
MiRNA-146a—A Key Player in Immunity and Diseases
 , , , , and
, , , , and 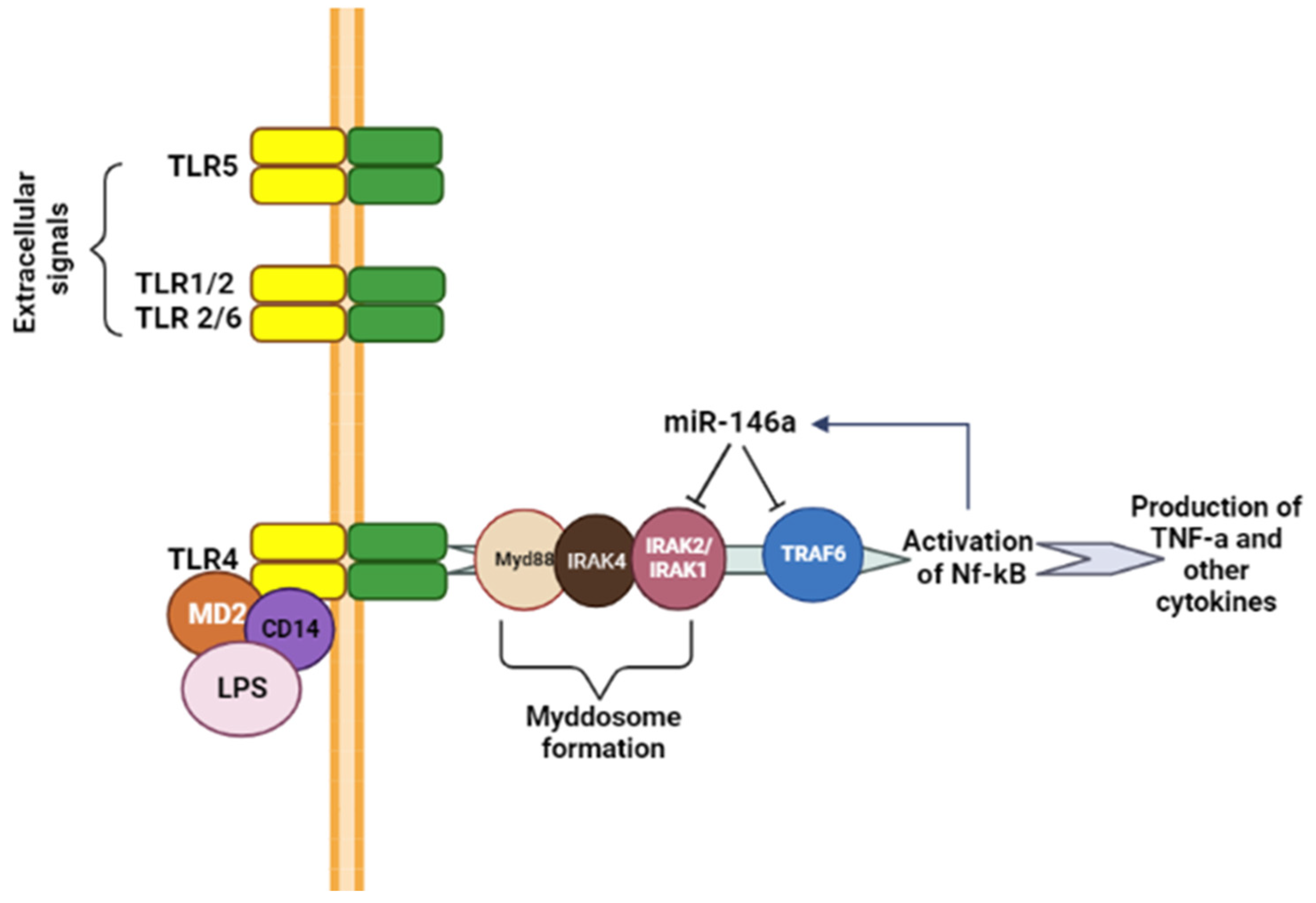{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
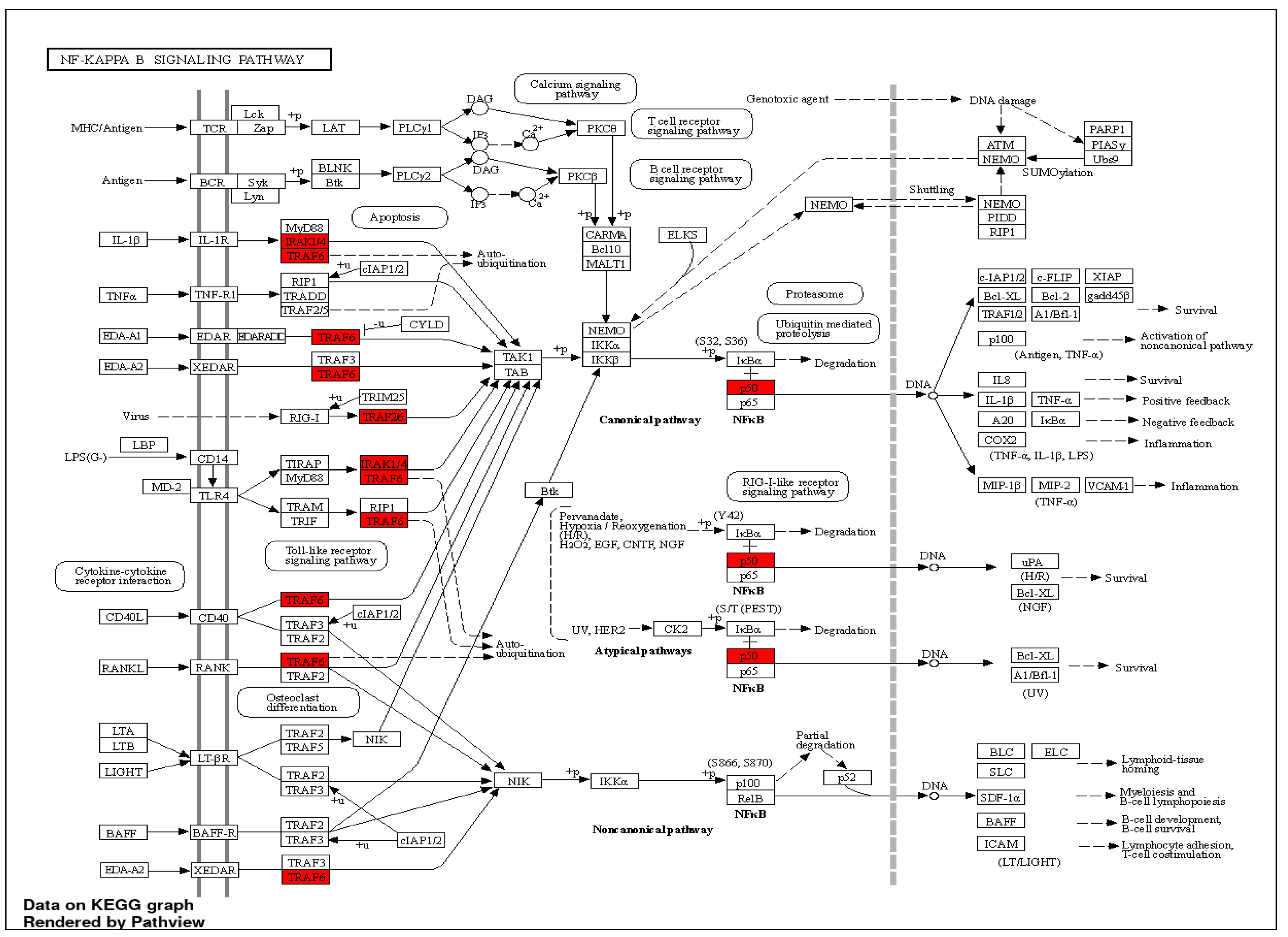
2. MiRNA-146a—Function, Role, Involvement in Innate Immunity, Development of Inflammatory Responses, and Haematopoiesis
3. Role of miRNA-146a in the Development of Autoimmune Diseases and Allergic Reactions
4. miRNA-146a in Infection Diseases
5. Participation of miRNA-146a in the Pathogenesis of Malignant Tumours of Various Localizations
5.1. miRNA-146a as a Tumour Suppressor
5.2. miRNA-146a as an Oncogene
5.3. Involvement of miRNA-146a in Tumour Metastasis
5.4. miRNA-146a as Regulator and the Role in Response to Cancer Therapy
6. Predicted Gene Targets of miRNA-146a
Immune Genes
7. Participation of miRNA-146a in the Development of Ischemic Stroke
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, H.; Wassmer, S.C. Harnessing the Potential of miRNAs in Malaria Diagnostic and Prevention. Front. Cell. Infect. Microbiol. 2021, 11, 793954. [Google Scholar] [CrossRef]
- Boldin, M.P.; Taganov, K.D.; Rao, D.S.; Yang, L.; Zhao, J.L.; Kalwani, M.; Garcia-Flores, Y.; Luong, M.; Devrekanli, A.; Xu, J.; et al. miR-146a is a significant brake on autoimmunity, myeloproliferation, and cancer in mice. J. Exp. Med. 2011, 208, 1189–1201. [Google Scholar] [CrossRef]
- Chen, C.-Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs Modulate Hematopoietic Lineage Differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbaye, C.; Testa, U. The emerging role of MIR-146A in the control of hematopoiesis, immune function and cancer. J. Hematol. Oncol. 2012, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.L.; Rao, D.S.; Boldin, M.P.; Taganov, K.D.; O’Connell, R.M.; Baltimore, D. NF-κB dysregulation in microRNA-146a–deficient mice drives the development of myeloid malignancies. Proc. Natl. Acad. Sci. USA 2011, 108, 9184–9189. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, L.A.; Sheedy, F.J.; McCoy, C.E. MicroRNAs: The fine-tuners of Toll-like receptor signalling. Nat. Rev. Immunol. 2011, 11, 163–175. [Google Scholar] [CrossRef]
- Esaba, R.; Sorensen, D.L.; Booth, S.A. MicroRNA-146a: A Dominant, Negative Regulator of the Innate Immune Response. Front. Immunol. 2014, 5, 578. [Google Scholar] [CrossRef]
- Chen, Y.; Zeng, Z.; Shen, X.; Wu, Z.; Dong, Y.; Cheng, J.C.-H. MicroRNA-146a-5p Negatively Regulates Pro-Inflammatory Cytokine Secretion and Cell Activation in Lipopolysaccharide Stimulated Human Hepatic Stellate Cells through Inhibition of Toll-Like Receptor 4 Signaling Pathways. Int. J. Mol. Sci. 2016, 17, 1076. [Google Scholar] [CrossRef] [Green Version]
- Nahid, A.; Pauley, K.M.; Satoh, M.; Chan, E.K.L. miR-146a Is Critical for Endotoxin-induced Tolerance. J. Biol. Chem. 2009, 284, 34590–34599. [Google Scholar] [CrossRef] [Green Version]
- Nahid, A.; Satoh, M.; Chan, E.K.L. Mechanistic Role of MicroRNA-146a in Endotoxin-Induced Differential Cross-Regulation of TLR Signaling. J. Immunol. 2011, 186, 1723–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, A.E.; Perry, M.M.; Moschos, S.A.; Larner-Svensson, H.M.; Lindsay, M.A. Role of miRNA-146a in the regulation of the innate immune response and cancer. Biochem. Soc. Trans. 2008, 36, 1211–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starczynowski, D.T.; Kuchenbauer, F.; Wegrzyn, J.; Rouhi, A.; Petriv, O.; Hansen, C.L.; Humphries, R.K.; Karsan, A. MicroRNA-146a disrupts hematopoietic differentiation and survival. Exp. Hematol. 2011, 39, 167–178.e4. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Volinia, S.; Liu, C.-G.; Fernandez-Cymering, C.; Palumbo, T.; Pichiorri, F.; Fabbri, M.; Coombes, K.; Alder, H.; Nakamura, T.; et al. MicroRNA signatures associated with cytogenetics and prognosis in acute myeloid leukemia. Blood 2008, 111, 3183–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, H.; Wang, H.-R.; Yang, S.; Zhong, J.-H.; Wang, T.; Wang, C.; Chen, F.-Y. Targeting Smad4 links microRNA-146a to the TGF-β pathway during retinoid acid induction in acute promyelocytic leukemia cell line. Int. J. Hematol. 2010, 92, 129–135. [Google Scholar] [CrossRef]
- Labbaye, C.; Spinello, I.; Quaranta, M.T.; Pelosi, E.; Pasquini, L.; Petrucci, E.; Biffoni, M.; Nuzzolo, E.R.; Billi, M.; Foà, R.; et al. A three-step pathway comprising PLZF/miR-146a/CXCR4 controls megakaryopoiesis. Nature 2008, 10, 788–801. [Google Scholar] [CrossRef]
- Landry, P.; Plante, I.; Ouellet, D.L.; Perron, M.P.; Rousseau, G.; Provost, P. Existence of a microRNA pathway in anucleate platelets. Nat. Struct. Mol. Biol. 2009, 16, 961–966. [Google Scholar] [CrossRef] [Green Version]
- Opalinska, J.B.; Bersenev, A.; Zhang, Z.; Schmaier, A.A.; Choi, J.; Yao, Y.; D’Souza, J.; Tong, W.; Weiss, M.J. MicroRNA expression in maturing murine megakaryocytes. Blood 2010, 116, e128–e138. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.L.; Starczynowski, D.T. Role of microRNA-146a in normal and malignant hematopoietic stem cell function. Front. Genet. 2014, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Dixon-McIver, A.; East, P.; Mein, C.A.; Cazier, J.-B.; Molloy, G.; Chaplin, T.; Lister, T.A.; Young, B.D.; Debernardi, S. Distinctive Patterns of MicroRNA Expression Associated with Karyotype in Acute Myeloid Leukaemia. PLoS ONE 2008, 3, e2141. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, H.; Lei, D. microRNA-146a Promotes Growth of Acute Leukemia Cells by Downregulating Ciliary Neurotrophic Factor Receptor and Activating JAK2/STAT3 Signaling. Yonsei Med. J. 2019, 60, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-L.; Wu, X.-X.; Wang, X.-F.; Di, D.-S.; Huang, Q.; Liu, R.-S.; Shuai, Z.-W.; Ye, D.-Q.; Leng, R.-X. Genetic variant in microRNA-146a gene is associated with risk of rheumatoid arthritis. Ann. Med. 2021, 53, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, H.; Chen, L.; Wang, Y.; Yao, X.; Jiang, X. Association of microRNAs genes polymorphisms with arthritis: A systematic review and meta-analysis. Biosci. Rep. 2019, 39, BSR20190298. [Google Scholar] [CrossRef]
