Google Scholar] [CrossRef]
- Benner, M.; Ferwerda, G.; Joosten, I.; van der Molen, R.G. How uterine microbiota might be responsible for a receptive, fertile endometrium. Hum. Reprod. Update 2018, 24, 393–415. [Google Scholar] [CrossRef]
- Ravel, J.; Moreno, I.; Simón, C. Bacterial vaginosis and its association with infertility, endometritis, and pelvic inflammatory disease. Am. J. Obstet. Gynecol. 2021, 224, 251–257. [Google Scholar] [CrossRef]
- Schwebke, J.R.; Richey, C.M.; Weiss, H.L. Correlation of behaviors with microbiological changes in vaginal flora. J. Infect. Dis. 1999, 180, 1632–1636. [Google Scholar] [CrossRef]
- Neggers, Y.H.; Nansel, T.R.; Andrews, W.W.; Schwebke, J.R.; Yu, K.F.; Goldenberg, R.L.; Klebanoff, M.A. Dietary intake of selected nutrients affects bacterial vaginosis in women. J. Nutr. 2007, 137, 2128–2133. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, P.; Andersen, L.O.; Lilje, B.; Johannesen, T.B.; Dahl, E.G.; Baig, S.; Jensen, J.S.; Falk, L. Changes in the vaginal microbiota following antibiotic treatment for Mycoplasma genitalium, Chlamydia trachomatis and bacterial vaginosis. PLoS ONE 2020, 15, e0236036. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Martinez, M.D.C.; Bernabeu, A.; Lledo, B.; Carratala-Munuera, C.; Quesada, J.A.; Lozano, F.M.; Ruiz, V.; Morales, R.; Llacer, J.; Ten, J.; et al. Impact of the Vaginal and Endometrial Microbiome Pattern on Assisted Reproduction Outcomes. J. Clin. Med. 2021, 10, 4063. [Google Scholar] [CrossRef] [PubMed]
- Haahr, T.; Jensen, J.S.; Thomsen, L.; Duus, L.; Rygaard, K.; Humaidan, P. Abnormal vaginal microbiota may be associated with poor reproductive outcomes: A prospective study in IVF patients. Hum. Reprod. 2016, 31, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Ichiyama, T.; Kuroda, K.; Nagai, Y.; Urushiyama, D.; Ohno, M.; Yamaguchi, T.; Nagayoshi, M.; Sakuraba, Y.; Yamasaki, F.; Hata, K.; et al. Analysis of vaginal and endometrial microbiota communities in infertile women with a history of repeated implantation failure. Reprod. Med. Biol. 2021, 20, 334–344. [Google Scholar] [CrossRef]
- Kong, Y.; Liu, Z.; Shang, Q.; Gao, Y.; Li, X.; Zheng, C.; Deng, X.; Chen, T. The Disordered Vaginal Microbiota Is a Potential Indicator for a Higher Failure of in vitro Fertilization. Front. Med. 2020, 7, 217. [Google Scholar] [CrossRef]
- Patel, N.; Patel, N.; Pal, S.; Nathani, N.; Pandit, R.; Patel, M.; Patel, N.; Joshi, C.; Parekh, B. Distinct gut and vaginal microbiota profile in women with recurrent implantation failure and unexplained infertility. BMC Women’s Health 2022, 22, 113. [Google Scholar] [CrossRef]
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The microbiota continuum along the female reproductive tract and its relation to uterine-related diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.N.; Eschenbach, D. Colonization of the upper genital tract by vaginal bacterial species in nonpregnant women. Am. J. Obstet. Gynecol. 2015, 212, e611–e619. [Google Scholar] [CrossRef]
- O’Callaghan, J.L.; Turner, R.; Dekker Nitert, M.; Barrett, H.L.; Clifton, V.; Pelzer, E.S. Re-assessing microbiomes in the low-biomass reproductive niche. BJOG 2020, 127, 147–158. [Google Scholar] [CrossRef]
- Zou, Y.; Liu, X.; Chen, P.; Wang, Y.; Li, W.; Huang, R. The endometrial microbiota profile influenced pregnancy outcomes in patients with repeated implantation failure: A retrospective study. J. Reprod. Immunol. 2023, 155, 103782. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; Matteo, M.; Tinelli, R.; Lepera, A.; Alfonso, R.; Indraccolo, U.; Marrocchella, S.; Greco, P.; Resta, L. Prevalence of chronic endometritis in repeated unexplained implantation failure and the IVF success rate after antibiotic therapy. Hum. Reprod. 2015, 30, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.G.; Ferrari, F.; Ciebiera, M.; Zgliczyńska, M.; Rapisarda, A.M.C.; Vecchio, G.M.; Pino, A.; Angelico, G.; Knafel, A.; Riemma, G.; et al. The Role of Genital Tract Microbiome in Fertility: A Systematic Review. Int. J. Mol. Sci. 2021, 23, 180. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ko, E.Y.; Wong, K.K.; Chen, X.; Cheung, W.C.; Law, T.S.; Chung, J.P.; Tsui, S.K.; Li, T.C.; Chim, S.S. Endometrial microbiota in infertile women with and without chronic endometritis as diagnosed using a quantitative and reference range-based method. Fertil. Steril. 2019, 112, 707–717.e1. [Google Scholar] [CrossRef]
