
Positive Regulation of S-Adenosylmethionine on Chondrocytic Differentiation via Stimulation of Polyamine Production and the Gene Expression of Chondrogenic Differentiation Factors
 , and
, and
Abstract
:1. Introduction
2. Results
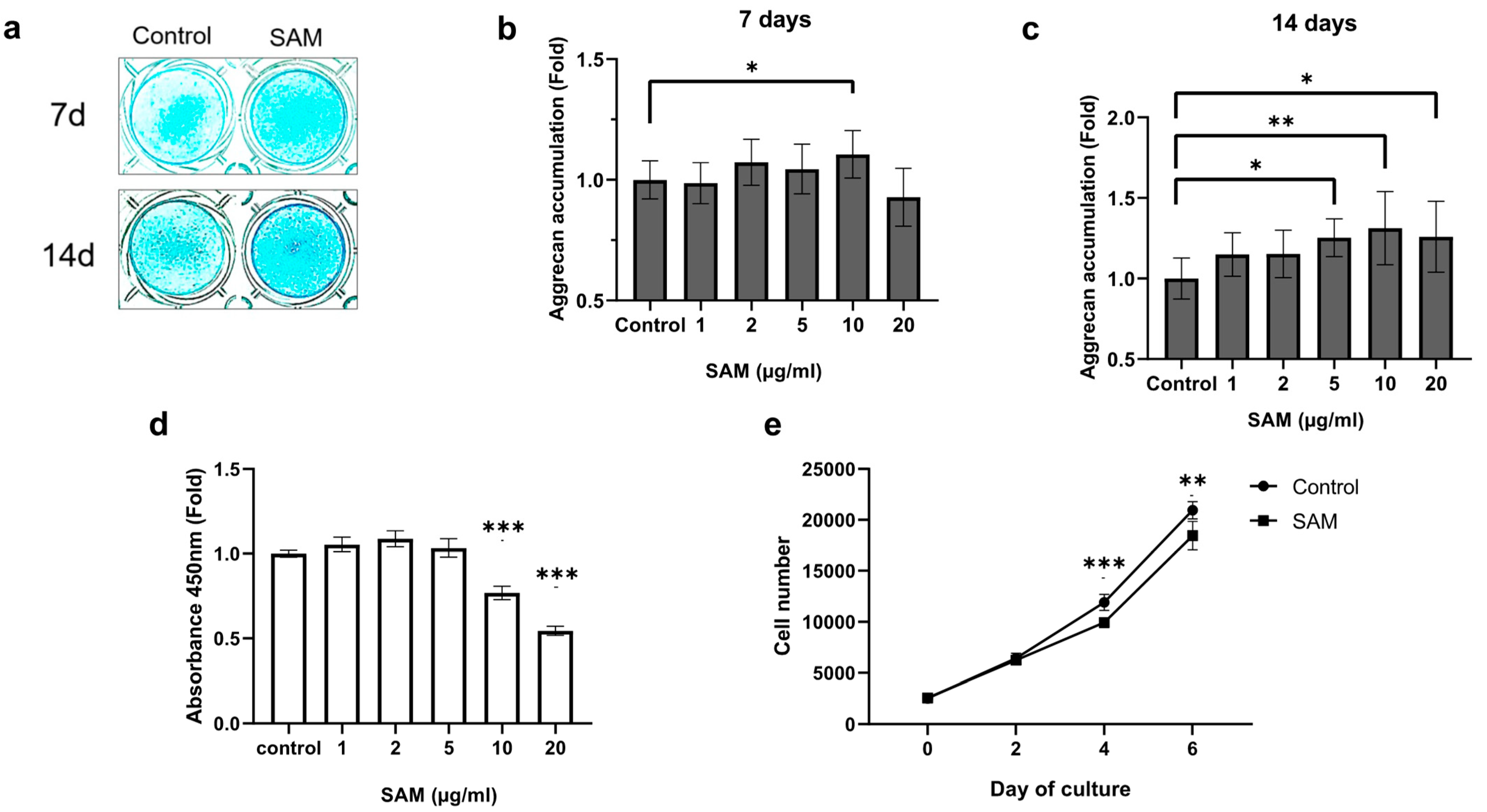
2.1. SAM Enhances Aggrecan Production, but Negatively Regulates Cell Proliferation of Chondrocytic HCS-2/8 Cells
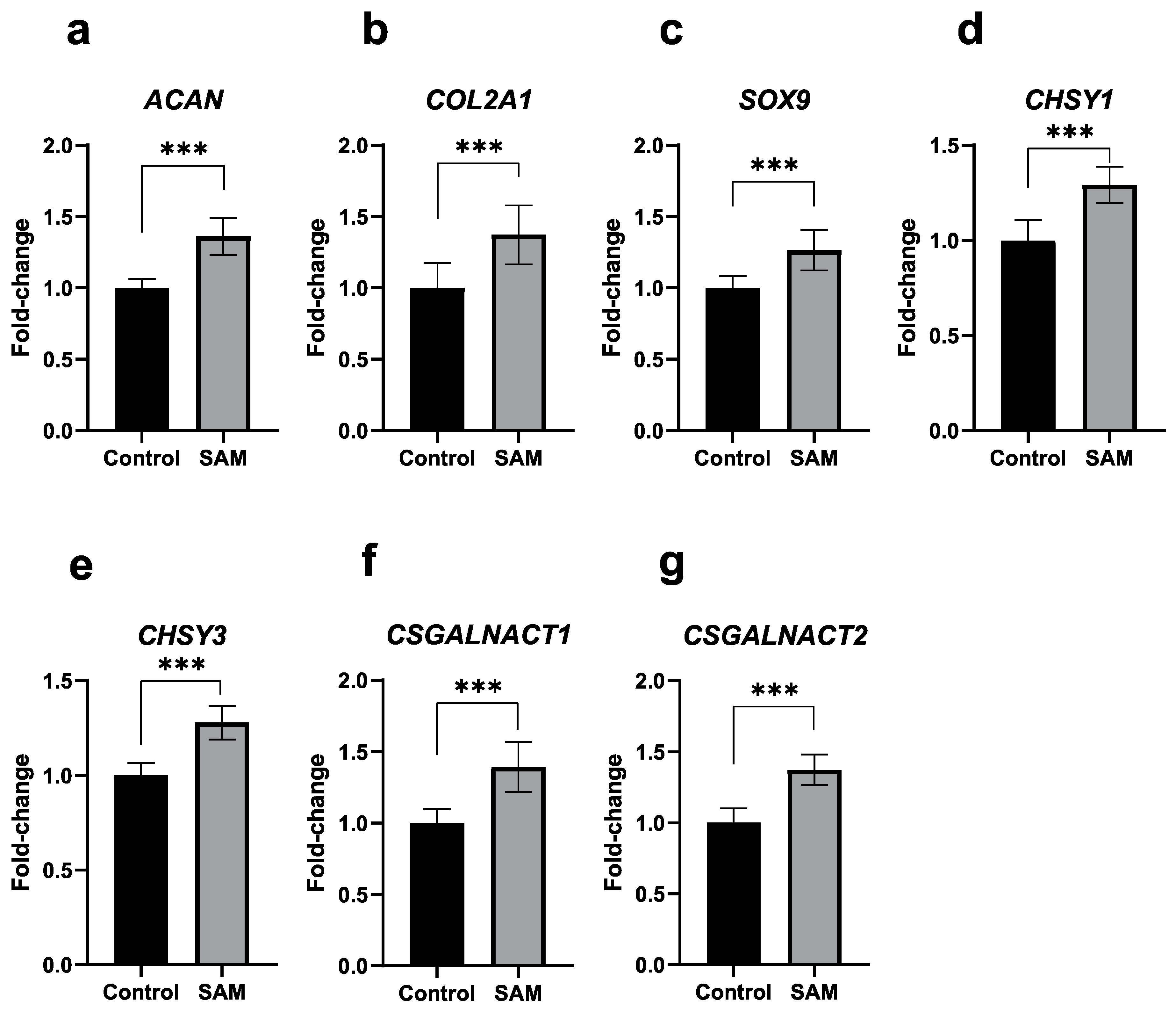
2.2. SAM Enhances the Gene Expression of Cartilage-Specific Markers, Chondrogenesis-Associated Factors, and Chondroitin Sulfate Synthesis-Involved Enzymes in HCS-2/8 Cells
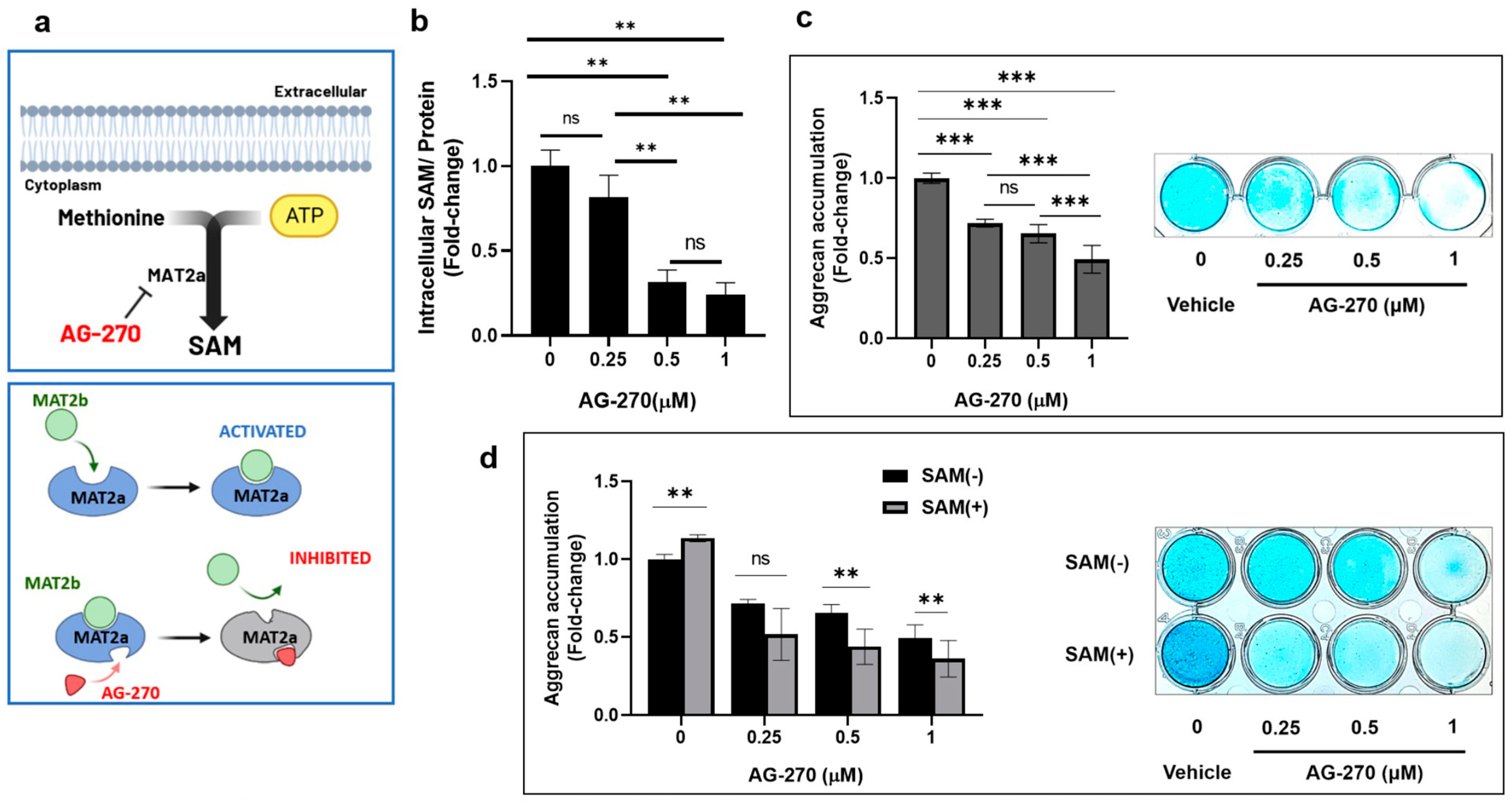
2.3. Inhibition of Intracellular SAM Synthesis Impedes Aggrecan Accumulation in HCS-2/8 Cells
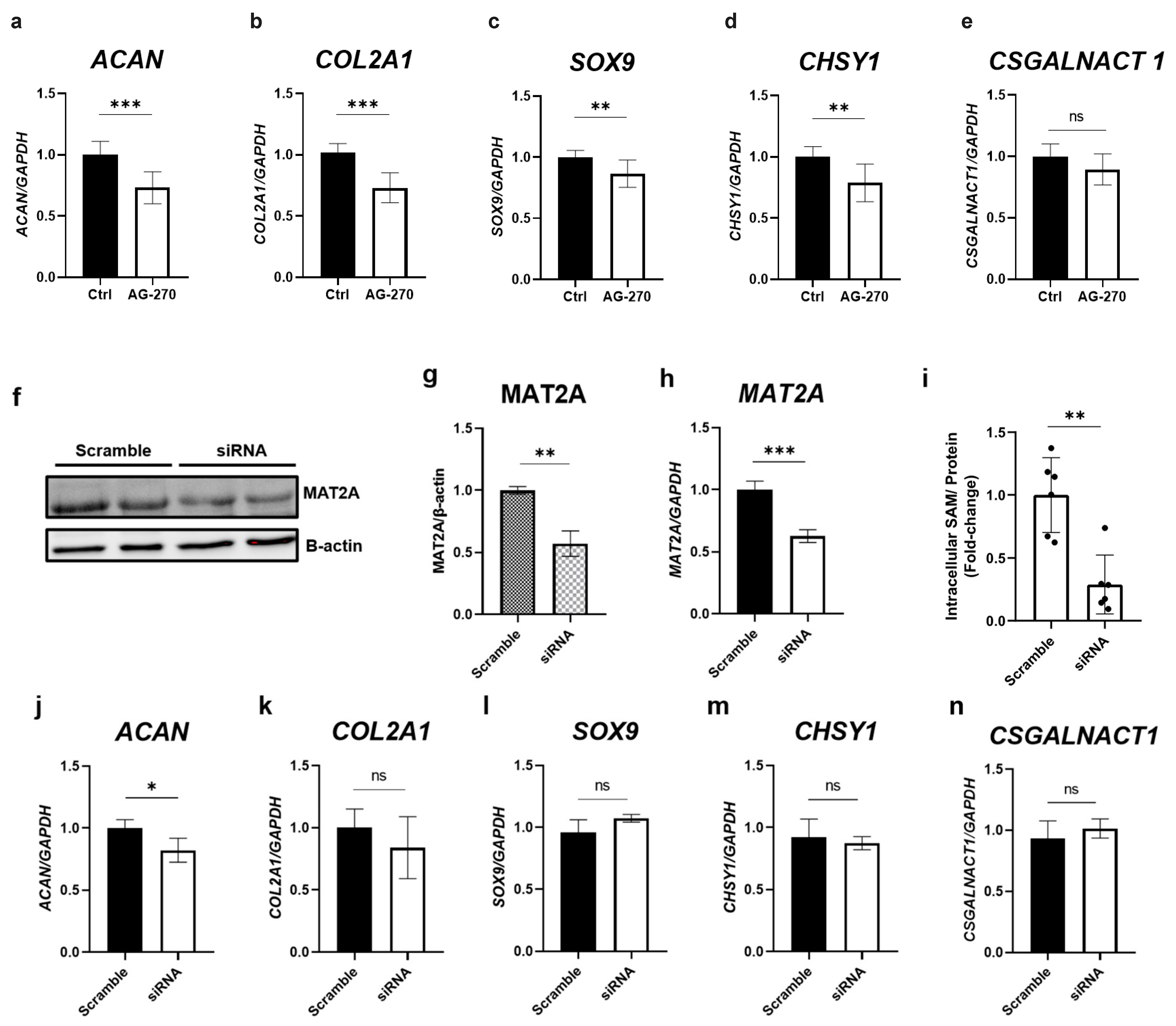
2.4. Inhibition of Intracellular SAM Synthesis Decreases the Gene Expression of Cartilage-Related Factors
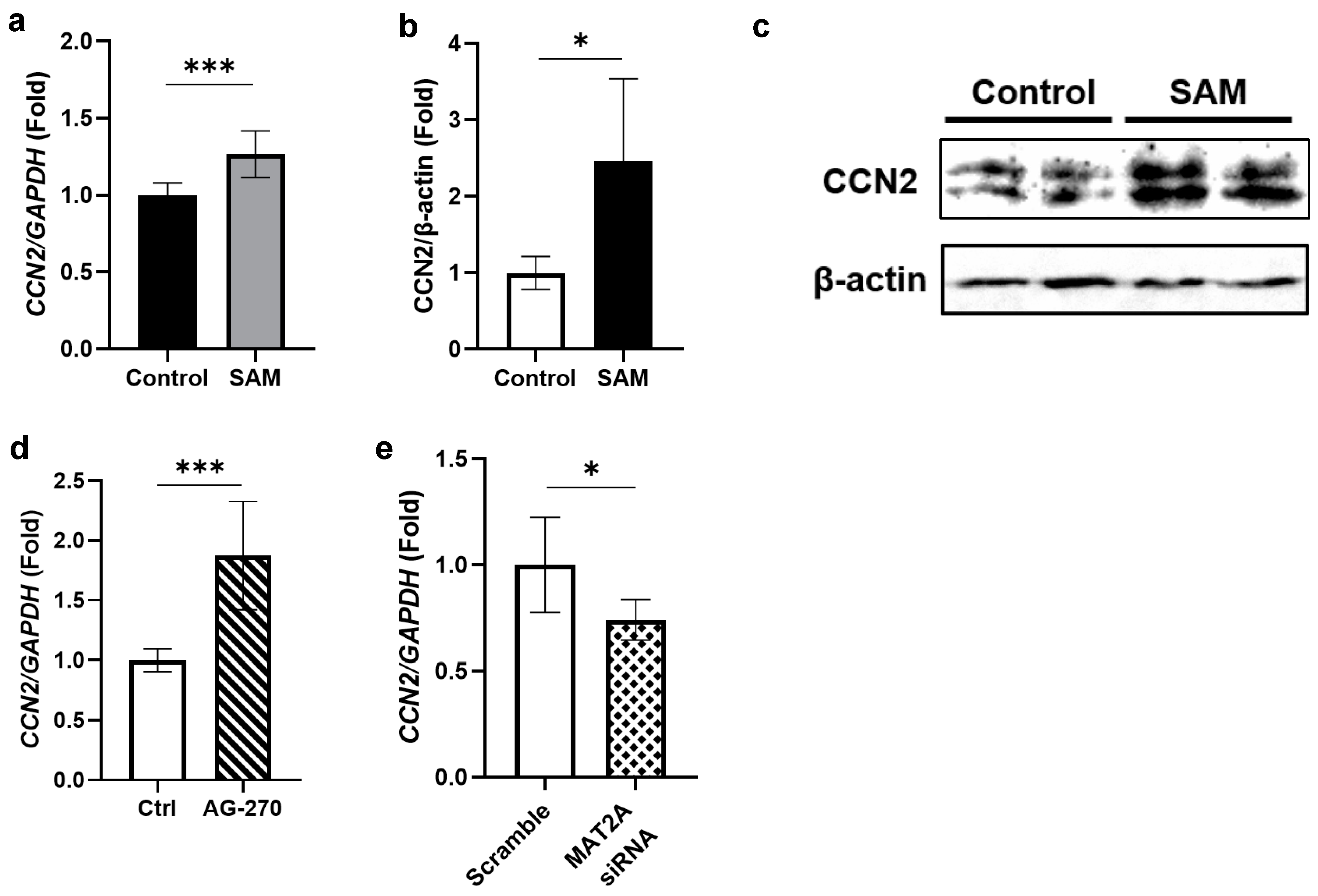
2.5. SAM Has Positive Effects on the Gene Expression and Protein Level of CCN2 in HCS-2/8 Cells
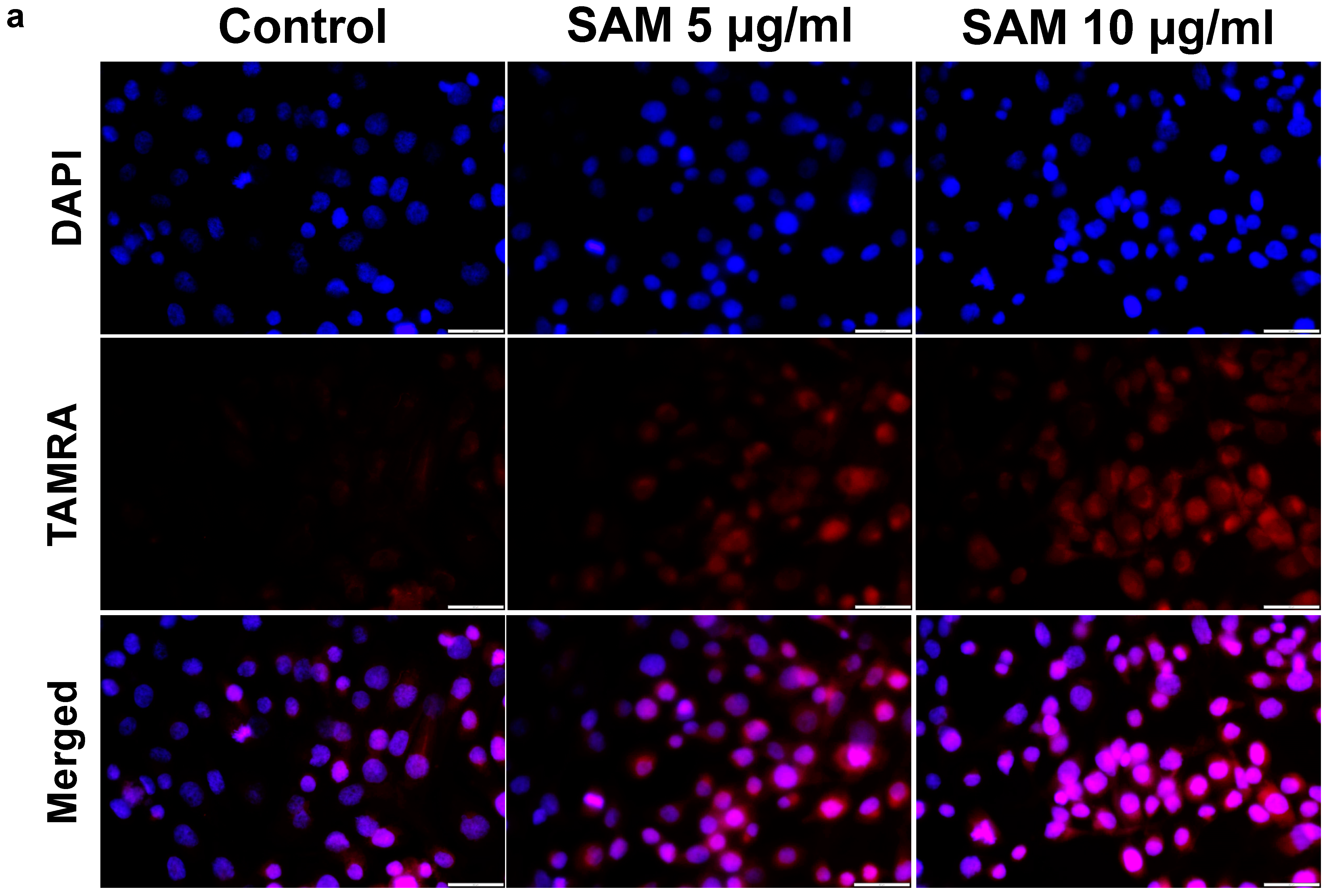
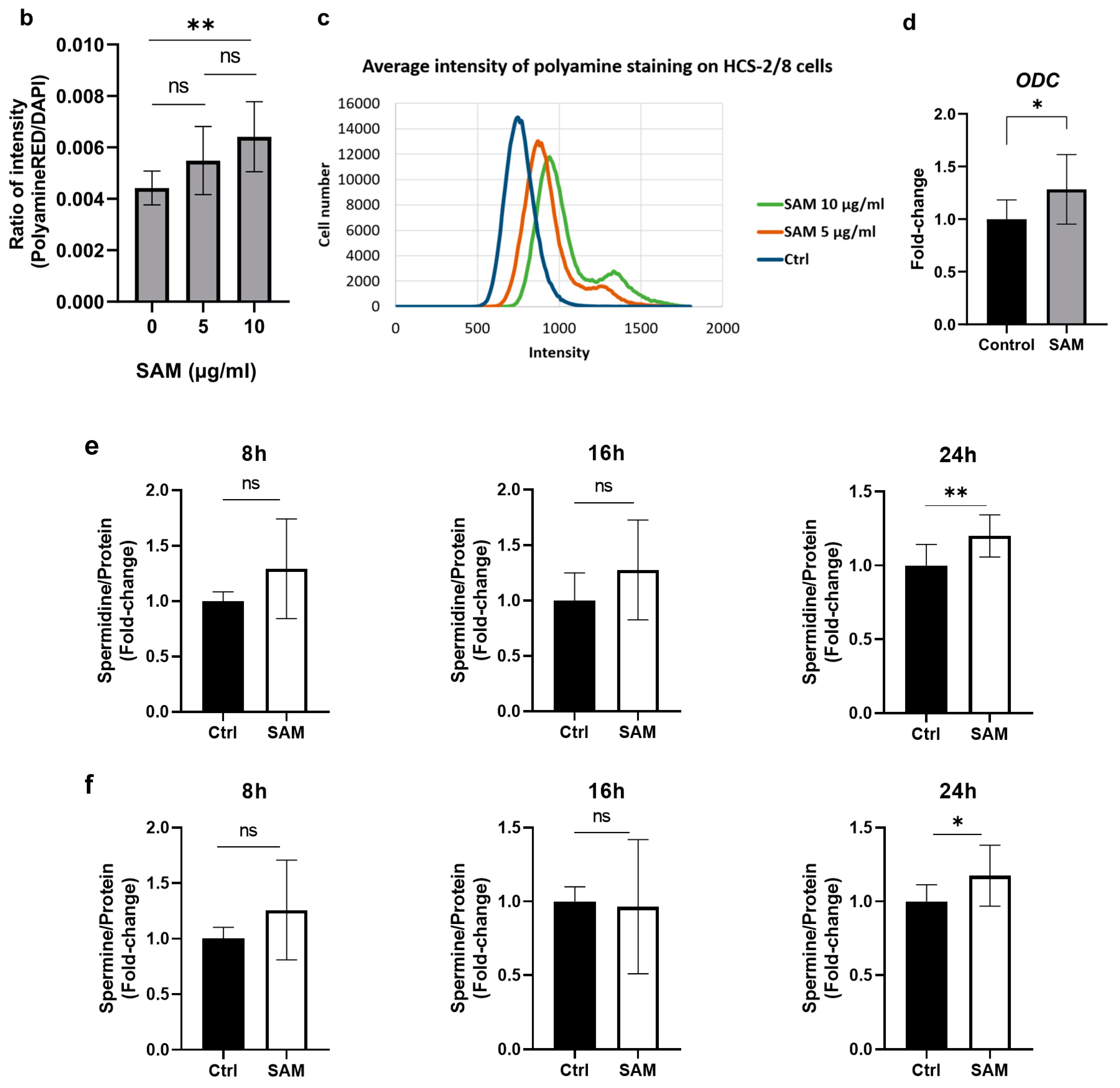
2.6. Exogenous SAM Increases Polyamine Levels in Chondrocytes
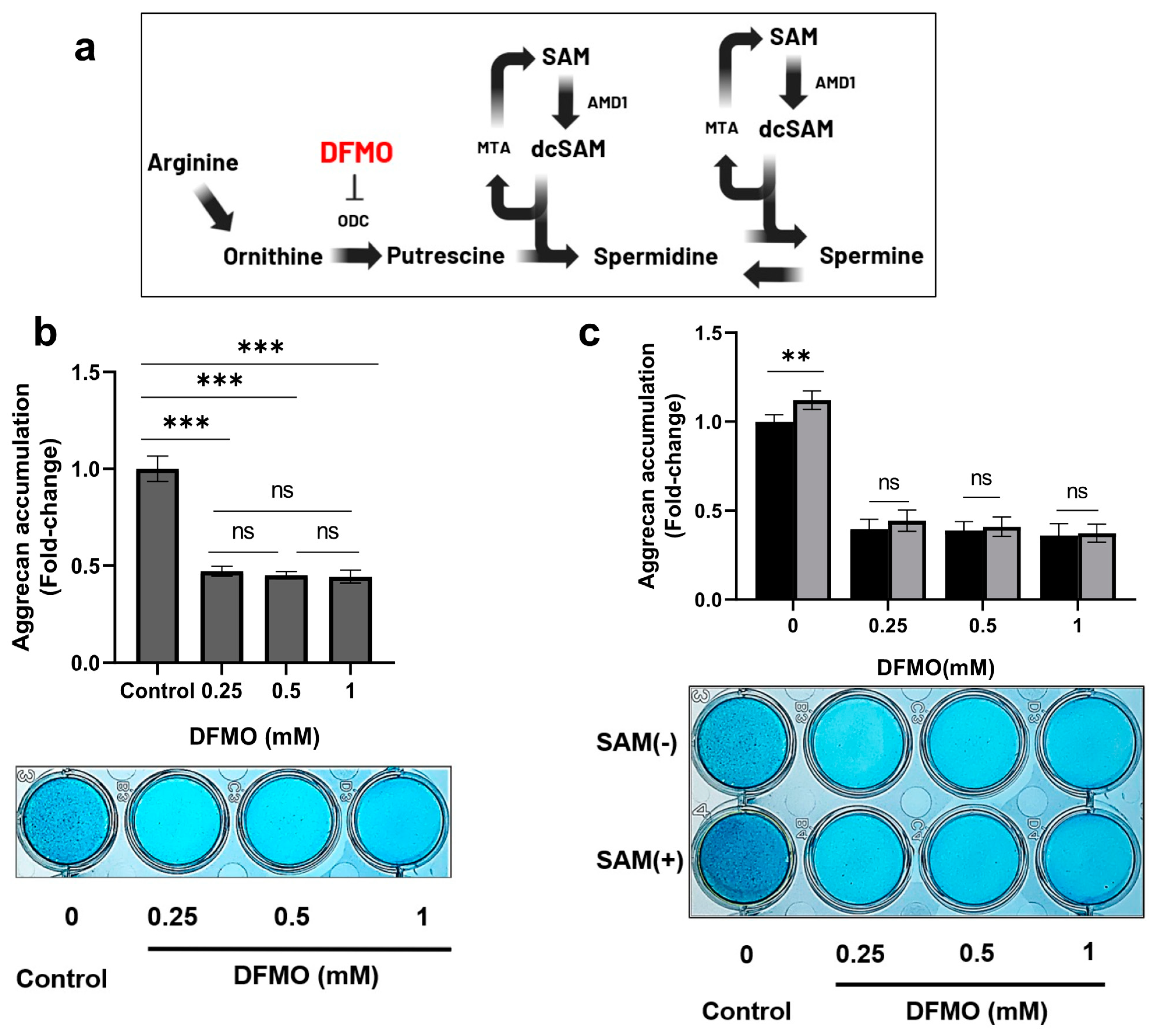
2.7. DFMO, an Inhibitor of ODC, Decreases SAM-Induced Aggrecan Production in Chondrocytes
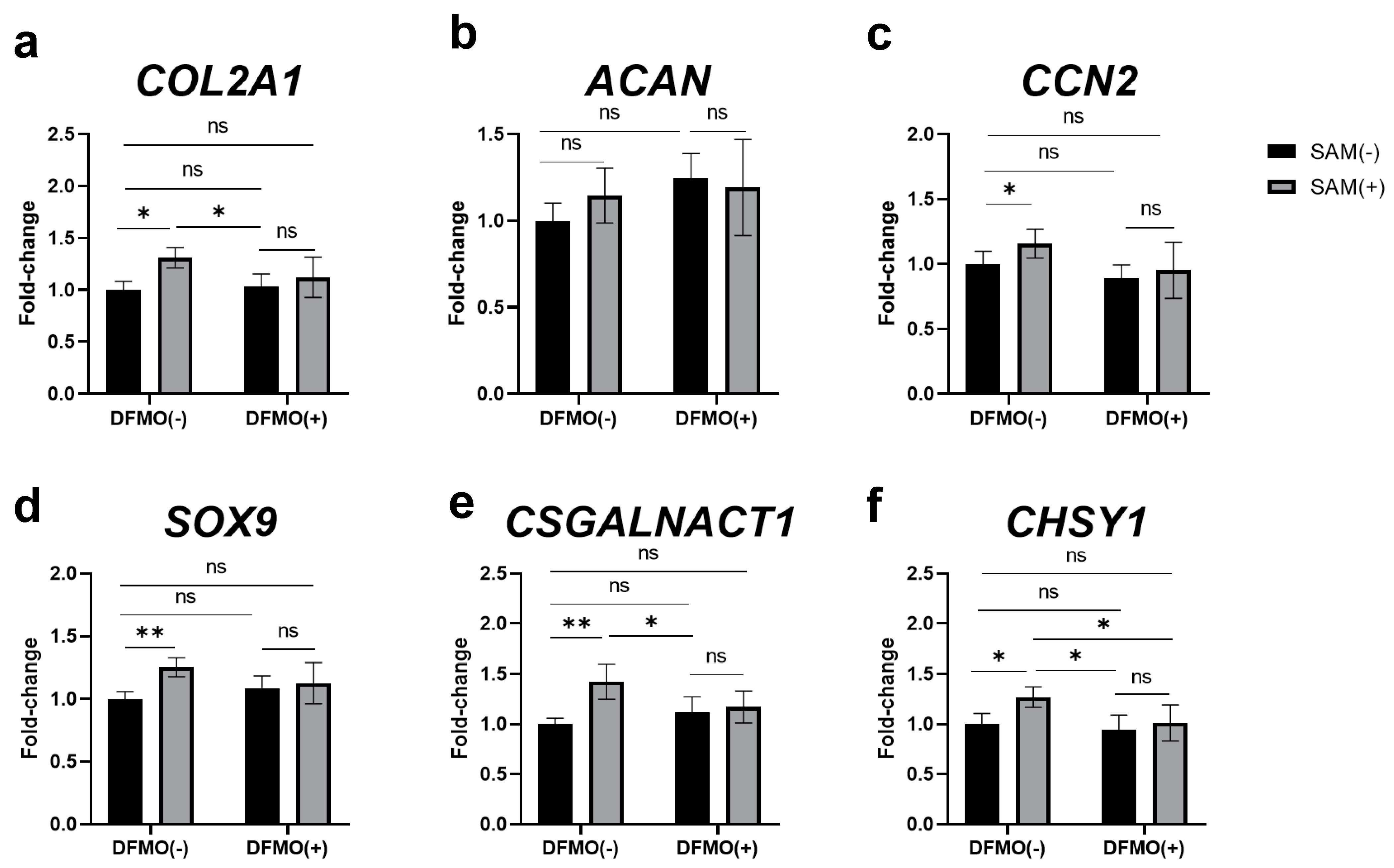
2.8. Inhibition of Polyamine Synthesis by DFMO Eliminates SAM-Induced Enhancement of Gene Expression in HCS-2/8 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Proliferation Assay
4.4. Alcian Blue Staining
4.5. Reverse-Transcription—Quantitative PCR
4.6. Western Blotting
4.7. Knockdown of mRNA Using MAT2A siRNA
4.8. Detection of Polyamine Level by PolyamineRED Staining
4.9. Determination of SAM Level by ELISA
4.10. Determination of Polyamine Level by High Performance Liquid Chromatography
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mato, J.M.; Martínez-Chantar, M.L.; Lu, S.C. S-Adenosylmethionine Metabolism and Liver Disease. Ann. Hepatol. 2013, 12, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Toennies, G. Sulfonium reactions of methionine and their possible metabolic significance. J. Biol. Chem. 1940, 132, 455–456. [Google Scholar] [CrossRef]