- Zhou, M.; Jiang, B.; Xiong, M.; Zhu, X. An Updated Meta-Analysis of the Associations Between MicroRNA Polymorphisms and Susceptibility to Rheumatoid Arthritis. Front. Physiol. 2018, 9, 1604. [Google Scholar] [CrossRef]
- Leng, R.-X.; Di, D.-S.; Ni, J.; Wu, X.-X.; Zhang, L.-L.; Wang, X.-F.; Liu, R.-S.; Huang, Q.; Fan, Y.-G.; Pan, H.-F.; et al. Identification of new susceptibility loci associated with rheumatoid arthritis. Ann. Rheum. Dis. 2020, 79, 1565–1571. [Google Scholar] [CrossRef]
- Fairhurst, A.; Wandstrat, A.E.; Wakeland, E.K. Systemic Lupus Erythematosus: Multiple Immunological Phenotypes in a Complex Genetic Disease. Adv. Immunol. 2006, 92, 1–69. [Google Scholar] [CrossRef] [PubMed]
- Kirou, K.A.; Lee, C.; George, S.; Louca, K.; Papagiannis, I.G.; Peterson, M.G.E.; Ly, N.; Woodward, R.N.; Fry, K.E.; Lau, A.Y.-H.; et al. Coordinate Overexpression of Interferon-Induced Genes in Systemic Lupus Erythematosus. Arthritis Rheum. 2004, 50, 3958–3967. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Yang, W.; Ye, D.-Q.; Cui, H.; Zhang, Y.; Hirankarn, N.; Qian, X.; Tang, Y.; Lau, Y.L.; de Vries, N.; et al. A Functional Variant in MicroRNA-146a Promoter Modulates Its Expression and Confers Disease Risk for Systemic Lupus Erythematosus. PLoS Genet. 2011, 7, e1002128. [Google Scholar] [CrossRef] [Green Version]
- Löfgren, E.S.; Frostegård, J.; Truedsson, L.; Pons-Estel, A.B.; D’Alfonso, S.; Witte, T.; Lauwerys, B.R.; Endreffy, E.; Kovács, L.; Vasconcelos, C.; et al. Genetic association of miRNA-146a with systemic lupus erythematosus in Europeans through decreased expression of the gene. Genes Immun. 2012, 13, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xin, S.; Wang, Y.; Ju, D.; Wu, Q.; Qiu, Y.; Niu, X.; Liu, W.; Li, J.; Ji, P. MicroRNA-146a-5p enhances T helper 17 cell differentiation via decreasing a disintegrin and metalloprotease 17 level in primary sjögren’s syndrome. Bioengineered 2021, 12, 310–324. [Google Scholar] [CrossRef]
- Hammad, R.H.M.; Hamed, D.H.E.D.; Eldosoky, M.A.E.R.; Ahmad, A.A.E.S.; Osman, H.M.; Elgalil, H.M.A.; Hassan, M.M.M. Plasma microRNA-21, microRNA-146a and IL-13 expression in asthmatic children. Innate Immun. 2018, 24, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.S.; Sivachandran, N.; Lau, A.; Boudreau, E.; Zhao, J.L.; Baltimore, D.; Delgado-Olguin, P.; Cybulsky, M.I.; Fish, J.E. MicroRNA-146 represses endothelial activation by inhibiting pro-inflammatory pathways. EMBO Mol. Med. 2013, 5, 1017–1034. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Liu, X.; Nelson, A.; Nakanishi, M.; Kanaji, N.; Wang, X.; Kim, M.; Li, Y.; Sun, J.; Michalski, J.; et al. Reduced miR-146a Increases Prostaglandin E2 in Chronic Obstructive Pulmonary Disease Fibroblasts. Am. J. Respir. Crit. Care Med. 2010, 182, 1020–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larner-Svensson, H.M.; Williams, E.A.; Tsitsiou, E.; Perry, M.M.; Jiang, X.; Chung, K.F.; Lindsay, A.M. Pharmacological studies of the mechanism and function of interleukin-1β-induced miRNA-146a expression in primary human airway smooth muscle. Respir. Res. 2010, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Laanesoo, A.; Urgard, E.; Periyasamy, K.; Laan, M.; Bochkov, Y.A.; Aab, A.; Magilnick, N.; Pooga, M.; Gern, J.E.; Johnston, S.L.; et al. Dual role of the miR-146 family in rhinovirus-induced airway inflammation and allergic asthma exacerbation. Clin. Transl. Med. 2021, 11, e427. [Google Scholar] [CrossRef] [PubMed]
- Crone, S.G.; Jacobsen, A.; Federspiel, B.; Bardram, L.; Krogh, A.; Lund, A.H.; Friis-Hansen, L. microRNA-146a inhibits G protein-coupled receptor-mediated activation of NF-κB by targeting CARD10 and COPS8 in gastric cancer. Mol. Cancer 2012, 11, 71. [Google Scholar] [CrossRef] [Green Version]
- Comer, B.S.; Camoretti-Mercado, B.; Kogut, P.C.; Halayko, A.J.; Solway, J.; Gerthoffer, W.T. MicroRNA-146a and microRNA-146b expression and anti-inflammatory function in human airway smooth muscle. Am. J. Physiol. Cell Mol. Physiol. 2014, 307, L727–L734. [Google Scholar] [CrossRef]
- Arghiani, N.; Nissan, T.; Matin, M.M. Role of microRNAs in COVID-19 with implications for therapeutics. Biomed. Pharmacother. 2021, 144, 112247. [Google Scholar] [CrossRef]
- Li, Y.; Tan, S.; Shen, Y.; Guo, L. miR-146a-5p negatively regulates the IL-1β-stimulated inflammatory response via downregulation of the IRAK1/TRAF6 signaling pathway in human intestinal epithelial cells. Exp. Ther. Med. 2022, 24, 615. [Google Scholar] [CrossRef]
- Balasubramanyam, M.; Aravind, S.; Gokulakrishnan, K.; Prabu, P.; Sathishkumar, C.; Ranjani, H.; Mohan, V. Impaired miR-146a expression links subclinical inflammation and insulin resistance in Type 2 diabetes. Mol. Cell. Biochem. 2011, 351, 197–205. [Google Scholar] [CrossRef]
- Deng, Y.; Yan, Y.; Tan, K.S.; Liu, J.; Chow, V.T.; Tao, Z.-Z.; Wang, D.-Y. MicroRNA-146a induction during influenza H3N2 virus infection targets and regulates TRAF6 levels in human nasal epithelial cells (hNECs). Exp. Cell Res. 2017, 352, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.-L.; Wang, X.; Mann, M.; Adamus, T.P.; Wang, D.; Moreira, D.F.; Zhang, Z.; Ouyang, C.; He, X.; Zhang, B.; et al. Myeloid cell–targeted miR-146a mimic inhibits NF-κB–driven inflammation and leukemia progression in vivo. Blood 2020, 135, 167–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.-J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [PubMed]
- Kivihall, A.; Aab, A.; Soja, J.; Sładek, K.; Sanak, M.; Altraja, A.; Jakiela, B.; Bochenek, G.; Rebane, A. Reduced expression of miR-146a in human bronchial epithelial cells alters neutrophil migration. Clin. Transl. Allergy 2019, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Rebane, A.; Runnel, T.; Aab, A.; Maslovskaja, J.; Rückert, B.; Zimmermann, M.; Plaas, M.; Kärner, J.; Treis, A.; Pihlap, M.; et al. MicroRNA-146a alleviates chronic skin inflammation in atopic dermatitis through suppression of innate immune responses in keratinocytes. J. Allergy Clin. Immunol. 2014, 134, 836–847.e11. [Google Scholar] [CrossRef]