- Chen, P.; Chen, P.; Guo, Y.; Fang, C.; Li, T. Interaction Between Chronic Endometritis Caused Endometrial Microbiota Disorder and Endometrial Immune Environment Change in Recurrent Implantation Failure. Front. Immunol. 2021, 12, 748447. [Google Scholar] [CrossRef] [PubMed]
- Borgdorff, H.; Gautam, R.; Armstrong, S.D.; Xia, D.; Ndayisaba, G.F.; van Teijlingen, N.H.; Geijtenbeek, T.B.; Wastling, J.M.; van de Wijgert, J.H. Cervicovaginal microbiome dysbiosis is associated with proteome changes related to alterations of the cervicovaginal mucosal barrier. Mucosal. Immunol. 2016, 9, 621–633. [Google Scholar] [CrossRef]
- Vomstein, K.; Reider, S.; Bottcher, B.; Watschinger, C.; Kyvelidou, C.; Tilg, H.; Moschen, A.R.; Toth, B. Uterine microbiota plasticity during the menstrual cycle: Differences between healthy controls and patients with recurrent miscarriage or implantation failure. J. Reprod. Immunol. 2022, 151, 103634. [Google Scholar] [CrossRef]
- Cela, V.; Daniele, S.; Obino, M.E.R.; Ruggiero, M.; Zappelli, E.; Ceccarelli, L.; Papini, F.; Marzi, I.; Scarfo, G.; Tosi, F.; et al. Endometrial Dysbiosis Is Related to Inflammatory Factors in Women with Repeated Implantation Failure: A Pilot Study. J. Clin. Med. 2022, 11, 2481. [Google Scholar] [CrossRef]
- Tuckerman, E.; Mariee, N.; Prakash, A.; Li, T.C.; Laird, S. Uterine natural killer cells in peri-implantation endometrium from women with repeated implantation failure after IVF. J. Reprod. Immunol. 2010, 87, 60–66. [Google Scholar] [CrossRef]
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK cells regulate key developmental processes at the human fetal-maternal interface. Nat. Med. 2006, 12, 1065–1074. [Google Scholar] [CrossRef]
- Vacca, P.; Mingari, M.C.; Moretta, L. Natural killer cells in human pregnancy. J. Reprod. Immunol. 2013, 97, 14–19. [Google Scholar] [CrossRef]
- Moffett, A.; Colucci, F. Uterine NK cells: Active regulators at the maternal-fetal interface. J. Clin. Investig. 2014, 124, 1872–1879. [Google Scholar] [CrossRef]
- Chen, X.; Mariee, N.; Jiang, L.; Liu, Y.; Wang, C.C.; Li, T.C.; Laird, S. Measurement of uterine natural killer cell percentage in the periimplantation endometrium from fertile women and women with recurrent reproductive failure: Establishment of a reference range. Am. J. Obstet. Gynecol. 2017, 217, 680.e1–680.e6. [Google Scholar] [CrossRef] [PubMed]
- Fukui, A.; Kwak-Kim, J.; Ntrivalas, E.; Gilman-Sachs, A.; Lee, S.K.; Beaman, K. Intracellular cytokine expression of peripheral blood natural killer cell subsets in women with recurrent spontaneous abortions and implantation failures. Fertil. Steril. 2008, 89, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Nakashima, A.; Myojo-Higuma, S.; Shiozaki, A. The balance between cytotoxic NK cells and regulatory NK cells in human pregnancy. J. Reprod. Immunol. 2008, 77, 14–22. [Google Scholar] [CrossRef] [PubMed]
- King, A. Uterine leukocytes and decidualization. Hum. Reprod. Update 2000, 6, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sung, N.; Gilman-Sachs, A.; Kwak-Kim, J. T Helper (Th) Cell Profiles in Pregnancy and Recurrent Pregnancy Losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef]
- Sasaki, Y.; Sakai, M.; Miyazaki, S.; Higuma, S.; Shiozaki, A.; Saito, S. Decidual and peripheral blood CD4+CD25+ regulatory T cells in early pregnancy subjects and spontaneous abortion cases. Mol. Hum. Reprod. 2004, 10, 347–353. [Google Scholar] [CrossRef]
- Kaminski, V.L.; Ellwanger, J.H.; Matte, M.C.C.; Savaris, R.F.; Vianna, P.; Chies, J.A.B. IL-17 blood levels increase in healthy pregnancy but not in spontaneous abortion. Mol. Biol. Rep. 2018, 45, 1565–1568. [Google Scholar] [CrossRef]
- Liang, P.Y.; Diao, L.H.; Huang, C.Y.; Lian, R.C.; Chen, X.; Li, G.G.; Zhao, J.; Li, Y.Y.; He, X.B.; Zeng, Y. The pro-inflammatory and anti-inflammatory cytokine profile in peripheral blood of women with recurrent implantation failure. Reprod. Biomed. Online 2015, 31, 823–826. [Google Scholar] [CrossRef]
