
Ceruloplasmin Reduces the Lactoferrin/Oleic Acid Antitumor Complex-Mediated Release of Heme-Containing Proteins from Blood Cells

Abstract
:1. Introduction
2. Results and Discussion
2.1. Interaction of Ceruloplasmin with Lactoferrin and with the Lactoferrin/Oleic Acid Complex
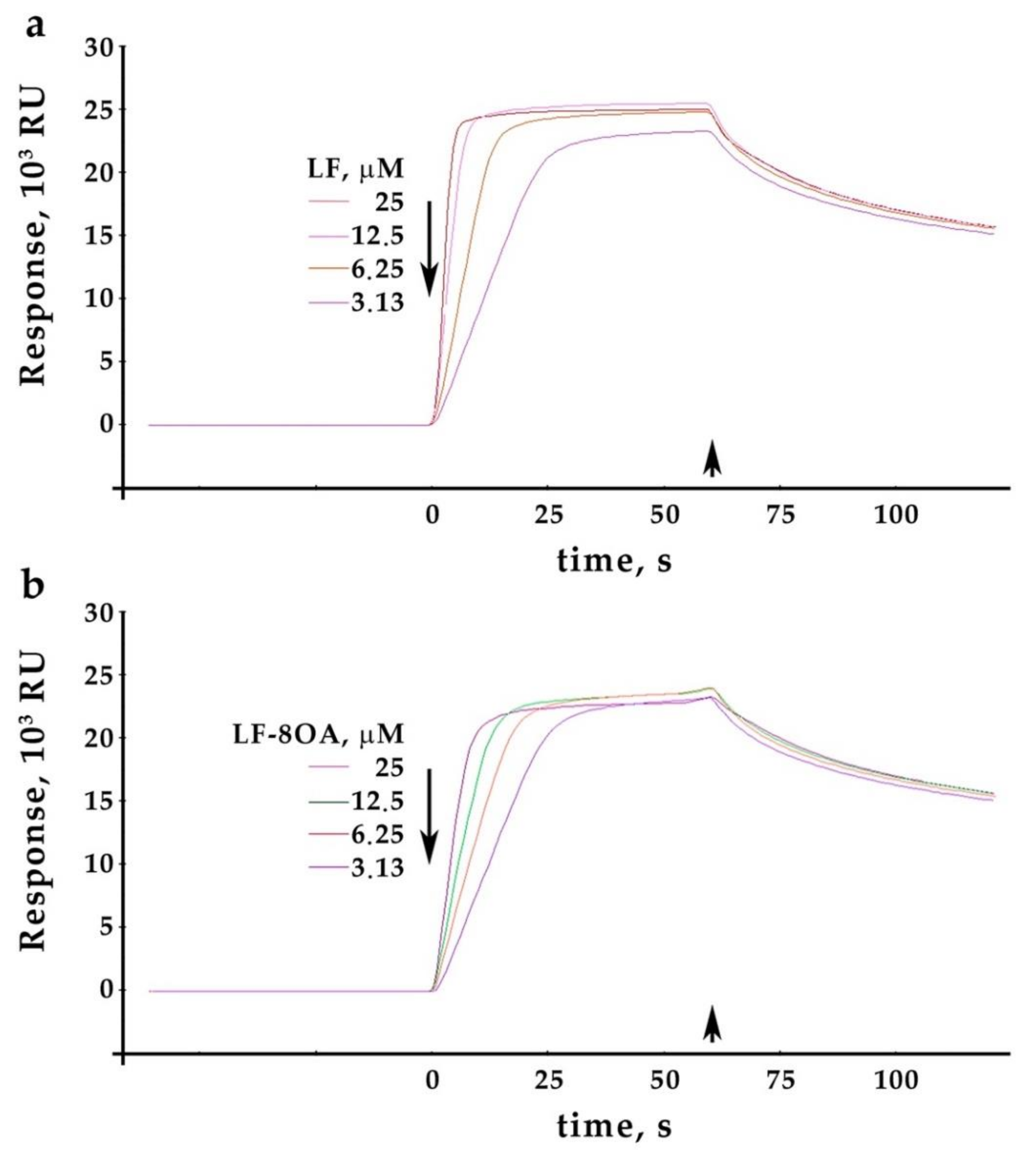
2.2. Ceruloplasmin Interacts with Lactoferrin and with the Lactoferrin/Oleic Acid Complex: Kinetics of Interaction
2.3. Comparing the Effect of Complexes of Lactoferrin or Albumin with Oleic Acid on the Lysis of Erythrocytes
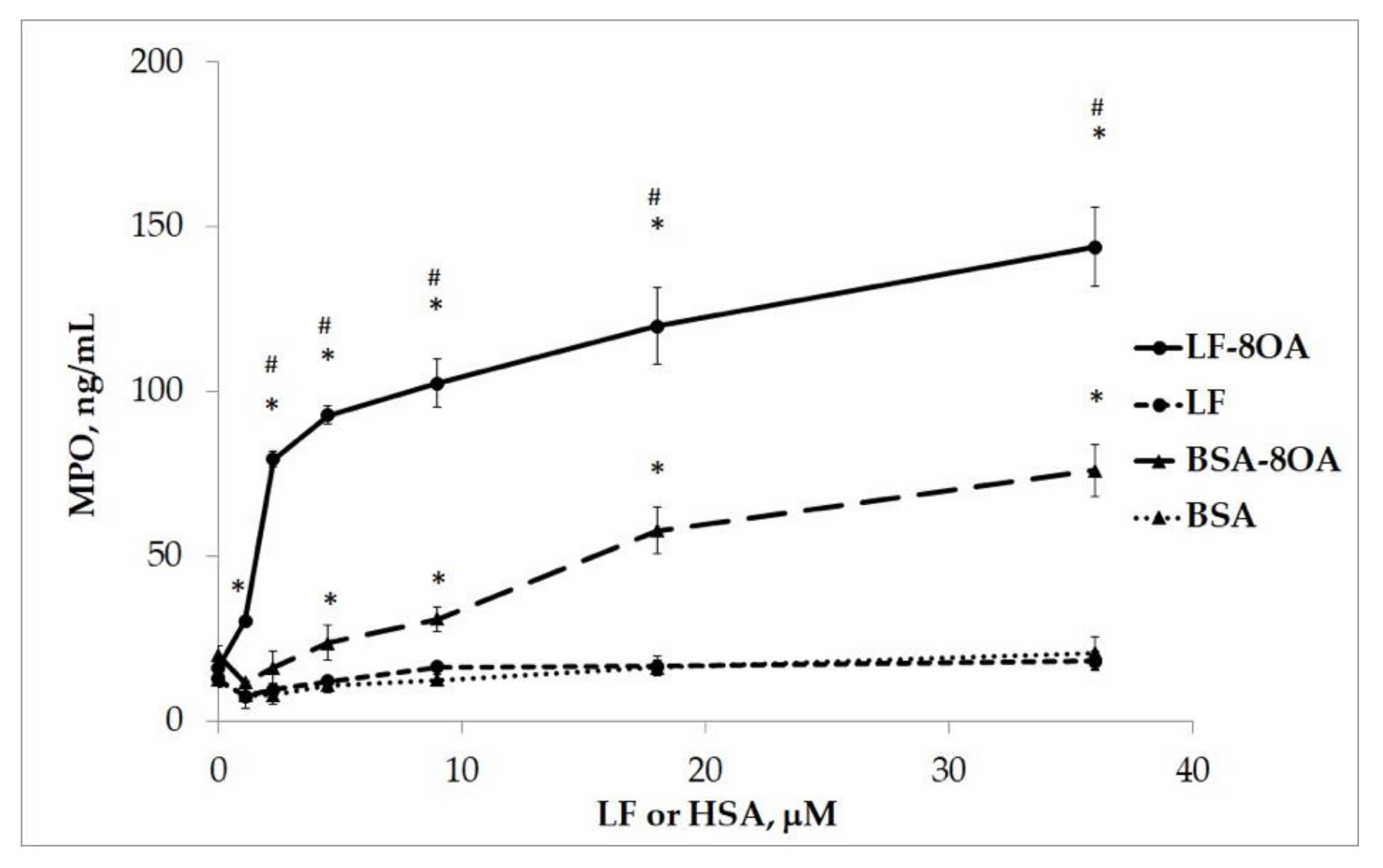
2.4. The Activation of Neutrophils in the Presence of the LF/OA Complex
3. Materials and Methods
3.1. Isolation and Purification of Human and Murine Ceruloplasmin
3.2. Isolation and Purification of Myeloperoxidase
3.3. Spectrophotometry for Measuring Concentration of Proteins
3.4. Obtaining Complexes of Proteins with Oleic Acid
3.5. Analysis of Stoichiometry in Complexes Formed by LF or BSA with OA
3.6. Isolation of Erythrocytes
3.7. Analysis of Hemolytic Activity
3.8. Isolation of Neutrophils
3.9. Myeloperoxidase Exocytosis
3.10. Studying Protein–Protein Interaction Using Biosensors Based on Surface Plasmon Resonance
3.11. Electrophoretic Methods
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Håkansson, A.; Zhivotovsky, B.; Orrenius, S.; Sabharwal, H.; Svanborg, C. Apoptosis induced by a human milk protein. Proc. Natl. Acad. Sci. USA 1995, 92, 8064–8068. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Mak, A.; Koch, K.; Moore, R.B. A molecular complex of bovine milk protein and oleic acid selectively kills cancer cells in vitro and inhibits tumour growth in an orthotopic rat bladder tumour model. BJU Int. 2013, 112, E201–E210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, F., Jr.; Yang, F.; Chen, J.; Zheng, C.Y.; Liang, Y. Cytotoxic aggregates of alpha-lactalbumin induced by unsaturated fatty acid induce apoptosis in tumor cells. Chem. Biol. Interact. 2009, 180, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Puthia, M.; Storm, P.; Nadeem, A.; Hsiung, S.; Svanborg, C. Prevention and treatment of colon cancer by peroral administration of HAMLET (human α-lactalbumin made lethal to tumour cells). Gut 2014, 63, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.; Gustafsson, L.; Mossberg, A.K.; Gronli, J.; Mork, S.; Bjerkvig, R.; Svanborg, C. Human alpha-lactalbumin made lethal to tumor cells (HAMLET) kills human glioblastoma cells in brain xenografts by an apoptosis-like mechanism and prolongs survival. Cancer Res. 2004, 64, 2105–2112. [Google Scholar] [CrossRef] [PubMed]