- Ferreira, L.R.P.; Frade, A.F.; Santos, R.H.B.; Teixeira, P.C.; Baron, M.A.; Navarro, I.C.; Benvenuti, L.A.; Fiorelli, A.I.; Bocchi, E.A.; Stolf, N.A.; et al. MicroRNAs miR-1, miR-133a, miR-133b, miR-208a and miR-208b are dysregulated in Chronic Chagas disease Cardiomyopathy. Int. J. Cardiol. 2014, 175, 409–417. [Google Scholar] [CrossRef]
- Navarro, I.C.; Ferreira, F.M.; Nakaya, H.I.; Baron, M.A.; Vilar-Pereira, G.; Pereira, I.R.; Silva, A.M.G.; Real, J.M.; De Brito, T.; Chevillard, C.; et al. MicroRNA Transcriptome Profiling in Heart of Trypanosoma cruzi-Infected Mice: Parasitological and Cardiological Outcomes. PLoS Negl. Trop. Dis. 2015, 9, e0003828. [Google Scholar] [CrossRef] [Green Version]
- Nonaka, C.K.V.; Macêdo, C.T.; Cavalcante, B.R.R.; de Alcântara, A.C.; Silva, D.N.; Bezerra, M.d.R.; Caria, A.C.I.; Tavora, F.R.F.; Neto, J.D.d.S.; Noya-Rabelo, M.M.; et al. Circulating miRNAs as Potential Biomarkers Associated with Cardiac Remodeling and Fibrosis in Chagas Disease Cardiomyopathy. Int. J. Mol. Sci. 2019, 20, 4064. [Google Scholar] [CrossRef] [Green Version]
- Jha, B.K.; Varikuti, S.; Seidler, G.R.; Volpedo, G.; Satoskar, A.R.; McGwire, B.S. MicroRNA-155 Deficiency Exacerbates Trypanosoma cruzi Infection. Infect. Immun. 2020, 88, e00948-19. [Google Scholar] [CrossRef]
- Houghton, J.M. Helicobacter species methods and protocols. Introduction. Methods Mol. Biol. 2012, 921, 1–5. [Google Scholar] [CrossRef]
- Hou, J.; Wang, P.; Lin, L.; Liu, X.; Ma, F.; An, H.; Wang, Z.; Cao, X. MicroRNA-146a Feedback Inhibits RIG-I-Dependent Type I IFN Production in Macrophages by Targeting TRAF6, IRAK1, and IRAK2. J. Immunol. 2009, 183, 2150–2158. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-Z.; Li, Y.-Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef]
- Boldin, M.P.; Baltimore, D. MicroRNAs, new effectors and regulators of NF-κB. Immunol. Rev. 2012, 246, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Ballinas-Verdugo, M.A.; Jiménez-Ortega, R.F.; Martínez-Martínez, E.; Rivas, N.; Contreras-López, E.A.; Carbó, R.; Sánchez, F.; Bojalil, R.; Márquez-Velasco, R.; Sánchez-Muñoz, F.; et al. Circulating miR-146a as a possible candidate biomarker in the indeterminate phase of Chagas disease. Biol. Res. 2021, 54, 21. [Google Scholar] [CrossRef] [PubMed]
- Rusca, N.; Monticelli, S. MiR-146a in Immunity and Disease. Mol. Biol. Int. 2011, 2011, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Zheng, S.G. Hall of Fame among Pro-inflammatory Cytokines: Interleukin-6 Gene and Its Transcriptional Regulation Mechanisms. Front. Immunol. 2016, 7, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Tang, R.; Sun, Y.; Wang, Y.-G.; Zhen, K.-Y.; Zhang, D.-M.; Pan, W.-Q. MicroR-146 blocks the activation of M1 macrophage by targeting signal transducer and activator of transcription 1 in hepatic schistosomiasis. Ebiomedicine 2016, 13, 339–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvamani, S.P.; Mishra, R.; Singh, S.K. Chikungunya Virus Exploits miR-146a to Regulate NF-κB Pathway in Human Synovial Fibroblasts. PLoS ONE 2014, 9, e103624. [Google Scholar] [CrossRef]
- Sharma, N.; Verma, R.; Kumawat, K.L.; Basu, A.; Singh, S.K. miR-146a suppresses cellular immune response during Japanese encephalitis virus JaOArS982 strain infection in human microglial cells. J. Neuroinflamm. 2015, 12, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Sun, X.; Zhu, Y.; Qin, W. Downregulation of miR-146a inhibits influenza A virus replication by enhancing the type I interferon response in vitro and in vivo. Biomed. Pharmacother. 2019, 111, 740–750. [Google Scholar] [CrossRef]
- Wu, S.; He, L.; Li, Y.; Wang, T.; Feng, L.; Jiang, L.; Zhang, P.; Huang, X. miR-146a facilitates replication of dengue virus by dampening interferon induction by targeting TRAF6. J. Infect. 2013, 67, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.V.; Diaz, A.; D’attilio, L.; Marchesini, M.M.; Bogue, C.; Bay, M.L.; Bottasso, O.A. Altered microRNA expression levels in mononuclear cells of patients with pulmonary and pleural tuberculosis and their relation with components of the immune response. Mol. Immunol. 2013, 53, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Jiang, J.; Cao, Z.; Yang, B.; Cheng, X. Modulation of T cell cytokine production by miR-144* with elevated expression in patients with pulmonary tuberculosis. Mol. Immunol. 2011, 48, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, M.M.M.; Mir, I.; Idrees, M.; Afzal, S.; Shahid, M. Role of MicroRNAs in Establishing Latency of Human Immunodeficiency Virus. Crit. Rev. Eukaryot. Gene Expr. 2020, 30, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Chen, L.; Luo, M.; Lv, H.; Luo, D.; Li, T.; Huang, S.; Xie, L.; Teng, Y.; Liu, Z.; et al. HIV-1-Induced miR-146a Attenuates Monocyte Migration by Targeting CCL5 in Human Primary Macrophages. AIDS Res. Hum. Retrovir. 2018, 34, 580–589. [Google Scholar] [CrossRef]
- Pogue, A.; Zhao, Y.; Lukiw, W. microRNA-146a as a biomarker for transmissible spongiform encephalopathy. Folia Neuropathol. 2022, 60, 24–34. [Google Scholar] [CrossRef]
- Huang, C.; Liu, X.-J.; Zhou, Q.; Xie, J.; Ma, T.-T.; Meng, X.-M.; Li, J. MiR-146a modulates macrophage polarization by inhibiting Notch1 pathway in RAW264.7 macrophages. Int. Immunopharmacol. 2016, 32, 46–54. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Sun, X.; Huang, C.; Long, X.-R.; Lin, X.; Zhang, L.; Lv, X.-W.; Li, J. MiR-146a Regulates IL-6 Production in Lipopolysaccharide-Induced RAW264.7 Macrophage Cells by Inhibiting Notch1. Inflammation 2014, 37, 71–82. [Google Scholar] [CrossRef]
- Fei, Y.; Chaulagain, A.; Wang, T.; Chen, Y.; Liu, J.; Yi, M.; Wang, Y.; Huang, Y.; Lin, L.; Chen, S.; et al. MiR-146a down-regulates inflammatory response by targeting TLR3 and TRAF6 in Coxsackievirus B infection. RNA 2020, 26, 91–100. [Google Scholar] [CrossRef]
- Fasseu, M.; Tréton, X.; Guichard, C.; Pedruzzi, E.; Cazals-Hatem, D.; Richard, C.; Aparicio, T.; Daniel, F.; Soulé, J.-C.; Moreau, R.; et al. Identification of Restricted Subsets of Mature microRNA Abnormally Expressed in Inactive Colonic Mucosa of Patients with Inflammatory Bowel Disease. PLoS ONE 2010, 5, e13160. [Google Scholar] [CrossRef]
