
Two-Dimensional Polyacrylamide Gel Electrophoresis Coupled with Nanoliquid Chromatography–Tandem Mass Spectrometry-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in the MCF7 Breast Cancer Cell Line Transfected for Jumping Translocation Breakpoint Protein Overexpression


 ,
,
Abstract
:1. Introduction
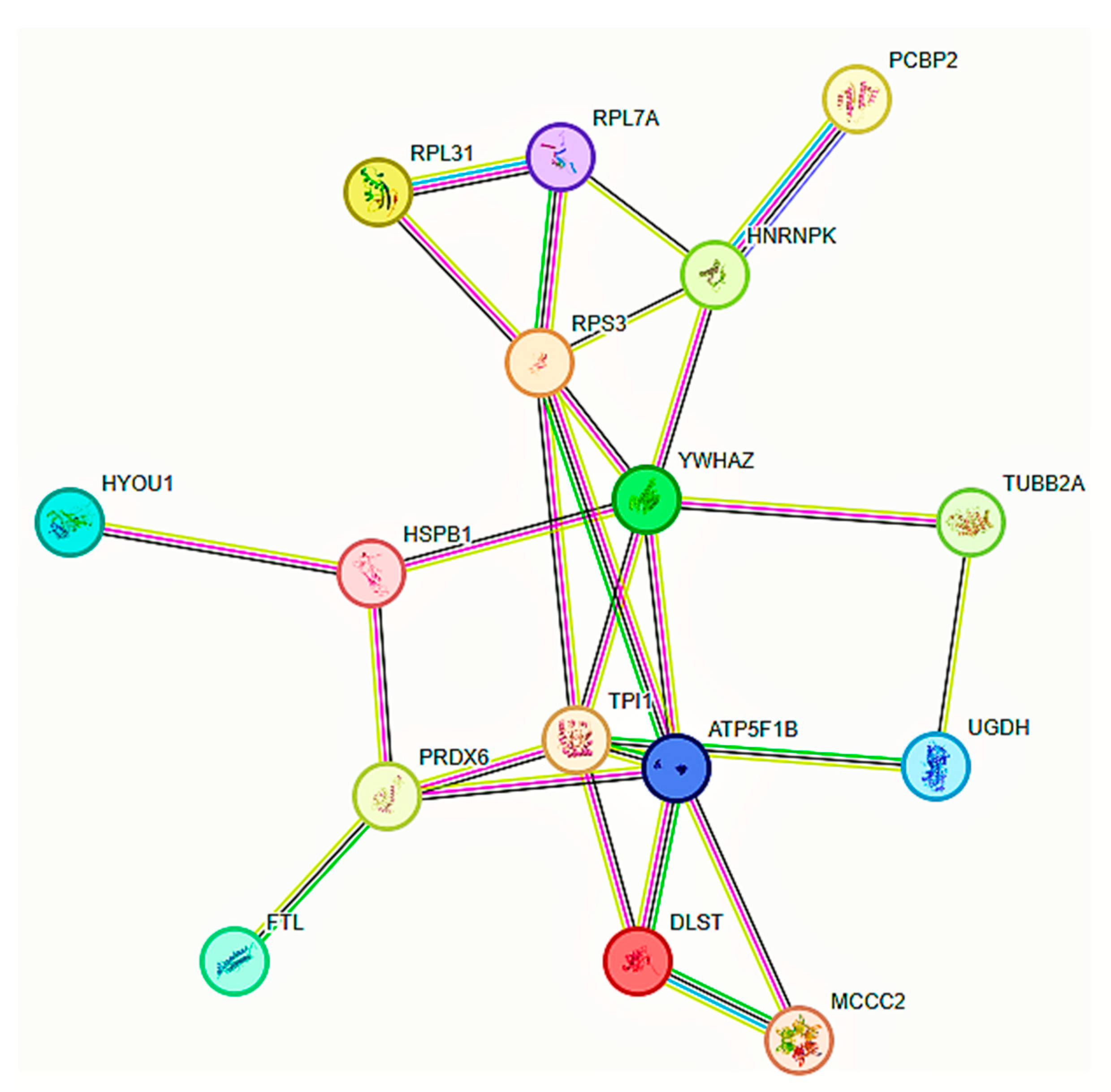
2. Results
3. Discussion
3.1. Pro-Tumorigenic (PT) JTB-Interacting Partners
3.2. Anti-Tumorigenic (AT) JTB-Interacting Partners
3.3. JTB-Interacting Partners with Controversial Neoplastic Functions
4. Materials and Methods
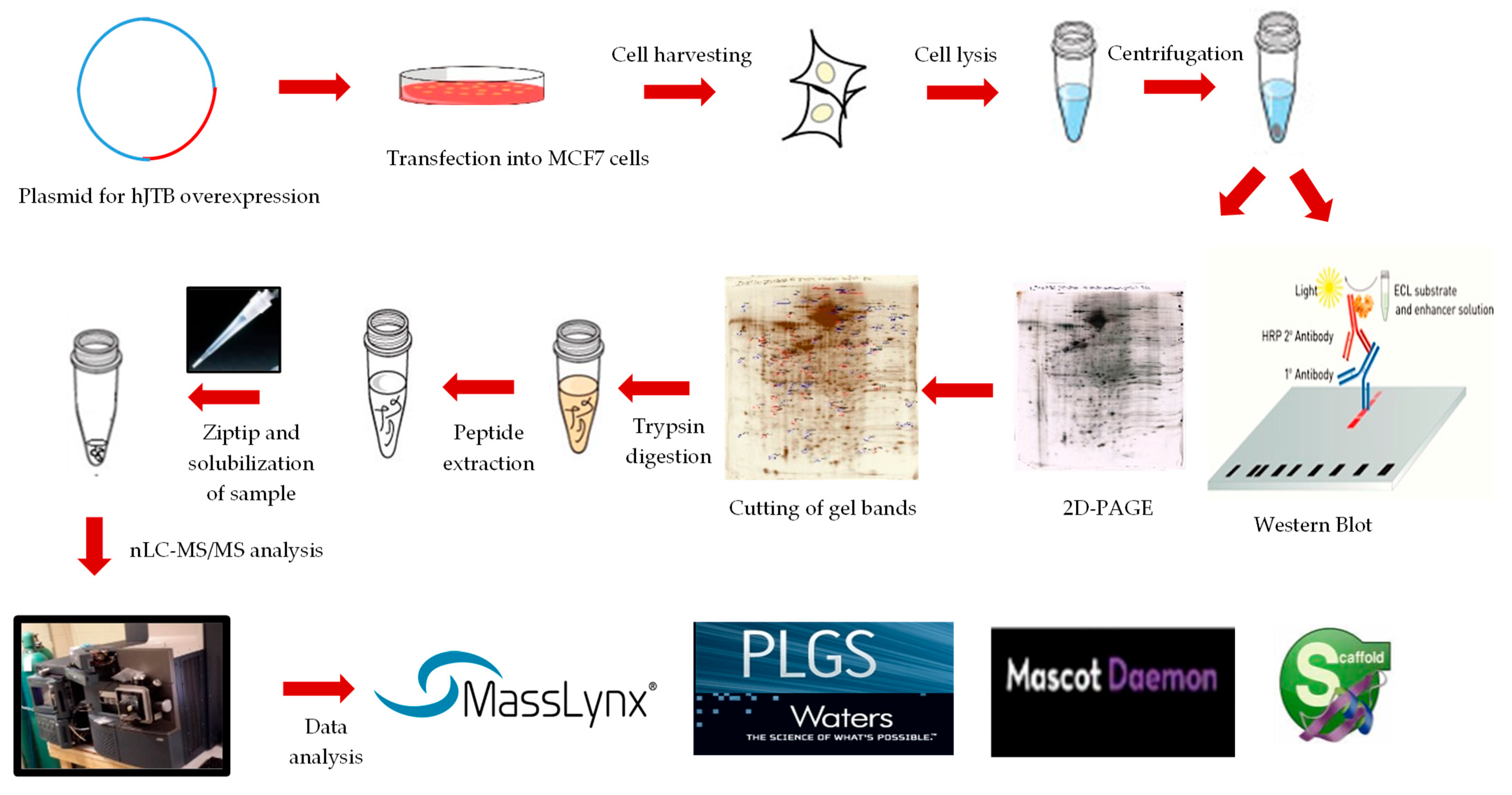
4.1. Cell Culture
4.2. Plasmids for Upregulation
4.3. Transfection into MCF7 Cells
4.4. Western Blot Analysis
4.5. 2D-PAGE and Proteomic Analysis
4.6. Data Sharing
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, R.; Sultan, A.; Ishrat, R.; Haque, S.; Khan, N.J.; Prieto, M.A. Identification of New Key Genes and Their Association with Breast Cancer Occurrence and Poor Survival Using In Silico and In Vitro Methods. Biomedicines 2023, 11, 1271. [Google Scholar] [CrossRef] [PubMed]
- Kanome, T.; Itoh, N.; Ishikawa, F.; Mori, K.; Kim-Kaneyama, J.R.; Nose, K.; Shibanuma, M. Characterization of Jumping translocation breakpoint (JTB) gene product isolated as a TGF-β1-inducible clone involved in regulation of mitochondrial function, cell growth and cell death. Oncogene 2007, 26, 5991–6001. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, S.; Osawa, M.; Omine, M.; Ishikawa, F. JTB: A novel membrane protein gene at 1q21 rearranged in a jumping translocation. Oncogene 1999, 18, 2085–2090. [Google Scholar] [CrossRef] [PubMed]
- Stankiewicz, P.; Cheung, S.W.; Shaw, C.J.; Saleki, R.; Szigeti, K.; Lupski, J.R. The donor chromosome breakpoint for a jumping translocation is associated with large low-copy repeats in 21q21.3. Cytogenet. Genome Res. 2003, 101, 118–123. [Google Scholar] [CrossRef]
- Pareja, F.; Selenica, P.; Brown, D.N.; Sebastiao, A.P.M.; da Silva, E.M.; Da Cruz Paula, A.; Del, A.; Fu, L.; Weigelt, B.; Brogi, E.; et al. Micropapillary variant of mucinous carcinoma of the breast shows genetic alterations intermediate between those of mucinous carcinoma and micropapillary carcinoma. Histopathology 2019, 75, 139–145. [Google Scholar] [CrossRef]
- Verras, G.-I.; Tchabashvili, L.; Mulita, F.; Grypari, I.M.; Sourouni, S.; Panagodimou, E.; Argentou, M.-I. Micropapillary Breast Carcinoma: From Molecular Pathogenesis to Prognosis. Breast Cancer 2022, 14, 41–61. [Google Scholar] [CrossRef]
- Elder, J.T.; Zhao, X. Evidence for local control of gene expression in the epidermal differentiation complex. Exp. Dermatol. 2002, 11, 406–412. [Google Scholar] [CrossRef]
- Poterlowicz, K.; Yarker, J.L.; Malashchuk, I.; Lajoie, B.R.; Mardaryev, A.N.; Gdula, M.R.; Sharov, A.A.; Kohwi-Shigematsu, T.; Botchkarev, V.A.; Fessing, M.Y. 5C analysis of the Epidermal Differentiation Complex locus reveals distinct chromatin interaction networks between gene-rich and gene-poor TADs in skin epithelial cells. PLoS Genet. 2017, 13, e1006966. [Google Scholar] [CrossRef]
- Abhishek, S.; Palamadai Krishnan, S. Epidermal Differentiation Complex: A Review on Its Epigenetic Regulation and Potential Drug Targets. Cell J. 2016, 18, 1–6. [Google Scholar] [CrossRef]
- Qin, D.; Ma, L.; Qin, L. Potential Role of the Epidermal Differentiation Complex in the Pathogenesis of Psoriasis. FBL 2022, 27, 325. [Google Scholar] [CrossRef]
- Tyszkiewicz, T.; Jarząb, M.; Szymczyk, C.; Halczok, M.; Krajewska, J.; Jaworska, M.; Fraczek, M.; Krajewska, A.; Hadas, E.; Swierniak, M.; et al. Epidermal differentiation complex (locus 1q21) gene expression in head and neck cancer and normal mucosa. Folia Histochem. Cytobiol. 2014, 52, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Platica, M.; Ionescu, A.; Ivan, E.; Holland, J.; Mandeli, J.; Platica, O. PAR, a protein involved in the cell cycle, is functionally related to chromosomal passenger proteins. Int. J. Oncol. 2011, 38, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Platica, O.; Chen, S.; Iván, E.; Lopingco, M.; Holland, J.; Platica, M. PAR, a novel androgen regulated gene, ubiquitously expressed in normal and malignant cells. Int. J. Oncol. 2000, 16, 1055–1061. [Google Scholar] [CrossRef]
- Wong, N.; Chan, A.; Lee, S.-W.; Lam, E.; To, K.-F.; Lai, P.B.S.; Li, X.-N.; Liew, C.-T.; Johnson, P.J. Positional mapping for amplified DNA sequences on 1q21–q22 in hepatocellular carcinoma indicates candidate genes over-expression. J. Hepatol. 2003, 38, 298–306. [Google Scholar] [CrossRef]
- Jayathirtha, M.; Neagu, A.-N.; Whitham, D.; Alwine, S.; Darie, C. Investigation of the effects of overexpression of jumping translocation breakpoint (JTB) protein in MCF7 cells for potential use as a biomarker in breast cancer. Am. J. Cancer Res. 2022, 12, 1784–1823. [Google Scholar] [PubMed]
- Jayathirtha, M.; Whitham, D.; Alwine, S.; Donnelly, M.; Neagu, A.-N.; Darie, C.C. Investigating the Function of Human Jumping Translocation Breakpoint Protein (hJTB) and Its Interacting Partners through In-Solution Proteomics of MCF7 Cells. Molecules 2022, 27, 8301. [Google Scholar] [CrossRef]
- Guo, Q.; Cheng, K.; Wang, X.; Li, X.; Yu, Y.; Hua, Y.; Yang, Z. Expression of HDAC1 and RBBP4 correlate with clinicopathologic characteristics and prognosis in breast cancer. Int. J. Clin. Exp. Pathol. 2020, 13, 563–572. [Google Scholar]
- Schultz-Rogers, L.E.; Thayer, M.L.; Kambakam, S.; Wierson, W.A.; Helmer, J.A.; Wishman, M.D.; Wall, K.A.; Greig, J.L.; Forsman, J.L.; Puchhalapalli, K.; et al. Rbbp4 loss disrupts neural progenitor cell cycle regulation independent of Rb and leads to Tp53 acetylation and apoptosis. Dev. Dyn. 2022, 251, 1267–1290. [Google Scholar] [CrossRef]
- Zheng, Z.; Yao, X.; Liu, Y. RBBP4 plays a vital role in the malignant progression of triple-negative breast cancer by regulating epithelial-mesenchymal transition. Genes Genom. 2022, 44, 1301–1309. [Google Scholar] [CrossRef]
- Li, Y.-D.; Lv, Z.; Zhu, W.-F. RBBP4 promotes colon cancer malignant progression via regulating Wnt/β-catenin pathway. World J. Gastroenterol. 2020, 26, 5328–5342. [Google Scholar] [CrossRef]
- Bayarkhangai, B.; Noureldin, S.; Yu, L.; Zhao, N.; Gu, Y.; Xu, H.; Guo, C. A comprehensive and perspective view of oncoprotein SET in cancer. Cancer Med. 2018, 7, 3084–3094. [Google Scholar] [CrossRef] [PubMed]
- Janghorban, M.; Farrell, A.S.; Allen-Petersen, B.L.; Pelz, C.; Daniel, C.J.; Oddo, J.; Langer, E.M.; Christensen, D.J.; Sears, R.C. Targeting c-MYC by antagonizing PP2A inhibitors in breast cancer. Proc. Natl. Acad. Sci. USA 2014, 111, 9157–9162. [Google Scholar] [CrossRef] [PubMed]
- Mody, H.R.; Hung, S.W.; Naidu, K.; Lee, H.; Gilbert, C.A.; Hoang, T.T.; Pathak, R.K.; Manoharan, R.; Muruganandan, S.; Govindarajan, R. SET contributes to the epithelial-mesenchymal transition of pancreatic cancer. Oncotarget 2017, 8, 67966–67979. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, H.; Ren, C.; Wang, L. High Expression of ABRACL Is Associated with Tumorigenesis and Affects Clinical Outcome in Gastric Cancer. Genet. Test. Mol. Biomark. 2019, 23, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, H. Actin-binding Rho activating C-terminal like (ABRACL) transcriptionally regulated by MYB proto-oncogene like 2 (MYBL2) promotes the proliferation, invasion, migration and epithelial-mesenchymal transition of breast cancer cells. Bioengineered 2022, 13, 9019–9031. [Google Scholar] [CrossRef] [PubMed]