- Cantoni, G.L. S-Adenosylmethionine: A new intermediate formed enzymatically from l-methionine and adenosinetriphosphate. J. Biol. Chem. 1953, 204, 403–416. [Google Scholar] [CrossRef]
- Cuomo, A.; Beccarini Crescenzi, B.; Bolognesi, S.; Goracci, A.; Koukouna, D.; Rossi, R.; Fagiolini, A. S-Adenosylmethionine (SAMe) in Major Depressive Disorder (MDD): A Clinician-Oriented Systematic Review. Ann. Gen. Psychiatry 2020, 19, 50. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, R.; Deng, Q.; Wang, W.; Cao, C.; Yu, C.; Li, S.; Shi, L.; Tian, J. S-Adenosylmethionine Improves Cognitive Impairment in D-Galactose-Induced Brain Aging by Inhibiting Oxidative Stress and Neuroinflammation. J. Chem. Neuroanat. 2023, 128, 102232. [Google Scholar] [CrossRef] [PubMed]
- Papakostas, G.I. Evidence for S-Adenosyl-L-Methionine (SAM-e) for the Treatment of Major Depressive Disorder. J. Clin. Psychiatry 2009, 70 (Suppl. 5), 18–22. [Google Scholar] [CrossRef]
- Soeken, K.L.; Lee, W.-L.; Bausell, R.B.; Agelli, M.; Berman, B.M. Safety and Efficacy of S-Adenosylmethionine (SAMe) for Osteoarthritis. J. Fam. Pract. 2002, 51, 425–430. [Google Scholar]
- Medici, V.; Virata, M.C.; Peerson, J.M.; Stabler, S.P.; French, S.W.; Gregory, J.F.; Albanese, A.; Bowlus, C.L.; Devaraj, S.; Panacek, E.A.; et al. S-adenosylmethionine treatment of alcoholic liver disease: A double-blind, randomized, placebo-controlled trial. Alcohol. Clin. Exp. Res. 2011, 35, 1960–1965. [Google Scholar] [CrossRef]
- Mosca, L.; Vitiello, F.; Pagano, M.; Coppola, A.; Veglia Tranchese, R.; Grillo, R.; Cacciapuoti, G.; Porcelli, M. S-Adenosylmethionine, a Promising Antitumor Agent in Oral and Laryngeal Cancer. Appl. Sci. 2022, 12, 1746. [Google Scholar] [CrossRef]
- Glorioso, S.; Todesco, S.; Mazzi, A.; Marcolongo, R.; Giordano, M.; Colombo, B.; Cherie-Ligniere, G.; Mattara, L.; Leardini, G.; Passeri, M. Double-Blind Multicentre Study of the Activity of S-Adenosylmethionine in Hip and Knee Osteoarthritis. Int. J. Clin. Pharmacol. Res. 1985, 5, 39–49. [Google Scholar]