- Amjadi, F.; Zandieh, Z.; Mehdizadeh, M.; Aghajanpour, S.; Raoufi, E.; Aghamajidi, A.; Aflatoonian, R. The uterine immunological changes may be responsible for repeated implantation failure. J. Reprod. Immunol. 2020, 138, 103080. [Google Scholar] [CrossRef]
- Rajaei, S.; Zarnani, A.H.; Jeddi-Tehrani, M.; Tavakoli, M.; Mohammadzadeh, A.; Dabbagh, A.; Mirahmadian, M. Cytokine profile in the endometrium of normal fertile and women with repeated implantation failure. Iran. J. Immunol. 2011, 8, 201–208. [Google Scholar] [PubMed]
- Hortal, M.; Lledo, B.; Ortiz, J.A.; Fabregat, A.; Moliner, B.; Bernabeu, A.; Bernabeu, R. O-261 The relationship of abnormal vaginal microbiome (dysbiosis of vaginal microbiome) and proinflammatory cytokines in recurrent implantation failure patients. Hum. Reprod. 2023, 38, dead093.315. [Google Scholar] [CrossRef]
- Fan, T.; Zhong, X.M.; Wei, X.C.; Miao, Z.L.; Luo, S.Y.; Cheng, H.; Xiao, Q. The alteration and potential relationship of vaginal microbiota and chemokines for unexplained recurrent spontaneous abortion. Medicine 2020, 99, e23558. [Google Scholar] [CrossRef] [PubMed]
- Kuon, R.J.; Togawa, R.; Vomstein, K.; Weber, M.; Goeggl, T.; Strowitzki, T.; Markert, U.R.; Zimmermann, S.; Daniel, V.; Dalpke, A.H.; et al. Higher prevalence of colonization with Gardnerella vaginalis and gram-negative anaerobes in patients with recurrent miscarriage and elevated peripheral natural killer cells. J. Reprod. Immunol. 2017, 120, 15–19. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, T.; Ma, Y.; Huang, Z.; He, Y.; Pan, H.; Fang, M.; Ding, H. Alteration of vaginal microbiota in patients with unexplained recurrent miscarriage. Exp. Ther. Med. 2019, 17, 3307–3316. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Zhang, L.; Du, D.; Wang, L.; Song, Q.; Liu, S. Alteration of vaginal microbiota in patients with recurrent miscarriage. J. Obstet. Gynaecol. 2022, 42, 248–255. [Google Scholar] [CrossRef]
- Zhao, F.; Chen, Y.; Gao, J.; Wu, M.; Li, C.; Wang, Z.; Huang, N.; Cui, L.; Du, M.; Ying, C. Characterization of Vaginal Microbiota in Women With Recurrent Spontaneous Abortion That Can Be Modified by Drug Treatment. Front. Cell. Infect. Microbiol. 2021, 11, 680643. [Google Scholar] [CrossRef]
- Peuranpaa, P.; Holster, T.; Saqib, S.; Kalliala, I.; Tiitinen, A.; Salonen, A.; Hautamaki, H. Female reproductive tract microbiota and recurrent pregnancy loss: A nested case-control study. Reprod. Biomed. Online 2022, 45, 1021–1031. [Google Scholar] [CrossRef]
- Skafte-Holm, A.; Humaidan, P.; Bernabeu, A.; Lledo, B.; Jensen, J.S.; Haahr, T. The Association between Vaginal Dysbiosis and Reproductive Outcomes in Sub-Fertile Women Undergoing IVF-Treatment: A Systematic PRISMA Review and Meta-Analysis. Pathogens 2021, 10, 295. [Google Scholar] [CrossRef]
- Mori, R.; Hayakawa, T.; Hirayama, M.; Ozawa, F.; Yoshihara, H.; Goto, S.; Kitaori, T.; Ozaki, Y.; Sugiura-Ogasawara, M. Cervicovaginal microbiome in patients with recurrent pregnancy loss. J. Reprod. Immunol. 2023, 157, 103944. [Google Scholar] [CrossRef]
- Chen, S.; Xue, X.; Zhang, Y.; Zhang, H.; Huang, X.; Chen, X.; Deng, G.; Luo, S.; Gao, J. Vaginal Atopobium is Associated with Spontaneous Abortion in the First Trimester: A Prospective Cohort Study in China. Microbiol. Spectr. 2022, 10, e0203921. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.T.; Yang, S.; Yang, Z.; Zhou, P.; Peng, T.; Yin, J.; Ye, Z.; Shan, H.; Yu, Y.; Li, R. An Altered Microbiota in the Lower and Upper Female Reproductive Tract of Women with Recurrent Spontaneous Abortion. Microbiol. Spectr. 2022, 10, e0046222. [Google Scholar] [CrossRef]
- Campisciano, G.; Iebba, V.; Zito, G.; Luppi, S.; Martinelli, M.; Fischer, L.; De Seta, F.; Basile, G.; Ricci, G.; Comar, M. Lactobacillus iners and gasseri, Prevotella bivia and HPV Belong to the Microbiological Signature Negatively Affecting Human Reproduction. Microorganisms 2020, 9, 39. [Google Scholar] [CrossRef]
- Nasioudis, D.; Forney, L.J.; Schneider, G.M.; Gliniewicz, K.; France, M.; Boester, A.; Sawai, M.; Scholl, J.; Witkin, S.S. Influence of Pregnancy History on the Vaginal Microbiome of Pregnant Women in their First Trimester. Sci. Rep. 2017, 7, 10201. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wong, K.K.; Ko, E.Y.; Chen, X.; Huang, J.; Tsui, S.K.; Li, T.C.; Chim, S.S. Systematic Comparison of Bacterial Colonization of Endometrial Tissue and Fluid Samples in Recurrent Miscarriage Patients: Implications for Future Endometrial Microbiome Studies. Clin. Chem. 2018, 64, 1743–1752. [Google Scholar] [CrossRef]
