

Berberine Protects against High-Energy and Low-Protein Diet-Induced Hepatic Steatosis: Modulation of Gut Microbiota and Bile Acid Metabolism in Laying Hens
 ,
,
Abstract
:1. Introduction
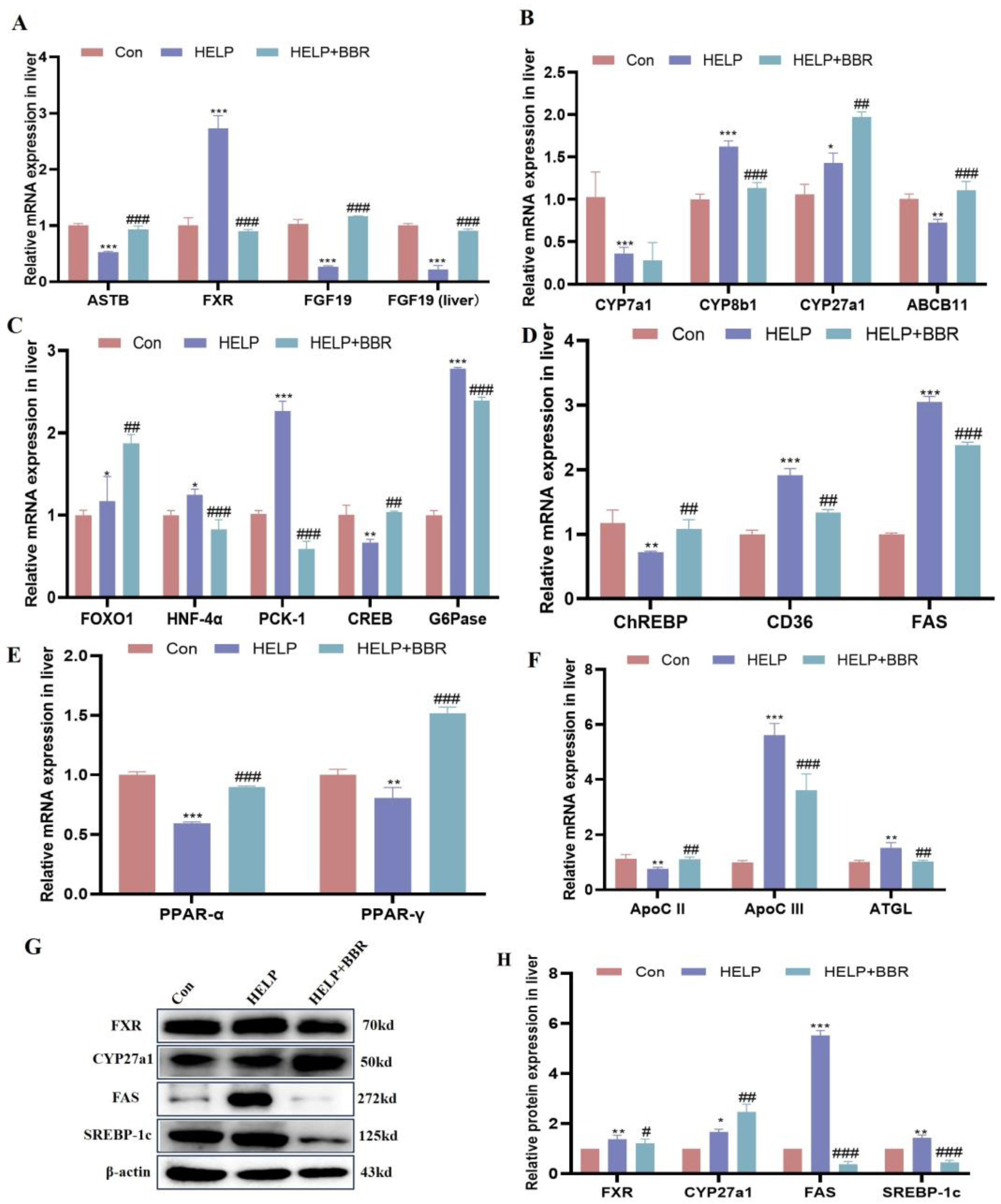
2. Results
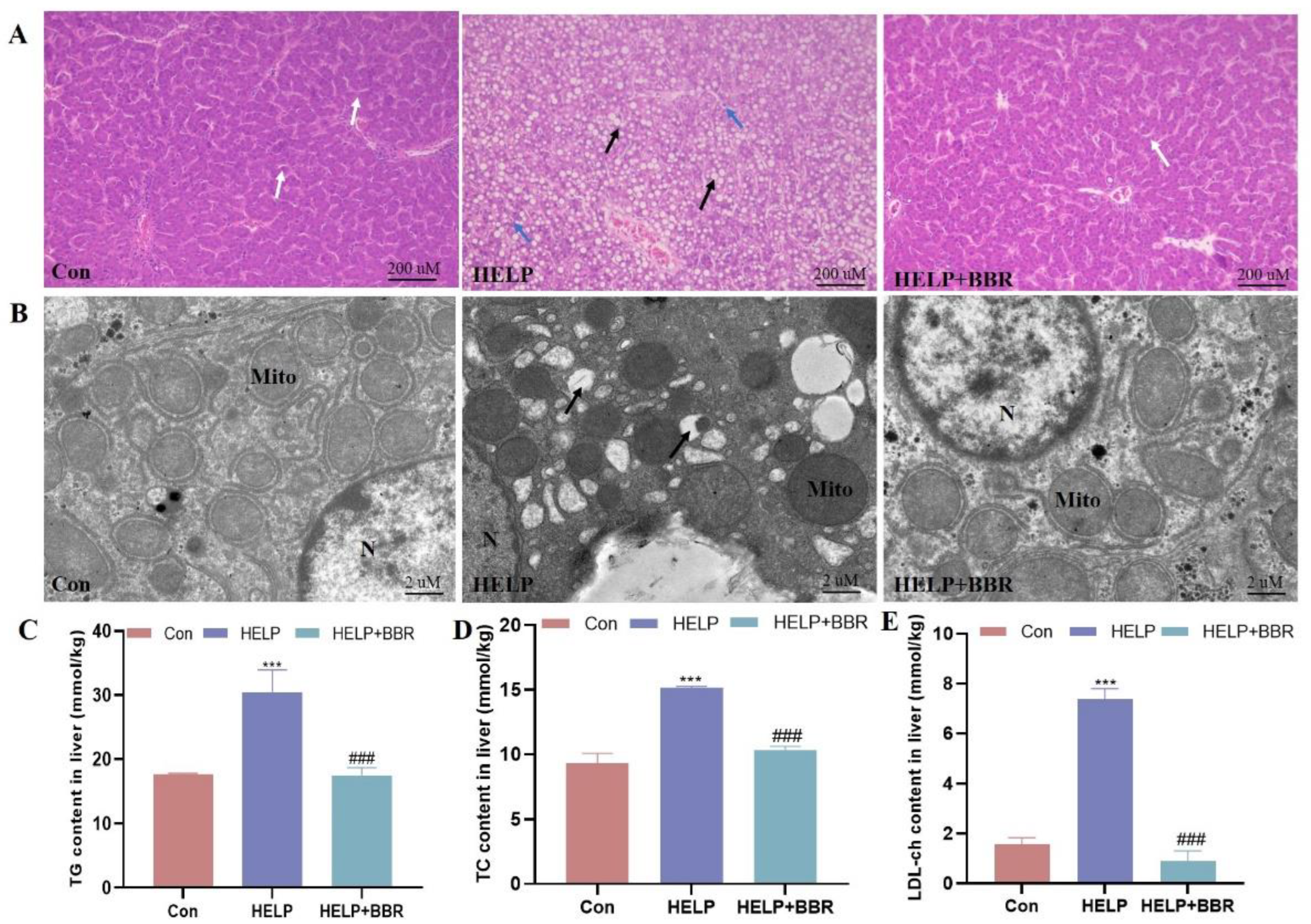
2.1. BBR Alleviated HELP-Induced Blood Lipid Metabolism and Hepatic Lipid Deposition
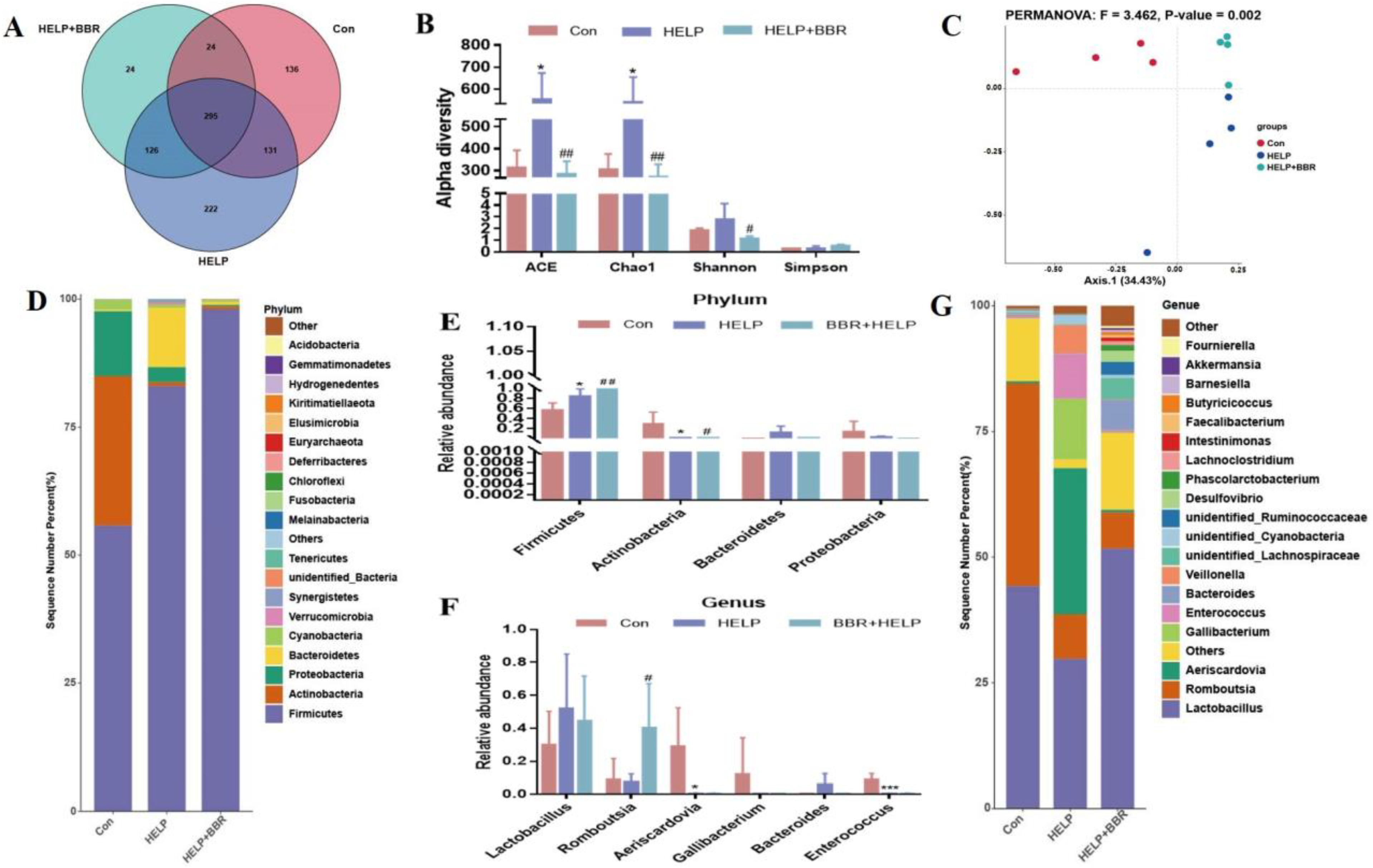
2.2. BBR Alters the Composition of Gut Microbiota in HELP-Fed Laying Hens
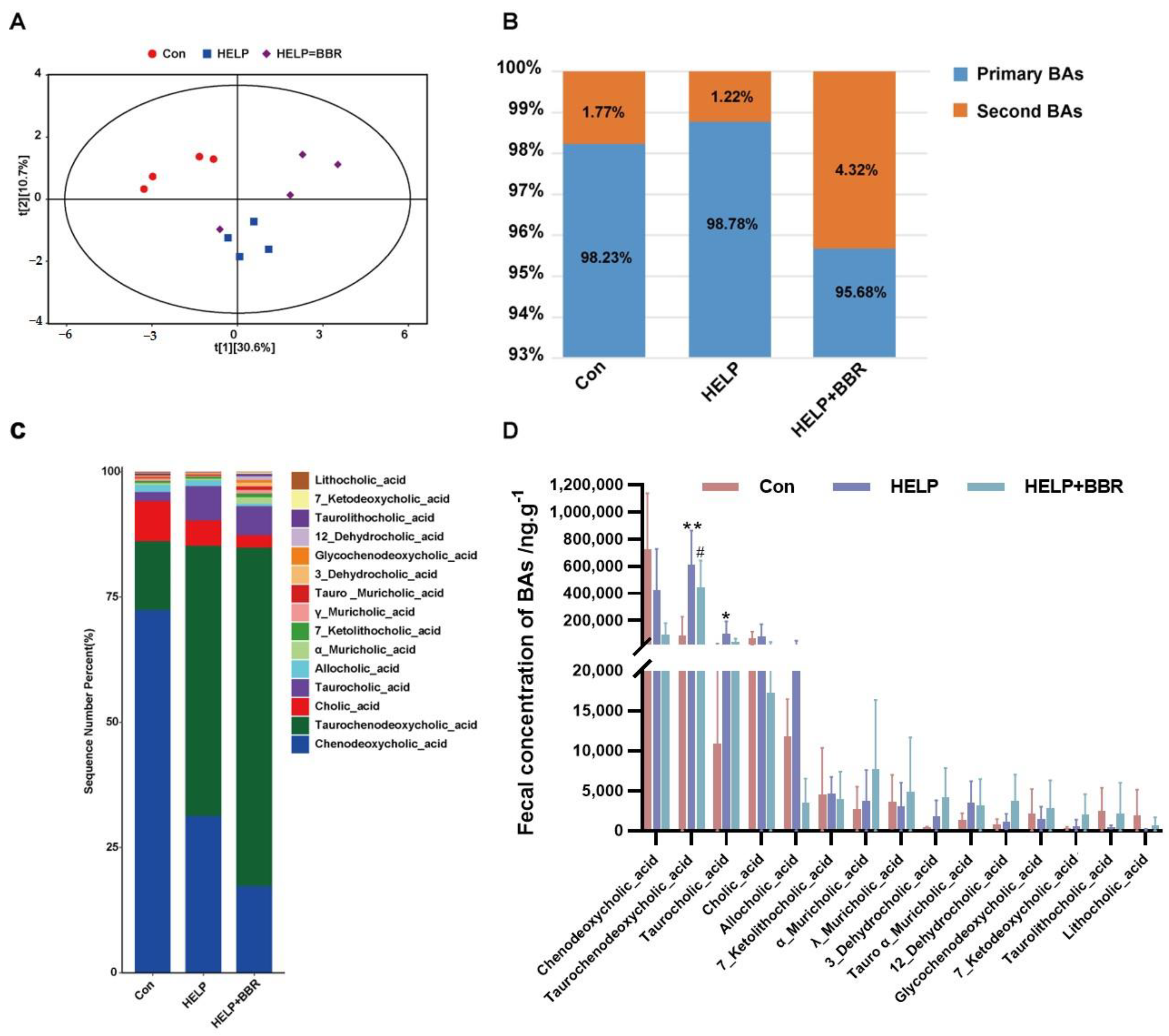
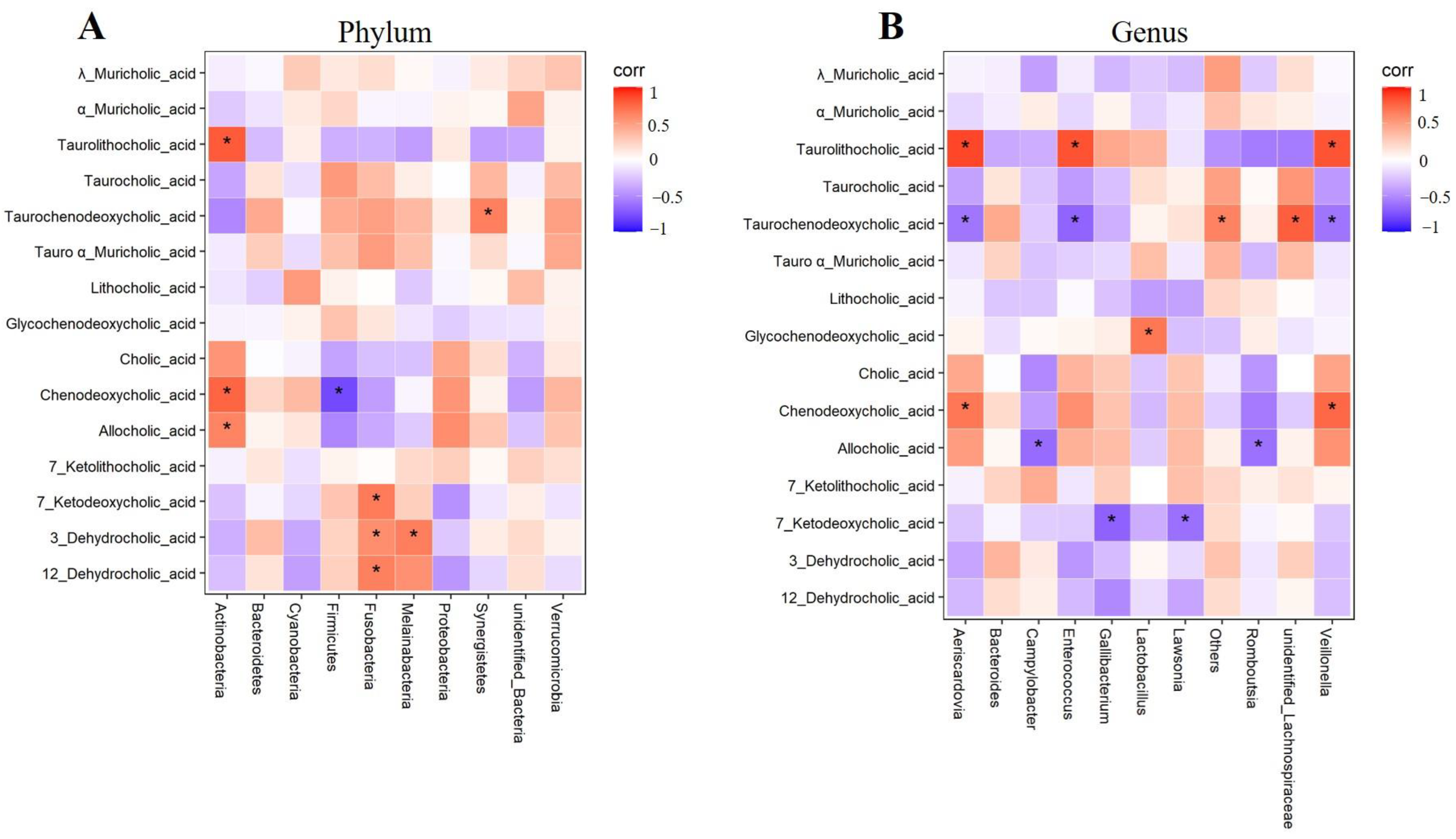
2.3. Regulation of BBR on Bile Acid Metabolism Disorder in Chicken Feces
2.4. BBR Alleviated HELP-Induced Abnormal Bile Acid Biosynthesis
2.5. BBR Alleviated HELP-Induced Abnormal Glucose and Lipid Metabolism
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Histopathological Examination
4.3. Determination of Liver Biochemical Indexes
4.4. Sequencing of 16S rDNA
4.5. Quantitative Analysis of BA Profile in Ileal Feces
4.6. Quantitative Real-Time PCR Analysis
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, W.; Shi, Y.; Li, G.; Huang, C.; Zhuang, Y.; Shu, B.; Cao, X.; Li, Z.; Hu, G.; Liu, P.; et al. Preparation of the peroxisome proliferator-activated receptor alpha polyclonal antibody: Its application in fatty liver hemorrhagic syndrome. Int. J. Biol. Macromol. 2021, 182, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Grimes, J.L.; Maurice, D.V.; Lightsey, S.F.; Bridges, W.J. Research note: Relationship of comb color to liver appearance and fat content in Single Comb White Leghorn laying hens. Poult. Sci. 1991, 70, 2544–2546. [Google Scholar] [CrossRef] [PubMed]
- Rozenboim, I.; Mahato, J.; Cohen, N.A.; Tirosh, O. Low protein and high-energy diet: A possible natural cause of fatty liver hemorrhagic syndrome in caged White Leghorn laying hens. Poult. Sci. 2016, 95, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Diaz, G.J.; Squires, E.J.; Julian, R.J. The use of selected plasma enzyme activities for the diagnosis of fatty liver-hemorrhagic syndrome in laying hens. Avian Dis. 1999, 43, 768–773. [Google Scholar] [CrossRef]
