
The Properties of Linezolid, Rifampicin, and Vancomycin, as Well as the Mechanism of Action of Pentamidine, Determine Their Synergy against Gram-Negative Bacteria
 , and
, and
Abstract
:1. Introduction
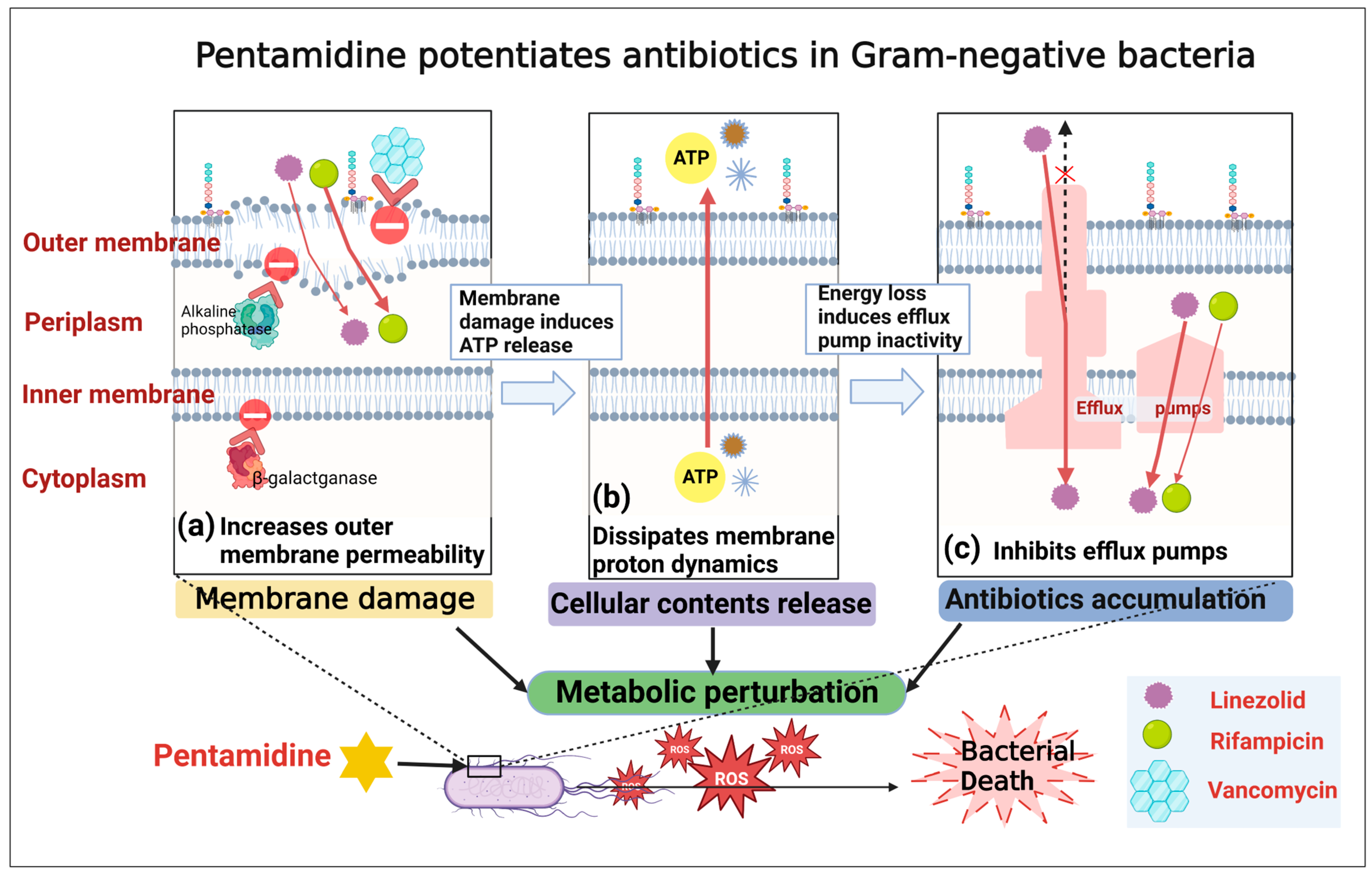
2. Results and Discussion
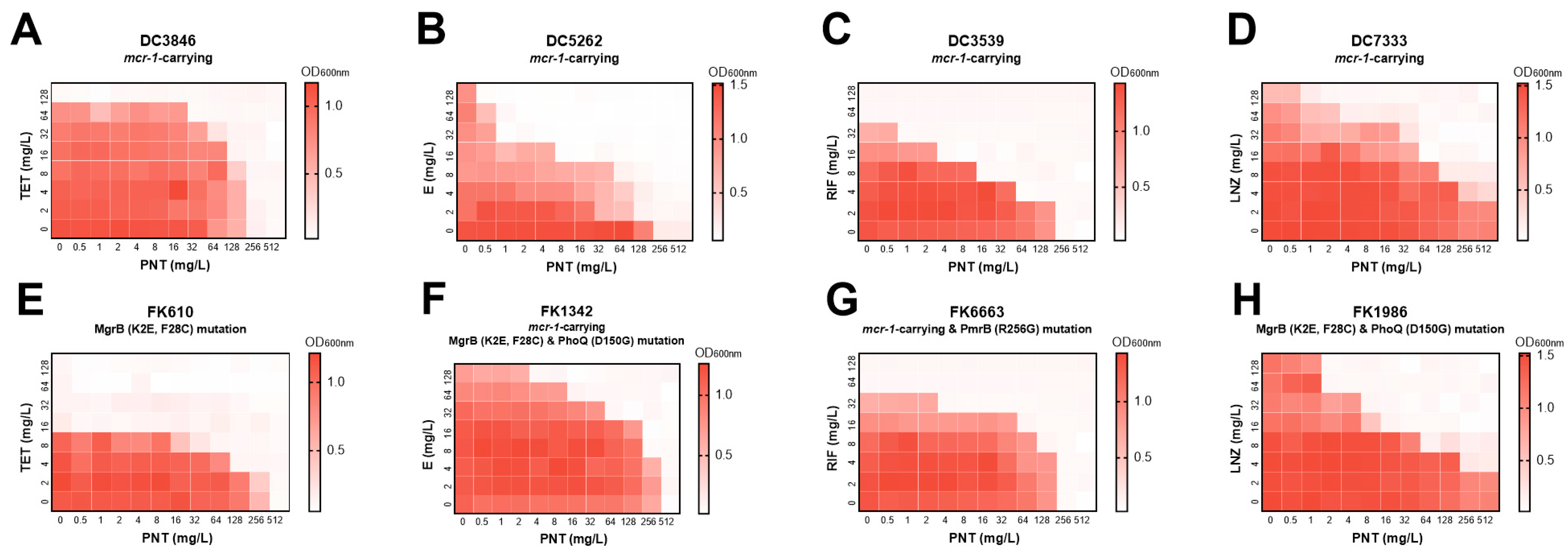
2.1. Pentamidine Potentiates Rifampicin, Linezolid, Erythromycin, and Tetracycline except for Vancomycin against Multiple GNB
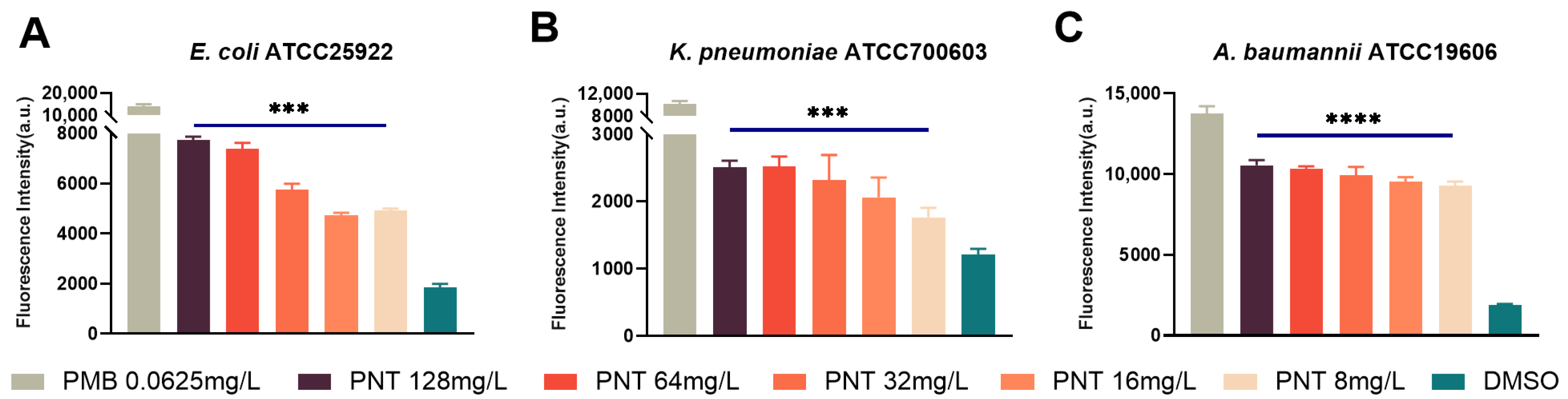
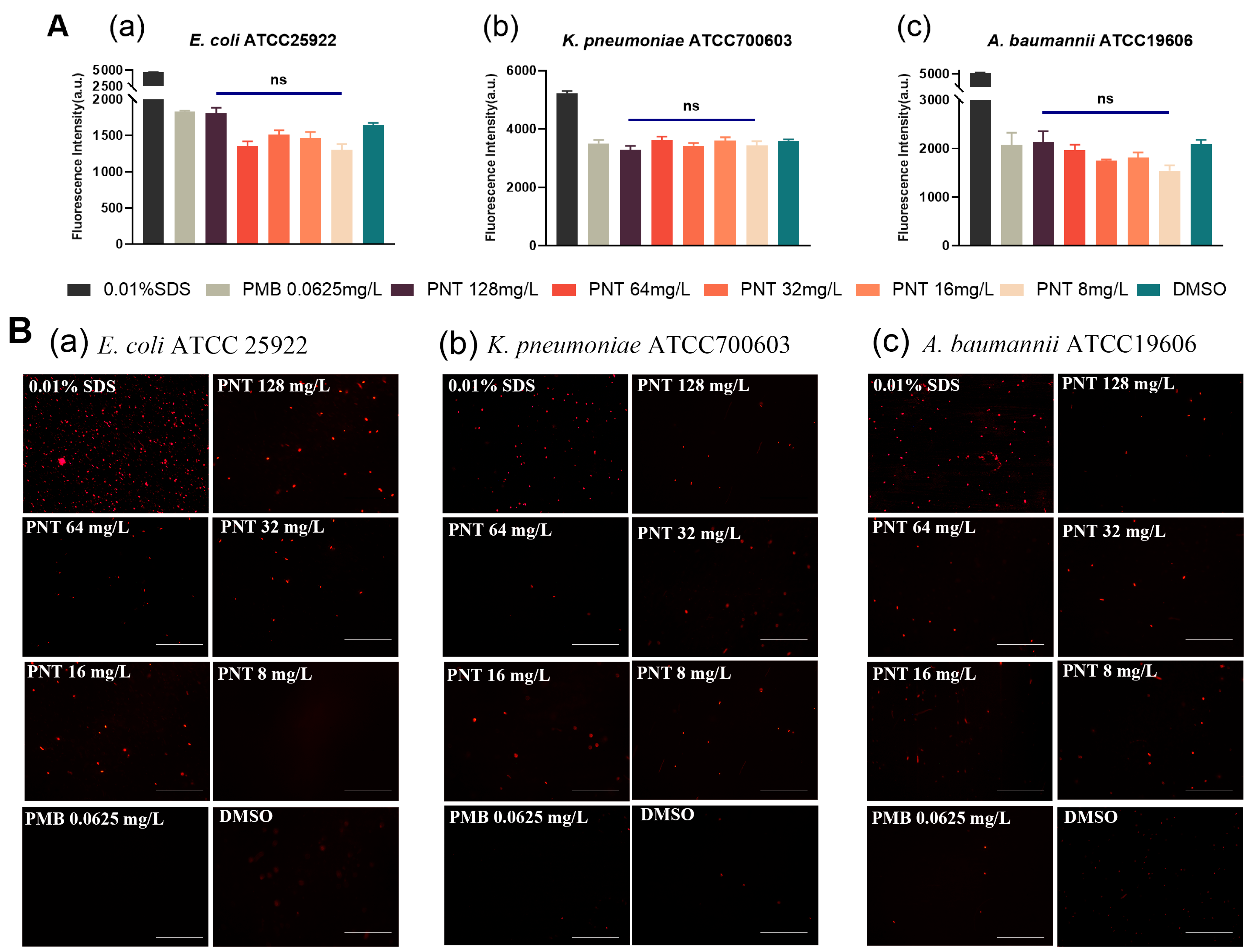
2.2. Pentamidine Preferentially Targets the Outer Membrane of GNB
2.3. Involvement of LPS Is Essential for Pentamidine to Disturb OM
2.4. Pentamidine Increases the OM Permeability but Does Not Demolish the OM
2.5. Pentamidine Does Not Disturb the Inner Membrane (IM)
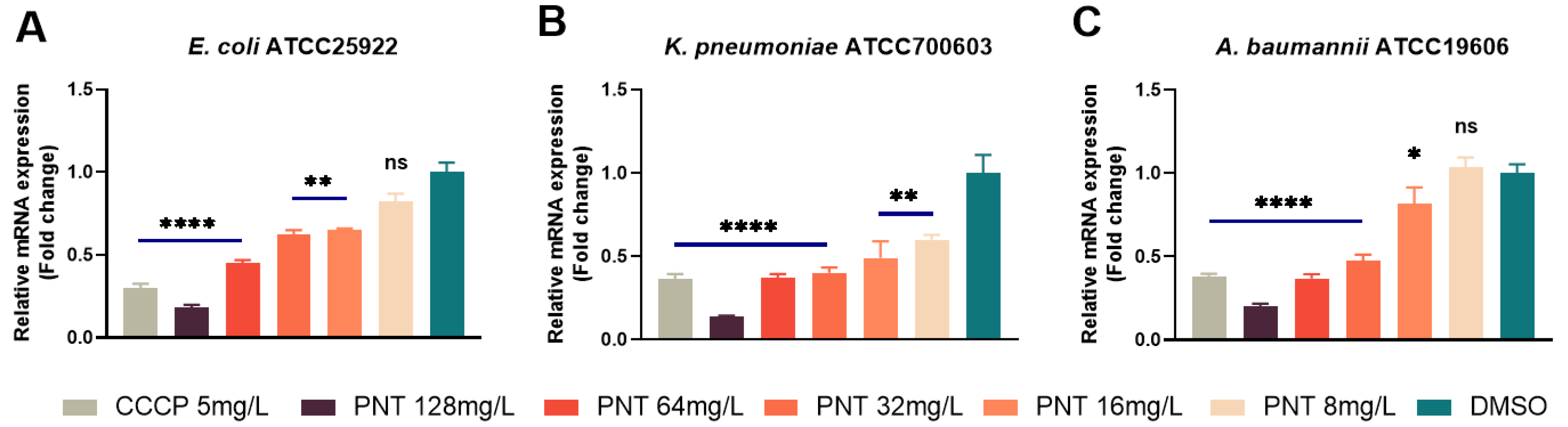
2.6. Pentamidine Dissipates the Membrane Proton Motive Force (PMF)
2.7. Pentamidine Inhibits Efflux Pumps
2.8. Reactive Oxygen Species (ROS) Production Is the Endpoint for Bacterial Death under Pentamidine
3. Materials and Methods
3.1. Bacterial Strains, Agents and MICs
3.2. Checkerboard Assay
3.3. Membrane Permeability Assay
3.4. Intracellular Enzyme Leakage Assays
3.5. LPS Neutralization Assay
3.6. Membrane Potential Assay
3.7. Acridine Orange (AO) Intracellular Accumulation Assay
3.8. Quantitative Real-Time PCR (RT-qPCR) Analysis
3.9. Reactive Oxygen Species (ROS) Detection
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Tong, Z.; Shi, J.; Li, R.; Upton, M.; Wang, Z. Drug repurposing for next-generation combination therapies against multidrug-resistant bacteria. Theranostics 2021, 11, 4910–4928. [Google Scholar] [CrossRef] [PubMed]
- Nguewa, P.A.; Fuertes, M.A.; Cepeda, V.; Iborra, S.; Carrion, J.; Valladares, B.; Alonso, C.; Perez, J.M. Pentamidine is an antiparasitic and apoptotic drug that selectively modifies ubiquitin. Chem. Biodivers. 2005, 2, 1387–1400. [Google Scholar] [CrossRef]
- Wu, C.; Xia, L.; Huang, W.; Xu, Y.; Gu, Y.; Liu, C.; Ji, L.; Li, W.; Wu, Y.; Zhou, K.; et al. Pentamidine sensitizes FDA-approved non-antibiotics for the inhibition of multidrug-resistant Gram-negative pathogens. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1771–1779. [Google Scholar] [CrossRef]