- Shi, Y.; Yamada, H.; Sasagawa, Y.; Tanimura, K.; Deguchi, M. Uterine endometrium microbiota and pregnancy outcome in women with recurrent pregnancy loss. J. Reprod. Immunol. 2022, 152, 103653. [Google Scholar] [CrossRef]
- Karami, N.; Boroujerdnia, M.G.; Nikbakht, R.; Khodadadi, A. Enhancement of peripheral blood CD56(dim) cell and NK cell cytotoxicity in women with recurrent spontaneous abortion or in vitro fertilization failure. J. Reprod. Immunol. 2012, 95, 87–92. [Google Scholar] [CrossRef]
- Lee, S.K.; Na, B.J.; Kim, J.Y.; Hur, S.E.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. Determination of clinical cellular immune markers in women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2013, 70, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Sunkara, S.K. Natural killer cells in female infertility and recurrent miscarriage: A systematic review and meta-analysis. Hum Reprod Update 2014, 20, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Shakhar, K.; Ben-Eliyahu, S.; Loewenthal, R.; Rosenne, E.; Carp, H. Differences in number and activity of peripheral natural killer cells in primary versus secondary recurrent miscarriage. Fertil. Steril. 2003, 80, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Katano, K.; Suzuki, S.; Ozaki, Y.; Suzumori, N.; Kitaori, T.; Sugiura-Ogasawara, M. Peripheral natural killer cell activity as a predictor of recurrent pregnancy loss: A large cohort study. Fertil. Steril. 2013, 100, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, H.; Shida, M.; Kondo, A.; Suzuki, T.; Sugi, T.; Izumi, S.; Hosaka, T.; Makino, T. Preconception peripheral natural killer cell activity as a predictor of pregnancy outcome in patients with unexplained infertility. Am. J. Reprod. Immunol. 2005, 53, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Calleja-Agius, J.; Jauniaux, E.; Pizzey, A.R.; Muttukrishna, S. Investigation of systemic inflammatory response in first trimester pregnancy failure. Hum. Reprod. 2012, 27, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Tian, Y.; Zheng, L.; Luu, T.; Kwak-Kim, J. The Update Immune-Regulatory Role of Pro- and Anti-Inflammatory Cytokines in Recurrent Pregnancy Losses. Int. J. Mol. Sci. 2022, 24, 132. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.P.; Klein, S.L. Pregnancy and pregnancy-associated hormones alter immune responses and disease pathogenesis. Horm. Behav. 2012, 62, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Bates, M.D.; Quenby, S.; Takakuwa, K.; Johnson, P.M.; Vince, G.S. Aberrant cytokine production by peripheral blood mononuclear cells in recurrent pregnancy loss? Hum. Reprod. 2002, 17, 2439–2444. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, J.Y.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. Th17 and regulatory T cells in women with recurrent pregnancy loss. Am. J. Reprod. Immunol. 2012, 67, 311–318. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, J.Y.; Hur, S.E.; Kim, C.J.; Na, B.J.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. An imbalance in interleukin-17-producing T and Foxp3(+) regulatory T cells in women with idiopathic recurrent pregnancy loss. Hum. Reprod. 2011, 26, 2964–2971. [Google Scholar] [CrossRef]
- Sereshki, N.; Gharagozloo, M.; Ostadi, V.; Ghahiri, A.; Roghaei, M.A.; Mehrabian, F.; Andalib, A.A.; Hassanzadeh, A.; Hosseini, H.; Rezaei, A. Variations in T-helper 17 and Regulatory T Cells during The Menstrual Cycle in Peripheral Blood of Women with Recurrent Spontaneous Abortion. Int. J. Fertil. Steril. 2014, 8, 59–66. [Google Scholar]
- Daher, S.; de Arruda Geraldes Denardi, K.; Blotta, M.H.; Mamoni, R.L.; Reck, A.P.; Camano, L.; Mattar, R. Cytokines in recurrent pregnancy loss. J. Reprod. Immunol. 2004, 62, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huang, L.; Lian, C.; Xue, H.; Lu, Y.; Chen, X.; Xia, Y. Vaginal Microbiota Diversity of Patients with Embryonic Miscarriage by Using 16S rDNA High-Throughput Sequencing. Int. J. Genom. 2020, 2020, 1764959. [Google Scholar] [CrossRef] [PubMed]