- Mossberg, A.K.; Hou, Y.; Svensson, M.; Holmqvist, B.; Svanborg, C. HAMLET treatment delays bladder cancer development. J. Urol. 2010, 183, 1590–1597. [Google Scholar] [CrossRef]
- Mossberg, A.K.; Wullt, B.; Gustafsson, L.; Månsson, W.; Ljunggren, E.; Svanborg, C. Bladder cancers respond to intravesical instillation of HAMLET (human alpha-lactalbumin made lethal to tumor cells). Int. J. Cancer 2007, 121, 1352–1359. [Google Scholar] [CrossRef]
- Gustafsson, L.; Leijonhufvud, I.; Aronsson, A.; Mossberg, A.K.; Svanborg, C. Treatment of skin papillomas with topical alpha-lactalbumin-oleic acid. N. Engl. J. Med. 2004, 350, 2663–2672. [Google Scholar] [CrossRef]
- Brisuda, A.; Ho, J.C.S.; Kandiyal, P.S.; Ng, J.T.; Ambite, I.; Butler, D.S.C.; Háček, J.; Wan, M.L.Y.; Tran, T.H.; Nadeem, A.; et al. Bladder cancer therapy using a conformationally fluid tumoricidal peptide complex. Nat. Commun. 2021, 12, 3427–3443. [Google Scholar] [CrossRef]
- Brinkmann, C.R.; Heegaard, C.W.; Petersen, T.E.; Jensenius, J.C.; Thiel, S. The toxicity of bovine α-lactalbumin made lethal to tumor cells is highly dependent on oleic acid and induces killing in cancer cell lines and noncancer-derived primary cells. FEBS J. 2011, 278, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Zhang, M.; Tian, M.; Ren, F.Z. Self-assembled β-lactoglobulin-oleic acid and β-lactoglobulin-linoleic acid complexes with antitumor activities. J. Dairy Sci. 2015, 98, 2898–2907. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, K.; Darinskas, A.; Noppe, W.; Duchardt, E.; Mok, K.H.; Vukojević, V.; Schleucher, J.; Morozova-Roche, L.A. Protein oligomerization induced by oleic acid at the solid-liquid interface--equine lysozyme cytotoxic complexes. FEBS J. 2009, 276, 3975–3989. [Google Scholar] [CrossRef] [PubMed]
- Atri, M.S.; Saboury, A.A.; Moosavi-Movahedi, A.A.; Goliaei, B.; Sefidbakht, Y.; Alijanvand, H.H.; Sharifzadeh, A.; Niasari-Naslaji, A. Structure and stability analysis of cytotoxic complex of camel α-lactalbumin and unsaturated fatty acids produced at high temperature. J. Biomol. Struct. Dyn. 2011, 28, 919–928. [Google Scholar] [CrossRef] [PubMed]