- Zhuang, Y.; Xing, C.; Cao, H.; Zhang, C.; Luo, J.; Guo, X.; Hu, G. Insulin resistance and metabonomics analysis of fatty liver haemorrhagic syndrome in laying hens induced by a high-energy low-protein diet. Sci. Rep. 2019, 9, 10141. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Sun, Y.; Zheng, X.; Yang, Y. Gastrodin attenuates perfluorooctanoic acid-induced liver injury by regulating gut microbiota composition in mice. Bioengineered 2021, 12, 11546–11556. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Ohashi, S.; Sasaki, K.; Nagata, R.; Pelpolage, S.; Fukuma, N.; Reed, J.D.; Shimada, K.I.; Kadoya, N.; Fukushima, M. Dietary adzuki bean paste dose-dependently reduces visceral fat accumulation in rats fed a normal diet. Food Res. Int. 2020, 130, 108890. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Marsh, S.; Hu, J.; Feng, W.; Wu, C. The pathogenesis of nonalcoholic fatty liver disease: Interplay between diet, gut microbiota, and genetic background. Gastroenterol. Res. Pract. 2016, 2016, 2862173. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J. MAFLD: A consensus-driven proposed nomenclature for metabolic associated fatty liver disease. Gastroenterology 2020, 158, 1999–2014. [Google Scholar] [CrossRef]
- Marchesini, G.; Brizi, M.; Bianchi, G.; Tomassetti, S.; Bugianesi, E.; Lenzi, M.; McCullough, A.J.; Natale, S.; Forlani, G.; Melchionda, N. Nonalcoholic fatty liver disease: A feature of the metabolic syndrome. Diabetes 2001, 50, 1844–1850. [Google Scholar] [CrossRef]
- Vuppalanchi, R.; Chalasani, N. Nonalcoholic fatty liver disease and nonalcoholic steatohepatitis: Selected practical issues in their evaluation and management. Hepatology 2009, 49, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Liu, D.; Gao, Q.; Ni, J.; Qian, L.; Ni, Y.; Fang, Q.; Jia, W.; Li, H. Bifidobacterium adolescents alleviates liver steatosis and steatohepatitis by increasing fibroblast growth factor 21 sensitivity. Front. Endocrinol. 2021, 12, 773340. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered fecal microbiota correlates with liver biochemistry in nonobese patients with Non-alcoholic fatty liver disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef] [PubMed]
- Abilos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef]
- Juarez-Hernandez, E.; Chavez-Tapia, N.C.; Uribe, M.; Barbero-Becerra, V.J. Role of bioactive fatty acids in nonalcoholic fatty liver disease. Nutr. J. 2016, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Presides, G.A.; Keaton, M.A.; Campeau, P.M.; Bessard, B.C.; Conner, M.E.; Hotez, P.J. The undernourished neonatal mouse metabolome reveals evidence of liver and biliary dysfunction, inflammation, and oxidative stress. J. Nutr. 2014, 144, 273–281. [Google Scholar] [CrossRef]