- Plevyak, M.; Hanna, N.; Mayer, S.; Murphy, S.; Pinar, H.; Fast, L.; Ekerfelt, C.; Ernerudh, J.; Berg, G.; Matthiesen, L.; et al. Deficiency of decidual IL-10 in first trimester missed abortion: A lack of correlation with the decidual immune cell profile. Am. J. Reprod. Immunol. 2002, 47, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Piccinni, M.P.; Robertson, S.A.; Saito, S. Editorial: Adaptive Immunity in Pregnancy. Front. Immunol. 2021, 12, 770242. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Feng, L.; Zhang, J. Interactions between gut microbiota and metabolites modulate cytokine network imbalances in women with unexplained miscarriage. NPJ Biofilms Microbiomes 2021, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen-gut microbiome axis: Physiological and clinical implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Demir, B.; Guven, S.; Guven, E.S.; Atamer, Y.; Gul, T. Serum IL-6 level may have role in the pathophysiology of unexplained infertility. Am. J. Reprod. Immunol. 2009, 62, 261–267. [Google Scholar] [CrossRef]
- Verstraelen, H.; Verhelst, R.; Claeys, G.; De Backer, E.; Temmerman, M.; Vaneechoutte, M. Longitudinal analysis of the vaginal microflora in pregnancy suggests that L. crispatus promotes the stability of the normal vaginal microflora and that L. gasseri and/or L. iners are more conducive to the occurrence of abnormal vaginal microflora. BMC Microbiol. 2009, 9, 116. [Google Scholar] [CrossRef]
- France, M.T.; Ma, B.; Gajer, P.; Brown, S.; Humphrys, M.S.; Holm, J.B.; Waetjen, L.E.; Brotman, R.M.; Ravel, J. VALENCIA: A nearest centroid classification method for vaginal microbial communities based on composition. Microbiome 2020, 8, 166. [Google Scholar] [CrossRef]
- Kervinen, K.; Holster, T.; Saqib, S.; Virtanen, S.; Stefanovic, V.; Rahkonen, L.; Nieminen, P.; Salonen, A.; Kalliala, I. Parity and gestational age are associated with vaginal microbiota composition in term and late term pregnancies. EBioMedicine 2022, 81, 104107. [Google Scholar] [CrossRef]
- Baud, D.; Pattaroni, C.; Vulliemoz, N.; Castella, V.; Marsland, B.J.; Stojanov, M. Sperm Microbiota and Its Impact on Semen Parameters. Front. Microbiol. 2019, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.L.; Chiu, C.M.; Lin, F.M.; Huang, W.C.; Liang, C.; Yang, T.; Yang, T.L.; Liu, C.Y.; Wu, W.Y.; Chang, Y.A.; et al. Bacterial communities in semen from men of infertile couples: Metagenomic sequencing reveals relationships of seminal microbiota to semen quality. PLoS ONE 2014, 9, e110152. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Luo, T.; Chen, T.; Wang, G. Seminal bacterial composition in patients with obstructive and non-obstructive azoospermia. Exp. Ther. Med. 2018, 15, 2884–2890. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, L.; Ding, J.; Cheng, Y.; Diao, L.; Li, L.; Zhang, Y.; Yin, T. Multiomics Studies Investigating Recurrent Pregnancy Loss: An Effective Tool for Mechanism Exploration. Front. Immunol. 2022, 13, 826198. [Google Scholar] [CrossRef]
- Banerjee, P.; Jana, S.K.; Pasricha, P.; Ghosh, S.; Chakravarty, B.; Chaudhury, K. Proinflammatory cytokines induced altered expression of cyclooxygenase-2 gene results in unreceptive endometrium in women with idiopathic recurrent spontaneous miscarriage. Fertil. Steril. 2013, 99, 179–187.e2. [Google Scholar] [CrossRef]
- Pan, H.T.; Ding, H.G.; Fang, M.; Yu, B.; Cheng, Y.; Tan, Y.J.; Fu, Q.Q.; Lu, B.; Cai, H.G.; Jin, X.; et al. Proteomics and bioinformatics analysis of altered protein expression in the placental villous tissue from early recurrent miscarriage patients. Placenta 2018, 61, 1–10. [Google Scholar] [CrossRef]
- Yin, X.J.; Hong, W.; Tian, F.J.; Li, X.C. Proteomic analysis of decidua in patients with recurrent pregnancy loss (RPL) reveals mitochondrial oxidative stress dysfunction. Clin. Proteomics 2021, 18, 9. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| First Author (Year) | Study Group | Country | Specimen Type | Composition: Phylum/Genus/Species | Conclusion |
|---|---|---|---|---|---|
| VAGINA | |||||
| Kitaya et al. (2019) | RIF patients (n = 28) | Japan | Vaginal swab | Lactobacillus (>90%) dominance in the RIF group. | There were no significant differences in the community composition of the vaginal microbiome between the RIF group and the control group. |