- Shukla, A.; Rastogi, M.; Singh, S.K. Zika virus NS1 suppresses the innate immune responses via miR-146a in human microglial cells. Int. J. Biol. Macromol. 2021, 193, 2290–2296. [Google Scholar] [CrossRef] [PubMed]
- Gilyazova, I.; Ivanova, E.; Pavlov, V.; Khasanova, G.; Khasanova, A.; Izmailov, A.; Asadullina, D.; Gilyazova, G.; Wang, G.; Gareev, I.; et al. Exosomal miRNA-155 and miRNA-146a are promising prognostic biomarkers of the severity of hemorrhagic fever with renal syndrome. Non-Coding RNA Res. 2023, 8, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qin, L.; Wang, J.; Xie, X.; Zhang, Y.; Li, C.; Guan, Z.; Qian, L.; Chen, L.; Hu, J.; et al. miR-146a Maintains Immune Tolerance of Kupffer Cells and Facilitates Hepatitis B Virus Persistence in Mice. J. Immunol. 2022, 208, 2558–2572. [Google Scholar] [CrossRef]
- Teng, Y.; Luo, M.; Yu, T.; Chen, L.; Huang, Q.; Chen, S.; Xie, L.; Zeng, Y.; Luo, F.; Xiong, H.; et al. CRISPR/Cas9-mediated deletion of miR-146a enhances antiviral response in HIV-1 infected cells. Genes Immun. 2019, 20, 327–337. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Z.; Su, J.; Chen, W.-S.; Wang, X.-W.; Bai, S.-X.; Zhang, J.-Z.; Yu, S.-Q.; Wu, Y.; He, H.; et al. Macrophage micro-RNA-155 promotes lipopolysaccharide-induced acute lung injury in mice and rats. Am. J. Physiol. Cell Mol. Physiol. 2016, 311, L494–L506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahamtan, A.; Teymoori-Rad, M.; Nakstad, B.; Salimi, V. Anti-Inflammatory MicroRNAs and Their Potential for Inflammatory Diseases Treatment. Front. Immunol. 2018, 9, 1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbatinelli, J.; Giuliani, A.; Matacchione, G.; Latini, S.; Laprovitera, N.; Pomponio, G.; Ferrarini, A.; Baroni, S.S.; Pavani, M.; Moretti, M.; et al. Decreased serum levels of the inflammaging marker miR-146a are associated with clinical non-response to tocilizumab in COVID-19 patients. Mech. Ageing Dev. 2021, 193, 111413. [Google Scholar] [CrossRef]
- Benz, F.; Roy, S.; Trautwein, C.; Roderburg, C.; Luedde, T. Circulating MicroRNAs as Biomarkers for Sepsis. Int. J. Mol. Sci. 2016, 17, 78. [Google Scholar] [CrossRef] [Green Version]
- Mambula, S.S.; Sau, K.; Henneke, P.; Golenbock, D.T.; Levitz, S.M. Toll-like Receptor (TLR) Signaling in Response toAspergillus fumigatus. J. Biol. Chem. 2002, 277, 39320–39326. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Duan, Z.; Zeng, R.; Du, L.; Xu, S.; Wang, L.; Liu, Y.; Chen, Q.; Chen, X.; Li, M. MiR-146a Negatively Regulates Aspergillus fumigatus-Induced TNF-α and IL-6 Secretion in THP-1 Macrophages. Mycopathologia 2021, 186, 341–354. [Google Scholar] [CrossRef]
- Lederhuber, H.; Baer, K.; Altiok, I.; Sadeghi, K.; Herkner, K.R.; Kasper, D.C. MicroRNA-146: Tiny Player in Neonatal Innate Immunity? Neonatology 2011, 99, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Rau, C.-S.; Yang, J.C.-S.; Chen, Y.-C.; Wu, C.-J.; Lu, T.-H.; Tzeng, S.-L.; Wu, Y.-C.; Hsieh, C.-H. Lipopolysaccharide-Induced microRNA-146a Targets CARD10 and Regulates Angiogenesis in Human Umbilical Vein Endothelial Cells. Toxicol. Sci. 2014, 140, 315–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Xu, B.; Zhang, J.; Kou, C.; Liu, J.; Wang, Q.; Zhang, R. Exosomal miR-146a-5p from Treponema pallidum-stimulated macrophages reduces endothelial cells permeability and monocyte transendothelial migration by targeting JAM-C. Exp. Cell Res. 2020, 388, 111823. [Google Scholar] [CrossRef] [PubMed]
- Mensà, E.; Giuliani, A.; Matacchione, G.; Gurău, F.; Bonfigli, A.R.; Romagnoli, F.; De Luca, M.; Sabbatinelli, J.; Olivieri, F. Circulating miR-146a in healthy aging and type 2 diabetes: Age- and gender-specific trajectories. Mech. Ageing Dev. 2019, 180, 1–10. [Google Scholar] [CrossRef]
- Ong, J.; Woldhuis, R.R.; Boudewijn, I.M.; Berg, A.v.D.; Kluiver, J.; Kok, K.; Terpstra, M.M.; Guryev, V.; de Vries, M.; Vermeulen, C.J.; et al. Age-related gene and miRNA expression changes in airways of healthy individuals. Sci. Rep. 2019, 9, 3765. [Google Scholar] [CrossRef] [Green Version]
- Jurkin, J.; Schichl, Y.M.; Koeffel, R.; Bauer, T.; Richter, S.; Konradi, S.; Gesslbauer, B.; Strobl, H. miR-146a Is Differentially Expressed by Myeloid Dendritic Cell Subsets and Desensitizes Cells to TLR2-Dependent Activation. J. Immunol. 2010, 184, 4955–4965. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, K.; Nakasa, T.; Miyaki, S.; Ishikawa, M.; Deie, M.; Adachi, N.; Yasunaga, Y.; Asahara, H.; Ochi, M. Expression of MicroRNA-146a in osteoarthritis cartilage. Arthritis Rheum. 2009, 60, 1035–1041. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Vargas, M.; Cortés-Rojo, C. Ferritin levels and COVID-19. Rev. Panam. Salud Pública 2020, 44, e72. [Google Scholar] [CrossRef]
- Arroyo, A.B.; Fernández-Pérez, M.P.; del Monte, A.; Águila, S.; Méndez, R.; Hernández-Antolín, R.; García-Barber, N.; Reyes-García, A.M.d.L.; González-Jiménez, P.; Arcas, M.I.; et al. miR-146a is a pivotal regulator of neutrophil extracellular trap formation promoting thrombosis. Haematologica 2020, 106, 1636–1646. [Google Scholar] [CrossRef]
- Jazdzewski, K.; Murray, E.L.; Franssila, K.; Jarzab, B.; Schoenberg, D.R.; de la Chapelle, A. Common SNP in pre-miR-146a decreases mature miR expression and predisposes to papillary thyroid carcinoma. Proc. Natl. Acad. Sci. USA 2008, 105, 7269–7274. [Google Scholar] [CrossRef]
- Zhou, Q.; Hou, S.; Liang, L.; Li, X.; Tan, X.; Wei, L.; Lei, B.; Kijlstra, A.; Yang, P. MicroRNA-146a and Ets-1 gene polymorphisms in ocular Behçet’s disease and Vogt–Koyanagi–Harada syndrome. Ann. Rheum. Dis. 2014, 73, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Li, Y.; Ma, G.; Wang, Y.; Cai, Y.; Liu, S.; Chen, Y.; Li, J.; Xie, Y.; Liu, G.; et al. A Functional Polymorphism in the Promoter Region of MicroRNA-146a Is Associated with the Risk of Alzheimer Disease and the Rate of Cognitive Decline in Patients. PLoS ONE 2014, 9, e89019. [Google Scholar] [CrossRef]