- Müller-Fassbender, H. Double-Blind Clinical Trial of S-Adenosylmethionine versus Ibuprofen in the Treatment of Osteoarthritis. Am. J. Med. 1987, 83, 81–83. [Google Scholar] [CrossRef]
- Najm, W.I.; Reinsch, S.; Hoehler, F.; Tobis, J.S.; Harvey, P.W. S-Adenosyl Methionine (SAMe) versus Celecoxib for the Treatment of Osteoarthritis Symptoms: A Double-Blind Cross-over Trial. [ISRCTN36233495]. BMC Musculoskelet. Disord. 2004, 5, 6. [Google Scholar] [CrossRef]
- Hosea Blewett, H.J. Exploring the Mechanisms behind S-Adenosylmethionine (SAMe) in the Treatment of Osteoarthritis. Crit. Rev. Food Sci. Nutr. 2008, 48, 458–463. [Google Scholar] [CrossRef]
- Harmand, M.-F.; Vilamitjana, J.; Maloche, E.; Duphil, R.; Ducassou, D. Effects of S-Adenosylmethionine on Human Articular Chondrocyte Differentiation: An in Vitro Study. Am. J. Med. 1987, 83, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Di Padova, C. S-Adenosylmethionine in the Treatment of Osteoarthritis: Review of the Clinical Studies. Am. J. Med. 1987, 83, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, L.; Zhan, Y.; Xie, C.; Xiang, Y.; Chen, D.; Wu, Y. Clinical Value and Expression of Homer 1, Homocysteine, S-Adenosyl-l-Homocysteine, Fibroblast Growth Factors 23 in Coronary Heart Disease. BMC Cardiovasc. Disord. 2022, 22, 215. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef]
- Canellakis, Z.N.; Marsh, L.L.; Bondy, P.K. Polyamines and Their Derivatives as Modulators in Growth and Differentiation. Yale, J. Biol. Med. 1989, 62, 481. [Google Scholar]
- He, Y.; Kashiwagi, K.; Fukuchi, J.; Terao, K.; Shirahata, A.; Igarashi, K. Correlation between the Inhibition of Cell Growth by Accumulated Polyamines and the Decrease of Magnesium and ATP. Eur. J. Biochem. 1993, 217, 89–96. [Google Scholar] [CrossRef]
- Sims, E.K.; Kulkarni, A.; Hull, A.; Woerner, S.E.; Cabrera, S.; Mastrandrea, L.D.; Hammoud, B.; Sarkar, S.; Nakayasu, E.S.; Mastracci, T.L.; et al. Inhibition of Polyamine Biosynthesis Preserves β Cell Function in Type 1 Diabetes. Cell Rep. Med. 2023, 4, 101261. [Google Scholar] [CrossRef]