- Ura, B.; Monasta, L.; Arrigoni, G.; Franchin, C.; Radillo, O.; Peterlunger, I.; Ricci, G.; Scrimin, F. A proteomic approach for the identification of biomarkers in endometrial cancer uterine aspirate. Oncotarget 2017, 8, 109536–109545. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Chen, P.; Li, S. MiR-145-5p Inhibits the Proliferation, Migration, and Invasion of Esophageal Carcinoma Cells by Targeting ABRACL. BioMed Res. Int. 2021, 2021, 6692544. [Google Scholar] [CrossRef]
- Hsiao, B.-Y.; Chen, C.-H.; Chi, H.-Y.; Yen, P.-R.; Yu, Y.-Z.; Lin, C.-H.; Pang, T.-L.; Lin, W.-C.; Li, M.-L.; Yeh, Y.-C.; et al. Human Costars Family Protein ABRACL Modulates Actin Dynamics and Cell Migration and Associates with Tumorigenic Growth. Int. J. Mol. Sci. 2021, 22, 2037. [Google Scholar] [CrossRef]
- Afzal, T.A.; Luong, L.A.; Chen, D.; Zhang, C.; Yang, F.; Chen, Q.; An, W.; Wilkes, E.; Yashiro, K.; Cutillas, P.R.; et al. NCK Associated Protein 1 Modulated by miRNA-214 Determines Vascular Smooth Muscle Cell Migration, Proliferation, and Neointima Hyperplasia. J. Am. Heart Assoc. 2016, 5, e004629. [Google Scholar] [CrossRef]
- Morris, D.C.; Popp, J.L.; Tang, L.K.; Gibbs, H.C.; Schmitt, E.; Chaki, S.P.; Bywaters, B.C.; Yeh, A.T.; Porter, W.W.; Burghardt, R.C.; et al. Nck deficiency is associated with delayed breast carcinoma progression and reduced metastasis. Mol. Biol. Cell 2017, 28, 3500–3516. [Google Scholar] [CrossRef]
- Kwon, M.R.; Lee, J.H.; Park, J.; Park, S.S.; Ju, E.J.; Ko, E.J.; Shin, S.H.; Son, G.W.; Lee, H.W.; Kim, Y.J.; et al. NCK-associated protein 1 regulates metastasis and is a novel prognostic marker for colorectal cancer. Cell Death Discov. 2023, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; He, L.; Shay, C.; Lang, L.; Loveless, J.; Yu, J.; Chemmalakuzhy, R.; Jiang, H.; Liu, M.; Teng, Y. Nck-associated protein 1 associates with HSP90 to drive metastasis in human non-small-cell lung cancer. J. Exp. Clin. Cancer Res. 2019, 38, 122. [Google Scholar] [CrossRef] [PubMed]
- Alaee, M.; Nool, K.; Pasdar, M. Plakoglobin restores tumor suppressor activity of p53(R175H) mutant by sequestering the oncogenic potential of β-catenin. Cancer Sci. 2018, 109, 1876–1888. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Gao, F.; Papageorgis, P.; Abdolmaleky, H.M.; Faller, D.V.; Thiagalingam, S. Aberrant activation of γ-catenin promotes genomic instability and oncogenic effects during tumor progression. Cancer Biol. Ther. 2007, 6, 1638–1643. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Ji, H.; Yin, L.; Niu, X.; Wang, Y.; Liu, Y.; Xuan, Q.; Li, L.; Zhang, H.; Zhou, X.; et al. High Expression of Plakoglobin Promotes Metastasis in Invasive Micropapillary Carcinoma of the Breast via Tumor Cluster Formation. J. Cancer 2019, 10, 2800–2810. [Google Scholar] [CrossRef] [PubMed]
- Spethmann, T.; Böckelmann, L.C.; Labitzky, V.; Ahlers, A.-K.; Schröder-Schwarz, J.; Bonk, S.; Simon, R.; Sauter, G.; Huland, H.; Kypta, R.; et al. Opposing prognostic relevance of junction plakoglobin in distinct prostate cancer patient subsets. Mol. Oncol. 2021, 15, 1956–1969. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.G.; Pearn, L.; Liddiard, K.; Pumford, S.L.; Burnett, A.K.; Tonks, A.; Darley, R.L. γ-Catenin is overexpressed in acute myeloid leukemia and promotes the stabilization and nuclear localization of β-catenin. Leukemia 2013, 27, 336–343. [Google Scholar] [CrossRef]
- Fang, W.-K.; Liao, L.-D.; Gu, W.; Chen, B.; Wu, Z.-Y.; Wu, J.-Y.; Shen, J.; Xu, L.-Y.; Li, E.-M. Down-regulated γ-catenin expression is associated with tumor aggressiveness in esophageal cancer. World J. Gastroenterol. 2014, 20, 5839–5848. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, L.; Qin, Y.; Liu, S.; Qiao, Y.; Wan, X.; Zeng, H.; Tang, X.; Liu, M.; Hou, Y. Effects of differential distributed-JUP on the malignancy of gastric cancer. J. Adv. Res. 2021, 28, 195–208. [Google Scholar] [CrossRef]
- Yang, L.; Chen, Y.; Cui, T.; Knösel, T.; Zhang, Q.; Albring, K.F.; Huber, O.; Petersen, I. Desmoplakin acts as a tumor suppressor by inhibition of the Wnt/β-catenin signaling pathway in human lung cancer. Carcinogenesis 2012, 33, 1863–1870. [Google Scholar] [CrossRef]
- Leick, K.M.; Rodriguez, A.B.; Melssen, M.M.; Benamar, M.; Lindsay, R.S.; Eki, R.; Du, K.-P.; Parlak, M.; Abbas, T.; Engelhard, V.H.; et al. The Barrier Molecules Junction Plakoglobin, Filaggrin, and Dystonin Play Roles in Melanoma Growth and Angiogenesis. Ann. Surg. 2019, 270, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Meng, Y.; Wang, Y.; Li, J.; Lam, M.; Wang, L.; Di, L.-j. Nuclear localization of Desmoplakin and its involvement in telomere maintenance. Int. J. Biol. Sci. 2019, 15, 2350–2362. [Google Scholar] [CrossRef] [PubMed]
- Morishita, E.C.; Murayama, K.; Kato-Murayama, M.; Ishizuka-Katsura, Y.; Tomabechi, Y.; Hayashi, T.; Terada, T.; Handa, N.; Shirouzu, M.; Akiyama, T.; et al. Crystal Structures of the Armadillo Repeat Domain of Adenomatous Polyposis Coli and Its Complex with the Tyrosine-Rich Domain of Sam68. Structure 2011, 19, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Minde, D.P.; Anvarian, Z.; Rüdiger, S.G.D.; Maurice, M.M. Messing up disorder: How do missense mutations in the tumor suppressor protein APC lead to cancer? Mol. Cancer 2011, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Taketo, M.M. Adenomatous polyposis coli (APC): A multi-functional tumor suppressor gene. J. Cell Sci. 2007, 120, 3327–3335. [Google Scholar] [CrossRef] [PubMed]
- Adrain, C.; Creagh, E.M.; Martin, S.J. Apoptosis-associated release of Smac/DIABLO from mitochondria requires active caspases and is blocked by Bcl-2. EMBO J. 2001, 20, 6627–6636. [Google Scholar] [CrossRef] [PubMed]
- Pluta, P.; Cebula-Obrzut, B.; Ehemann, V.; Pluta, A.; Wierzbowska, A.; Piekarski, J.H.; Bilski, A.; Nejc, D.; Kordek, R.; Robak, T.; et al. Correlation of Smac/DIABLO protein expression with the clinico-pathological features of breast cancer patients. Neoplasma 2011, 58, 430–435. [Google Scholar] [CrossRef]
- Paul, A.; Krelin, Y.; Arif, T.; Jeger, R.; Shoshan-Barmatz, V. A New Role for the Mitochondrial Pro-apoptotic Protein SMAC/Diablo in Phospholipid Synthesis Associated with Tumorigenesis. Mol. Ther. 2018, 26, 680–694. [Google Scholar] [CrossRef]
- Pandey, S.K.; Shteinfer-Kuzmine, A.; Chalifa-Caspi, V.; Shoshan-Barmatz, V. Non-apoptotic activity of the mitochondrial protein SMAC/Diablo in lung cancer: Novel target to disrupt survival, inflammation, and immunosuppression. Front. Oncol. 2022, 12, 992260. [Google Scholar] [CrossRef]
- Martin, J.; Magnino, F.; Schmidt, K.; Piguet, A.-C.; Lee, J.-S.; Semela, D.; St-Pierre, M.V.; Ziemiecki, A.; Cassio, D.; Brenner, C.; et al. Hint2, a mitochondrial apoptotic sensitizer down-regulated in hepatocellular carcinoma. Gastroenterology 2006, 130, 2179–2188. [Google Scholar] [CrossRef]
- Martin, J.; Maurhofer, O.; Bellance, N.; Benard, G.; Graber, F.; Hahn, D.; Galinier, A.; Hora, C.; Gupta, A.; Ferrand, G.; et al. Disruption of the histidine triad nucleotide-binding hint2 gene in mice affects glycemic control and mitochondrial function. Hepatology 2013, 57, 2037–2048. [Google Scholar] [CrossRef]