- Ichimura-Shimizu, M.; Watanabe, S.; Kashirajima, Y.; Nagatomo, A.; Wada, H.; Tsuneyama, K.; Omagari, K. Dietary cholic acid exacerbates liver fibrosis in NASH model of sprague-dawley rats Fed a high-fat and high-cholesterol diet. Int. J. Mol. Sci. 2022, 23, 9268. [Google Scholar] [CrossRef]
- Huang, W.; Ma, K.; Zhang, J.; Qatan ani, M.; Cuvillier, J.; Liu, J.; Dong, B.; Huang, X.; Moore, D.D. Nuclear receptor-dependent bile acid signaling is required for normal liver regeneration. Science 2006, 312, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Safari, H.; Kaczorowski, N.; Felder, M.L.; Brannon, E.R.; Varghese, M.; Singer, K.; Eniola-Adefeso, O. Biodegradable, bile salt microparticles for localized fat dissolution. Sci. Adv. 2020, 6, eabd8019. [Google Scholar] [CrossRef]
- Sinal, C.J.; Tohkin, M.; Miyata, M.; Ward, J.M.; Lambert, G.; Gonzalez, F.J. Targeted disruption of the nuclear receptor FXR/BAR impairs bile acid and lipid homeostasis. Cell 2000, 102, 731–744. [Google Scholar] [CrossRef]
- Li, R.; Palmiotti, A.; de Vries, H.D.; Hovingh, M.V.; Koehorst, M.; Mulder, N.L.; Zhang, Y.; Kats, K.; Bloks, V.W.; Fu, J.; et al. Low production of 12alpha-hydroxylated bile acids prevents hepatic steatosis in Cyp2c70(-/-) mice by reducing fat absorption. J. Lipid. Res. 2021, 62, 100134. [Google Scholar] [CrossRef]
- Park, M.Y.; Kim, S.J.; Ko, E.K.; Ahn, S.H.; Seo, H.; Sung, M.K. Gut microbiota-associated bile acid deconjugation accelerates hepatic steatosis in ob/ob mice. J. Appl. Microbiol. 2016, 121, 800–810. [Google Scholar] [CrossRef]
- Yao, J.; Zhou, C.S.; Ma, X.; Fu, B.Q.; Tao, L.S.; Chen, M.; Xu, Y.P. FXR agonist GW4064 alleviates endotoxin-induced hepatic inflammation by repressing macrophage activation. World J. Gastroenterol. 2014, 20, 14430–14441. [Google Scholar] [CrossRef]
- Tarantino, G.; Balsano, C.; Santini, S.J.; Brienza, G.; Clemente, I.; Cosimini, B.; Sinatti, G. It is high time physicians thought of natural products for alleviating NAFLD. Is there sufficient evidence to use them? Int. J. Mol. Sci. 2021, 22, 13424. [Google Scholar] [CrossRef]
- Pirillo, A.; Catapano, A.L. Berberine, a plant alkaloid with lipid- and glucose-lowering properties: From in vitro evidence to clinical studies. Atherosclerosis 2015, 243, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Hao, J.; Fan, D. Biological properties and clinical applications of berberine. Front. Med. 2020, 14, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Berberine pharmacology and the gut microbiota: A hidden therapeutic link. Pharmacol. Res. 2020, 155, 104722. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Huang, K.; Bai, Y.; Feng, X.; Gong, L.; Wei, C.; Huang, H.; Zhang, H. Dietary supplementation with berberine improves growth performance and modulates the composition and function of cecal microbiota in yellow-feathered broilers. Poult. Sci. 2021, 100, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Dehau, T.; Cherlet, M.; Croubels, S.; van Immerseel, F.; Goossens, E. A high dose of dietary berberine improves gut wall morphology, despite an expansion of Enterobacteriaceae and a reduction in beneficial microbiota in broiler chickens. mSystems 2023, 8, e0123922. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Yang, N.; Kong, B.; Cao, B.; Feng, D.; Yu, X.; Ge, C.; Huang, J.; Shen, J.; Wang, P.; et al. Orally administered berberine modulates hepatic lipid metabolism by altering microbial bile acid metabolism and the intestinal FXR signaling pathway. Mol. Pharmacol. 2017, 91, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.W.; Jun, M.S.; Yang, H.K.; Lee, B.C. Cellular and molecular mechanisms and effects of berberine on obesity-induced inflammation. Biomedicines 2022, 10, 1739. [Google Scholar] [CrossRef]
- Li, D.; Zheng, J.; Hu, Y.; Hou, H.; Hao, S.; Liu, N.; Wang, Y. Amelioration of intestinal barrier dysfunction by berberine in the treatment of nonalcoholic fatty liver disease in rats. Pharmacogn. Mag. 2017, 13, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Zhang, J.; Lin, S.; Wang, Y.; Chen, Y.; Fan, J. Prolyl endopeptidase gene disruption improves gut dysbiosis and non-alcoholic fatty liver disease in mice induced by a high-fat diet. Front. Cell Dev. Biol. 2021, 9, 628143. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clement, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef]
- Zhou, D.; Pan, Q.; Shen, F.; Cao, H.X.; Ding, W.J.; Chen, Y.W.; Fan, J.G. Total fecal microbiota transplantation alleviates high-fat diet-induced steatohepatitis in mice via beneficial regulation of gut microbiota. Sci. Rep. 2017, 7, 1529. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Feng, S.; He, K.; Guo, L.; Chen, W.; Wang, M.; Zhong, L.; Wu, C.; Peng, X.; et al. Differential effects of dietary white meat and red meat on NAFLD progression by modulating gut microbiota and metabolites in rats. Oxidative Med. Cell. Longev. 2022, 2022, 6908934. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Gao, Z.; Zhang, Q.; Gu, C. The mechanism of berberine alleviating metabolic disorder based on gut microbiome. Front. Cell. Infect. Microbiol. 2022, 12, 854885. [Google Scholar] [CrossRef]