- Stokes, J.M.; MacNair, C.R.; Ilyas, B.; French, S.; Cote, J.P.; Bouwman, C.; Farha, M.A.; Sieron, A.O.; Whitfield, C.; Coombes, B.K.; et al. Pentamidine sensitizes Gram-negative pathogens to antibiotics and overcomes acquired colistin resistance. Nat. Microbiol. 2017, 2, 17028. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Espejo, S.; Cebrero-Cangueiro, T.; Labrador-Herrera, G.; Pachon, J.; Pachon-Ibanez, M.E.; Alvarez-Marin, R. In Vitro Activity of Pentamidine Alone and in Combination with Antibiotics against Multidrug-Resistant Clinical Pseudomonas aeruginosa Strains. Antibiotics 2020, 9, 885. [Google Scholar] [CrossRef]
- Cebrero-Cangueiro, T.; Alvarez-Marin, R.; Labrador-Herrera, G.; Smani, Y.; Cordero-Matia, E.; Pachon, J.; Pachon-Ibanez, M.E. In vitro Activity of Pentamidine Alone and in Combination With Aminoglycosides, Tigecycline, Rifampicin, and Doripenem Against Clinical Strains of Carbapenemase-Producing and/or Colistin-Resistant Enterobacteriaceae. Front. Cell. Infect. Microbiol. 2018, 8, 363. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Qian, C.; Zhang, X.; Liu, Y.; Pan, W.; Yao, Z.; Zeng, W.; Xu, C.; Zhou, T. When Combined with Pentamidine, Originally Ineffective Linezolid Becomes Active in Carbapenem-Resistant Enterobacteriaceae. Microbiol. Spectr. 2023, 11, e0313822. [Google Scholar] [CrossRef]
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef]
- Brown, J.S.; Mohamed, Z.J.; Artim, C.M.; Thornlow, D.N.; Hassler, J.F.; Rigoglioso, V.P.; Daniel, S.; Alabi, C.A. Antibacterial isoamphipathic oligomers highlight the importance of multimeric lipid aggregation for antibacterial potency. Commun. Biol. 2018, 1, 220. [Google Scholar] [CrossRef]
- Armengol, E.; Domenech, O.; Fuste, E.; Perez-Guillen, I.; Borrell, J.H.; Sierra, J.M.; Vinas, M. Efficacy of combinations of colistin with other antimicrobials involves membrane fluidity and efflux machinery. Infect. Drug Resist. 2019, 12, 2031–2038. [Google Scholar] [CrossRef]
- Armengol, E.; Asuncion, T.; Vinas, M.; Sierra, J.M. When Combined with Colistin, an Otherwise Ineffective Rifampicin-Linezolid Combination Becomes Active in Escherichia coli, Pseudomonas aeruginosa, and Acinetobacter baumannii. Microorganisms 2020, 8, 86. [Google Scholar] [CrossRef]