- Li, W.; Cai, S.; Wang, L.; Yang, C.; Zhou, B.; Wang, H. HINT2 downregulation promotes colorectal carcinoma migration and metastasis. Oncotarget 2017, 8, 13521–13531. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Sun, Q.; Zhou, D.; Song, W.; Yang, Q.; Ju, B.; Zhang, L.; Xie, H.; Zhou, L.; Hu, Z.; et al. HINT2 triggers mitochondrial Ca2+ influx by regulating the mitochondrial Ca2+ uniporter (MCU) complex and enhances gemcitabine apoptotic effect in pancreatic cancer. Cancer Lett. 2017, 411, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.; St-Pierre, M.V.; Dufour, J.-F. Hit proteins, mitochondria and cancer. Biochim. Biophys. Acta—Bioenerg. 2011, 1807, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yu, J.; Wu, R.; Shi, L.; Zhang, X.; Zhang, W.; Zhong, X.; Wang, Y.; Li, H.; Shen, Y.; et al. GRP78 determines glioblastoma sensitivity to UBA1 inhibition-induced UPR signaling and cell death. Cell Death Dis. 2021, 12, 733. [Google Scholar] [CrossRef] [PubMed]
- Groen, E.J.N.; Gillingwater, T.H. UBA1: At the Crossroads of Ubiquitin Homeostasis and Neurodegeneration. Trends Mol. Med. 2015, 21, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Yang, G.; Huang, H.; Zhou, Y.; Hu, X.; Lu, Q.; Guo, P.; Hou, J.; Cao, L.; Tian, F.; et al. Ubiquitin-Like Modifier Activating Enzyme 1 as a Novel Diagnostic and Prognostic Indicator That Correlates With Ferroptosis and the Malignant Phenotypes of Liver Cancer Cells. Front. Oncol. 2020, 10, 592413. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Turner, T.H.; Cai, J.; Floros, K.V.; Yu, A.K.; Coon, C.M.; Khatri, R.; Alzubi, M.A.; Jakubik, C.T.; Bouck, Y.M.; et al. Genomic screening reveals ubiquitin-like modifier activating enzyme 1 as a potent and druggable target in c-MYC-high triple negative breast cancer models. PNAS Nexus 2022, 1, pgac232. [Google Scholar] [CrossRef]
- Majeed, S.; Aparnathi, M.K.; Nixon, K.C.J.; Venkatasubramanian, V.; Rahman, F.; Song, L.; Weiss, J.; Barayan, R.; Sugumar, V.; Barghout, S.H.; et al. Targeting the Ubiquitin–Proteasome System Using the UBA1 Inhibitor TAK-243 is a Potential Therapeutic Strategy for Small-Cell Lung Cancer. Clin. Cancer Res. 2022, 28, 1966–1978. [Google Scholar] [CrossRef]
- Xu, W.; Lukkarila, J.L.; da Silva, S.R.; Paiva, S.-L.; Gunning, P.T.; Schimmer, A.D. Targeting the Ubiquitin E1 as a Novel Anti-Cancer Strategy. Curr. Pharm. Des. 2013, 19, 3201–3209. [Google Scholar] [CrossRef]
- Gan, Y.; Ye, F.; He, X.-X. The role of YWHAZ in cancer: A maze of opportunities and challenges. J. Cancer 2020, 11, 2252–2264. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-C.; Li, C.-F.; Chen, I.H.; Lai, M.-T.; Lin, Z.-J.; Korla, P.K.; Chai, C.-Y.; Ko, G.; Chen, C.-M.; Hwang, T.; et al. YWHAZ amplification/overexpression defines aggressive bladder cancer and contributes to chemo-/radio-resistance by suppressing caspase-mediated apoptosis. J. Pathol. 2019, 248, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Park, J.; Han, D.; Moon, J.H.; Ryu, H.S.; Kim, Y. Identification of TUBB2A by quantitative proteomic analysis as a novel biomarker for the prediction of distant metastatic breast cancer. Clin. Proteom. 2020, 17, 16. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, K.; Yin, X.; Li, H.; Li, Y.; Zhang, Q.; Wang, H.; Qiu, Y. Lower Expression of Gelsolin in Colon Cancer and Its Diagnostic Value in Colon Cancer Patients. J. Cancer 2019, 10, 1288–1296. [Google Scholar] [CrossRef]
- Chaponnier, C.; Gabbiani, G. Gelsolin modulation in epithelial and stromal cells of mammary carcinoma. Am. J. Pathol. 1989, 134, 597–603. [Google Scholar] [PubMed]
- Winston, J.S.; Asch, H.L.; Zhang, P.J.; Edge, S.B.; Hyland, A.; Asch, B.B. Downregulation of Gelsolin Correlates with the Progression to Breast Carcinoma. Breast Cancer Res. Treat. 2001, 65, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Shirkoohi, R.; Nakagawa, K.; Qiao, H.; Fujita, H.; Okada, F.; Hamada, J.-i.; Kuzumaki, S.; Takimoto, M.; Kuzumaki, N. siRNA gelsolin knockdown induces epithelial-mesenchymal transition with a cadherin switch in human mammary epithelial cells. Int. J. Cancer 2006, 118, 1680–1691. [Google Scholar] [CrossRef]
- Bianchi-Smiraglia, A.; Paesante, S.; Bakin, A.V. Integrin β5 contributes to the tumorigenic potential of breast cancer cells through the Src-FAK and MEK-ERK signaling pathways. Oncogene 2013, 32, 3049–3058. [Google Scholar] [CrossRef]
- Zhang, L.-y.; Guo, Q.; Guan, G.-f.; Cheng, W.; Cheng, P.; Wu, A.-H. Integrin Beta 5 Is a Prognostic Biomarker and Potential Therapeutic Target in Glioblastoma. Front. Oncol. 2019, 9, 904. [Google Scholar] [CrossRef]
- Shi, W.; He, J.; Huang, Y.; Zeng, Z.; Feng, Z.; Xu, H.; Nie, Y. Integrin β5 enhances the malignancy of human colorectal cancer by increasing the TGF-β signaling. Anti-Cancer Drugs 2021, 32, 717–726. [Google Scholar] [CrossRef]
- Konda, J.D.; Olivero, M.; Musiani, D.; Lamba, S.; Di Renzo, M.F. Heat-shock protein 27 (HSP27, HSPB1) is synthetic lethal to cells with oncogenic activation of MET, EGFR and BRAF. Mol. Oncol. 2017, 11, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Cayado-Gutiérrez, N.; Moncalero, V.L.; Rosales, E.M.; Berón, W.; Salvatierra, E.E.; Alvarez-Olmedo, D.; Radrizzani, M.; Ciocca, D.R. Downregulation of Hsp27 (HSPB1) in MCF-7 human breast cancer cells induces upregulation of PTEN. Cell Stress Chaperones 2013, 18, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, X.; Wang, H.; Wang, Y.; Chen, P.; Wang, L. Heat Shock Protein 27 Enhances SUMOylation of Heat Shock Protein B8 to Accelerate the Progression of Breast Cancer. Am. J. Pathol. 2020, 190, 2464–2477. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ou, Z.; Xie, M.; Kang, R.; Fan, Y.; Niu, X.; Wang, H.; Cao, L.; Tang, D. HSPB1 as a novel regulator of ferroptotic cancer cell death. Oncogene 2015, 34, 5617–5625. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, E.; Miki, Y.; Suzuki, T.; Hirakawa, H.; Ishida, T.; Sasano, H. Heterogeneous Nuclear Ribonucleoprotein K Is Involved in the Estrogen-Signaling Pathway in Breast Cancer. Int. J. Mol. Sci. 2021, 22, 2581. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Iwabuchi, E.; Takagi, K.; Suzuki, T.; Sasano, H.; Yaegashi, N.; Ito, K. Co-expression of nuclear heterogeneous nuclear ribonucleic protein K and estrogen receptor α in endometrial cancer. Pathol. Res. Pract. 2022, 231, 153795. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, H.; Wan, J.; Ma, Q.; Qi, Y.; Gu, Z. The hnRNPK/A1/R/U Complex Regulates Gene Transcription and Translation and is a Favorable Prognostic Biomarker for Human Colorectal Adenocarcinoma. Front. Oncol. 2022, 12, 845931. [Google Scholar] [CrossRef]
- Gallardo, M.; Hornbaker, M.J.; Zhang, X.; Hu, P.; Bueso-Ramos, C.; Post, S.M. Aberrant hnRNP K expression: All roads lead to cancer. Cell Cycle 2016, 15, 1552–1557. [Google Scholar] [CrossRef]
- Peng, W.-z.; Liu, J.-x.; Li, C.-f.; Ma, R.; Jie, J.-Z. hnRNPK promotes gastric tumorigenesis through regulating CD44E alternative splicing. Cancer Cell Int. 2019, 19, 335. [Google Scholar] [CrossRef]
- Li, M.; Zhang, W.; Yang, X.; Liu, H.; Cao, L.; Li, W.; Wang, L.; Zhang, G.; Gao, R. Downregulation of HNRNPK in human cancer cells inhibits lung metastasis. Anim. Models Exp. Med. 2019, 2, 291–296. [Google Scholar] [CrossRef]