- Yu, M.; Alimujiang, M.; Hu, L.; Liu, F.; Bao, Y.; Yin, J. Berberine alleviates lipid metabolism disorders via inhibition of mitochondrial complex I in gut and liver. Int. J. Biol. Sci. 2021, 17, 1693–1707. [Google Scholar] [CrossRef]
- Yue, S.J.; Liu, J.; Wang, A.T.; Meng, X.T.; Yang, Z.R.; Peng, C.; Guan, H.S.; Wang, C.Y.; Yan, D. Berberine alleviates insulin resistance by reducing peripheral branched-chain amino acids. Am. J. Physiol.-Endocrinol. Metab. 2019, 316, E73–E85. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Sun, X.; Cheng, H.; Xu, S.; Li, D.; Xie, Z. Large yellow tea extract ameliorates metabolic syndrome by suppressing lipogenesis through SIRT6/SREBP1 pathway and modulating microbiota in leptin receptor knockout rats. Foods 2022, 11, 1638. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Cuesta, M.C.; Del, C.R.; Garriga-Garcia, M.; Pelaez, C.; Requena, T. Taxonomic characterization and short-chain fatty acids production of the obese microbiota. Front. Cell. Infect. Microbiol. 2021, 11, 598093. [Google Scholar] [CrossRef]
- Zeng, Q.; Li, D.; He, Y.; Li, Y.; Yang, Z.; Zhao, X.; Liu, Y.; Wang, Y.; Sun, J.; Feng, X.; et al. Discrepant gut microbiota markers for the classification of obesity-related metabolic abnormalities. Sci. Rep. 2019, 9, 13424. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Moreno, M.; Perez-Herrera, A.; Locia-Morales, D.; Dizzel, S.; Meyre, D.; Stearns, J.C.; Cruz, M. Association of gut microbiome with fasting triglycerides, fasting insulin and obesity status in Mexican children. Pediatr. Obes. 2021, 16, e12748. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.W.; Xiao, M.; Wu, X.; Li, X.W.; Li, X.X.; Zhao, T.; Yu, L.; Chen, X.Q. Ilexsaponin A (1) ameliorates diet-induced nonalcoholic fatty liver disease by regulating bile acid metabolism in mice. Front. Pharmacol. 2021, 12, 771976. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Xie, C.; Lv, Y.; Li, J.; Krausz, K.W.; Shi, J.; Brocker, C.N.; Desai, D.; Amin, S.G.; Bisson, W.H.; et al. Intestine-selective farnesoid X receptor inhibition improves obesity-related metabolic dysfunction. Nat. Commun. 2015, 6, 10166. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; An, Y.; Tang, H.; Wang, Y. Alterations of bile acids and gut microbiota in obesity induced by high fat diet in rat model. J. Agric. Food. Chem. 2019, 67, 3624–3632. [Google Scholar] [CrossRef]