- El-Fakharany, E.M.; Abu-Serie, M.M.; Litus, E.A.; Permyakov, S.E.; Permyakov, E.A.; Uversky, V.N.; Redwan, E.M. The Use of Human, Bovine, and Camel Milk Albumins in Anticancer Complexes with Oleic Acid. Protein J. 2018, 37, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Zhang, M.; Tian, M.; Jiang, L.; Guo, H.Y.; Ren, F.Z. Bovine lactoferrin binds oleic acid to form an anti-tumor complex similar to HAMLET. Biochim. Biophys. Acta 2014, 1841, 535–543. [Google Scholar] [CrossRef]
- Yao, Q.; Li, H.; Fan, L.; Huang, S.; Wang, J.; Zheng, N. The combination of lactoferrin and linolenic acid inhibits colorectal tumor growth through activating AMPK/JNK-related apoptosis pathway. PeerJ 2021, 9, e11072. [Google Scholar] [CrossRef] [PubMed]
- Metz-Boutigue, M.H.; Jollès, J.; Mazurier, J.; Schoentgen, F.; Legrand, D.; Spik, G.; Montreuil, J.; Jollès, P. Human lactotransferrin: Amino acid sequence and structural comparisons with other transferrins. Eur. J. Biochem. 1984, 145, 659–676. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F.; Dive, C. An iron binding protein common to many external secretion. Clin. Chim. Acta 1996, 14, 735–739. [Google Scholar] [CrossRef]
- Masson, P.L.; Heremans, J.F. Lactoferrin in milk from different species. Comp. Biochem. Physiol. B 1971, 39, 119–129. [Google Scholar] [CrossRef]
- Cutone, A.; Rosa, L.; Ianiro, G.; Lepanto, M.S.; Bonaccorsi di Patti, M.C.; Valenti, P.; Musci, G. Lactoferrin’s Anti-Cancer Properties: Safety, Selectivity, and Wide Range of Action. Biomolecules 2020, 10, 456. [Google Scholar] [CrossRef] [PubMed]
- Fillebeen, C.; Descamps, L.; Dehouck, M.P.; Fenart, L.; Benaïssa, M.; Spik, G.; Cecchelli, R.; Pierce, A. Receptor-mediated transcytosis of lactoferrin through the blood-brain barrier. J. Biol. Chem. 1999, 274, 7011–7017. [Google Scholar] [CrossRef] [PubMed]
- Kamemori, N.; Takeuchi, T.; Sugiyama, A.; Miyabayashi, M.; Kitagawa, H.; Shimizu, H.; Ando, K.; Harada, E. Trans-endothelial and trans-epithelial transfer of lactoferrin into the brain through BBB and BCSFB in adult rats. J. Vet. Med. Sci. 2008, 70, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Zorina, V.N. The prospects for application of lactoferrin and its derivatives in the treatment of cancer. Probl. Oncol. 2019, 65, 785–790. [Google Scholar]