- Chen, C.; Lei, J.; Zheng, Q.; Tan, S.; Ding, K.; Yu, C. Poly(rC) binding protein 2 (PCBP2) promotes the viability of human gastric cancer cells by regulating CDK2. FEBS Open Bio 2018, 8, 764–773. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Qin, H.; Zheng, L. Research progress on RNA−binding proteins in breast cancer. Front. Oncol. 2022, 12, 974523. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Guo, Q.; Wang, H.; Yuan, X.; Wang, B.; Lobie, P.E.; Zhu, T.; Tan, S.; Wu, Z. PCBP2 Posttranscriptional Modifications Induce Breast Cancer Progression via Upregulation of UFD1 and NT5E. Mol. Cancer Res. 2021, 19, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Zhang, X.-N.; Xu, C.-Z.; Zhou, D.-H.; Chen, J.; Liu, Z.-X.; Sun, Y.; Huang, W.; Qu, L.-S. MCCC2 promotes HCC development by supporting leucine oncogenic function. Cancer Cell Int. 2021, 21, 22. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yuan, Z.; Song, C. Methylcrotonoyl-CoA carboxylase 2 overexpression predicts an unfavorable prognosis and promotes cell proliferation in breast cancer. Biomark. Med. 2019, 13, 427–436. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Mao, Y.-H.; Huang, W.-t.; Li, M.-z.; Zhang, H.; Qing, Y.; Lu, S.; Xiao, H.; Li, K. Methylcrotonoyl-CoA Carboxylase 2 Promotes Proliferation, Migration and Invasion and Inhibits Apoptosis of Prostate Cancer Cells through Regulating GLUD1-P38 MAPK Signaling Pathway. OncoTargets Ther. 2020, 13, 7317–7327. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Feng, H.; Lee, D. MCCC2 overexpression predicts poorer prognosis and promotes cell proliferation in colorectal cancer. Exp. Mol. Pathol. 2020, 115, 104428. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.M.; Gu, F.; Ambati, C.R.; Rasaily, U.; Ramirez-Pena, E.; Joseph, R.; Manikkam, M.; San Martin, R.; Charles, C.; Pan, Y.; et al. UDP-glucose 6-dehydrogenase regulates hyaluronic acid production and promotes breast cancer progression. Oncogene 2020, 39, 3089–3101. [Google Scholar] [CrossRef]
- Lin, L.-H.; Chou, H.-C.; Chang, S.-J.; Liao, E.-C.; Tsai, Y.-T.; Wei, Y.-S.; Chen, H.-Y.; Lin, M.-W.; Wang, Y.-S.; Chien, Y.-A.; et al. Targeting UDP-glucose dehydrogenase inhibits ovarian cancer growth and metastasis. J. Cell. Mol. Med. 2020, 24, 11883–11902. [Google Scholar] [CrossRef]
- Oyinlade, O.; Wei, S.; Lal, B.; Laterra, J.; Zhu, H.; Goodwin, C.R.; Wang, S.; Ma, D.; Wan, J.; Xia, S. Targeting UDP-α-D-glucose 6-dehydrogenase inhibits glioblastoma growth and migration. Oncogene 2018, 37, 2615–2629. [Google Scholar] [CrossRef]
- Saha, S.; Yao, S.; Elakad, O.; Lois, A.-M.; Henric-Petri, H.; Büntzel, J.; Hinterthaner, M.; Danner, B.; Ströbel, P.; Emmert, A.; et al. UDP-glucose 6-dehydrogenase expression as a predictor of survival in patients with pulmonary adenocarcinoma. Int. J. Surg. Oncol. 2020, 5, e85. [Google Scholar] [CrossRef]
- Jin, X.; Wang, D.; Lei, M.; Guo, Y.; Cui, Y.; Chen, F.; Sun, W.; Chen, X. TPI1 activates the PI3K/AKT/mTOR signaling pathway to induce breast cancer progression by stabilizing CDCA5. J. Transl. Med. 2022, 20, 191. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Sun, S.-J.; Ai, Y.-J.; Feng, X.; Zheng, Y.-M.; Gao, Y.; Zhang, J.-Y.; Zhang, L.; Sun, Y.-P.; Xiong, Y.; et al. Elevated nuclear localization of glycolytic enzyme TPI1 promotes lung adenocarcinoma and enhances chemoresistance. Cell Death Dis. 2022, 13, 205. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xu, Y.; Zhou, Y.; Lang, R.; Shi, Z.; Zhao, J.; Meng, Y.; Bao, L. Integrated Analyses Reveal the Multi-Omics and Prognostic Characteristics of ATP5B in Breast Cancer. Front. Genet. 2021, 12, 652474. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Li, Y.; Li, W.; Kang, W.; Tang, R.; Wu, W.; Xing, Z.; Zhou, L. The function of Cav-1 in MDA-MB-231 breast cancer cell migration and invasion induced by ectopic ATP5B. Med. Oncol. 2021, 38, 73. [Google Scholar] [CrossRef]
- Wang, X.; Chang, X.; He, C.; Fan, Z.; Yu, Z.; Yu, B.; Wu, X.; Hou, J.; Li, J.; Su, L.; et al. ATP5B promotes the metastasis and growth of gastric cancer by activating the FAK/AKT/MMP2 pathway. FASEB J. 2021, 35, e20649. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-H.; Wu, C.-H.; Tsai, C.-C.; Chen, T.-Y.; Tsai, K.-J.; Hung, C.-M.; Hsu, C.-Y.; Wu, C.-W.; Hsieh, T.-H. Effects of Luteolin on Human Breast Cancer Using Gene Expression Array: Inferring Novel Genes. Curr. Issues Mol. Biol. 2022, 44, 2107–2121. [Google Scholar] [CrossRef]
- Anderson, N.M.; Qin, X.; Finan, J.M.; Lam, A.; Athoe, J.; Missiaen, R.; Skuli, N.; Kennedy, A.; Saini, A.S.; Tao, T.; et al. Metabolic Enzyme DLST Promotes Tumor Aggression and Reveals a Vulnerability to OXPHOS Inhibition in High-Risk Neuroblastoma. Cancer Res. 2021, 81, 4417–4430. [Google Scholar] [CrossRef]
- Shen, N.; Korm, S.; Karantanos, T.; Li, D.; Zhang, X.; Ritou, E.; Xu, H.; Lam, A.; English, J.; Zong, W.-X.; et al. DLST-dependence dictates metabolic heterogeneity in TCA-cycle usage among triple-negative breast cancer. Commun. Biol. 2021, 4, 1289. [Google Scholar] [CrossRef]
- Wu, T.; Li, Y.; Liu, B.; Zhang, S.; Wu, L.; Zhu, X.; Chen, Q. Expression of Ferritin Light Chain (FTL) Is Elevated in Glioblastoma, and FTL Silencing Inhibits Glioblastoma Cell Proliferation via the GADD45/JNK Pathway. PLoS ONE 2016, 11, e0149361. [Google Scholar] [CrossRef]
- Liu, J.; Gao, L.; Zhan, N.; Xu, P.; Yang, J.a.; Yuan, F.e.; Xu, Y.; Cai, Q.; Geng, R.; Chen, Q. Hypoxia induced ferritin light chain (FTL) promoted epithelia mesenchymal transition and chemoresistance of glioma. J. Exp. Clin. Cancer Res. 2020, 39, 137. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, J.; Chen, H.; Zhang, Y.; Shi, H.; Huang, L.; Tao, J.; Shen, R.; Wang, T. Ferritin Light Chain (FTL) competes with long noncoding RNA Linc00467 for miR-133b binding site to regulate chemoresistance and metastasis of colorectal cancer. Carcinogenesis 2019, 41, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Tamura, R.E.; de Vasconcellos, J.F.; Sarkar, D.; Libermann, T.A.; Fisher, P.B.; Zerbini, L.F. GADD45 proteins: Central players in tumorigenesis. Curr. Mol. Med. 2012, 12, 634–651. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.S.; Patil, S.; Majumdar, B.; Oswal, R.G. Comparison of special stains for keratin with routine hematoxylin and eosin stain. J. Int. Oral Health JIOH 2015, 7, 1–5. [Google Scholar] [PubMed]
- Wang, J.-M.; Jiang, J.-Y.; Zhang, D.-L.; Du, X.; Wu, T.; Du, Z.-X. HYOU1 facilitates proliferation, invasion and glycolysis of papillary thyroid cancer via stabilizing LDHB mRNA. J. Cell. Mol. Med. 2021, 25, 4814–4825. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Gong, L.; Liu, X.; Zhu, T.; Zhou, W.; Kong, L.; Luo, J. Identification of peroxiredoxin 6 as a direct target of withangulatin A by quantitative chemical proteomics in non-small cell lung cancer. Redox Biol. 2021, 46, 102130. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Lu, E.; Pan, C.; Xu, Y.; Zhu, X. Overexpression and biological function of PRDX6 in human cervical cancer. J. Cancer 2020, 11, 2390–2400. [Google Scholar] [CrossRef]
- Huang, W.-S.; Huang, C.-Y.; Hsieh, M.-C.; Kuo, Y.-H.; Tung, S.-Y.; Shen, C.-H.; Hsieh, Y.-Y.; Teng, C.-C.; Lee, K.; Lee, K.-F.; et al. Expression of PRDX6 Correlates with Migration and Invasiveness of Colorectal Cancer Cells. Cell. Physiol. Biochem. 2018, 51, 2616–2630. [Google Scholar] [CrossRef]
- Chang, X.-Z.; Li, D.-Q.; Hou, Y.-F.; Wu, J.; Lu, J.-S.; Di, G.-H.; Jin, W.; Ou, Z.-L.; Shen, Z.-Z.; Shao, Z.-M. Identification of the functional role of peroxiredoxin 6 in the progression of breast cancer. Breast Cancer Res. 2007, 9, R76. [Google Scholar] [CrossRef]