- Hung, P.-S.; Chang, K.-W.; Kao, S.-Y.; Chu, T.-H.; Liu, C.-J.; Lin, S.-C. Association between the rs2910164 polymorphism in pre-mir-146a and oral carcinoma progression. Oral Oncol. 2012, 48, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Labib, A.D.; Shaker, O.G.; El Refai, R.M.; Ghoniem, A.S.; Elmazny, A. Association between miRNA-146a and Polymorphisms of its Target Gene, IRAK1, Regarding Susceptibility to and Clinical Features of Systemic Lupus Erythematous and Multiple Sclerosis. Lab. Med. 2019, 50, 34–41. [Google Scholar] [CrossRef]
- Li, L.; Chen, X.-P.; Li, Y.-J. MicroRNA-146a and Human Disease. Scand. J. Immunol. 2010, 71, 227–231. [Google Scholar] [CrossRef]
- Li, Y.; Du, C.; Wang, W.; Ma, G.; Cui, L.; Zhou, H.; Tao, H.; Yao, L.; Zhao, B.; Li, K. Genetic Association of MiR-146a with Multiple Sclerosis Susceptibility in the Chinese Population. Cell. Physiol. Biochem. 2015, 35, 281–291. [Google Scholar] [CrossRef]
- Ahmadi, K.; Soleimani, A.; Soleimani Motlagh, S.; Baharvand Ahmadi, S.; Almasian, M.; Kiani, A.A. Polymorphisms of Pre-MiR-499 Rs3746444 T/C and Pre-MiRNA-146a Rs2910164 C/G in the Autoimmune Diseases of Rheumatoid Arthritis and Systemic Lupus Erythematosus in the West of Iran. Iran J. Public Health 2020, 49, 782–790. [Google Scholar]
- Shao, Y.; Li, J.; Cai, Y.; Xie, Y.; Ma, G.; Li, Y.; Chen, Y.; Liu, G.; Zhao, B.; Cui, L.; et al. The Functional Polymorphisms of miR-146a Are Associated with Susceptibility to Severe Sepsis in the Chinese Population. Mediat. Inflamm. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, T.; Song, X.; Qucuo, M.; Yang, B.; Zhang, J.; Wang, J.; Ying, B.; Tao, C.; Wang, L. Genetic study of two single nucleotide polymorphisms within corresponding microRNAs and susceptibility to tuberculosis in a Chinese Tibetan and Han population. Hum. Immunol. 2011, 72, 598–602. [Google Scholar] [CrossRef]
- Ahmad, M.; Ahmad, S.; Rahman, B.; Haq, T.U.; Jalil, F.; Shah, A.A. Association of MIR146A rs2910164 variation with a predisposition to sporadic breast cancer in a Pakistani cohort. Ann. Hum. Genet. 2019, 83, 325–330. [Google Scholar] [CrossRef]
- Keewan, E.; Naser, S.A. MiR-146a rs2910164 G > C polymorphism modulates Notch-1/IL-6 signaling during infection: A possible risk factor for Crohn’s disease. Gut Pathog. 2020, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, S.; Afshar, S.; Karami, M.; Saidijam, M.; Keramat, F.; Hashemi, S.H.; Alikhani, M.Y. Association between risk of brucellosis and genetic variations in MicroRNA-146a. BMC Infect. Dis. 2021, 21, 1070. [Google Scholar] [CrossRef] [PubMed]
- Xion, Y.; Chen, S.; Chen, R.; Lin, W.; Ni, J. Association between microRNA polymorphisms and humoral immunity to hepatitis B vaccine. Hum. Vaccines Immunother. 2013, 9, 1673–1678. [Google Scholar] [CrossRef] [Green Version]
- Cezar-De-Mello, P.F.T.; Toledo-Pinto, T.G.; Marques, C.S.; Arnez, L.E.A.; Cardoso, C.C.; Guerreiro, L.T.A.; Antunes, S.L.G.; Jardim, M.M.; Covas, C.d.J.F.; Illaramendi, X.; et al. Pre-miR-146a (rs2910164 G>C) Single Nucleotide Polymorphism Is Genetically and Functionally Associated with Leprosy. PLoS Neglected Trop. Dis. 2014, 8, e3099. [Google Scholar] [CrossRef]
- van Loon, W.; Gai, P.P.; Hamann, L.; Bedu-Addo, G.; Mockenhaupt, F.P. MiRNA-146a polymorphism increases the odds of malaria in pregnancy. Malar. J. 2019, 18, 7. [Google Scholar] [CrossRef] [Green Version]
- Gupta, H.; Sahu, P.K.; Pattnaik, R.; Mohanty, A.; Majhi, M.; Mohanty, A.K.; Pirpamer, L.; Hoffmann, A.; Mohanty, S.; Wassmer, S.C. Plasma levels of hsa-miR-3158-3p microRNA on admission correlate with MRI findings and predict outcome in cerebral malaria. Clin. Transl. Med. 2021, 11, e396. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef]
- He, H.; Jazdzewski, K.; Li, W.; Liyanarachchi, S.; Nagy, R.; Volinia, S.; Calin, G.A.; Liu, C.-G.; Franssila, K.; Suster, S.; et al. The role of microRNA genes in papillary thyroid carcinoma. Proc. Natl. Acad. Sci. USA 2005, 102, 19075–19080. [Google Scholar] [CrossRef]
- Bonci, D.; Coppola, V.; Musumeci, M.; Addario, A.; Giuffrida, R.; Memeo, L.; D’Urso, L.; Pagliuca, A.; Biffoni, M.; Labbaye, C.; et al. The miR-15a–miR-16-1 cluster controls prostate cancer by targeting multiple oncogenic activities. Nat. Med. 2008, 14, 1271–1277. [Google Scholar] [CrossRef]
- Pacifico, F.; Crescenzi, E.; Mellone, S.; Iannetti, A.; Porrino, N.; Liguoro, D.; Moscato, F.; Grieco, M.; Formisano, S.; Leonardi, A. Nuclear Factor-κB Contributes to Anaplastic Thyroid Carcinomas through Up-Regulation of miR-146a. J. Clin. Endocrinol. Metab. 2010, 95, 1421–1430. [Google Scholar] [CrossRef]
- Hurst, D.R.; Edmonds, M.D.; Scott, G.K.; Benz, C.C.; Vaidya, K.S.; Welch, D.R. Breast Cancer Metastasis Suppressor 1 Up-regulates miR-146, Which Suppresses Breast Cancer Metastasis. Cancer Res. 2009, 69, 1279–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulis, M.; Esteller, M. DNA Methylation and Cancer. Adv. Genet. 2010, 70, 27–56. [Google Scholar] [PubMed]
- Iacona, J.R.; Monteleone, N.J.; Lutz, C.S. miR-146a suppresses 5-lipoxygenase activating protein (FLAP) expression and Leukotriene B4 production in lung cancer cells. Oncotarget 2018, 9, 26751–26769. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Y.; Sun, X.-X.; Ma, X.; Chen, Z.-N. microRNA-146a inhibits cancer metastasis by downregulating VEGF through dual pathways in hepatocellular carcinoma. Mol. Cancer 2015, 14, 5. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-T.; Fang, L.; Cheng, Y.-X.; Sun, Q. LncRNA PVT1 regulates prostate cancer cell growth by inducing the methylation of miR-146a. Cancer Med. 2016, 5, 3512–3519. [Google Scholar] [CrossRef]
- Yap, E.; Norziha, Z.; Simbun, A.; Tumian, N.; Cheong, S.; Leong, C.; Wong, C. MicroRNAs that affect the Fanconi Anemia/BRCA pathway are downregulated in imatinib-resistant chronic myeloid leukemia patients without detectable BCR-ABL kinase domain mutations. Leuk. Res. 2017, 59, 32–40. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, S.; Yang, J.; Chen, X.; Huang, W. Identification of miR-146a is Associated with the Aggressiveness and Suppresses Proliferation via Targeting CDKN2A in Breast Cancer. Pathol. Oncol. Res. 2018, 26, 245–251. [Google Scholar] [CrossRef]