- Zhu, K.; Nie, S.; Gong, D.; Xie, M. Effect of polysaccharide from Ganoderma atrum on the serum metabolites of type 2 diabetic rats. Food Hydrocoll. 2016, 53, 31–36. [Google Scholar] [CrossRef]
- He, Z.; Ma, Y.; Yang, S.; Zhang, S.; Liu, S.; Xiao, J.; Wang, Y.; Wang, W.; Yang, H.; Li, S.; et al. Gut microbiota-derived ursodeoxycholic acid from neonatal dairy calves improves intestinal homeostasis and colitis to attenuate extended-spectrum beta-lactamase-producing enteroaggregative Escherichia coli infection. Microbiome 2022, 10, 79. [Google Scholar] [CrossRef]
- Sirvent, A.; Claudel, T.; Martin, G.; Brozek, J.; Kosykh, V.; Darteil, R.; Hum, D.W.; Fruchart, J.C.; Staels, B. The farnesoid X receptor induces very low-density lipoprotein receptor gene expression. FEBS Lett. 2004, 566, 173–177. [Google Scholar] [CrossRef]
- Staels, B.; Fonseca, V.A. Bile acids and metabolic regulation: Mechanisms and clinical responses to bile acid sequestration. Diabetes Care 2009, 32 (Suppl. S2), S237–S245. [Google Scholar] [CrossRef] [PubMed]
- Neuschwander-Tetri, B.A.; Loomba, R.; Sanyal, A.J.; Lavine, J.E.; Van Natta, M.L.; Abdelmalek, M.F.; Chalasani, N.; Dasarathy, S.; Diehl, A.M.; Hameed, B.; et al. Farnesoid X nuclear receptor ligand obeticholic acid for non-cirrhotic, non-alcoholic steatohepatitis (FLINT): A multicentre, randomised, placebo-controlled trial. Lancet 2015, 385, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, W.; Xiao, M.; Yu, L.; Chen, Q.; Hu, X.; Zhao, Y.; Xiong, L.; Chen, X.; Wang, X.; et al. Penthorum chinense Pursh. extract attenuates non-alcholic fatty liver disease by regulating gut microbiota and bile acid metabolism in mice. J. Ethnopharmacol. 2022, 294, 115333. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Wan, M.; Xu, D.; Pan, D.; Xia, H.; Yang, L.; Sun, G. Flaxseed powder attenuates non-alcoholic steatohepatitis via modulation of gut microbiota and bile acid metabolism through gut-liver axis. Int. J. Mol. Sci. 2021, 22, 10858. [Google Scholar] [CrossRef] [PubMed]
- Haczeyni, F.; Poekes, L.; Wang, H.; Mridha, A.R.; Barn, V.; Geoffrey, H.W.; Ioannou, G.N.; Yeh, M.M.; Leclercq, I.A.; Teoh, N.C.; et al. Obeticholic acid improves adipose morphometry and inflammation and reduces steatosis in dietary but not metabolic obesity in mice. Obesity 2017, 25, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.M.; Guo, H.; Yousef, B.A.; Guerram, M.; Hamdi, A.M.; Zhang, L.; Jiang, Z. Role of inflammatory and oxidative stress, cytochrome P450 2E1, and bile acid disturbance in rat liver injury induced by isoniazid and lipopolysaccharide cotreatment. Antimicrob. Agents. Chemother. 2016, 60, 5285–5293. [Google Scholar] [CrossRef]