- Mehta, D.; Saini, V.; Aggarwal, B.; Khan, A.; Bajaj, A. Unlocking the bacterial membrane as a therapeutic target for next-generation antimicrobial amphiphiles. Mol. Aspects Med. 2021, 81, 100999. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Kim, M.C.; Park, S.J.; Kim, H.S.; Sung, H.; Kim, M.N.; Kim, S.H.; Lee, S.O.; Choi, S.H.; Woo, J.H.; et al. In Vitro Synergistic Activity of Antimicrobial Agents in Combination against Clinical Isolates of Colistin-Resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 6774–6779. [Google Scholar] [CrossRef]
- Zhou, Y.; Huang, W.; Lei, E.; Yang, A.; Li, Y.; Wen, K.; Wang, M.; Li, L.; Chen, Z.; Zhou, C.; et al. Cooperative Membrane Damage as a Mechanism for Pentamidine-Antibiotic Mutual Sensitization. ACS Chem. Biol. 2022, 17, 3178–3190. [Google Scholar] [CrossRef] [PubMed]
- Hashemian, S.M.R.; Farhadi, T.; Ganjparvar, M. Linezolid: A review of its properties, function, and use in critical care. Drug Des. Devel. Ther. 2018, 12, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Palzkill, T. Deep Mutational Scanning Reveals the Active-Site Sequence Requirements for the Colistin Antibiotic Resistance Enzyme MCR-1. mBio 2021, 12, e0277621. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wu, D.; Wang, C.; Shan, A.; Bi, C.; Li, Y.; Gan, W. Hybridization with Insect Cecropin A (1-8) Improve the Stability and Selectivity of Naturally Occurring Peptides. Int. J. Mol. Sci. 2020, 21, 1470. [Google Scholar] [CrossRef]
- Dezanet, C.; Kempf, J.; Mingeot-Leclercq, M.P.; Decout, J.L. Amphiphilic Aminoglycosides as Medicinal Agents. Int. J. Mol. Sci. 2020, 21, 7411. [Google Scholar] [CrossRef]
- Tang, H.; Zhang, P.; Kieft, T.L.; Ryan, S.J.; Baker, S.M.; Wiesmann, W.P.; Rogelj, S. Antibacterial action of a novel functionalized chitosan-arginine against Gram-negative bacteria. Acta Biomater. 2010, 6, 2562–2571. [Google Scholar] [CrossRef] [PubMed]
- Nagy, T.A.; Crooks, A.L.; Quintana, J.L.J.; Detweiler, C.S. Clofazimine Reduces the Survival of Salmonella enterica in Macrophages and Mice. ACS Infect. Dis. 2020, 6, 1238–1249. [Google Scholar] [CrossRef]
- Dombach, J.L.; Quintana, J.L.J.; Nagy, T.A.; Wan, C.; Crooks, A.L.; Yu, H.; Su, C.C.; Yu, E.W.; Shen, J.; Detweiler, C.S. A small molecule that mitigates bacterial infection disrupts Gram-negative cell membranes and is inhibited by cholesterol and neutral lipids. PLoS Pathog. 2020, 16, e1009119. [Google Scholar] [CrossRef] [PubMed]