- Brooks, W. Increased Polyamines Alter Chromatin and Stabilize Autoantigens in Autoimmune Diseases. Front. Immunol. 2013, 4, 91. [Google Scholar] [CrossRef] [PubMed]
- Facchini, A.; Borzì, R.M.; Olivotto, E.; Platano, D.; Pagani, S.; Cetrullo, S.; Flamigni, F. Role of Polyamines in Hypertrophy and Terminal Differentiation of Osteoarthritic Chondrocytes. Amino Acids 2012, 42, 667–678. [Google Scholar] [CrossRef]
- Takigawa, M.; Takano, T.; Suzuki, F. Effects of Parathyroid Hormone and Cyclic AMP Analogues on the Activity of Ornithine Decarboxylase and Expression of the Differentiated Phenotype of Chondrocytes in Culture. J. Cell. Physiol. 1981, 106, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Takigawa, M.; Ishida, H.; Takano, T.; Suzuki, F. Polyamine and Differentiation: Induction of Ornithine Decarboxylase by Parathyroid Hormone Is a Good Marker of Differentiated Chondrocytes. Proc. Natl. Acad. Sci. USA 1980, 77, 1481–1485. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Takigawa, M.; Suzuki, F. Role of Polyamines in Expression of the Differentiated Phenotype of Chondrocytes in Culture. Med. Biol. 1981, 59, 423–427. [Google Scholar]
- Takigawa, M. CCN Proteins (Cellular Communication Network Factors): Expanding Their Repertoire Toward a New Concept. Methods Mol. Biol. 2023, 2582, 1–10. [Google Scholar] [CrossRef]
- Charrier, A.; Brigstock, D.R. Regulation of Pancreatic Function by Connective Tissue Growth Factor (CTGF, CCN2). Cytokine Growth Factor Rev. 2013, 24, 59–68. [Google Scholar] [CrossRef]
- Arnott, J.A.; Lambi, A.G.; Mundy, C.M.; Hendesi, H.; Pixley, R.A.; Owen, T.A.; Safadi, F.F.; Popoff, S.N. The Role of Connective Tissue Growth Factor (CTGF/CCN2) in Skeletogenesis. Crit. Rev. Eukaryot. Gene Expr. 2011, 21, 43–69. [Google Scholar] [CrossRef]
- Takigawa, M. CCN2: A Master Regulator of the Genesis of Bone and Cartilage. J. Cell Commun. Signal. 2013, 7, 191–201. [Google Scholar] [CrossRef]
- Kubota, S.; Takigawa, M. Role of CCN2/CTGF/Hcs24 in Bone Growth. In International Review of Cytology; Academic Press: Cambridge, MA, USA, 2007; Volume 257. [Google Scholar] [CrossRef]