- Kasai, Y.; Toriguchi, K.; Hatano, E.; Nishi, K.; Ohno, M.; Yoh, T.; Fukuyama, K.; Nishio, T.; Okuno, M.; Iwaisako, K.; et al. Nardilysin promotes hepatocellular carcinoma through activation of signal transducer and activator of transcription 3. Cancer Sci. 2017, 108, 910–917. [Google Scholar] [CrossRef]
- Kanda, K.; Sakamoto, J.; Matsumoto, Y.; Ikuta, K.; Goto, N.; Morita, Y.; Ohno, M.; Nishi, K.; Eto, K.; Kimura, Y.; et al. Nardilysin controls intestinal tumorigenesis through HDAC1/p53–dependent transcriptional regulation. JCI Insight 2018, 3, e91316. [Google Scholar] [CrossRef] [PubMed]
- Hospital, V.; Prat, A. Nardilysin, A Basic Residues Specific Metallopeptidase That Mediates Cell Migration and Proliferation. Protein Pept. Lett. 2004, 11, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.-s.; Geng, H.; Li, L.; Ying, J.; Ma, C.; Wang, Y.; Poon, F.F.; Wang, X.; Ying, Y.; Yeo, W.; et al. The epigenetic modifier PRDM5 functions as a tumor suppressor through modulating WNT/β-catenin signaling and is frequently silenced in multiple tumors. PLoS ONE 2011, 6, e27346. [Google Scholar] [CrossRef]
- Guo, J.; Yang, Q.; Wei, S.; Shao, J.; Zhao, T.; Guo, L.; Liu, J.; Chen, J.; Wang, G. Low expression of PRDM5 predicts poor prognosis of esophageal squamous cell carcinoma. BMC Cancer 2022, 22, 745. [Google Scholar] [CrossRef] [PubMed]
- Teng, J.-J.; Zhao, W.-J.; Zhang, X.-L.; Zhao, D.-K.; Qiu, X.-Y.; Chen, X.-D.; Yang, L. Downregulation of promoter methylation gene PRDM5 contributes to the development of tumor proliferation and predicts poor prognosis in gastric cancer. J. Cancer 2021, 12, 6921–6930. [Google Scholar] [CrossRef] [PubMed]
- Casamassimi, A.; Rienzo, M.; Di Zazzo, E.; Sorrentino, A.; Fiore, D.; Proto, M.C.; Moncharmont, B.; Gazzerro, P.; Bifulco, M.; Abbondanza, C. Multifaceted Role of PRDM Proteins in Human Cancer. Int. J. Mol. Sci. 2020, 21, 2648. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Song, J.; Chen, W.; Yuan, D.; Wang, W.; Chen, X.; Liu, H.; Su, H.; Zhu, J. Expression and Role of Dickkopf-1 (Dkk1) in Tumors: From the Cells to the Patients. Cancer Manag. Res. 2021, 13, 659–675. [Google Scholar] [CrossRef]
- Qi, L.; Sun, B.; Liu, Z.; Li, H.; Gao, J.; Leng, X. Dickkopf-1 inhibits epithelial-mesenchymal transition of colon cancer cells and contributes to colon cancer suppression. Cancer Sci. 2012, 103, 828–835. [Google Scholar] [CrossRef]
- Fezza, M.; Moussa, M.; Aoun, R.; Haber, R.; Hilal, G. DKK1 promotes hepatocellular carcinoma inflammation, migration and invasion: Implication of TGF-β1. PLoS ONE 2019, 14, e0223252. [Google Scholar] [CrossRef]
- Teng, Y.; Qin, H.; Bahassan, A.; Bendzunas, N.G.; Kennedy, E.J.; Cowell, J.K. The WASF3-NCKAP1-CYFIP1 Complex Is Essential for Breast Cancer Metastasis. Cancer Res. 2016, 76, 5133–5142. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, M.Q.; Cheney, R.; Sharma, S.; Cowell, J.K. Inactivation of the WASF3 gene in prostate cancer cells leads to suppression of tumorigenicity and metastases. Br. J. Cancer 2010, 103, 1066–1075. [Google Scholar] [CrossRef]
- Zhu, Y.; Lin, H.; Li, Z.; Wang, M.; Luo, J. Modulation of expression of ribosomal protein L7a (rpL7a) by ethanol in human breast cancer cells. Breast Cancer Res. Treat. 2001, 69, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Li, X.; Nian, W.; Wang, J.; Wang, X.; Sun, L.; Zhu, Y.; Tong, Z. Ribosome Proteins Represented by RPL27A Mark the Development and Metastasis of Triple-Negative Breast Cancer in Mouse and Human. Front. Cell Dev. Biol. 2021, 9, 716730. [Google Scholar] [CrossRef] [PubMed]
- Vaarala, M.; Porvari, K.; Kyllönen, A.; Mustonen, M.; Lukkarinen, O.; Vihko, P. Several genes encoding ribosomal proteins are over-expressed in prostate-cancer cell lines: Confirmation of L7a and L37 over-expression in prostate-cancer tissue samples. Int. J. Cancer 1998, 78, 27–32. [Google Scholar] [CrossRef]
- Maruyama, Y.; Miyazaki, T.; Ikeda, K.; Okumura, T.; Sato, W.; Horie-Inoue, K.; Okamoto, K.; Takeda, S.; Inoue, S. Short hairpin RNA library-based functional screening identified ribosomal protein L31 that modulates prostate cancer cell growth via p53 pathway. PLoS ONE 2014, 9, e108743. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, W.; Nasr, Z. Deregulation of ribosomal proteins in human cancers. Biosci. Rep. 2021, 41, BSR20211577. [Google Scholar] [CrossRef] [PubMed]
- Alam, E.; Maaliki, L.; Nasr, Z. Ribosomal protein S3 selectively affects colon cancer growth by modulating the levels of p53 and lactate dehydrogenase. Mol. Biol. Rep. 2020, 47, 6083–6090. [Google Scholar] [CrossRef] [PubMed]
- Ono, H.; Iizumi, Y.; Goi, W.; Sowa, Y.; Taguchi, T.; Sakai, T. Ribosomal protein S3 regulates XIAP expression independently of the NF-κB pathway in breast cancer cells. Oncol. Rep. 2017, 38, 3205–3210. [Google Scholar] [CrossRef]
- Kopecki, Z.; Ludwig, R.J.; Cowin, A.J. Cytoskeletal Regulation of Inflammation and Its Impact on Skin Blistering Disease Epidermolysis Bullosa Acquisita. Int. J. Mol. Sci. 2016, 17, 1116. [Google Scholar] [CrossRef]
- Mukherjee, R.; Yun, J.W. Long chain acyl CoA synthetase 1 and gelsolin are oppositely regulated in adipogenesis and lipogenesis. Biochem. Biophys. Res. Commun. 2012, 420, 588–593. [Google Scholar] [CrossRef]
- Gyamfi, J.; Eom, M.; Koo, J.-S.; Choi, J. Multifaceted Roles of Interleukin-6 in Adipocyte-Breast Cancer Cell Interaction. Transl. Oncol. 2018, 11, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ren, Y.; Dai, Z.-J.; Wu, C.-J.; Ji, Y.-H.; Xu, J. IL-6, IL-8 and TNF-α levels correlate with disease stage in breast cancer patients. Adv. Clin. Exp. Med. 2017, 26, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Casneuf, T.; Axel, A.E.; King, P.; Alvarez, J.D.; Werbeck, J.L.; Verhulst, T.; Verstraeten, K.; Hall, B.M.; Sasser, A.K. Interleukin-6 is a potential therapeutic target in interleukin-6 dependent, estrogen receptor-α-positive breast cancer. Breast Cancer 2016, 8, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Mercogliano, M.F.; Bruni, S.; Elizalde, P.V.; Schillaci, R. Tumor Necrosis Factor α Blockade: An Opportunity to Tackle Breast Cancer. Front. Oncol. 2020, 10, 584. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Chen, X.; Shu, Y. Gene expression of the invasive phenotype of TNF-α-treated MCF-7 cells. Biomed. Pharmacother. 2009, 63, 421–428. [Google Scholar] [CrossRef]
- Martinez-Ruiz, G.; Maldonado, V.; Ceballos-Cancino, G.; Grajeda, J.P.R.; Melendez-Zajgla, J. Role of Smac/DIABLO in cancer progression. J. Exp. Clin. Cancer Res. 2008, 27, 48. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, M.; Lizárraga, F.; Vazquez-Santillan, K.; Hidalgo-Miranda, A.; Piña-Sánchez, P.; Torres, J.; García-Ramírez, R.; Lagunas, V.; Meléndez-Zajgla, J.; Ceballos-Cancino, G. Coexpression of Smac/DIABLO and Estrogen Receptor in breast cancer. Cancer Biomark. 2021, 30, 429–446. [Google Scholar] [CrossRef]
- Comşa, S.; Anca Maria, C.; Raica, M. The Story of MCF-7 Breast Cancer Cell Line: 40 years of Experience in Research. Anticancer Res. 2015, 35, 3147–3154. [Google Scholar]
- Tran, B.; Bédard, P. Luminal-B breast cancer and novel therapeutic targets. Breast Cancer Res. BCR 2011, 13, 221. [Google Scholar] [CrossRef]