| Control n = 18 (undergoing first IVF attempt) | |||||
| Ichiyama et al. (2021) [65] | Women with RIF (n = 145) | Japan | Vaginal swabs | Atopobium, Megasphaera, Gardneralla, Prevotella ↑* | The vaginal microbiota of the RIF group had lower levels of Lactobacillus than in the control group. |
| Healthy controls (n = 21) | Lactobacillus ↓* | The dysbiotic microbiota from the RIF group consisted of significantly higher levels of 4 other genera. | |||
| Diaz-Martinez et al. (2021) [63] | IVF patients with a RIF history (n = 23) | Spain | Vaginal swabs | Genus | There was no significant difference in Lactobacillus between the groups with RIF in history and without. |
| Streptococcus ↓ | |||||
| IVF patients without RIF in history (n = 25) | Preovotella spp., Ureaplasma spp. and Dialister spp. ↑ | ||||
| Species | |||||
| L. helveticus ↑ | |||||
| Patel et al. (2022) [67] | Women with RIF (n = 10), unexplained infertility (n = 10) and healthy controls (n = 10) | India | Vaginal swabs | The dominance of Lactobacillaceae in all the groups Lactobacillus iners AB-1 dominance in the unexplained infertility group. | There was no difference in lactobacillus between the group with RIF, unexplained infertility, and healthy controls. |
| Leptotrichia and Sneathia ↑ in control group * | |||||
| ENDOMETRIUM | |||||
| Kitaya et al. (2019) | RIF patients (n = 28) | Japan | Endometrial fluid | Genus | The endometrial fluid microbiota showed a significant difference in community composition between the RIF group and the control group. Burkholderia was only detected in the RIF group. |
| Lactobacillus (>90%) dominance Gardnerella ↑ | |||||
| Control (infertile patients undergoing first IVF attempt n = 18) | Burkholderia ↑* | ||||
| Ichiyama et al. (2021) [65] | Women with RIF (n = 145) | Japan | Endometrial tissue with Pipette | Atopobium, Megasphaera, Gardnerella, Prevotella, Schlegelella, Delftia, Burkholderia, Sphingobacterium, Dietzia, Enterococcus, Micrococcus, Ralstonia, Leucobacter, and Hydrogenophaga ↑* | The dysbiotic microbiota from the RIF group consisted of significantly higher levels of 14 other genera. |
| Healthy controls (n = 21) | |||||
| Diaz-Martinez et al. (2021) [63] | Patients with a RIF history (n = 23) | Spain | Transcervical endometrial tissue (Tao Brush IMC Endometrial Sampler) | Genus | The RIF group had significantly different compositions of the endometrial fluid microbiota. A higher abundance of Prevotella was detected in RIF patients. |
| Prevotella and Sneathia amnii ↑* | |||||
| and without RIF in history (n = 25) | Species | ||||
| L. helveticus ↑* | |||||
| Keburiya et al. (2022) | Women with RIF (n = 91) | Russia | Endometrial tissue (Embryo transfer catheter tips) | Obligate anaerobes (streptococci, enterobacteria) were found in the RIF group but with low concentrations and were insignificant. | G. Vaginalis was significantly higher in the naïve IVF patients but did not have a significant impact on embryo implantation. There was no significant difference in Lactobacillus between the RIF group and naïve IVF patients. |
| Women with first IVF attempt (n = 39) | |||||
| Chen et al. (2022) [75] | Women with RIF (n = 75) | China | Transcervical endometrial tissue | Phylum | There was a significant difference in community composition at the genus level of the RIF group compared to the control group. The dominance of Sphingomonas was found in the endometrial microbiota of the RIF group |
| Dominance of Firmicutes, Proteobacteria, Bacteroidetes, Cyanobacteria, and Actinobacteria | |||||
| Healthy controls (n = 36) | Genus | ||||
| Sphingomonas, Brevundimonas, DMER64, | |||||
| Methylobacterium, Rhodoferax, Caulobacter ↑* | |||||
| Lactobacillus, Pseudomonas, Delftia ↓* | |||||
| Vomstein et al. (2022) [77] | Women with RIF (n = 20) | Finland | Transcervical endometrial tissue | Phylum | Women with RIF fail to attenuate their endometrial microbiome diversity and richness throughout the menstrual cycle |
| Firmicutes further decrease after ovulation. | |||||
| Healthy controls (n = 10) | Proteobacteria increase after ovulation. | ||||
| Bacteroidetes peak in the follicular phase. | |||||