- Wesseling, C.M.J.; Slingerland, C.J.; Veraar, S.; Lok, S.; Martin, N.I. Structure-Activity Studies with Bis-Amidines That Potentiate Gram-Positive Specific Antibiotics against Gram-Negative Pathogens. ACS Infect. Dis. 2021, 7, 3314–3335. [Google Scholar] [CrossRef] [PubMed]
- van Groesen, E.; Slingerland, C.J.; Innocenti, P.; Mihajlovic, M.; Masereeuw, R.; Martin, N.I. Vancomyxins: Vancomycin-Polymyxin Nonapeptide Conjugates That Retain Anti-Gram-Positive Activity with Enhanced Potency against Gram-Negative Strains. ACS Infect. Dis. 2021, 7, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Mozaheb, N.; Van Der Smissen, P.; Opsomer, T.; Mignolet, E.; Terrasi, R.; Paquot, A.; Larondelle, Y.; Dehaen, W.; Muccioli, G.G.; Mingeot-Leclercq, M.P. Contribution of Membrane Vesicle to Reprogramming of Bacterial Membrane Fluidity in Pseudomonas aeruginosa. mSphere 2022, 7, e0018722. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.E.; Rhomberg, P.R.; Lister, T.; Cotroneo, N.; Parr, T.R.; Castanheira, M. In Vitro Activity Analysis of a New Polymyxin, SPR741, Tested in Combination with Antimicrobial Agents against a Challenge Set of Enterobacteriaceae, Including Molecularly Characterized Strains. Antimicrob. Agents Chemother. 2020, 65, e00742-20. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, C.S.; Wright, A. Fusions of secreted proteins to alkaline phosphatase: An approach for studying protein secretion. Proc. Natl. Acad. Sci. USA 1985, 82, 5107–5111. [Google Scholar] [CrossRef]
- Elfageih, R.; Karyolaimos, A.; Kemp, G.; de Gier, J.W.; von Heijne, G.; Kudva, R. Cotranslational folding of alkaline phosphatase in the periplasm of Escherichia coli. Protein Sci. 2020, 29, 2028–2037. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.J.; Ingram, J.M.; Costerton, J.W. Interactions of alkaline phosphatase and the cell wall of Pseudomonas aeruginosa. J. Bacteriol. 1971, 107, 325–336. [Google Scholar] [CrossRef]
- Hyun, S.; Choi, Y.; Jo, D.; Choo, S.; Park, T.W.; Park, S.J.; Kim, S.; Lee, S.; Park, S.; Jin, S.M.; et al. Proline Hinged Amphipathic alpha-Helical Peptide Sensitizes Gram-Negative Bacteria to Various Gram-Positive Antibiotics. J. Med. Chem. 2020, 63, 14937–14950. [Google Scholar] [CrossRef] [PubMed]