- Pang, H.; Rowan, B.G.; Al-Dhaheri, M.; Faber, L.E. Epidermal growth factor suppresses induction by progestin of the adhesion protein desmoplakin in T47D breast cancer cells. Breast Cancer Res. 2004, 6, R239. [Google Scholar] [CrossRef]
- Aktary, Z.; Alaee, M.; Pasdar, M. Beyond cell-cell adhesion: Plakoglobin and the regulation of tumorigenesis and metastasis. Oncotarget 2017, 8, 32270–32291. [Google Scholar] [CrossRef]
- Leick, K.M.; Obeid, J.M.; Bekiranov, S.; Slingluff, C.L. Systems analysis of barrier molecule and ARNT-related gene expression regulation in melanoma. Oncoimmunology 2019, 8, e1665978. [Google Scholar] [CrossRef]
- Aslebagh, R.; Channaveerappa, D.; Arcaro, K.F.; Darie, C.C. Comparative two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) of human milk to identify dysregulated proteins in breast cancer. Electrophoresis 2018, 39, 1723–1734. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Pathways | NES | FDR q-Val | |
|---|---|---|---|
| Upregulated | Hypoxia | 1.49 | 0.535 |
| EMT | 1.08 | 1 | |
| Downregulated | Mitotic_spindle | −1.17 | 1 |
| UPR | −1.09 | 1 |
| Gene Name | Gene Description | Role in Biological Processes | Expression in Malignancies and Putative Neoplastic Effects | Pathways | Neoplastic Behavior | |
|---|---|---|---|---|---|---|
| Upregulated proteins | ||||||
| RBBP4/RbAp48 | retinoblastoma-binding protein 4/histone-binding protein RBBP4 | tumor-specific, histone/DNA-binding protein involved in chromatin remodeling, DNA replication, nucleosome assembly, histone modification, transcription regulation, cell cycle and proliferation, cell differentiation and signaling, cell motility, protein-containing complex assembly [17,18] | overexpressed in embryonal and glial brain cancers [18], TNBC tissues and cell lines [19], CC cell lines [20] | PT | EMT [19], nuclear accumulation of β-catenin, activating Wnt/β-catenin signaling [20] | cell proliferation, invasion, migration, anti-apoptosis [17] |
| SET/TAF-I β isoform CRA_c/INHAT | SET nuclear proto-oncogene/template-activating factor-I β/inhibitor of histone acetyltransferase | multitasking histone/DNA-binding protein involved in DNA replication, chromatin organization, transcription regulation, nucleosome assembly, histone chaperoning (uniprot.org), protein-containing complex assembly, cell cycle, apoptosis, DNA repair [21] | overexpressed in 50–60% of BC [22], pancreatic cancer, leukemias, lymphomas, nephroblastoma, hepatoma, choriocarcinoma [23] | PT | EMT, Rac1/JNK/c-Jun, MEK/ERK [23] | N-cadherin activation, cell growth, proliferation, migration, invasion, colony formation [23] |
| ABRACL/HSCP280/ABRA C-terminal-like | actin-binding Rho-activating C-terminal-like | actin cytoskeleton remodeling, cell motility [24], cell cycle [25] | overexpressed in BC cells [25], endometrial cancer [26], esophageal carcinoma [27], GC [24], colon cancer [28] | PT | EMT [25], proteasome degradation, mitochondrial pathway [24] | proliferation, invasion, migration [25,28] |
| NCKAP1/NAP1 | noncatalytic region of tyrosine kinase (Nck)-associated protein 1 | cytoskeleton/actin dynamics regulator, part of WAVE complex that regulates lamellipodia/invadopodia formation and cell mobility [29], cell adhesion and migration [30] | overexpressed in CRC [31], NSCLC [32], BC [30] | PT | EMT activation [31] by HSP90-mediated invasion and metastasis by provoking MMP9 and MMP14 activation [32] | promotes primary tumor growth and progression to metastatic disease by directing the polarized interaction of BC cells with collagen fibrils; increases cell migration, invasion [31], and MMP9 secretion [32] |
| JUP/γ-catenin | junction plakoglobin | member of Armadillo family, homolog of β-catenin, common junctional plaque protein involved in cell–cell junction and signaling [33] | overexpressed in many cancers [34]: IMPC tissue and cells [35], PCa [36], AML [37], ESCC [38] | tumor suppressor (AT) in ESCC [38]; when overexpressed and located in nucleus, PT/ oncoprotein [35] | AJ; activation of PI3K/Akt/Bcl-2 and cluster cells survival [35], JUP/EGFR/AKT/GSK3β involved in tumor metastasis via inducing nuclear β-catenin translocation to upregulation of MMP7 expression in GC, promoting EMT and increasing invasion potential [39] | downregulation of CDH1, overexpression of VIM, increases cell mobility and migration, enhances pAKT and pERK, increases in PTTG and c-Myc protein levels, chromosomal instability and uncontrolled proliferation [34], tumor cluster regulator, metastasis promoter, and apoptosis downregulation [35] |
| DSPI//DP isoform I | desmoplakin I | tumor-suppressor; component of desmosomal plaques that interacts with JUPl; DSP overexpression enhances JUP expression [40] | inactivated in NSCLC [40], overexpressed in melanoma cell line [41] | AT or controversial, depending on the localization; in nucleus, it is involved in telomere maintenance [42] | when overexpressed, reduced expression of Wnt/β-catenin [40] | depletion induces DNA damage response and cell apoptosis, blocking the normal function of centrosomes; overexpression may have opposite effects |
| APC/3NMW | chain A, Armadillo repeats domain of adenomatous polyposis coli (APC) | multi-functional tumor suppressor; regulates cell–cell adhesion, cell polarization, and migration [43]; cell proliferation and differentiation [44]; organization of actin and microtubule networks, spindle formation, and chromosome segregation [45] | mutated in colon cancer and liver cancer [45] | AT | negative regulator of canonical Wnt signaling [43,45] | deregulation involved in carcinogenesis |
| DIABLO/ SMAC | direct IAP-binding protein with low pI/second mitochondrial-derived activator of caspase | mitochondrial protein that promotes caspase-dependent apoptosis by inhibition of IAP family members [46] | overexpressed in GC, CRC, and ovarian cancer; downregulated in PCa, lung, and soft tissue cancers; more downregulated in BC tissues than in control samples; expression decreases with BC progression [47] | AT or PT | APOPTOSIS, phospholipid biosynthetic pathways [48] | pro- and non-apoptotic functions; SMAC DEPs are associated with lipids, lipid-signaling molecules, metabolism, DNA/RNA-associated proteins, transport and intracellular trafficking, cellular signaling, immunity, TME reorganization [49] |
| HINT2 | histidine triad nucleotide-binding protein 2, mitochondrial precursor | tumor suppressor in mitochondrial matrix; sensitizes cells to apoptosis [50]; positively regulates lipid and glucose metabolism and mitochondrial respiration [51] | downregulated in HCC [50], CRC [52], pancreatic cancer [53], BC [54] | AT | overexpression induces an anti-EMT gene expression profile in cancer cells [52] | overexpression inhibits cell migration, metastasis, and invasion [52]; induces cell apoptosis; decreases mitochondrial membrane potential; promotes intracellular ROS production; elevates mitochondrial Ca2+ levels [53] |
| Downregulated proteins | ||||||
| UBA1 | ubiquitin-like modifier-activating enzyme 1 | initiation of ubiquitination cascade [55], regulator of proteostasis [56] | involved in development of HCC [57], TNBC [58], SCLC [59] | AT | ubiquitin-conjugation pathway (UCP) [59]; ferroptosis regulator [57] | silencing inhibits cell proliferation, migration, and invasion; upregulates proteolytic and DNA damage stress, the Fe2+ content in cells and ferroptosis; and suppresses NRF2 signaling pathway [57]; elicits UPR and induces cell death [60]; highly toxic for TNBC models; upregulation of ER stress; pro-apoptotic [58] |