- Szustak, M.; Gendaszewska-Darmach, E. Nanocellulose-Based Scaffolds for Chondrogenic Differentiation and Expansion. Front. Bioeng. Biotechnol. 2021, 9, 736213. [Google Scholar] [CrossRef]
- LoGiudice, N.; Le, L.; Abuan, I.; Leizorek, Y.; Roberts, S.C. Alpha-Difluoromethylornithine, an Irreversible Inhibitor of Polyamine Biosynthesis, as a Therapeutic Strategy against Hyperproliferative and Infectious Diseases. Med. Sci. Basel Switz. 2018, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Rigueur, D.; Lyons, K.M. Whole-Mount Skeletal Staining. Methods Mol. Biol. 2014, 1130, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bi, T.; Liu, L.; Gao, Q.; Shen, G.; Qin, L. S-Adenosylmethionine Synergistically Enhances the Antitumor Effect of Gemcitabine against Pancreatic Cancer through JAK2/STAT3 Pathway. Naunyn. Schmiedebergs Arch. Pharmacol. 2019, 392, 615–622. [Google Scholar] [CrossRef]
- Yan, L.; Liang, X.; Huang, H.; Zhang, G.; Liu, T.; Zhang, J.; Chen, Z.; Zhang, Z.; Chen, Y. S-Adenosylmethionine Affects Cell Cycle Pathways and Suppresses Proliferation in Liver Cells. J. Cancer 2019, 10, 4368–4379. [Google Scholar] [CrossRef]
- Módis, K.; Coletta, C.; Asimakopoulou, A.; Szczesny, B.; Chao, C.; Papapetropoulos, A.; Hellmich, M.R.; Szabo, C. Effect of S-Adenosyl-L-Methionine (SAM), an Allosteric Activator of Cystathionine-β-Synthase (CBS) on Colorectal Cancer Cell Proliferation and Bioenergetics in Vitro. Nitric Oxide Biol. Chem. 2014, 41, 146–156. [Google Scholar] [CrossRef]
- Cooper, G.M. The Cell: A Molecular Approach, 2nd ed.; Oxford University Press: Sunderland MA, USA, 2000. [Google Scholar]
- Sharma, A.; Wood, L.D.; Richardson, J.B.; Roberts, S.; Kuiper, N.J. Glycosaminoglycan Profiles of Repair Tissue Formed Following Autologous Chondrocyte Implantation Differ from Control Cartilage. Arthritis Res. Ther. 2007, 9, R79. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Kimata, K.; Sato, T.; Gotoh, M.; Narimatsu, H.; Shinomiya, K.; Watanabe, H. Chondroitin Sulfate N-Acetylgalactosaminyltransferase-1 Plays a Critical Role in Chondroitin Sulfate Synthesis in Cartilage. J. Biol. Chem. 2007, 282, 4152–4161. [Google Scholar] [CrossRef]