- Van Moll, L.; De Smet, J.; Paas, A.; Tegtmeier, D.; Vilcinskas, A.; Cos, P.; Van Campenhout, L. In Vitro Evaluation of Antimicrobial Peptides from the Black Soldier Fly (Hermetia Illucens) against a Selection of Human Pathogens. Microbiol. Spectr. 2022, 10, e0166421. [Google Scholar] [CrossRef]
- Tyagi, A.K.; Malik, A. Morphostructural Damage in Food-Spoiling Bacteria due to the Lemon Grass Oil and Its Vapour: SEM, TEM, and AFM Investigations. Evid. Based Complement. Alternat. Med. 2012, 2012, 692625. [Google Scholar] [CrossRef]
- Majumdar, S.; Hingorani, T.; Srirangam, R. Evaluation of active and passive transport processes in corneas extracted from preserved rabbit eyes. J. Pharm. Sci. 2010, 99, 1921–1930. [Google Scholar] [CrossRef]
- Mohiuddin, S.G.; Ghosh, S.; Kavousi, P.; Orman, M.A. Proton Motive Force Inhibitors Are Detrimental to Methicillin-Resistant Staphylococcus aureus Strains. Microbiol. Spectr. 2022, 10, e0202422. [Google Scholar] [CrossRef]
- Chung, E.M.C.; Dean, S.N.; Propst, C.N.; Bishop, B.M.; van Hoek, M.L. Komodo dragon-inspired synthetic peptide DRGN-1 promotes wound-healing of a mixed-biofilm infected wound. NPJ Biofilms Microbiomes 2017, 3, 9. [Google Scholar] [CrossRef]
- Cox, G.; Koteva, K.; Wright, G.D. An unusual class of anthracyclines potentiate Gram-positive antibiotics in intrinsically resistant Gram-negative bacteria. J. Antimicrob. Chemother. 2014, 69, 1844–1855. [Google Scholar] [CrossRef] [PubMed]
- Kascakova, S.; Maigre, L.; Chevalier, J.; Refregiers, M.; Pages, J.M. Antibiotic transport in resistant bacteria: Synchrotron UV fluorescence microscopy to determine antibiotic accumulation with single cell resolution. PLoS ONE 2012, 7, e38624. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Karim, S.A.M.; El-Ganiny, A.M.A.; El-Sayed, M.A.; Abbas, H.A.A. Promising FDA-approved drugs with efflux pump inhibitory activities against clinical isolates of Staphylococcus aureus. PLoS ONE 2022, 17, e0272417. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.A.; Rolain, J.M. Efflux pump inhibitor CCCP to rescue colistin susceptibility in mcr-1 plasmid-mediated colistin-resistant strains and Gram-negative bacteria. J. Antimicrob. Chemother. 2018, 73, 1862–1871. [Google Scholar] [CrossRef] [PubMed]
