
Tocotrienol-Rich Fraction Attenuates Blue Light-Induced Oxidative Stress and Melanogenesis in B16-F1 Melanocytes via Anti-Oxidative and Anti-Tyrosinase Properties
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
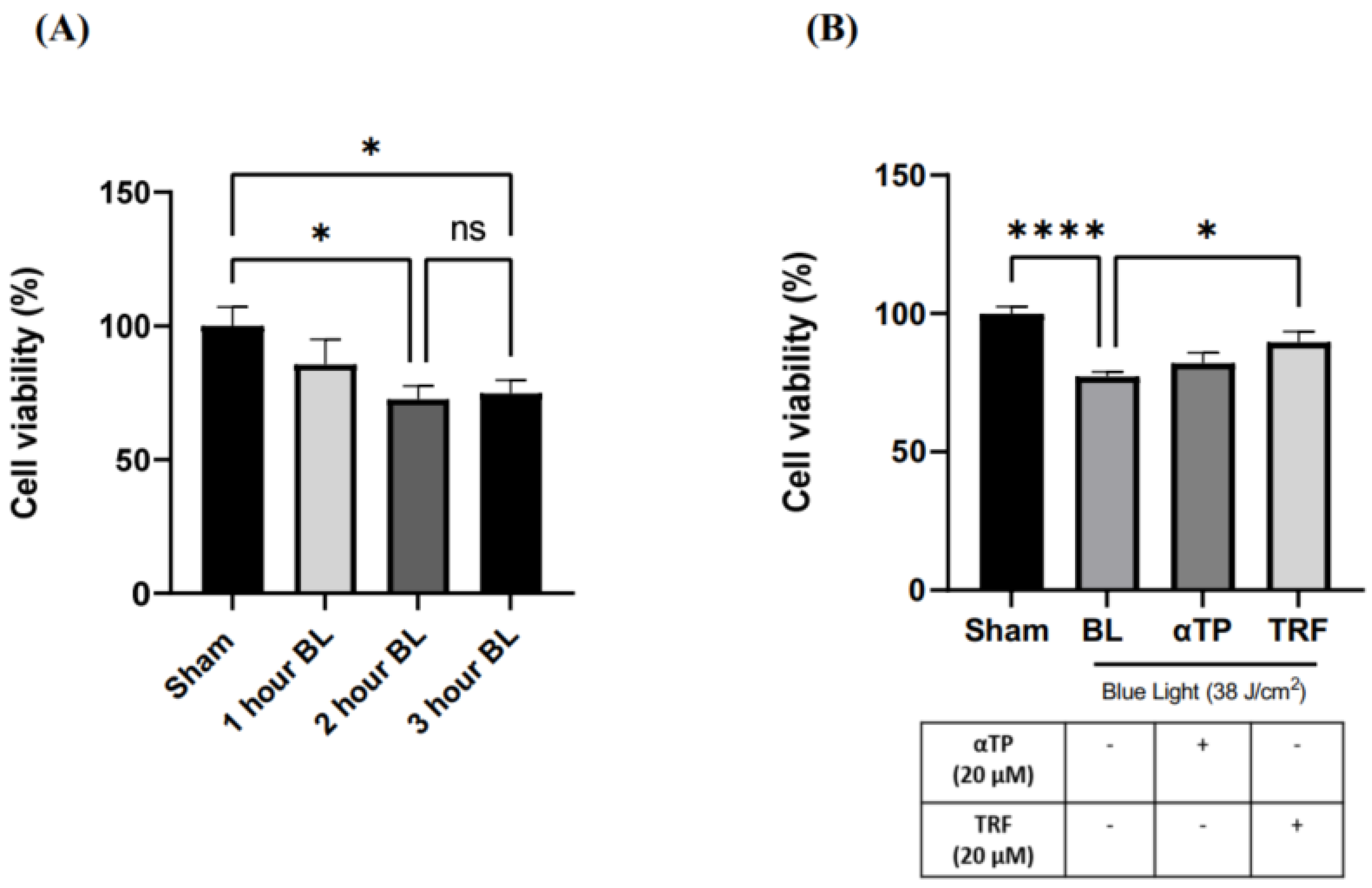
2.1. TRF Improved Cell Viability of BL-Induced Melanocytes
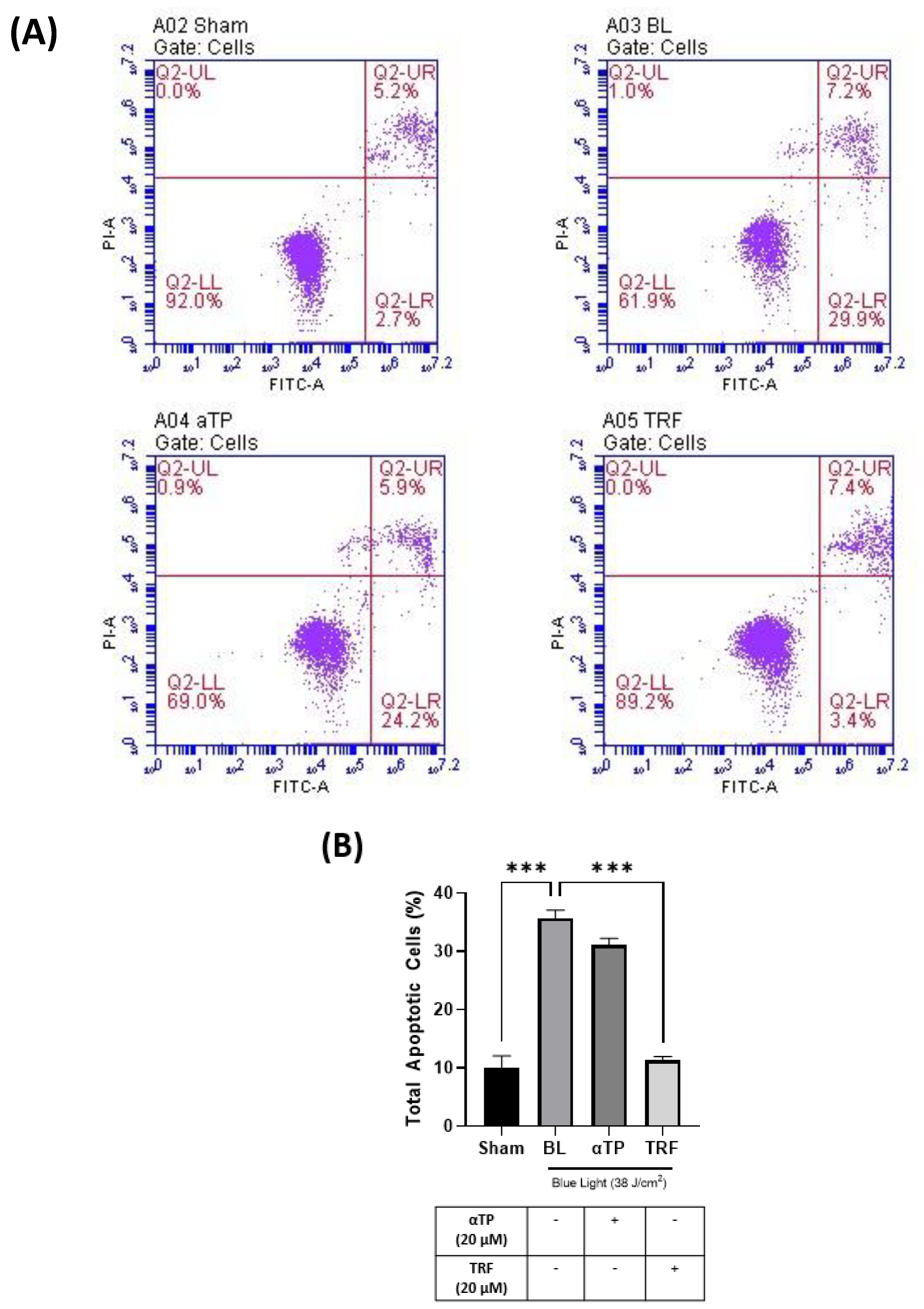
2.2. TRF Inhibited BL-Induced Cell Apoptosis
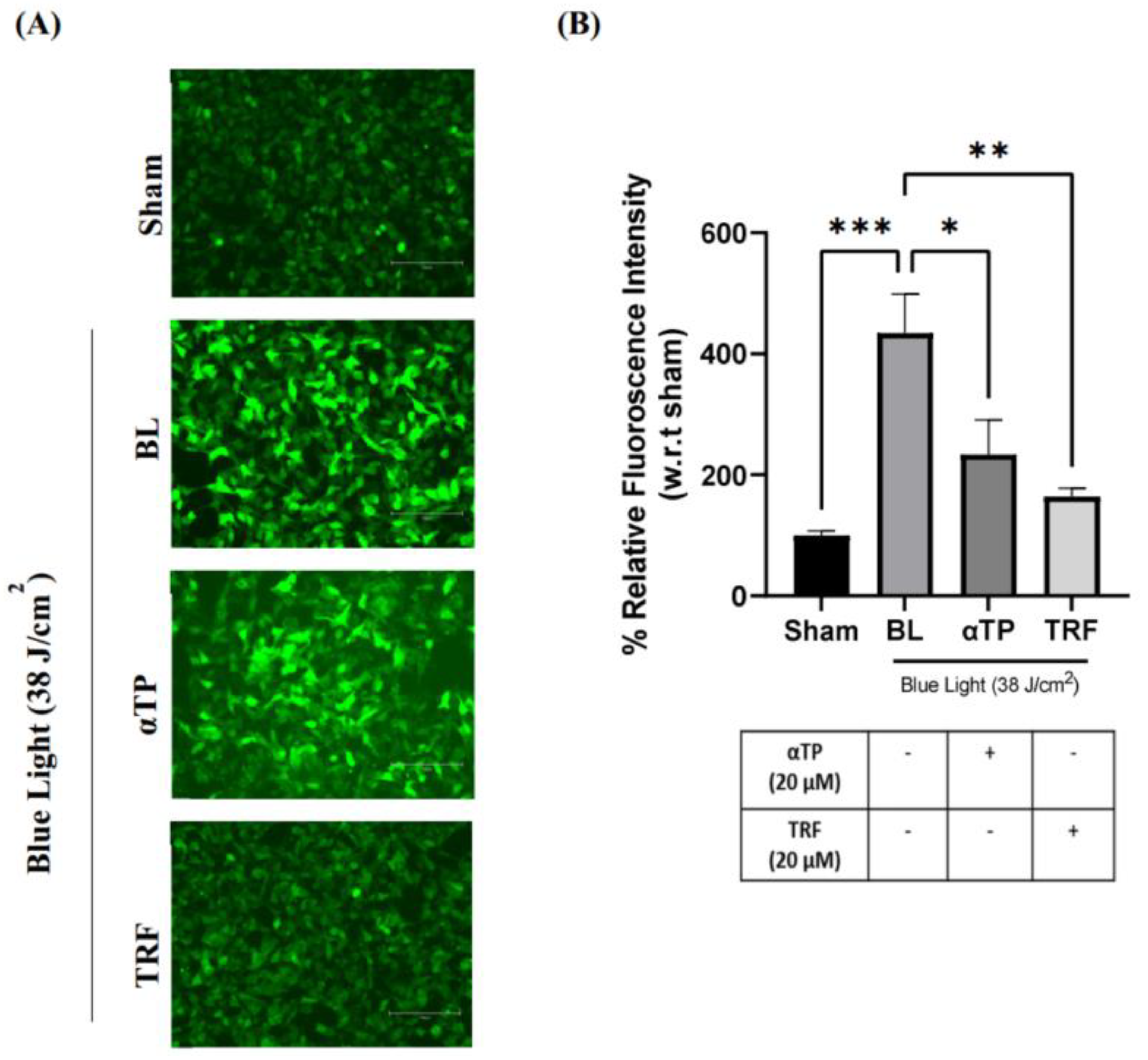
2.3. TRF Exhibited Anti-Oxidative Effects against Oxidative Stress Induced by BL
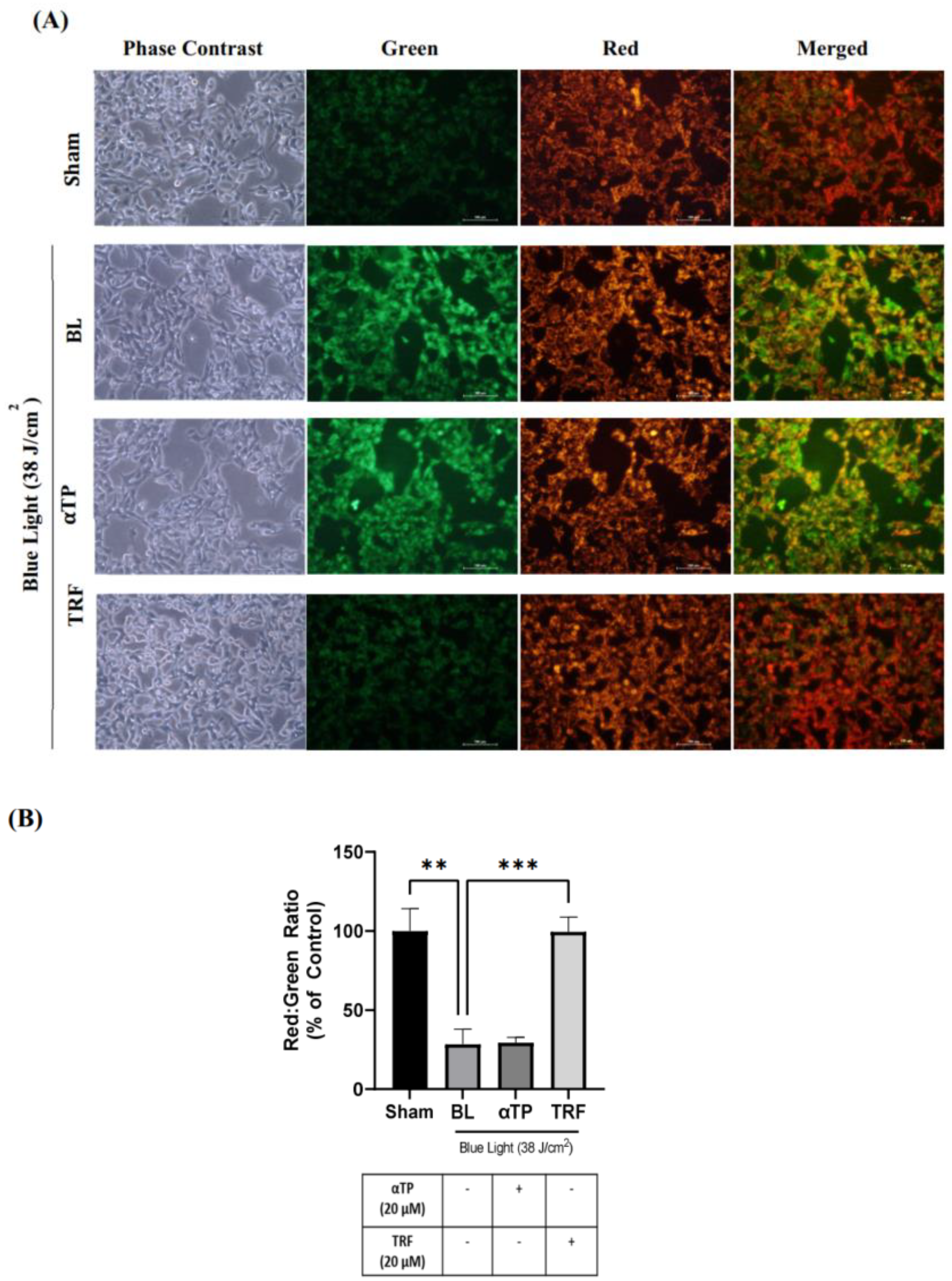
2.4. TRF Regulated BL-Induced Mitochondrial Membrane Potential Alterations
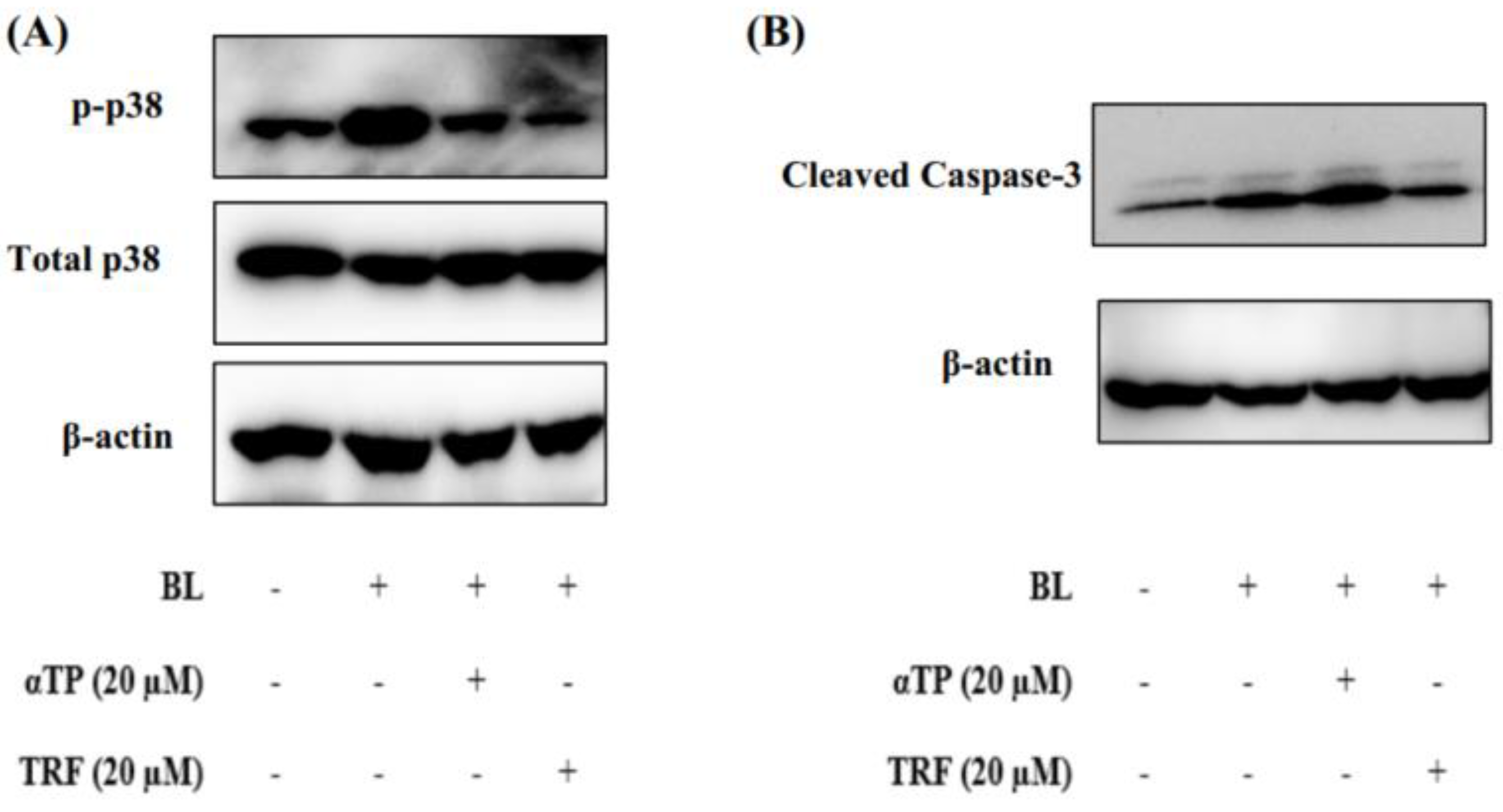
2.5. TRF Attenuated Cellular Death through Modulation of p38-MAPK Regulated Mitochondrial Apoptotic Pathway
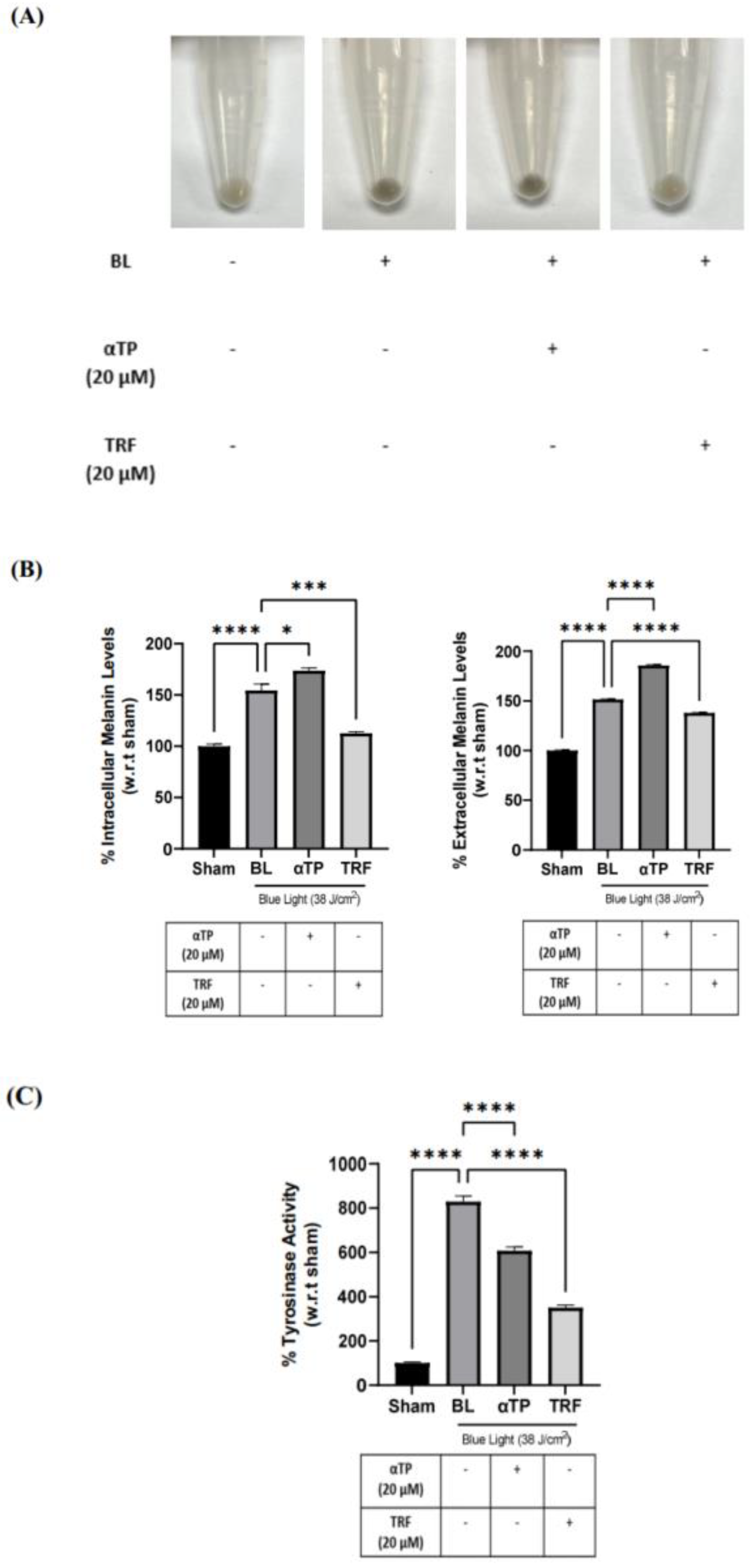
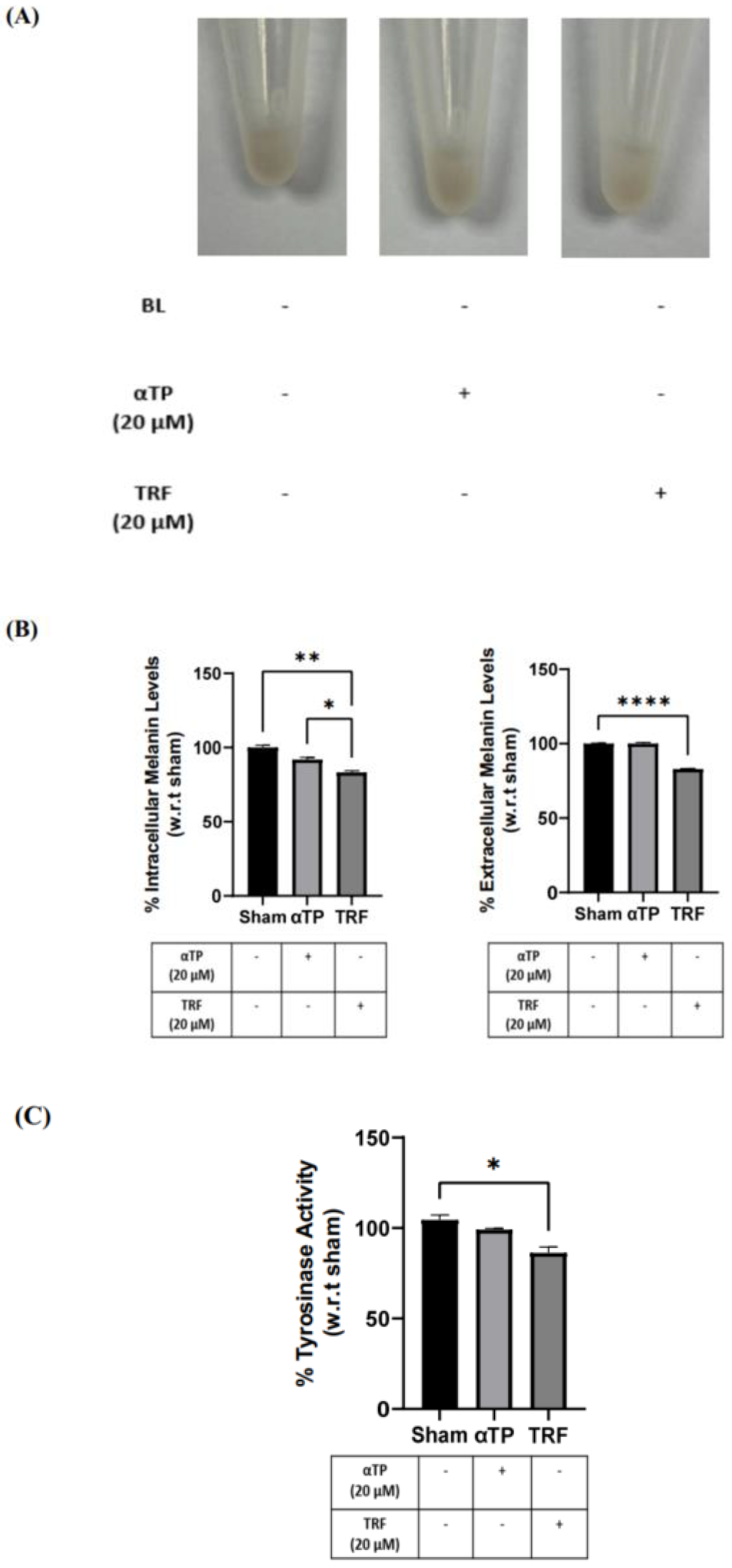
2.6. TRF Prevented Pigmentation Induced by BL through Regulation of Tyrosinase Activity
2.7. TRF Exhibited Anti-Melanogenic Effects
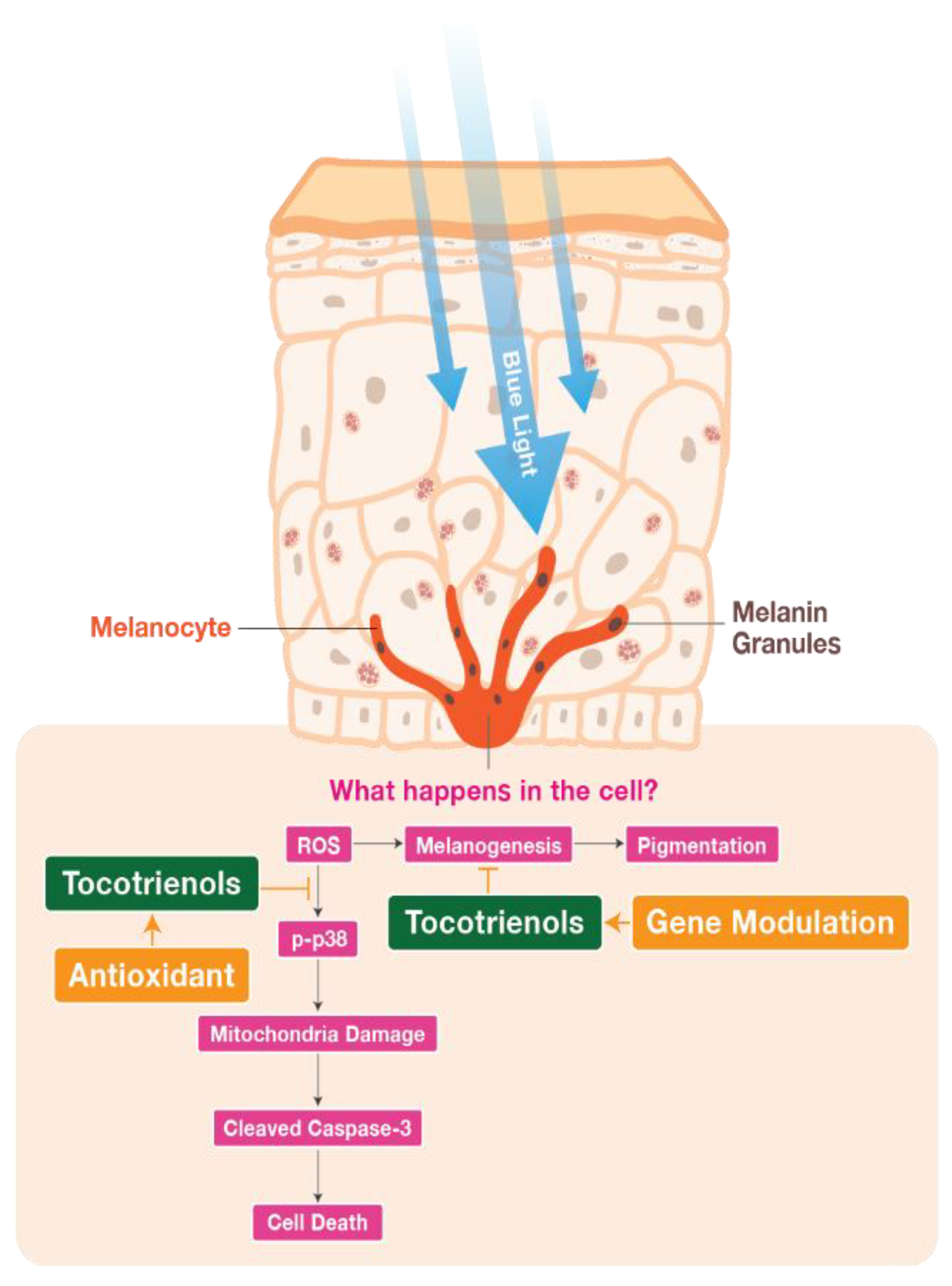
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. Natural Extract and Cell Treatment
4.3. Cell Irradiation
4.4. MTT Cell Viability Assay
4.5. Annexin-V Assay by Flow Cytometry