- Wu, H.; Liu, Q.; Cai, T.; Chen, Y.-D.; Wang, Z.-F. Induction of microRNA-146a is involved in curcumin-mediated enhancement of temozolomide cytotoxicity against human glioblastoma. Mol. Med. Rep. 2015, 12, 5461–5466. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, A.; Gupta, N.; Tang, C.; Mallilankaraman, K.; Silambarasan, M.; Shi, M.; Lu, L.; Ang, B.T.; Ling, E.-A.; Dheen, S.T. Microglial SMAD4 regulated by microRNA-146a promotes migration of microglia which support tumor progression in a glioma environment. Oncotarget 2018, 9, 24950–24969. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, W.; Zhang, L.; Chen, X.; Liu, F.; Zhang, J.; Guan, S.; Sun, Y.; Chen, P.; Wang, D.; et al. miR-146a-5p mediates epithelial–mesenchymal transition of oesophageal squamous cell carcinoma via targeting Notch2. Br. J. Cancer 2016, 115, 1548–1554. [Google Scholar] [CrossRef]
- Iacona, J.R.; Lutz, C.S. miR-146a-5p: Expression, regulation, and functions in cancer. Wiley Interdiscip. Rev. RNA 2019, 10, e1533. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tang, S.; Le, S.-Y.; Lu, R.; Rader, J.S.; Meyers, C.; Zheng, Z.-M. Aberrant Expression of Oncogenic and Tumor-Suppressive MicroRNAs in Cervical Cancer Is Required for Cancer Cell Growth. PLoS ONE 2008, 3, e2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Song, J.; Ding, B.; Cui, Y.; Liang, J.; Han, S. miR-146a promotes cervical cancer cell viability via targeting IRAK1 and TRAF6. Oncol. Rep. 2018, 39, 3015–3024. [Google Scholar] [CrossRef] [Green Version]
- Forloni, M.; Dogra, S.K.; Dong, Y.; Conte, D.; Ou, J.; Zhu, L.J.; Deng, A.; Mahalingam, M.; Green, M.R.; Wajapeyee, N. miR-146a promotes the initiation and progression of melanoma by activating Notch signaling. Elife 2014, 3, e01460. [Google Scholar] [CrossRef] [PubMed]
- Raimo, M.; Orso, F.; Grassi, E.; Cimino, D.; Penna, E.; De Pittà, C.; Stadler, M.B.; Primo, L.; Calautti, E.; Quaglino, P.; et al. miR-146a Exerts Differential Effects on Melanoma Growth and Metastatization. Mol. Cancer Res. 2016, 14, 548–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, W.; Shang, Y.; Shao, Q.; Yuan, X. miR-146a promotes cell migration and invasion in melanoma by directly targeting SMAD4. Oncol. Lett. 2018, 15, 7111–7117. [Google Scholar] [CrossRef] [PubMed]
- Reddemann, K.; Gola, D.; Schillert, A.; Knief, J.; Kuempers, C.; Ribbat-Idel, J.; Ber, S.; Schemme, J.; Bernard, V.; Gebauer, N.; et al. Dysregulation of MiRNAs in Angioimmunoblastic T-Cell Lymphoma. Anticancer Res. 2015, 35, 2055–2061. [Google Scholar]
- Inoue, J.-I.; Gohda, J.; Akiyama, T.; Semba, K. NF-?B activation in development and progression of cancer. Cancer Sci. 2007, 98, 268–274. [Google Scholar] [CrossRef]
- Kim, J.; Yao, F.; Xiao, Z.; Sun, Y.; Ma, L. MicroRNAs and metastasis: Small RNAs play big roles. Cancer Metastasis Rev. 2018, 37, 5–15. [Google Scholar] [CrossRef]
- Lou, W.; Liu, J.; Gao, Y.; Zhong, G.; Chen, D.; Shen, J.; Bao, C.; Xu, L.; Pan, J.; Cheng, J.; et al. MicroRNAs in cancer metastasis and angiogenesis. Oncotarget 2017, 8, 115787–115802. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Cao, Y.; He, Z.; He, J.; Hu, C.; Duan, H.; Jiang, J. Serum Levels of miR-19b and miR-146a as Prognostic Biomarkers for Non-Small Cell Lung Cancer. Tohoku J. Exp. Med. 2014, 232, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleau, A.-M.; Redrado, M.; Nistal-Villan, E.; Villalba, M.; Exposito, F.; Redin, E.; de Aberasturi, A.L.; Larzabal, L.; Freire, J.; Gomez-Roman, J.; et al. miR-146a targets c-met and abolishes colorectal cancer liver metastasis. Cancer Lett. 2018, 414, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Bhaumik, D.; Scott, G.K.; Schokrpur, S.; Patil, C.K.; Campisi, J.; Benz, C.C. Expression of microRNA-146 suppresses NF-κB activity with reduction of metastatic potential in breast cancer cells. Oncogene 2008, 27, 5643–5647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-García, E.I.; Meza-Sosa, K.F.; López-Sevilla, Y.; Camacho-Concha, N.; Sánchez, N.C.; Pérez-Martínez, L.; Pedraza-Alva, G. Merlin negative regulation by miR-146a promotes cell transformation. Biochem. Biophys. Res. Commun. 2015, 468, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Liao, Y.; Qiu, Y.; Liu, H.; Tan, D.; Wu, T.; Tang, M.; Zhang, S.; Wang, H. miRNA 146a promotes chemotherapy resistance in lung cancer cells by targeting DNA damage inducible transcript 3 (CHOP). Cancer Lett. 2018, 428, 55–68. [Google Scholar] [CrossRef]
- Hu, W.; Tan, C.; He, Y.; Zhang, G.; Xu, Y.; Tang, J. Functional miRNAs in breast cancer drug resistance. OncoTargets Ther. 2018, 11, 1529–1541. [Google Scholar] [CrossRef] [Green Version]
- Wilczyński, M.; Żytko, E.; Szymańska, B.; Dzieniecka, M.; Nowak, M.; Danielska, J.; Stachowiak, G.; Wilczyński, J.R. Expression of miR-146a in patients with ovarian cancer and its clinical significance. Oncol. Lett. 2017, 14, 3207–3214. [Google Scholar] [CrossRef] [Green Version]
- Marschner, D.; Falk, M.; Javorniczky, N.R.; Hanke-Müller, K.; Rawluk, J.; Schmitt-Graeff, A.; Simonetta, F.; Haring, E.; Dicks, S.; Ku, M.; et al. MicroRNA-146a regulates immune-related adverse events caused by immune checkpoint inhibitors. J. Clin. Investig. 2020, 5, e132334. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Lu, Z.; Tian, J.; Wang, G.; Gao, Z. Effect of a functional polymorphism in the pre-miR-146a gene on the risk and prognosis of renal cell carcinoma. Mol. Med. Rep. 2015, 12, 6997–7004. [Google Scholar] [CrossRef]
- Ivanova, E.; Asadullina, D.; Rakhimov, R.; Izmailov, A.; Gilyazova, G.; Galimov, S.; Pavlov, V.; Khusnutdinova, E.; Gilyazova, I. Exosomal miRNA-146a is downregulated in clear cell renal cell carcinoma patients with severe immune-related adverse events. Non-Coding RNA Res. 2022, 7, 159–163. [Google Scholar] [CrossRef]