- Sionov, R.V.; Steinberg, D. Targeting the Holy Triangle of Quorum Sensing, Biofilm Formation, and Antibiotic Resistance in Pathogenic Bacteria. Microorganisms 2022, 10, 1239. [Google Scholar] [CrossRef] [PubMed]
- Klenotic, P.A.; Moseng, M.A.; Morgan, C.E.; Yu, E.W. Structural and Functional Diversity of Resistance-Nodulation-Cell Division Transporters. Chem. Rev. 2021, 121, 5378–5416. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.L.; Bavro, V.N. Mutations in the TolC Periplasmic Domain Affect Substrate Specificity of the AcrAB-TolC Pump. Front. Mol. Biosci. 2020, 7, 166. [Google Scholar] [CrossRef]
- Blair, J.M.; Piddock, L.J. How to Measure Export via Bacterial Multidrug Resistance Efflux Pumps. mBio 2016, 7, e00840-16. [Google Scholar] [CrossRef] [PubMed]
- Kalsy, M.; Tonk, M.; Hardt, M.; Dobrindt, U.; Zdybicka-Barabas, A.; Cytrynska, M.; Vilcinskas, A.; Mukherjee, K. The insect antimicrobial peptide cecropin A disrupts uropathogenic Escherichia coli biofilms. NPJ Biofilms Microbiomes 2020, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Maiden, M.M.; Waters, C.M. Triclosan depletes the membrane potential in Pseudomonas aeruginosa biofilms inhibiting aminoglycoside induced adaptive resistance. PLoS Pathog. 2020, 16, e1008529. [Google Scholar] [CrossRef]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Mode of action of the antimicrobial peptide Mel4 is independent of Staphylococcus aureus cell membrane permeability. PLoS ONE 2019, 14, e0215703. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Comparative mode of action of the antimicrobial peptide melimine and its derivative Mel4 against Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 7063. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Xing, L.; Qu, P.; Tan, T.; Yang, N.; Li, D.; Chen, H.; Feng, X. Identification of a novel cathelicidin antimicrobial peptide from ducks and determination of its functional activity and antibacterial mechanism. Sci. Rep. 2015, 5, 17260. [Google Scholar] [CrossRef]
- Xia, Y.; Cebrian, R.; Xu, C.; Jong, A.; Wu, W.; Kuipers, O.P. Elucidating the mechanism by which synthetic helper peptides sensitize Pseudomonas aeruginosa to multiple antibiotics. PLoS Pathog. 2021, 17, e1009909. [Google Scholar] [CrossRef]
- Dohi, N.; Yamaguchi, M.; Hase, R.; Suzuki, R.; Wakabayashi, Y.; Nishiyama, R.; Saito, S.Y.; Ishikawa, T. Quantitative real-time measurement of endothelin-1-induced contraction in single non-activated hepatic stellate cells. PLoS ONE 2021, 16, e0255656. [Google Scholar] [CrossRef]