4.6. Intracellular ROS Assay
4.7. Mitochondrial Membrane Potential
4.8. Intracellular and Extracellular Melanin Content
4.9. Tyrosinase Activity
4.10. Western Blot
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernard, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin barrier immunity and ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Passeron, T.; Coelho, S.G.; Miyamura, Y.; Takahashi, K.; Hearing, V.J. Immunohistochemistry and in situ hybridization in the study of human skin melanocytes. Exp. Dermatol. 2007, 16, 162–170. [Google Scholar] [CrossRef]
- de Assis, L.V.M.; Tonolli, P.N.; Moraes, M.N.; Baptista, M.S.; de Lauro Castrucci, A.M. How does the skin sense sun light? An integrative view of light sensing molecules. J. Photochem. Photobiol. C Photochem. Rev. 2021, 47, 100403. [Google Scholar] [CrossRef]
- Modenese, A.; Korpinen, L.; Gobba, F. Solar radiation exposure and outdoor work: An underestimated occupational risk. Int. J. Environ. Res. Public Health 2018, 15, 2063. [Google Scholar] [CrossRef]
- O’Hagan, J.B.; Khazova, M.; Price, L.L. Low-energy light bulbs, computers, tablets and the blue light hazard. Eye 2016, 30, 230–233. [Google Scholar] [CrossRef]