| YWHAZ/14-3-3ζ | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta | central hub protein involved in many signal transduction pathways [61] | oncogene overexpressed in multiple cancers: HCC, CRC, LUAD, BC [61], urothelial carcinomas [62] | AT | UPR | knockdown decreases cell growth, proliferation, and invasion; enhances apoptosis and tamoxifen-induced inhibition of cell viability [61] |
| TUBB2A | tubulin beta-2A chain | associated with cellular proliferation, movement, and adhesion; involved in mitotic cell cycle, cytoskeleton organization, and cell migration; novel biomarker for the prediction of distant metastatic BC [63] | overexpressed in invasive BC cell lines [63] | AT | UPR TNFA_SIGNALING_VIA_NFKB | decreased invasiveness and cell migration [63] |
| GSN | gelsolin | actin-binding protein/actin regulator | dysregulated in various cancers; downregulated in CC tissues, HCC, GC, cervical cancer, ovarian cancer [64], BC [65,66] | PT | knockdown leads to EMT in mammary epithelial cells [67] | cell motility [66]; controls CDH1 to N-cadherin conversion via Snail [67]; deficiency increases with progression from ADH to DCIS to IBC [66] |
| ITGB5 | integrin subunit beta 5 | integral transmembrane protein involved in cell adhesion | oncogenic factor overexpressed in aggressive tumors: primary and metastatic TNBC cells [68]; GBM [69], CRC [70] | AT | overexpressed, mediates TGF-β/SMAD signaling and facilitates EMT in cancer cells [69,70]; deficiency leads to inhibition of Src-FAK and MEK-ERK signaling [68] | depletion reduces tumor growth, survival, proliferation, migration, invasion, and angiogenesis [68,69,70] |
| HSPB1/ HSP27 | heat shock protein beta-1 | stress-inducible chaperone | overexpressed in many cancers [71], BC tissues [72], and MCF7 cells [73] | AT | APOPTOSIS; downregulation in MCF7 cells induces upregulation of PTEN (tumor suppressor) that negatively regulates PI3K/AKT signaling [72] | negative regulator of ferroptotic cancer cell death [74] and apoptotic death [71]; HSP27 downregulation was correlated with increased PTEN expression [72] |
| HNRNPK | heterogeneous nuclear ribonucleoprotein K | multifunctional RNA-binding protein (RBP), contributes to chromatin remodeling, transcription, splicing, and translation [75] | tumor suppressor or oncogene [76] overexpressed in many cancers: melanoma, PCa, BC (especially in ER+ and/or PR+ BC, including MCF7 cells [75]), lung, CRC [77], HCC, ESCC [78], GC [79] | AT | ER-mediated signaling pathway [75]; high expression was accompanied by high levels of c-Myc in BC [80] | downregulation inhibits lung metastasis [80] and ERα expression [75] |
| PCBP2 | poly(rC)-binding protein 2 isoform b | RNA-binding protein, contributes to transcriptional and translational regulation [81,82] | oncogene, promoter of GC [81], HCC, GBM, BC tissues, and cell lines [83] | AT | overexpression increases cholesterol synthesis and facilitates the stemness of BCSCs via activating PI3K/Akt signaling [82] | depletion decreases GC cells viability and proliferation [81]; inhibits cell proliferation, colony formation, migration, invasion, in vivo tumor growth, and metastasis in BC [83] |
| MCCC2 | methylcrotonyl-CoA carboxylase 2 | mitochondrial member of the biotin-dependent carboxylase superfamily | oncogene overexpressed in HCC [84], BC [85], PCa [86], CRC [87] | AT | downregulation of survival-dependent leucine metabolism [84] | knockdown expression reduces cell proliferation, migration, and invasion and glycolysis markers, glucose consumption, lactate secretion, and acetyl-CoA level [84]; promotes apoptosis [86] |
| UGDH isoform 2 | UDP-glucose 6-dehydrogenase | metabolic enzyme associated with mesenchymal-like gene expression [88] | upregulated in epithelial cancers, such as BC [88]; highly metastatic ovarian cancer cell lines [89]; GBM [90]; lung cancer [91] | AT | EMT inhibition, inactivation of ERK/MAPK, metabolic reprogramming, ECM remodeling [88] | knockdown decreases cell motility, invasion, GAGs synthesis, and cell migration [90]; tumor growth, HA production, colony formation [88]; induces cell cycle arrest in G0/G1 phase [89] |
| TPI1 | triosephosphate isomerase 1 | glycolytic enzyme involved in metabolic reprogramming | oncogenic function when translocated to cell nucleus induced by stress condition; overexpressed in multiple cancers: BC tissues and cell lines [92], LUAD [93] | AT | MTORC1_SIGNALING; GLYCOLYSIS; HYPOXIA; EMT [92] | knockdown reduces cell migration, colony formation and xenograft tumor growth [93] |
| ATP5F1B/ATP5B | adenosine triphosphate synthase F1 subunit beta, mitochondrial precursor | metabolic enzyme in inner mitochondrial membrane, produces ATP via OXPHOS, ectopically expressed on the surface of various cancer cells [94] | participant in carcinogenesis in several tumors, overexpressed in BC, especially in luminal and HER2+ subtypes [94]; plasma membrane of highly invasive cells, including MDA-MB-231 BC cells [95]; GC [96] | AT | OXPHOS; overexpression induces cancer progression via FAK/AKT/MMP2 pathway [96] | overexpression increases intracellular ATP in cancer cells, promoting migration, invasion [95], and proliferation [94]; inhibition suppresses cancer cell metastasis and growth [96] |
| DLST mitochondrial | dihydrolipoamide S-succinyltransferase | metabolic enzyme | oncogene highly expressed in BC, including MCF7 and MDA-MB-231 BC cell lines [97]; overexpressed in neuroblastoma [98]; TNBC [99] | AT | OXPHOS; ROS [99] | depletion impedes disease initiation and progression; impaires OXPHOS; suppresses growth and TCA-cycle; increases ROS levels; induces apoptosis; decreases burden and invasion [98,99] |
| FTL | ferritin light chain | iron metabolism regulator [100], hypoxia-responsive gene [101] | oncogene overexpressed in various malignant tumors [101]: GBM cells and serum [100], and CRC tissues and cell lines [102] | AT | interacts with PI3K/Akt, GADD45/JNK, TGF-β signaling, and cell cycle proteins; FTL knockdown decreases the expression of Wnt target genes, cyclin D1, and c-Myc [100]; represses EMT by regulation of Akt/GSK3β/β-catenin signaling [101] | knockdown results in inhibition of cancer cell growth and viability by activation of GADD45A expression that impairs cell proliferation (tumor suppressor [103]) [100]; reduces cell migration, invasion, and cancer cell survival rate and increases apoptosis [101] |
| HYOU1/GRP170/ORP150 | hypoxia upregulated 1 (HSP70 protein family) | chaperone with cytoprotective role, involved in protein folding in ER under stressful conditions [104,105] | upregulated in many cancers: BC, PTC, NPC, EOC, KS [105] | AT | silencing promotes OXPHOS and inhibits aerobic glycolysis [105] | silencing suppressed proliferation, migration, and invasion [105] |
| PRDX6 | peroxiredoxin 6 | antioxidant enzyme | overexpressed in various cancers [106]; overexpressed in cervical cancer [107]; CRC [108]; MDA-MB-231HM BC cell line [109] | AT | ROS | knockdown inhibits proliferation, migration, and invasion and stimulates apoptosis [107] |