- Elizarova, A.; Sokolov, A.; Kostevich, V.; Kisseleva, E.; Zelenskiy, E.; Zakharova, E.; Panasenko, O.; Budevich, A.; Semak, I.; Egorov, V.; et al. Interaction of Lactoferrin with Unsaturated Fatty Acids: In Vitro and In Vivo Study of Human Lactoferrin/Oleic Acid Complex Cytotoxicity. Materials 2021, 14, 1602. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.V.; Pulina, M.O.; Zakharova, E.T.; Susorova, A.S.; Runova, O.L.; Vasilyev, V.B.; Kolodkin, N.I. Identification and isolation from breast milk of ceruloplasmin—Lactoferrin complex. Biochemistry 2006, 71, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.V.; Pulina, M.O.; Runova, O.L.; Zakharova, E.T.; Vasilyev, V.B. Complex of ceruloplasmin and lactoferrin in human lacrimal fluid. Med. Acad. J. 2013, 13, 39–43. [Google Scholar]
- Saenko, E.L.; Yaropolov, A.I. The protective effect of various human ceruloplasmin isoforms during copper-induced erythrocyte lysis. Biochemistry 1991, 56, 648–654. [Google Scholar]
- Varfolomeeva, E.Y.; Semenova, E.V.; Sokolov, A.V.; Aplin, K.D.; Timofeeva, K.E.; Vasilyev, V.B.; Filatov, M.V. Ceruloplasmin decreases respiratory burst reaction during pregnancy. Free. Radic. Res. 2016, 50, 909–919. [Google Scholar] [CrossRef]
- Bakkenist, A.R.; Wever, R.; Vulsma, T.; Plat, H.; van Gelder, B.F. Isolation procedure and some properties of myeloperoxidase from human leucocytes. Biochim. Biophys. Acta 1978, 524, 45–54. [Google Scholar] [CrossRef]
- Panasenko, O.M.; Gorudko, I.V.; Sokolov, A.V. Hypochlorous acid as a precursor of free radicals in living systems. Biochemistry 2013, 78, 1466–1489. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C.; Bryant, R.R.; Lim, S.; Scott, L.E.; Moor, J.E. Ceruloplasmin elevation and synthesis in rats with transplantable tumors. Enzyme 1979, 24, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Segelmark, M.; Persson, B.; Hellmark, T.; Wieslander, J. Binding and inhibition of myeloperoxidase (MPO): A major function of ceruloplasmin? Clin. Exp. Immunol. 1997, 108, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Chapman, A.L.; Mocatta, T.J.; Shiva, S.; Seidel, A.; Chen, B.; Khalilova, I.; Paumann-Page, M.E.; Jameson, G.N.L.; Winterbourn, C.C.; Kettle, A.J. Ceruloplasmin is an endogenous inhibitor of myeloperoxidase. J. Biol. Chem. 2013, 288, 6465–6477. [Google Scholar] [CrossRef] [PubMed]