- Lorrio, S.; Rodriguez-Luna, A.; Delgado-Wicke, P.; Mascaraque, M.; Gallego, M.; Perez-Davo, A.; González, S.; Juarranz, Á. Protective effect of the aqueous extract of deschampsia antarctica (EDAFENCE((R))) on skin cells against blue light emitted from digital devices. Int. J. Mol. Sci. 2020, 21, 988. [Google Scholar] [CrossRef]
- Portillo, M.; Mataix, M.; Alonso-Juarranz, M.; Lorrio, S.; Villalba, M.; Rodriguez-Luna, A.; González, S. The aqueous extract of polypodium leucotomos (Fernblock((R))) regulates opsin 3 and prevents photooxidation of melanin precursors on skin cells exposed to blue light emitted from digital devices. Antioxidants 2021, 10, 400. [Google Scholar] [CrossRef]
- Oh, P.S.; Na, K.S.; Hwang, H.; Jeong, H.S.; Lim, S.; Sohn, M.H.; Jeong, H.J. Effect of blue light emitting diodes on melanoma cells: Involvement of apoptotic signaling. J. Photochem. Photobiol. B 2015, 142, 197–203. [Google Scholar] [CrossRef]
- Sato, K.; Minai, Y.; Watanabe, H. Effect of monochromatic visible light on intracellular superoxide anion production and mitochondrial membrane potential of B16F1 and B16F10 murine melanoma cells. Cell Biol. Int. 2013, 37, 633–637. [Google Scholar] [CrossRef]