- Song, M.; Liu, Y.; Huang, X.; Ding, S.; Wang, Y.; Shen, J.; Zhu, K. A broad-spectrum antibiotic adjuvant reverses multidrug-resistant Gram-negative pathogens. Nat. Microbiol. 2020, 5, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- MacNair, C.R.; Farha, M.A.; Serrano-Wu, M.H.; Lee, K.K.; Hubbard, B.; Cote, J.P.; Carfrae, L.A.; Tu, M.M.; Gaulin, J.L.; Hunt, D.K.; et al. Preclinical Development of Pentamidine Analogs Identifies a Potent and Nontoxic Antibiotic Adjuvant. ACS Infect. Dis. 2022, 8, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, Y.; Zhang, X.; Chen, L.; Xu, C.; Liu, S.; Cao, J.; Zheng, X.; Jia, H.; Chen, L.; et al. Combining colistin with furanone C-30 rescues colistin resistance of gram-negative bacteria in vitro and in vivo. Microbiol. Spectr. 2021, 9, e0123121. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strains | Monotherapy (mg/L) | Combination (mg/L) | FICI | Interpretation | ||
|---|---|---|---|---|---|---|---|
| PNT | LNZ | PNT | LNZ | ||||
| E. coli | ATCC25922 | 256 | >64 | 32 | 16 | <0.375 | Synergistic |
| K. pneumoniae | ATCC700603 | >512 | >64 | 128 | 8 | <0.375 | Synergistic |
| A. baumannii | ATCC19606 | 512 | >64 | 32 | 8 | <0.1875 | Synergistic |
| K. pneumoniae | CR FK7921 | 512 | 512 | 64 | 32 | 0.1875 | Synergistic |
| E. cloacae | CR CG1779 | 2048 | 1024 | 64 | 128 | 0.1563 | Synergistic |
| E. coli | Col-R DC7333 | >512 | >128 | 32 | 32 | 0.3125 | Synergistic |
| K. pneumoniae | Col-R FK1986 | >512 | >128 | 16 | 16 | 0.1563 | Synergistic |
| P. aeruginosa | ATCC27853 | 256 | >64 | 256 | >64 | 2 | No interaction |
| PNT | RIF | PNT | RIF | ||||
| E. coli | ATCC25922 | 256 | 16 | 32 | 4 | 0.375 | Synergistic |
| K. pneumoniae | ATCC700603 | >512 | 32 | 64 | 8 | <0.375 | Synergistic |
| A. baumannii | ATCC19606 | 512 | 16 | 128 | 8 | 0.75 | Additive |
| E. coli | Col-R DC3539 | 256 | 64 | 16 | 8 | 0.1875 | Synergistic |
| K. pneumoniae | Col-R FK6663 | 256 | 64 | 64 | 16 | 0.5 | Synergistic |
| PNT | E | PNT | E | ||||
| E. coli | ATCC25922 | 256 | 128 | 64 | 32 | 0.5 | Synergistic |
| A. baumannii | ATCC19606 | 512 | 128 | 128 | 32 | 0.5 | Synergistic |
| E. coli | Col-R DC5262 | 256 | >128 | 8 | 16 | 0.1563 | Synergistic |