- Maas, R.R.; Soukup, K.; Fournier, N.; Massara, M.; Galland, S.; Kornete, M.; Wischnewski, V.; Lourenco, J.; Croci, D.; Álvarez-Prado, Á.F.; et al. The local microenvironment drives activation of neutrophils in human brain tumors. Cell 2023, 186, 4546–4566. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Kantarci, A.; Yagi, M.; Jackson, T.; Hasturk, H.; Kurihara, H.; Van Dyke, T.E. Ceruloplasmin induces polymorphonuclear leukocyte priming in localized aggressive periodontitis. J. Periodontol. 2009, 80, 1300–1306. [Google Scholar] [CrossRef] [PubMed]
- Griffin, S.V.; Chapman, P.T.; Lianos, E.A.; Lockwood, C.M. The inhibition of myeloperoxidase by ceruloplasmin can be reversed by anti-myeloperoxidase antibodies. Kidney Int. 1999, 55, 917–925. [Google Scholar] [CrossRef]
- Reiding, K.R.; Franc, V.; Huitema, M.G.; Brouwer, E.; Heeringa, P.; Heck, A.J.R. Neutrophil myeloperoxidase harbors distinct site-specific peculiarities in its glycosylation. J. Biol. Chem. 2019, 294, 20233–20245. [Google Scholar] [CrossRef]
- Tjondro, H.C.; Ugonotti, J.; Kawahara, R.; Chatterjee, S.; Loke, I.; Chen, S.; Soltermann, F.; Hinneburg, H.; Parker, B.L.; Venkatakrishnan, V.; et al. Hyper-truncated Asn355- and Asn391-glycans modulate the activity of neutrophil granule myeloperoxidase. J. Biol. Chem. 2021, 296, 100144. [Google Scholar] [CrossRef]
- Van Berkel, P.H.; Geerts, M.E.; van Veen, H.A.; Mericskay, M.; de Boer, H.A.; Nuijens, J.H. N-terminal stretch Arg2, Arg3, Arg4 and Arg5 of human lactoferrin is essential for binding to heparin, bacterial lipopolysaccharide, human lysozyme and DNA. Biochem. J. 1997, 328 Pt 1, 145–151. [Google Scholar] [CrossRef]
- Britigan, B.E.; Lewis, T.S.; Waldschmidt, M.; McCormick, M.L.; Krieg, A.M. Lactoferrin binds CpG-containing oligonucleotides and inhibits their immunostimulatory effects on human B cells. J. Immunol. 2001, 167, 2921–2928. [Google Scholar] [CrossRef] [PubMed]