| Gardnerella and Dialister were increased around ovulation | |||||
| Cela et al. (2022) [78] | Women with RIF (n = 26) | Italy | Transcervical endometrial tissue | Genus | Lactobacillus abundance is found to be negatively correlated with anti-inflammatory cytokines and positively correlated with pro-inflammatory cytokines. |
| With LD (n = 13) | Lactobacillus ↓* | ||||
| And NLDM (n = 13) | Gardnerella, Streptococcus and Bifidobacterium ↑ | ||||
| Zou et al. (2023) [71] | Women with (n = 141) | China | Transcervical endometrial tissue | Genus | Diverse pathogenic bacteria were found in high abundance in the endometrial microbiota of women with RIF. |
| Streptococcus, Staphylococcus, Neisseria, and Klebsiella ↑ | |||||
| Iwami et al. (2023) | RIF patients with endometrial microbiota data (n = 131): n = 30 (23%) with abnormal microbiota. | Japan | Endometrial tissue with pipelle | Genus | Treatment with probiotics in women with RIF and abnormal endometrial microbiota might improve IVF outcomes. |
| Streptococcus, Gardnerella, Atopobium and Bifidobacterium ↑ | |||||
| Lozano et al. (2023) | Women with RIF (n = 27) | Spain | Transcervical endometrial tissue (Tao Brush IUMC Endometrial sampler) | Genus | Lactobacillus is adversely associated with pathogenic bacteria (Prevotella, Dialister, and Streptococcus), and this dysbiosis might be the cause of an increased risk of implantation failure. |
| Lactobacillus ↑* (control 97.96% vs. RIF 92.27%) | |||||
| Women without RIF (n = 18) | Prevotella ↑* (control 0.00% vs. RIF 2.19%) | ||||
| Dialister ↑* (control 0.06% vs. RIF 0.15%) | |||||
| Streptococcus ↑* (control 0.05% vs. RIF 0.18%) | |||||
| VAGINA | |||||
|---|---|---|---|---|---|
| First Author (Year) | Study Group | Country | Specimen Type | Composition: Phylum/Genus/Species | Conclusion |
| Kuon et al. (2017) [95] | Women with RPL (n = 243) | Germany | Vaginal swabs | Genus | RPL patients with elevated peripheral natural killer cells suffer more often from colonization by G. vaginalis and gram-negative anaerobes. |
| Group B Streptococcus ↑ | |||||
| Enterobacteriaceae ↑ | |||||
| Species | |||||
| Gardnerella vaginalis ↑ | |||||
| Zhang et al. (2019) [96] | History of recurrent miscarriages (n = 10) | China | Vaginal swabs | Phylum | The dysbiotic microbiota from the RPL group consisted of an increased abundance of Atopobium, Prevotella, and Streptococcus and a decreased abundance of Lactobacillus. |
| Firmicutes ↑* | |||||
| Actinobacteria ↓* | |||||
| Bacteroidetes ↓* | |||||
| Healthy controls (n = 10) | Genus | ||||
| Atopobium, Prevotella and Streptococcus ↑* | |||||
| Lactobacillus ↓ | |||||
| Al-Memar et al. (2020) [30] | Women with first term miscarriages (n = 64) | UK | Vaginal swabs | Genus | A reduced abundance of Lactobacillus was found in first-trimester miscarriages compared to women with viable pregnancies. Community State Type IV was more often found in miscarriages. |
| Streptococcus, Prevotella, Ureaplasma, Peptoniphilus, and Dialaster ↑ | |||||
| Women with full term pregnancies (n = 83) | Lactobacillus spp. ↓ | ||||
| Fan et al. (2020) [94] | Women with unexplained recurrent spontaneous abortion (n = 31) | China | Vaginal swabs | Phylum | The vaginal microbiota of women with unexplained RPL has much higher alpha diversity and a higher abundance of Proteobacteria. |
| Proteobacteria ↑* | |||||
| Normal pregnancy induced abortion (n = 27) | Genus | ||||
| ↑ Gammaproteobacteria, Proteobacteria, Pseudomonas, Moraxella, Ruminococcus, Collinsella aerofaciens, Alteromonadaceae, Cellvibrio, Arthrobacter, Roseburia, and Micrococcaceae | |||||
| Peuranpaa et al. (2022) [99] | Women with RPL (n = 47) | Finland | Vaginal swabs | Species | The vaginal microbiota of women with RPL has a higher abundance of G. vaginalis compared with healthy controls. |
| Healthy controls (n = 39) | Gardnerella vaginalis ↑ | ||||
| Caliskan et al. (2022) | Women with recurrent miscarriages (n = 25) | Turkey | Vaginal swabs | Genus | A decrease in Lactobacillus spp. And an increase in anaerobic microorganisms were found in the vaginal microbiota of women with RPL. |
| Lactobacillus ↓ | |||||
| Healthy controls (n = 25) | Enterobacterium spp., Eubacterium spp. | ||||