- Gao, X.; Liu, P.; Wu, C.; Wang, T.; Liu, G.; Cao, H.; Zhang, C.; Hu, G.; Guo, X. Effects of fatty liver hemorrhagic syndrome on the AMP-activated protein kinase signaling pathway in laying hens. Poult. Sci. 2019, 98, 2201–2210. [Google Scholar] [CrossRef]
- Huang, F.; Zheng, X.; Ma, X.; Jiang, R.; Zhou, W.; Zhou, S.; Zhang, Y.; Lei, S.; Wang, S.; Kuang, J.; et al. Theabrownin from Pu-erh tea attenuates hypercholesterolemia via modulation of gut microbiota and bile acid metabolism. Nat. Commun. 2019, 10, 4971. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Huang, F.; Zhao, A.; Lei, S.; Zhang, Y.; Xie, G.; Chen, T.; Qu, C.; Rajani, C.; Dong, B.; et al. Bile acid is a significant host factor shaping the gut microbiome of diet-induced obese mice. BMC Biol. 2017, 15, 120. [Google Scholar] [CrossRef]
- Xie, G.; Wang, Y.; Wang, X.; Zhao, A.; Chen, T.; Ni, Y.; Wong, L.; Zhang, H.; Zhang, J.; Liu, C.; et al. Profiling of serum bile acids in a healthy Chinese population using UPLC-MS/MS. J. Proteome Res. 2015, 14, 850–859. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition of Diet % | Control Group | HELP Group |
|---|---|---|
| Corn | 64.00 | 70.00 |
| Wheat Bran | 2.00 | 1.20 |
| Soybean Meal | 24.00 | 14.58 |
| Fat-Soybean Oil | 0 | 4.22 |
| Calcium | 8.00 | 8.00 |
| * Premix | 2.00 | 2.00 |
| Total | 100.00 | 100.00 |
| Nutrient level | ||
| Crude Protein CP | 15.86 | 12.00 |
| Available Phosphorus (AP) | 0.51 | 0.46 |
| Arginine | 1.03 | 0.74 |
| Methionine | 0.37 | 0.32 |
| Valine | 0.77 | 0.58 |
| Metabolic Energy (kcal/kg) | 2678.99 | 3100.00 |
| Met + Cys | 0.67 | 0.56 |
| Gene Name | Accession Number | Primer Sequences (5′ to 3′) |
|---|---|---|
| FXR | AF49249.7 | Forward: CTCTCGCAAAATGGGGCAGT |
| Reverse: CGCGGGAATTCGATTGGC | ||
| ASBT | AB970773.1 | Forward: ACCATGAAATTGAAACAAGAGTGAA |
| Reverse: TGGGATAACTTTAGCCTGTCCA | ||
| FGF19 | NM_204674.3 | Forward: GCCAGAGGTCTACTCATCGC |
| Reverse: ACCTGCAACATTCTGCGGTA | ||
| CYP7a1 | NM_001001753.2 | Forward: GCTCCGCATGTTCCTGAATG |
| Reverse: ATGGTGTTAGCTTGCGAGGC | ||
| CYP8b1 | NM_001389480.2 | Forward: TACCAAGGGACAGGGAACAAGGAG |
| Reverse: GGAGGCAACACGGCATAGGC | ||
| ABCB11 | XM_046921923.1 | Forward: ATCTTGGCCATCCAGCAAGG |
| Reverse: ACTGGCTCTTGCTCAACAACACC | ||