- Baek, J.; Lee, M.G. Oxidative stress and antioxidant strategies in dermatology. Redox Rep. 2016, 21, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Mohd Zaffarin, A.S.; Ng, S.F.; Ng, M.H.; Hassan, H.; Alias, E. Pharmacology and pharmacokinetics of vitamin E: Nanoformulations to enhance bioavailability. Int. J. Nanomed. 2020, 15, 9961–9974. [Google Scholar] [CrossRef] [PubMed]
- Berardesca, E.; Cameli, N. Vitamin E supplementation in inflammatory skin diseases. Dermatol. Ther. 2021, 34, e15160. [Google Scholar] [CrossRef] [PubMed]
- Thiele, J.J.; Ekanayake-Mudiyanselage, S. Vitamin E in human skin: Organ-specific physiology and considerations for its use in dermatology. Mol. Asp. Med. 2007, 28, 646–667. [Google Scholar] [CrossRef]
- Kanchi, M.M.; Shanmugam, M.K.; Rane, G.; Sethi, G.; Kumar, A.P. Tocotrienols: The unsaturated sidekick shifting new paradigms in vitamin E therapeutics. Drug Discov. Today 2017, 22, 1765–1781. [Google Scholar] [CrossRef]
- Neo, J.R.E.; Teo, Z.N.; Yeo, J.S.E.; Ng, C.K.S.; Teo, C.W.L.; Ung, Y.W.; Yap, W.N. Tocotrienols improve urban particulate matter-induced skin damages by regulating skin barrier function and ROS/MAPK signalling pathway in keratinocytes. Atmos. Pollut. Res. 2022, 13, 101564. [Google Scholar] [CrossRef]