| NRDC1 | nardilysin/N-arginine dibasic convertase 1 | nuclear, cytoplasmatic, or cell-surface associated metallo-/zinc-endopeptidase of the M16 family; transcriptional co-regulator [110]; cell-surface receptor for HB-EGF; epigenetic regulatory function [111] | promotion of BC, GC, ESCC, and HCC tissue and serum [110]; overexpressed in CRC [111] | AT | activates IL-6-Signal Transducer and Activator of Transcription 3 (STAT3) signaling [110]; regulates tumor development via HDAC/p53 pathway [111] | involved in cell migration and proliferation [112]; deficiency diminishes tumor size, suppresses carcinogenesis/proliferation [111], spheroid growth, and STAT3 phosphorylation [110] |
| PRDM5/ PFM2 | PRDI-BF1 and RIY domain containing 1 | zinc finger protein acts as an epigenetic modifier [113] | tumor suppressor, frequently silenced/downregulated/inactivated by methylation in multiple carcinoma lines: NPC, ESCC [114]; GC [115]; HCC, ovarian, cervical, BC [113,116] | PT | stress-responsive gene, epigenetic regulation of multiple oncogenes, antagonizes Wnt/β-catenin signaling, possible involvement in EMT process [113,114] | knockdown increases cell growth, proliferation, migration, invasion, tumor initiation, and progression [113,114,115]; opposite role in melanoma [116] |
| DKK1 | Dickkopf Wnt signaling pathway inhibitor 1 | secretory antagonist of β-catenin-dependent Wnt signaling pathway [117] | tumor suppressor in CC [118] or oncogene abnormally expressed in tumor cells, overexpressed in many cell lines: HCC [119]; lung, BC (incl. in serum), cervical cancers, glioma [117] | AT | WNT_BETA_CATENIN_SIGNALING; overexpression inhibits EMT in CC [118] | knockdown may inhibit migration, invasion, proliferation, cancer stem cell-like proprieties, tumor growth, and angiogenesis; enhances apoptosis and tumor regression [117] |
| NCKAP1/NAP1 | NCK-associated protein 1, isoform CRA_a | member of the Wiskott–Aldrich syndrome protein family member (WASF) regulatory complex (WRC); involved in actin cytoskeleton organization, lamellipodia formation, and cell motility and adhesion [32] | overexpressed in high-grade tumors: BC, PCa, colon cancer [120], NSCLC tissue [32] | AT | WASF3 stability, invasion potential [120]; HSP90-mediated invasion by MMP9 activation, VIM upregulation, and EMT in cancer cells [32] | silencing destabilizes WASF3 complex involved in actin cytoskeletal reorganization, cell movement, and invasion [121]; suppresses invasiveness and metastasis [120]; reduces MMP9 secretion [32] |
| RPL7A | 60S ribosomal protein L7a | ribosomal protein | overexpressed in BC [122], including TNBC, especially metastatic TNBC cells [123], PCa cell lines [124] | AT | activates TRK oncogene [122]; EIF2 signaling [123] | involved in cellular transformation, tumor growth, aggressiveness, and metastasis [122]; blocking RPL7A reduced cell migration and invasion [123] |
| RPL31 | 60S ribosomal protein L31 | ribosomal protein | overexpressed in PCa [125], CC [126] | AT | knockdown decreases degradation of tumor suppressor p53 and its targets [125] | knockdown enhances levels of p53 and p21, decreases cell growth and cell cycle [125] |
| RPS3 | 40S ribosomal protein S3 | ribosomal protein involved in ribosomal maturation and translation initiation, DNA damage repair, apoptosis, survival, transcription, and tumorigenesis [127] | oncogenic protein overexpressed in colon adenocarcinoma [127], BC [128] | AT | MYC_TARGETS_v1; ribosome signaling pathway, knockdown increases level of tumor suppressor p53 and induces G1 cell cycle arrest [127] | knockdown promotes ribosomal stress, which impairs ribosomal biogenesis; impedes cell proliferation, invasion, and migration; and increases apoptosis [127] |
| Biological Processes/ Pathways Involved in Carcinogenesis and Tumor Progression | EMT | Increase in Cell Growth | Cytoskeleton Organization | Inflammatory Response | Cell–cell Adhesion | Chromatin Remodelling | Chromosome/Genomic Instability | Transcription Regulation | Stimulation of Cell Proliferation | Cell Cycle | Increased Cell Invasion | Increased Cell Motility/Migration | Anti-Apoptosis | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DEPs | ||||||||||||||
| Pro-tumorigenic (PT) functions | ||||||||||||||
| RBBP4up | X | X | X | X | X | X | X | X | ||||||
| SETup | X | X | X | X | X | X | X | |||||||
| ABRACLup | X | X | X | X | X | X | ||||||||
| NCKAP1up | X | X | X | X | X | |||||||||
| GSNdown | X | X | X | X | ||||||||||
| PRDM5down | X | X | X | X | X | |||||||||
| Biological Processes/ Pathways Involved in Carcinogenesis and Tumor Progression | EMT Inhibition | Proteostasis Regulation | Inhibition of Cell/Tumor Growth | Cytoskeleton Organization | Cell–cell Adhesion | Chromatin Remodelling | Transcription Regulation | Inhibition of Cell Proliferation | Cell Cycle Arrest | Inhibition of Cell Invasion | Inhibition of Cell Motility/Migration | Pro-Apoptosis | Reduced Glycolysis | Increased ROS Production | UPR Increasing | Angiogenesis Inhibition | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DEPs | |||||||||||||||||
| Anti-tumorigenic functions (AT) | |||||||||||||||||
| APCup | X | X | X | ||||||||||||||
| HINT2up | X | X | X | X | X | ||||||||||||
| UBA1down | X | X | X | X | X | X | |||||||||||
| YWHAZdown | X | X | X | X | |||||||||||||
| TUBB2Adown | X | X | |||||||||||||||
| ITGB5down | X | X | X | X | X | X | X | X | |||||||||
| HSPB1/HSP27down | X | ||||||||||||||||
| HNRNPKdown | X | X | |||||||||||||||
| PCBP2down | X | X | X | X | X | ||||||||||||
| MCCC2down | X | X | X | X | X | ||||||||||||
| UGDHdown | X | X | X | X | X | ||||||||||||
| TPI1down | X | X | X | X | |||||||||||||
| ATP5F1Bdown | X | X | X | X | |||||||||||||
| DLSTdown | X | X | X | X | |||||||||||||
| FTLdown | X | X | X | X | X | X | X | ||||||||||
| HYOU1down | X | X | X | X | |||||||||||||
| PRDX6down | X | X | X | X | |||||||||||||
| NRDC1down | X | X | X | X | |||||||||||||
| DKK1down | X | X | X | X | X | X | |||||||||||
| NCKAP1down | X | X | X | ||||||||||||||
| RPL7Adown | X | X | X | ||||||||||||||
| RPL31down | X | X | X | X | X | X | |||||||||||
| RPS3down | X | X | X | X | X | X | |||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayathirtha, M.; Jayaweera, T.; Whitham, D.; Petre, B.A.; Neagu, A.-N.; Darie, C.C. Two-Dimensional Polyacrylamide Gel Electrophoresis Coupled with Nanoliquid Chromatography–Tandem Mass Spectrometry-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in the MCF7 Breast Cancer Cell Line Transfected for Jumping Translocation Breakpoint Protein Overexpression. Int. J. Mol. Sci. 2023, 24, 14714. https://doi.org/10.3390/ijms241914714
Jayathirtha M, Jayaweera T, Whitham D, Petre BA, Neagu A-N, Darie CC. Two-Dimensional Polyacrylamide Gel Electrophoresis Coupled with Nanoliquid Chromatography–Tandem Mass Spectrometry-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in the MCF7 Breast Cancer Cell Line Transfected for Jumping Translocation Breakpoint Protein Overexpression. International Journal of Molecular Sciences. 2023; 24(19):14714. https://doi.org/10.3390/ijms241914714
Chicago/Turabian StyleJayathirtha, Madhuri, Taniya Jayaweera, Danielle Whitham, Brîndușa Alina Petre, Anca-Narcisa Neagu, and Costel C. Darie. 2023. "Two-Dimensional Polyacrylamide Gel Electrophoresis Coupled with Nanoliquid Chromatography–Tandem Mass Spectrometry-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in the MCF7 Breast Cancer Cell Line Transfected for Jumping Translocation Breakpoint Protein Overexpression" International Journal of Molecular Sciences 24, no. 19: 14714. https://doi.org/10.3390/ijms241914714