- Sato, T.; Gotoh, M.; Kiyohara, K.; Akashima, T.; Iwasaki, H.; Kameyama, A.; Mochizuki, H.; Yada, T.; Inaba, N.; Togayachi, A.; et al. Differential Roles of TwoN-Acetylgalactosaminyltransferases, CSGalNAcT-1, and a Novel Enzyme, CSGalNAcT-2. J. Biol. Chem. 2003, 278, 3063–3071. [Google Scholar] [CrossRef]
- Clarke, S.G. 16 Inhibition of Mammalian Protein Methyltransferases by 5’-Methylthioadenosine (MTA): A Mechanism of Action of Dietary Same? The Enzymes 2006, 24, 467–493. [Google Scholar] [CrossRef]
- Barceló, H.A.; Wiemeyer, J.C.M.; Sagasta, C.L.; Macias, M.; Barreira, J.C. Effect of S-Adenosylmethionine on Experimental Osteoarthritis in Rabbits. Am. J. Med. 1987, 83 (Suppl. 1), 55–59. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, H.; Liu, D.; Liu, W.; Du, J.; Wen, D.; Li, L.; Zhang, A.; Jiang, J.; Zeng, L. CTGF Promotes the Repair and Regeneration of Alveoli after Acute Lung Injury by Promoting the Proliferation of Subpopulation of AEC2s. Respir. Res. 2023, 24, 227. [Google Scholar] [CrossRef] [PubMed]
- Leask, A.; Parapuram, S.K.; Shi-wen, X.; Abraham, D.J. Connective Tissue Growth Factor (CTGF, CCN2) Gene Regulation: A Potent Clinical Bio-Marker of Fibroproliferative Disease? J. Cell Commun. Signal. 2009, 3, 89–94. [Google Scholar] [CrossRef]
- Guo, F.; Carter, D.E.; Leask, A. Mechanical Tension Increases CCN2/CTGF Expression and Proliferation in Gingival Fibroblasts via a TGFβ-Dependent Mechanism. PLoS ONE 2011, 6, e19756. [Google Scholar] [CrossRef]
- Abd El Kader, T.; Kubota, S.; Nishida, T.; Hattori, T.; Aoyama, E.; Janune, D.; Hara, E.S.; Ono, M.; Tabata, Y.; Kuboki, T.; et al. The Regenerative Effects of CCN2 Independent Modules on Chondrocytes in Vitro and Osteoarthritis Models in Vivo. Bone 2014, 59, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Mihara, A.; Ohara, T.; Hata, J.; Chen, S.; Honda, T.; Tamrakar, S.; Isa, A.; Wang, D.; Shimizu, K.; Katakura, Y.; et al. Association of Serum S-Adenosylmethionine, s-Adenosylhomocysteine, and Their Ratio with the Risk of Dementia and Death in a Community. Sci. Rep. 2022, 12, 12427. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, K.; Lefebvre, V.; Zhou, G.; Garofalo, S.; Kimura, J.H.; de Crombrugghe, B. Use of a New Rat Chondrosarcoma Cell Line to Delineate a 119-Base Pair Chondrocyte-Specific Enhancer Element and to Define Active Promoter Segments in the Mouse Pro-A1(II) Collagen Gene. J. Biol. Chem. 1995, 270, 27711–27719. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Nishiyama, T.; Miura, Y.; Fujishiro, T.; Kanzaki, N.; Hashimoto, S.; Matsumoto, T.; Kurosaka, M.; Kuroda, R. DcR3 Induces Cell Proliferation through MAPK Signaling in Chondrocytes of Osteoarthritis. Osteoarthr. Cartil. 2011, 19, 903–910. [Google Scholar] [CrossRef]
- Farquharson, C.; Whitehead, C.C.; Loveridge, N. Alterations in Glycosaminoglycan Concentration and Sulfation during Chondrocyte Maturation. Calcif. Tissue Int. 1994, 54, 296–303. [Google Scholar] [CrossRef]
- Yamamoto, K.; Kishida, T.; Nakai, K.; Sato, Y.; Kotani, S.; Nishizawa, Y.; Yamamoto, T.; Kanamura, N.; Mazda, O. Direct Phenotypic Conversion of Human Fibroblasts into Functional Osteoblasts Triggered by a Blockade of the Transforming Growth Factor-β Signal. Sci. Rep. 2018, 8, 8463. [Google Scholar] [CrossRef]
- Ono, K.; Okusha, Y.; Tran, M.T.; Umemori, K.; Eguchi, T. Western Blot Protocols for Analysis of CCN Proteins and Fragments in Exosomes, Vesicle-Free Fractions, and Cells. Methods Mol. Biol. 2023, 2582, 39–57. [Google Scholar] [CrossRef]
- Takamatsu, A.; Ohkawara, B.; Ito, M.; Masuda, A.; Sakai, T.; Ishiguro, N.; Ohno, K. Verapamil Protects against Cartilage Degradation in Osteoarthritis by Inhibiting Wnt/β-Catenin Signaling. PLoS ONE 2014, 9, e92699. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M.; Márquez, J.; García-Caballero, M.; Núñez de Castro, I.; Sánchez-Jiménez, F. Simultaneous Fluorometric Determination of Intracellular Polyamines Separated by Reversed-Phase High-Performance Liquid Chromatography. Agents Actions 1992, 36, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Harada, Y.; Moriyama, S.; Furuta, K.; Tanaka, S.; Miyaji, T.; Omote, H.; Moriyama, Y.; Hiasa, M. Vesicular Polyamine Transporter Mediates Vesicular Storage and Release of Polyamine from Mast Cells. J. Biol. Chem. 2017, 292, 3909–3918. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences | Species |