- Yap, W.N.; Zaiden, N.; Xu, C.H.; Chen, A.; Ong, S.; Teo, V.; Yap, Y.L. Gamma- and delta-tocotrienols inhibit skin melanin synthesis by suppressing constitutive and UV-induced tyrosinase activation. Pigment Cell Melanoma Res. 2010, 23, 688–692. [Google Scholar] [CrossRef]
- Rascalou, A.; Lamartine, J.; Poydenot, P.; Demarne, F.; Bechetoille, N. Mitochondrial damage and cytoskeleton reorganization in human dermal fibroblasts exposed to artificial visible light similar to screen-emitted light. J. Dermatol. Sci. 2018, 91, 195–205. [Google Scholar] [CrossRef]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the Mitochondrial Membrane Potential Using the Cationic JC-1 Dye as a Sensitive Fluorescent Probe. Bio Protoc. 2019, 9, e3128. [Google Scholar] [CrossRef]
- Cui, S.; Nian, Q.; Chen, G.; Wang, X.; Zhang, J.; Qiu, J.; Zhang, Z. Ghrelin ameliorates A549 cell apoptosis caused by paraquat via p38-MAPK regulated mitochondrial apoptotic pathway. Toxicology 2019, 426, 152267. [Google Scholar] [CrossRef]
- Solano, F. Photoprotection and skin pigmentation: Melanin-related molecules and some other new agents obtained from natural sources. Molecules 2020, 25, 1537. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Ohta, S.; Wolf, A.M. Blue light-induced oxidative stress in live skin. Free Radic. Biol. Med. 2017, 108, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Yepes, J.; Burns, M.; Anandhan, A.; Khalimonchuk, O.; del Razo, L.M.; Quintanilla-Vega, B.; Pappa, A.; Panayiotidis, M.I.; Franco, R. Oxidative stress, redox signaling, and autophagy: Cell death versus survival. Antioxid. Redox Signal. 2014, 21, 66–85. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, R.; Mathai, M.; Zulli, A. Revisiting the therapeutic potential of tocotrienol. Biofactors 2022, 48, 813–856. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Grab, J.; Rybniker, J. The expanding role of p38 mitogen-activated protein kinase in programmed host cell death. Microbiol. Insights 2019, 12, 1178636119864594. [Google Scholar] [CrossRef]
- Eskandari, E.; Eaves, C.J. Paradoxical roles of caspase-3 in regulating cell survival, proliferation, and tumorigenesis. J. Cell Biol. 2022, 221, e202201159. [Google Scholar] [CrossRef] [PubMed]
- Satyamitra, M.; Ney, P.; Graves, J., 3rd; Mullaney, C.; Srinivasan, V. Mechanism of radioprotection by delta-tocotrienol: Pharmacokinetics, pharmacodynamics and modulation of signalling pathways. Br. J. Radiol. 