- Van Berkel, P.H.; van Veen, H.A.; Geerts, M.E.; Nuijens, J.H. Characterization of monoclonal antibodies against human lactoferrin. J. Immunol. Methods 2002, 267, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Y.; Shi, H.; Peng, Y.; Fan, X.; Li, C. The molecular mechanisms of copper metabolism and its roles in human diseases. Pflug. Arch. Eur. J. Physiol. 2020, 472, 1415–1429. [Google Scholar] [CrossRef] [PubMed]
- Geng, R.; Ke, N.; Wang, Z.; Mou, Y.; Xiang, B.; Zhang, Z.; Ji, X.; Zou, J.; Wang, D.; Yin, Z.; et al. Copper deprivation enhances the chemosensitivity of pancreatic cancer to rapamycin by mTORC1/2 inhibition. Chem.-Biol. Interact. 2023, 382, 110546. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.N.; Chen, L.H.; Yang, X.L.; Dong, J.Y.; Wu, W.B.; Chen, D.; Geng, R.M.; Ke, N.W.; Liu, J. Sichuan da xue xue bao. Yi Xue Ban J. Sichuan University. Med. Sci. Ed. 2020, 51, 643–649. [Google Scholar] [CrossRef]
- Li, Y.; Fang, M.; Xu, Z.; Li, X. Tetrathiomolybdate as an old drug in a new use: As a chemotherapeutic sensitizer for non-small cell lung cancer. J. Inorg. Biochem. 2022, 233, 111865. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wan, Y.; Chen, G.; Huang, Y. Multitargeting Strategy Using Tetrathiomolybdate and Lenvatinib: Maximizing Antiangiogenesis Activity in a Preclinical Liver Cancer Model. Anti-Cancer Agents Med. Chem. 2023, 23, 786–793. [Google Scholar] [CrossRef]
- Skvortsov, A.N.; Il’icheva, E.I.U.; Zatulovskiĭ, E.A.; Savel’ev, A.N.; Tsymbalenko, N.B.; Shavlovskiĭ, M.M.; Puchkova, L.V. Characteristics of rat ceruloplasmin from the serum of animals, which received salts of silver with food. Tsitologiia 2010, 52, 960–966. [Google Scholar]
- Zatulovskiy, E.A.; Skvortsov, A.N.; Rusconi, P.; Ilyechova, E.Y.; Babich, P.S.; Tsymbalenko, N.V.; Broggini, M.; Puchkova, L.V. Serum depletion of holo-ceruloplasmin induced by silver ions in vivo reduces uptake of cisplatin. J. Inorg. Biochem. 2012, 116, 88–96. [Google Scholar] [CrossRef]
- Semak, I.; Budzevich, A.; Maliushkova, E.; Kuzniatsova, V.; Popkov, N.; Zalutsky, I.; Ivashkevich, O. Development of dairy herd of transgenic goats as biofactory for large-scale production of biologically active recombinant human lactoferrin. Transgenic Res. 2019, 28, 465–478. [Google Scholar] [CrossRef]
- Gorbunov, N.P.; Kostevich, V.A.; Sokolov, A.V. Obtaining of monoclonal antibody for immunoaffinity purification of myeloperoxidase and prototype of hemosorbent. Russ. J. Immunol. 2019, 13, 215–217. [Google Scholar]