|---|---|---|
| CCN2 | F-GCAGGCTAGAGAAGCAGAGC R-ATGTCTTCATGCTGGTGCAG | Human |
| ACAN | F-TTCGGGCAGAAGAAGGAC R-CGTGAGCTCCGCTTCTGT | Human |
| COL2a1 | F-ATGGCTTCCAGAGGCC GAC R-TTGCTGCATGAGTTGCCACGCA | Human |
| ODC | F-ATGGCTTCCAGAGGCC GAC R-TTGCTGCATGAGTTGCCACGCA | Human |
| SOX9 | F-AGGCAAGCAAAGGAGATGAA R-TGGTGTTCTGAGAGGCACAG | Human |
| GAPDH | F-GCCAAAAGGGTCATCATCTC R-GTCTTCTGGGTGGCAGTGAT | Human |
| CHSY1 | F-CGACAGGAACTTTCTCTTCGTGG R-GGTACAGATGTGTCAGAACCCTC | Human |
| CHSY3 | F-AGTTGGAGCGGGCTTACAGTGA R-CAGCACCTCAAAGCGAGAGTGT | Human |
| CSGALNACT1 | F-GGATGACGTGTCAGTATCGGTC R-CCGTACCACTATGAGGTTGCTG | Human |
| CSGALNACT2 | F-TCATCTCACAGTGGTGTATTTTGG R-GCACCCACATTTAGTCCTCGTC | Human |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoang, L.D.; Aoyama, E.; Hiasa, M.; Omote, H.; Kubota, S.; Kuboki, T.; Takigawa, M. Positive Regulation of S-Adenosylmethionine on Chondrocytic Differentiation via Stimulation of Polyamine Production and the Gene Expression of Chondrogenic Differentiation Factors. Int. J. Mol. Sci. 2023, 24, 17294. https://doi.org/10.3390/ijms242417294
Hoang LD, Aoyama E, Hiasa M, Omote H, Kubota S, Kuboki T, Takigawa M. Positive Regulation of S-Adenosylmethionine on Chondrocytic Differentiation via Stimulation of Polyamine Production and the Gene Expression of Chondrogenic Differentiation Factors. International Journal of Molecular Sciences. 2023; 24(24):17294. https://doi.org/10.3390/ijms242417294
Chicago/Turabian StyleHoang, Loc Dinh, Eriko Aoyama, Miki Hiasa, Hiroshi Omote, Satoshi Kubota, Takuo Kuboki, and Masaharu Takigawa. 2023. "Positive Regulation of S-Adenosylmethionine on Chondrocytic Differentiation via Stimulation of Polyamine Production and the Gene Expression of Chondrogenic Differentiation Factors" International Journal of Molecular Sciences 24, no. 24: 17294. https://doi.org/10.3390/ijms242417294