2012, 85, e1093–e1103. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y. Skin pigmentation abnormalities and their possible relationship with skin aging. Int. J. Mol. Sci. 2021, 22, 3727. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Shim, S.Y.; Lee, Y.E.; Lee, M. Antioxidant compounds, kirenol and methyl ent-16alpha, 17-dihydroxy-kauran-19-oate bioactivity-guided isolated from siegesbeckia glabrescens attenuates MITF-mediated melanogenesis via inhibition of intracellular ROS production. Molecules 2021, 26, 1940. [Google Scholar] [CrossRef]
- Yu, F.; Lu, Y.; Zhong, Z.; Qu, B.; Wang, M.; Yu, X.; Chen, J. Mitf involved in innate immunity by activating tyrosinase-mediated melanin synthesis in pteria penguin. Front. Immunol. 2021, 12, 626493. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, D.W.; Kim, S.; Choi, H.I.; Lee, Y.; Kim, C.D.; Lee, J.H.; Do Lee, S.; Lee, Y.H. Downregulation of NFAT2 promotes melanogenesis in B16 melanoma cells. Anat. Cell Biol. 2010, 43, 303–309. [Google Scholar] [CrossRef]
- Suitthimeathegorn, O.; Yang, C.; Ma, Y.; Liu, W. Direct and indirect effects of blue light exposure on skin: A review of published literature. Skin Pharmacol. Physiol. 2022, 35, 305–318. [Google Scholar] [CrossRef]