- Huber, V.; Vallacchi, V.; Fleming, V.; Hu, X.; Cova, A.; Dugo, M.; Shahaj, E.; Sulsenti, R.; Vergani, E.; Filipazzi, P.; et al. Tumor-derived microRNAs induce myeloid suppressor cells and predict immunotherapy resistance in melanoma. J. Clin. Investig. 2018, 128, 5505–5516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Li, X.; Zhao, Z.; Yang, X.; Xiao, S.; Zhou, Y. MicroRNA-146a affects the chemotherapeutic sensitivity and prognosis of advanced gastric cancer through the regulation of LIN52. Oncol. Lett. 2017, 13, 1386–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; She, K.; Tian, D.; Zhang, P.; Xin, X. miR-146a Inhibits Proliferation and Enhances Chemosensitivity in Epithelial Ovarian Cancer via Reduction of SOD2. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2016, 23, 275–282. [Google Scholar] [CrossRef]
- Shi, L.; Xu, Z.; Wu, G.; Chen, X.; Huang, Y.; Wang, Y.; Jiang, W.; Ke, B. Up-regulation of miR-146a increases the sensitivity of non-small cell lung cancer to DDP by downregulating cyclin J. BMC Cancer 2017, 17, 138. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Umelo, I.A.; Lv, S.; Teugels, E.; Fostier, K.; Kronenberger, P.; Dewaele, A.; Sadones, J.; Geers, C.; De Grève, J. miR-146a Inhibits Cell Growth, Cell Migration and Induces Apoptosis in Non-Small Cell Lung Cancer Cells. PLoS ONE 2013, 8, e60317. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; He, R.; Rong, M.; Dang, Y.; Chen, G. Synergistic Effect of MiR-146a Mimic and Cetuximab on Hepatocellular Carcinoma Cells. BioMed. Res. Int. 2014, 2014, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Chu, S.; Li, H.; Qiu, Y. MicroRNA-146a-5p enhances ginsenoside Rh2-induced anti-proliferation and the apoptosis of the human liver cancer cell line HepG2. Oncol. Lett. 2018, 16, 5367–5374. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Si, Z.-Z.; Li, T.; Li, J.-Q.; Zhang, Z.-Q.; Chen, G.-S.; Qi, H.-Z.; Yao, H.-L. MicroRNA-146a-5p enhances radiosensitivity in hepatocellular carcinoma through replication protein A3-induced activation of the DNA repair pathway. Am. J. Physiol. Physiol. 2019, 316, C299–C311. [Google Scholar] [CrossRef]
- Petrović, N.; Stanojković, T.P.; Nikitović, M. MicroRNAs in Prostate Cancer Following Radiotherapy: Towards Predicting Response to Radiation Treatment. Curr. Med. Chem. 2022, 29, 1543–1560. [Google Scholar] [CrossRef]
- Yan, W.; Zhang, W.; Sun, L.; Liu, Y.; You, G.; Wang, Y.; Kang, C.; You, Y.; Jiang, T. Identification of MMP-9 specific microRNA expression profile as potential targets of anti-invasion therapy in glioblastoma multiforme. Brain Res. 2011, 1411, 108–115. [Google Scholar] [CrossRef]
- Fisher, J.F.; Mobashery, S. Mechanism-Based Profiling of MMPs. Methods Mol. Biol. 2010, 622, 471–487. [Google Scholar] [CrossRef] [PubMed]
- Shomali, N.; Shirafkan, N.; Duijf, P.H.; Ghasabi, M.; Babaloo, Z.; Yousefi, M.; Mansoori, B.; Asadi, M.; Shanehbandi, D.; Baghbani, E.; et al. Downregulation of miR-146a promotes cell migration in Helicobacter pylori–negative gastric cancer. J. Cell. Biochem. 2019, 120, 9495–9505. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-X.; Liu, Y.-S.; Zhang, X.-J. TargetScore used to reveal potential targets of miRNA203 and miRNA-146a in psoriasis by integrating microRNA overexpression and microarray data. Medicine 2018, 97, e12671. [Google Scholar] [CrossRef] [PubMed]
- Thurmond, R.L.; Gelfand, E.W.; Dunford, P.J. The role of histamine H1 and H4 receptors in allergic inflammation: The search for new antihistamines. Nat. Rev. Drug Discov. 2008, 7, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Petit-Bertron, A.-F.; Machavoine, F.; Defresne, M.P.; Gillard, M.; Chatelain, P.; Mistry, P.; Schneider, E.; Dy, M. H4 Histamine Receptors Mediate Cell Cycle Arrest in Growth Factor-Induced Murine and Human Hematopoietic Progenitor Cells. PLoS ONE 2009, 4, e6504. [Google Scholar] [CrossRef]
- Gschwandtner, M.; Mommert, S.; Köther, B.; Werfel, T.; Gutzmer, R. The Histamine H4 Receptor Is Highly Expressed on Plasmacytoid Dendritic Cells in Psoriasis and Histamine Regulates Their Cytokine Production and Migration. J. Investig. Dermatol. 2011, 131, 1668–1676. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Wang, Y.; Dong, X.; Sheng, M.; Wang, H.; Shi, J.; Sheng, Y.; Liu, L.; Jiang, Q.; Chen, Y.; et al. Downregulation of miRNA-146a-5p promotes malignant transformation of mesenchymal stromal/stem cells by glioma stem-like cells. Aging 2020, 12, 9151–9172. [Google Scholar] [CrossRef]
- Yajima, M.; Miyata, M.; Ikuta, K.; Hasegawa, Y.; Oneyama, C.; Kanda, T. Efficient Epstein-Barr Virus Progeny Production Mediated by Cancer-Derived LMP1 and Virally-Encoded microRNAs. Microorganisms 2019, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- Perry, M.M.; Moschos, S.A.; Williams, A.E.; Shepherd, N.J.; Larner-Svensson, H.M.; Lindsay, M.A. Rapid Changes in MicroRNA-146a Expression Negatively Regulate the IL-1β-Induced Inflammatory Response in Human Lung Alveolar Epithelial Cells. J. Immunol. 2008, 180, 5689–5698. [Google Scholar] [CrossRef] [Green Version]
- Rom, S.; Rom, I.; Passiatore, G.; Pacifici, M.; Radhakrishnan, S.; Del Valle, L.; Khalili, K.; Eletto, D.; Peruzzi, F.; Piña-Oviedo, S. CCL8/MCP-2 is a target for mir-146a in HIV-1-infected human microglial cells. FASEB J. 2010, 24, 2292–2300. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hamblin, M.H.; Yin, K.-J. Noncoding RNAs and Stroke. Neuroscientist 2019, 25, 22–26. [Google Scholar] [CrossRef]
- Ouyang, Y.-B.; Stary, C.M.; Yang, G.-Y.; Giffard, R. microRNAs: Innovative Targets for Cerebral Ischemia and Stroke. Curr. Drug Targets 2013, 14, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Chavda, V.; Madhwani, K. Coding and non-coding nucleotides’: The future of stroke gene therapeutics. Genomics 2021, 113, 1291–1307. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Cui, J.G.; Dua, P.; Pogue, A.I.; Bhattacharjee, S.; Lukiw, W.J. Differential expression of miRNA-146a-regulated inflammatory genes in human primary neural, astroglial and microglial cells. Neurosci. Lett. 2011, 499, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.-F.; Sun, Z.-W.; Zhang, Y.; Wang, S.; Yu, Q.-G.; Wu, Z.-B. Regulatory effects of miR-146a/b on the function of endothelial progenitor cells in acute ischemic stroke in mice. Kaohsiung J. Med. Sci. 2017, 33, 369–378. [Google Scholar] [CrossRef]