| G6Pase | BM439740.1 | Forward: TCCAGCACATCCACTCCATCTACC |
| Reverse: TCAACACCAAGCATCCGCAGAAG | ||
| CREB | CAJNRD030001119.1 | Forward: ACCTGCCATTGCCACTGTTACG |
| Reverse: CTCCATCCGTGCCGTTGTTAGAC | ||
| FOXO1 | NM_204328.2 | Forward: ACACAGTGAACCCCATGTCA |
| Reverse: AGGGGCATACGGGTTCATAG | ||
| HNF-4α | AY700581.1 | Forward: AGGATGTCTTGCTGCTAGGG |
| Reverse: GCAGGCGTATTCATTGTCGT | ||
| FAS | AB495724.1 | Forward: ACTGTGGGCTCCAAATCTTCA |
| Reverse: CAAGGAGCCATCGTGTAAAGC | ||
| ATGL | EU240627.2 | Forward: GCTGATCCAGGCCTGTGTCT |
| Reverse: TGGAGGTATCTGCCCACAGTAGA | ||
| PPAR-γ | AB045597.1 | Forward: CACTGCAGGAACAGAACAAAGAA Reverse: TCCACAGAGCGAAACTGACATC |
| CD36 | NM_001030731.1 | Forward: CTGGGAAGGTTACTGCGATT Reverse: GCGAGGAACTGTGAAACGATA |
| ChREBP | EU152408.1 | Forward: GATGAGCACCGCAAACCAGAGG Reverse: TCGGAGCCGCTTCTTGTAGTAGG |
| PPAR-α | AF163809.1 | Forward: GACACCCTTTCACCAGCATC Reverse: CCCTTACAACCTTCACAAGCA |
| ApoC II | CM040951.1 | Forward: CCTCCCAGCTCACCCAATT Reverse: CAGGATCCCGGTGTAAGTCA |
| ApoC III | NM_001302127.2 | Forward: AAGGTGCAGGAGTACGTCAA Reverse: GCGTTGTCTGACAGCCATTT |
| CYP27a1 | XM_040676620.2 | Forward: CTTCCCCAAGAACACCCTCT Reverse: AAGGGATGGAGCTGAAAGGG |
| PCK-1 | NM_205471.2 | Forward: TCAACACCAGATTCCCAGGC Reverse: CCTCATGCTAGCCACCACAT |
| β-actin | L08165.1 | Forward: ATTGCTGCGCTCGTTGTT Reverse: CTTTTGCTCTGGGCTTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Yang, Y.; Chen, J.; Dai, X.; Xing, C.; Zhang, C.; Cao, H.; Guo, X.; Hu, G.; Zhuang, Y. Berberine Protects against High-Energy and Low-Protein Diet-Induced Hepatic Steatosis: Modulation of Gut Microbiota and Bile Acid Metabolism in Laying Hens. Int. J. Mol. Sci. 2023, 24, 17304. https://doi.org/10.3390/ijms242417304
Wang C, Yang Y, Chen J, Dai X, Xing C, Zhang C, Cao H, Guo X, Hu G, Zhuang Y. Berberine Protects against High-Energy and Low-Protein Diet-Induced Hepatic Steatosis: Modulation of Gut Microbiota and Bile Acid Metabolism in Laying Hens. International Journal of Molecular Sciences. 2023; 24(24):17304. https://doi.org/10.3390/ijms242417304
Chicago/Turabian StyleWang, Chang, Yitian Yang, Jinyan Chen, Xueyan Dai, Chenghong Xing, Caiying Zhang, Huabin Cao, Xiaoquan Guo, Guoliang Hu, and Yu Zhuang. 2023. "Berberine Protects against High-Energy and Low-Protein Diet-Induced Hepatic Steatosis: Modulation of Gut Microbiota and Bile Acid Metabolism in Laying Hens" International Journal of Molecular Sciences 24, no. 24: 17304. https://doi.org/10.3390/ijms242417304