| Megasphaera spp. and Sneathia spp. ↑ | |||||
| Jiao et al. (2022) [97] | History of recurrent miscarriages (n = 16) | China | Vaginal swabs | Genus | A decrease in Lactobacillus and an increase in the abundance of Atopobium and Prevotella were found in the vaginal microbiota of women with RPL. |
| Lactobacillus ↓* | |||||
| Healthy controls (n = 20) | Gardnerella ↓* | ||||
| Atopobium ↑* | |||||
| Liu et al. (2022) [103] | Women with RPL (n = 25, with 50% chronic endometritis) | China | Vaginal swabs | Genus | Lactobacillus was the dominant genus in the RPL and control groups. No significant difference was found between the vaginal microbiota of the two groups. |
| Lactobacillus (>90%) | |||||
| Bidifidobacterium ↓ | |||||
| Healthy controls (n = 25, with 24% chronic endometritis) | Gardnerella spp. ↓ | ||||
| Atopobium ↑ | |||||
| Ncib et al. (2022) | Women with unexplained RPL (n = 65) | Italia | Vaginal swabs | Genus | There is a high prevalence of aerobic vaginitis-causing bacteria in women with RPL (65%). |
| Enterococcus, | |||||
| Healthy controls with at least two live births and no miscarriages (n = 50) | Staphylococcus, | ||||
| Streptococcus ↑ | |||||
| Mori et al. (2023) [101] | Patients with unexplained RPL (n = 88) | Japan | Vaginal swabs | Genus | There is no difference in Lactobacillus and bacterial diversity between patients with RPL and live birth. L. iners dominance of the vaginal microbiota in the Japanese study population. |
| Healthy controls with no history of miscarriage (n = 17) | Dominance of Lactobacillus iners, Gardnerella vaginalis, Atopobium vaginae and Bifidobacterium breve. | ||||
| ENDOMETRIUM | |||||
| First Author (Year) | Study group | Country | Sampling | Composition: Phylum/Genus/Species | Conclusion |
| Verstraelen et al. (2016) | 19 women with RIF (n = 11), RPL (n = 7) or both (n = 1) | Belgium | Transcervical endometrial tissue (Tao Brush IUMC Endometrial sampler) | Phylum | 90% of the women with either RPL or RIF had a uterine microbiota with a predominance of the phylum Bacteroidetes. |
| Bacteroidetes ↑ | |||||
| Proteobacteria ↑ | |||||
| Species | |||||
| Bacteroides xylanisolvens, Bacteroides thetaiotaomicron, Bacteroides fragilis ↑ | |||||
| Peuranpaa et al. (2022) [99] | Women with RPL (n = 47) | Finland | Transcervical endometrial tissue | Species | L. crispatus is less abundant in women with RPL compared to controls (17.2% versus 45.6%). G. vaginalis is more abundant in the RPL group. |
| L. crispatus ↓ | |||||
| Healthy controls (n = 39) | L. iners dominance | ||||
| Gardnerella vaginalis ↑ | |||||
| Liu et al. (2022) [103] | Women with RPL (n = 25, with 50% chronic endometritis) | China | Transcervical endometrial tissue | Phylum | A lower abundance of Lactobacillus spp. was found, and Acinetobacter spp. were predominant. |
| Healthy controls (n = 25, with 24% chronic endometritis) | Proteobacteria ↑ | ||||
| Genus | |||||
| Lactobacillus (<10%) | |||||
| Anaerobacillus ↑* | |||||
| Erysipelothrix ↑* | |||||
| Hydrogenophilus↑* | |||||
| Shi et al. (2022) [107] | Patients with RPL (n = 63) | Japan | Transcervical endometrial tissue | Genus | A higher abundance of Ureaplasma species in the endometrium was associated with an increased risk of miscarriage with an euploid karyotype. |
| Ureaplasma spp. ↑ | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Louwers, Y.V.; Laven, J.S.E.; Schoenmakers, S. Clinical Relevance of Vaginal and Endometrial Microbiome Investigation in Women with Repeated Implantation Failure and Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2024, 25, 622. https://doi.org/10.3390/ijms25010622
Gao X, Louwers YV, Laven JSE, Schoenmakers S. Clinical Relevance of Vaginal and Endometrial Microbiome Investigation in Women with Repeated Implantation Failure and Recurrent Pregnancy Loss. International Journal of Molecular Sciences. 2024; 25(1):622. https://doi.org/10.3390/ijms25010622
Chicago/Turabian StyleGao, Xushan, Yvonne V. Louwers, Joop S. E. Laven, and Sam Schoenmakers. 2024. "Clinical Relevance of Vaginal and Endometrial Microbiome Investigation in Women with Repeated Implantation Failure and Recurrent Pregnancy Loss" International Journal of Molecular Sciences 25, no. 1: 622. https://doi.org/10.3390/ijms25010622