- Sokolov, A.V.; Kostevich, V.A.; Romanico, D.N.; Zakharova, E.T.; Vasilyev, V.B. Two-stage method for purification of ceruloplasmin based on its interaction with neomycin. Biochemistry 2012, 77, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, A.V.; Pulina, M.O.; Ageeva, K.V.; Ayrapetov, M.I.; Berlov, M.N.; Volgin, G.N.; Markov, A.G.; Yablonsky, P.K.; Kolodkin, N.I.; Zakharova, E.T.; et al. Interaction of ceruloplasmin, lactoferrin, and myeloperoxidase. Biochemistry 2007, 72, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Noyer, M.; Dwulet, F.E.; Hao, Y.L.; Putnam, F.W. Purification and characterization of undegraded human ceruloplasmin. Anal. Biochem. 1980, 102, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Gifford, J.L.; Ishida, H.; Vogel, H.J. Structural characterization of the interaction of human lactoferrin with calmodulin. PLoS ONE 2012, 7, e51026. [Google Scholar] [CrossRef] [PubMed]
- Phopin, K.; Ruankham, W.; Prachayasittikul, S.; Prachayasittikul, V.; Tantimongcolwat, T. Insight into the Molecular Interaction of Cloxyquin (5-chloro-8-hydroxyquinoline) with Bovine Serum Albumin: Biophysical Analysis and Computational Simulation. Int. J. Mol. Sci. 2019, 21, 249. [Google Scholar] [CrossRef] [PubMed]
- Sud’ina, G.F.; Brock, T.G.; Pushkareva, M.A.; Galkina, S.I.; Turutin, D.V.; Peters-Golden, M.; Ullrich, V. Sulphatides trigger polymorphonuclear granulocyte spreading on collagen-coated surfaces and inhibit subsequent activation of 5-lipoxygenase. Biochem. J. 2001, 359 Pt 3, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Churashova, I.A.; Sokolov, A.V.; Kostevich, V.A.; Gorbunov, N.P.; Runova, O.L.; Firova, E.M.; Vasilyev, V.B. Myeloperoxidase/high-density lipoprotein cholesterol ratio in patients with arterial hypertension and chronic coronary heart disease. Med. Acad. J. 2021, 21, 75–86. [Google Scholar] [CrossRef]
- Davis, B.J. Disc electrophoresis. II. Method and application to human serum proteins. Ann. N. Y. Acad. Sci. 1964, 121, 404–427. [Google Scholar] [CrossRef]
- Maurer, H.R. Disc Electrophoresis and Related Techniques of Polyacrylamide Gel Electrophoresis, 2nd ed.; de Gruyter, W., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1972. [Google Scholar] [CrossRef]