- He, Y.; Lv, B.; Huan, Y.; Liu, B.; Li, Y.; Jia, L.; Qu, C.; Wang, D.; Yu, H.; Yuan, H. Zhenbao pill protects against acute spinal cord injury via miR-146a-5p regulating the expression of GPR17. Biosci. Rep. 2018, 38, BSR20171132. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-H.; Li, J.-P.; Chen, L.; Liu, J.-L. miR-146a induces apoptosis in neuroblastoma cells by targeting BCL11A. Med. Hypotheses 2018, 117, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Liu, C.; Zhang, Q.; Li, X.; Qin, G.; Huang, Q.; Meng, Y.; Chen, L.; Wei, J. Association between polymorphisms in microRNAs and ischemic stroke in an Asian population: Evidence based on 6,083 cases and 7,248 controls. Clin. Interv. Aging 2018, 13, 1709–1726. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-H.; Su, S.-Y.; Liu, J.-L. Differential Regulation of microRNAs in Patients with Ischemic Stroke. Curr. Neurovasc. Res. 2015, 12, 214–221. [Google Scholar] [CrossRef]
- Li, S.-H.; Chen, L.; Pang, X.-M.; Su, S.-Y.; Zhou, X.; Chen, C.-Y.; Huang, L.-G.; Li, J.-P.; Liu, J.-L. Decreased miR-146a expression in acute ischemic stroke directly targets the Fbxl10 mRNA and is involved in modulating apoptosis. Neurochem. Int. 2017, 107, 156–167. [Google Scholar] [CrossRef]
- Kotb, H.G.; Ibrahim, A.H.; Mohamed, E.F.; Ali, O.M.; Hassanein, N.; Badawy, D.; Aly, E.A. The expression of microRNA 146a in patients with ischemic stroke: An observational study. Int. J. Gen. Med. 2019, 12, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Zhu, R.; Liu, X.; He, Z.; Li, Q. miR-146a and miR-196a2 Polymorphisms in Patients with Ischemic Stroke in the Northern Chinese Han Population. Neurochem. Res. 2014, 39, 1709–1716. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Kim, O.J.; Kim, S.Y.; Oh, S.H.; Oh, D.; Shin, B.S.; Kim, N.K. Association of the miR-146a, miR-149, miR-196a2, and miR-499 Polymorphisms with Ischemic Stroke and Silent Brain Infarction Risk. Arter. Thromb. Vasc. Biol. 2013, 33, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Su, S.; Li, S.; Pang, X.; Chen, C.; Li, J.; Liu, J. MicroRNA-146a down-regulation correlates with neuroprotection and targets pro-apoptotic genes in cerebral ischemic injury in vitro. Brain Res. 2016, 1648, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.S.; Chopp, M.; Pan, W.L.; Wang, X.L.; Fan, B.Y.; Zhang, Y.; Kassis, H.; Zhang, R.L.; Zhang, X.M.; Zhang, Z.G. MicroRNA-146a Promotes Oligodendrogenesis in Stroke. Mol. Neurobiol. 2017, 54, 227–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peplow, P.; Martinez, B. Immunomodulators and microRNAs as neurorestorative therapy for ischemic stroke. Neural Regen. Res. 2017, 12, 865–874. [Google Scholar] [CrossRef]
- Collins, V.P.; Jones, D.T.W.; Giannini, C. Pilocytic astrocytoma: Pathology, molecular mechanisms and markers. Acta Neuropathol. 2015, 129, 775–788. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.A.; Jeyapalan, J.N.; Forshew, T.; Tatevossian, R.G.; Lawson, A.R.J.; Patel, S.N.; Doctor, G.T.; Mumin, M.A.; Picker, S.R.; Phipps, K.P.; et al. Molecular analysis of pediatric brain tumors identifies microRNAs in pilocytic astrocytomas that target the MAPK and NF-κB pathways. Acta Neuropathol. Commun. 2015, 3, 86. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.-Y.; Xi, J.; Zhang, Y.-H.; Zhang, C.-N.; Song, L.; Song, Y.; Hui, R.-T.; Chen, J.-Z. Association of the MicroRNA-146a SNP rs2910164 with Ischemic Stroke Incidence and Prognosis in a Chinese Population. Int. J. Mol. Sci. 2016, 17, 660. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Zhou, S.; Zhang, Y.; Lv, Z.; Li, S.; Xie, C.; Ke, Y.; Deng, P.; Geng, Y.; Zhang, Q.; et al. Association of the Genetic Polymorphisms in Pre-MicroRNAs with Risk of Ischemic Stroke in a Chinese Population. PLoS ONE 2015, 10, e0117007. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Wang, K.; Huang, J.; Zheng, G.; Lv, Y.; Luo, N.; Liang, M.; Huang, L. Upregulated Serum MiR-146b Serves as a Biomarker for Acute Ischemic Stroke. Cell. Physiol. Biochem. 2018, 45, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Li, Y.; Zhang, J.; Zhang, W.; Guo, R. Genetic Variants of MiRNA-146a and MiR-499 and Risk of Ischemic Stroke in the Chinese Population: A Meta-Analysis and Trial Sequential Analysis. Int. J. Clin. Exp. Med. 2019, 12, 7964–7974. [Google Scholar]
- Gangwar, R.S.; Rajagopalan, S.; Natarajan, R.; Deiuliis, A.J. Noncoding RNAs in Cardiovascular Disease: Pathological Relevance and Emerging Role as Biomarkers and Therapeutics. Am. J. Hypertens. 2018, 31, 150–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Ray, K.; Ludwig, N.; Lenhof, H.-P.; Meese, E.; et al. miRTargetLink 2.0—Interactive miRNA target gene and target pathway networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef]
- Chang, L.; Zhou, G.; Soufan, O.; Xia, J. miRNet 2.0: Network-based visual analytics for miRNA functional analysis and systems biology. Nucleic Acids Res. 2020, 48, W244–W251. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilyazova, I.; Asadullina, D.; Kagirova, E.; Sikka, R.; Mustafin, A.; Ivanova, E.; Bakhtiyarova, K.; Gilyazova, G.; Gupta, S.; Khusnutdinova, E.; et al. MiRNA-146a—A Key Player in Immunity and Diseases. Int. J. Mol. Sci. 2023, 24, 12767. https://doi.org/10.3390/ijms241612767
Gilyazova I, Asadullina D, Kagirova E, Sikka R, Mustafin A, Ivanova E, Bakhtiyarova K, Gilyazova G, Gupta S, Khusnutdinova E, et al. MiRNA-146a—A Key Player in Immunity and Diseases. International Journal of Molecular Sciences. 2023; 24(16):12767. https://doi.org/10.3390/ijms241612767
Chicago/Turabian StyleGilyazova, Irina, Dilara Asadullina, Evelina Kagirova, Ruhi Sikka, Artur Mustafin, Elizaveta Ivanova, Ksenia Bakhtiyarova, Gulshat Gilyazova, Saurabh Gupta, Elza Khusnutdinova, and et al. 2023. "MiRNA-146a—A Key Player in Immunity and Diseases" International Journal of Molecular Sciences 24, no. 16: 12767. https://doi.org/10.3390/ijms241612767