- Godley, B.F.; Shamsi, F.A.; Liang, F.Q.; Jarrett, S.G.; Davies, S.; Boulton, M. Blue light induces mitochondrial DNA damage and free radical production in epithelial cells. J. Biol. Chem. 2005, 280, 21061–21066. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neo, J.R.E.; Teo, C.W.L.; Ung, Y.W.; Yap, W.N. Tocotrienol-Rich Fraction Attenuates Blue Light-Induced Oxidative Stress and Melanogenesis in B16-F1 Melanocytes via Anti-Oxidative and Anti-Tyrosinase Properties. Int. J. Mol. Sci. 2023, 24, 15373. https://doi.org/10.3390/ijms242015373
Neo JRE, Teo CWL, Ung YW, Yap WN. Tocotrienol-Rich Fraction Attenuates Blue Light-Induced Oxidative Stress and Melanogenesis in B16-F1 Melanocytes via Anti-Oxidative and Anti-Tyrosinase Properties. International Journal of Molecular Sciences. 2023; 24(20):15373. https://doi.org/10.3390/ijms242015373
Chicago/Turabian StyleNeo, Juvenia Rui En, Cheryl Wei Ling Teo, Yee Wei Ung, and Wei Ney Yap. 2023. "Tocotrienol-Rich Fraction Attenuates Blue Light-Induced Oxidative Stress and Melanogenesis in B16-F1 Melanocytes via Anti-Oxidative and Anti-Tyrosinase Properties" International Journal of Molecular Sciences 24, no. 20: 15373. https://doi.org/10.3390/ijms242015373