| K. pneumoniae | Col-R FK1342 | 512 | >128 | 64 | 32 | <0.375 | Synergistic |
| PNT | TET | PNT | TET | ||||
| K. pneumoniae | ATCC700603 | >512 | 32 | 64 | 8 | <0.375 | Synergistic |
| K. pneumoniae | Col-R FK610 | 512 | 16 | 128 | 4 | 0.5 | Synergistic |
| E. coli | Col-R DC3846 | 256 | 128 | 64 | 32 | 0.5 | Synergistic |
| K. pneumoniae | Col-R FK6663 | 256 | 64 | 64 | 16 | 0.5 | Synergistic |
| PNT | VA | PNT | VA | ||||
| E. coli | ATCC25922 | 256 | >128 | 256 | >128 | 2 | No interaction |
| K. pneumoniae | CR FK7921 | 512 | 128 | 512 | 128 | 2 | No interaction |
| A. baumannii | ATCC19606 | 512 | >128 | 512 | >128 | 2 | No interaction |
| Strain | RIF | RIF + PMB a | RIF + CCCP b | LNZ | LNZ + PMB a | LNZ + CCCP b | VA | VA + PMB a | VA + CCCP b |
|---|---|---|---|---|---|---|---|---|---|
| RIF MIC (mg/L) | LNZ MIC (mg/L) | VA MIC (mg/L) | |||||||
| E. coli ATCC25922 | 16 | <0.5 | 8 | 1024 | 512 | <0.5 | 1024 | 64 | 1024 |
| K. pneumoniae ATCC700603 | 32 | <0.5 | 8 | 512 | 256 | <0.5 | 512 | 32 | 512 |
| P. aeruginosa ATCC27853 | 16 | <0.5 | 4 | 256 | 128 | <0.5 | 512 | 16 | 512 |
| A. baumannii ATCC19606 | 16 | <0.5 | 8 | 1024 | 512 | 64 | 1024 | 128 | 1024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, M.; Zhao, D.; Liu, S.; Zhang, X.; Yao, Z.; Chen, H.; Zhou, C.; Zhou, T.; Xu, C. The Properties of Linezolid, Rifampicin, and Vancomycin, as Well as the Mechanism of Action of Pentamidine, Determine Their Synergy against Gram-Negative Bacteria. Int. J. Mol. Sci. 2023, 24, 13812. https://doi.org/10.3390/ijms241813812
Tang M, Zhao D, Liu S, Zhang X, Yao Z, Chen H, Zhou C, Zhou T, Xu C. The Properties of Linezolid, Rifampicin, and Vancomycin, as Well as the Mechanism of Action of Pentamidine, Determine Their Synergy against Gram-Negative Bacteria. International Journal of Molecular Sciences. 2023; 24(18):13812. https://doi.org/10.3390/ijms241813812
Chicago/Turabian StyleTang, Miran, Deyi Zhao, Sichen Liu, Xiaotuan Zhang, Zhuocheng Yao, Hule Chen, Cui Zhou, Tieli Zhou, and Chunquan Xu. 2023. "The Properties of Linezolid, Rifampicin, and Vancomycin, as Well as the Mechanism of Action of Pentamidine, Determine Their Synergy against Gram-Negative Bacteria" International Journal of Molecular Sciences 24, no. 18: 13812. https://doi.org/10.3390/ijms241813812