- Owen, C.A.; Smith, H. Detection of ceruloplasmin after zone electrophoresis. Clin. Chim. Acta 1961, 6, 441–444. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of Bacteriophag. Nature 1970, 227, 680–686. [Google Scholar] [CrossRef] [PubMed]
- White, K.N.; Conesa, C.; Sánchez, L.; Amini, M.; Farnaud, S.; Lorvoralak, C.; Evans, R.W. The transfer of iron between ceruloplasmin and transferrins. Biochim. Et Biophys. Acta 2012, 1820, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lu, M.; Chen, C.; Tong, X.; Li, Y.; Yang, K.; Lv, H.; Xu, J.; Qin, L. Holo-lactoferrin: The link between ferroptosis and radiotherapy in triple-negative breast cancer. Theranostics 2021, 11, 3167–3182. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Concentration, μM | ka, 104/m × c | kd, 1/c | Rmax, RU | KD, μM | U-Value |
|---|---|---|---|---|---|---|
| LF | 3.125–25.0 | 9.048 ± 1.1 | 0.03324 ± 0.0036 | (23.74 ± 0.18) ×103 | 0.367 | 7 |
| LF/8OA | 7.532 ± 0.58 | 0.03069 ± 0.002 | (22.34 ± 0.12) ×103 | 0.407 | 4 |
| Protein Concentration, μM | LF/8OA | BSA/8OA |
|---|---|---|
| MPO Concentration, ng/mL | ||
| 0 | 16.1 ± 3.5 | 20.3 ± 2.4 |
| 0.6 | 14.45 ± 1.0 | 15.9 ± 2.7 |
| 1.1 | 30.5 ± 2.4 | 13.6 ± 4.9 |
| 2.3 | 79.4 ± 2.9 | 16.3 ± 5.2 |
| 4.6 | 92.9 ± 7.2 | 23.4 ± 3.7 |
| 9.1 | 102.4 ± 11.5 | 31.1 ± 7.1 |
| 18.3 | 119.9 ± 12.1 | 57.9 ± 7.8 |
| 36.5 | 143.9 ± 9.5 | 76.1 ± 6.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elizarova, A.Y.; Sokolov, A.V.; Vasilyev, V.B. Ceruloplasmin Reduces the Lactoferrin/Oleic Acid Antitumor Complex-Mediated Release of Heme-Containing Proteins from Blood Cells. Int. J. Mol. Sci. 2023, 24, 16711. https://doi.org/10.3390/ijms242316711
Elizarova AY, Sokolov AV, Vasilyev VB. Ceruloplasmin Reduces the Lactoferrin/Oleic Acid Antitumor Complex-Mediated Release of Heme-Containing Proteins from Blood Cells. International Journal of Molecular Sciences. 2023; 24(23):16711. https://doi.org/10.3390/ijms242316711
Chicago/Turabian StyleElizarova, Anna Yu., Alexey V. Sokolov, and Vadim B. Vasilyev. 2023. "Ceruloplasmin Reduces the Lactoferrin/Oleic Acid Antitumor Complex-Mediated Release of Heme-Containing Proteins from Blood Cells" International Journal of Molecular Sciences 24, no. 23: 16711. https://doi.org/10.3390/ijms242316711